Novel Molecular Mechanisms of Gangliosides in the Nervous System Elucidated by Genetic Engineering

 , ,
, ,
Abstract
:1. Introduction
2. Roles of Gangliosides
2.1. Gangliosides in Development and Growth
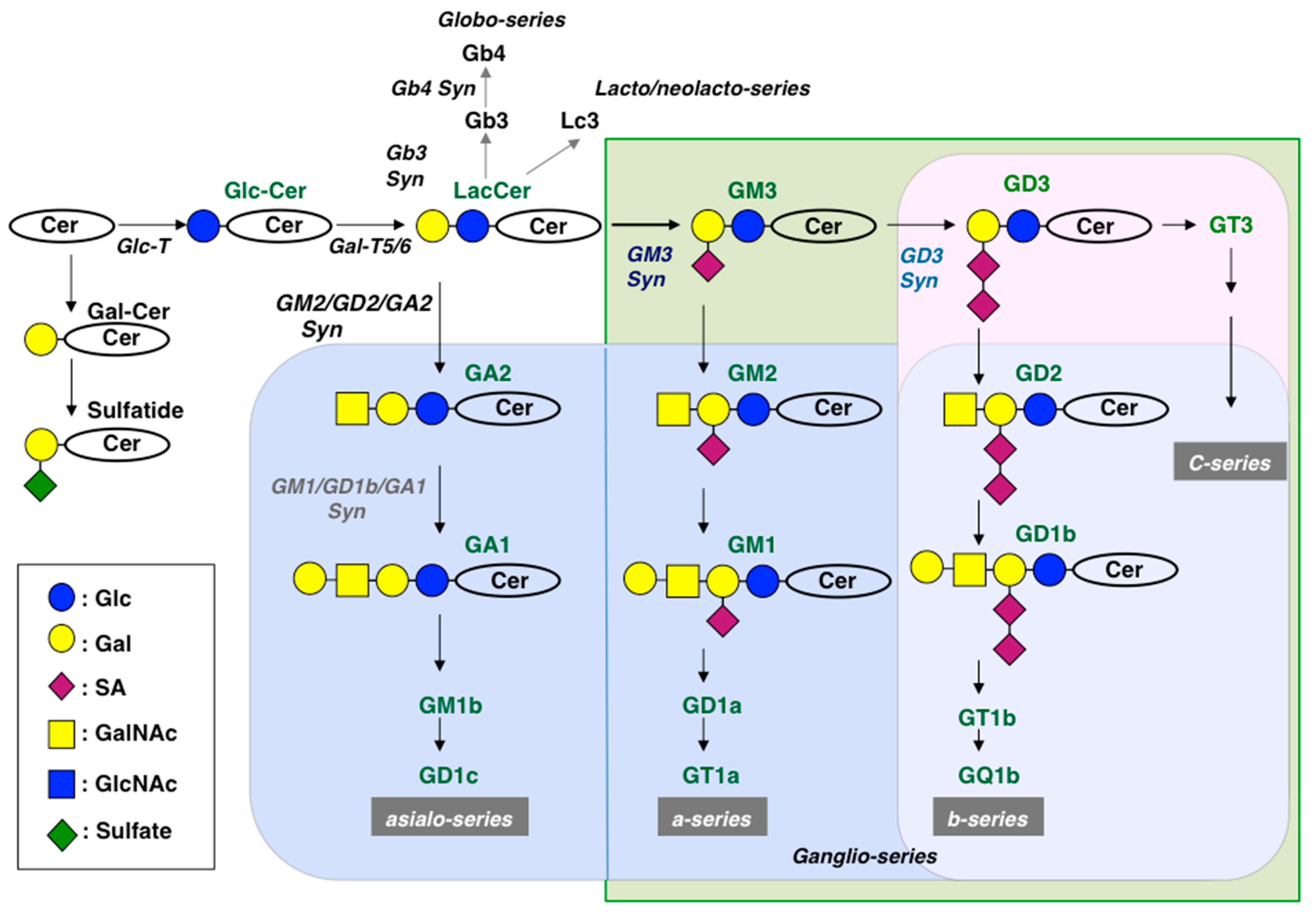
2.2. Function of Monosialylgangliosides and Disialylgangliosides
3. Aging, Neurodegeneration, and Gangliosides
3.1. Changes of Gangliosides in Central Nervous Systems with Aging
3.2. Gangliosides in Alzheimer’s Disease
3.3. Parkinson Disease and Gangliosides
3.4. Inflammatory Reaction and Gangliosides
4. Functions of Glycolipids Elucidated in Ganglioside-Deficient (Knockout) Mice
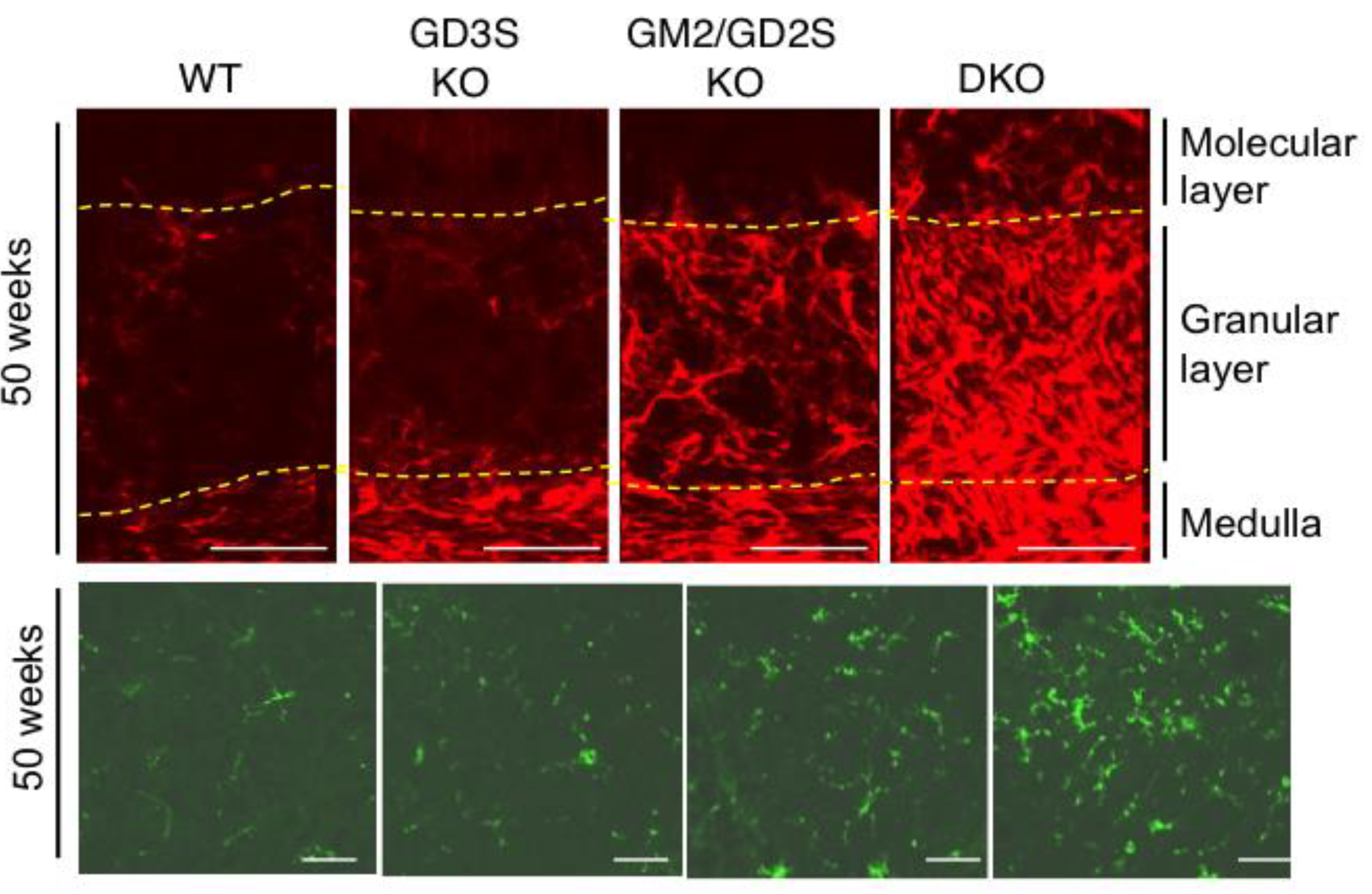
4.1. Abnormal Phenotypes Exhibited by Knockout Mice and Inflammatory Reaction
4.2. Neuro-Inflammation Corresponding to the Degree of Ganglioside Deficiency
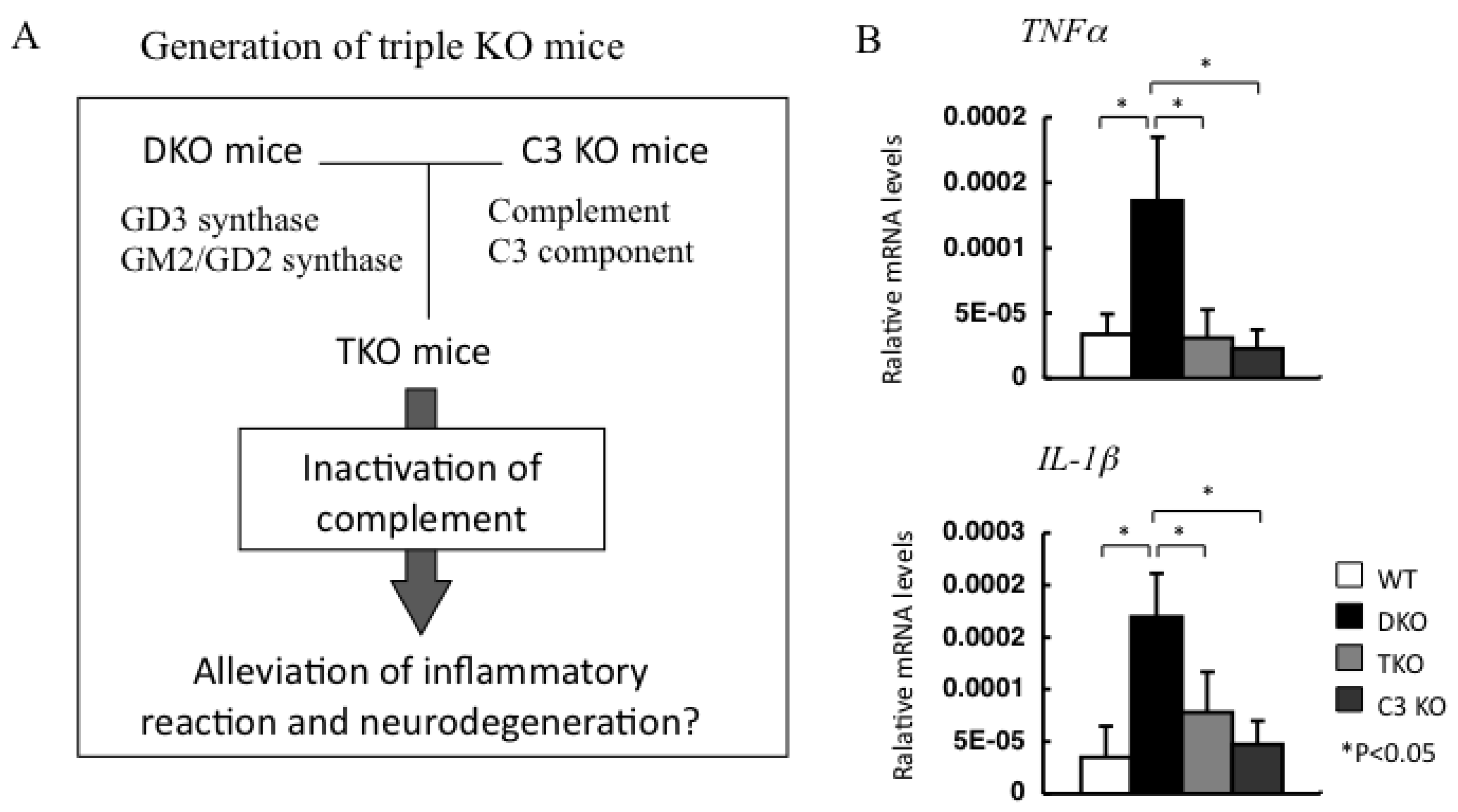
4.3. Involvement of Complement System in the Inflammatory Reaction
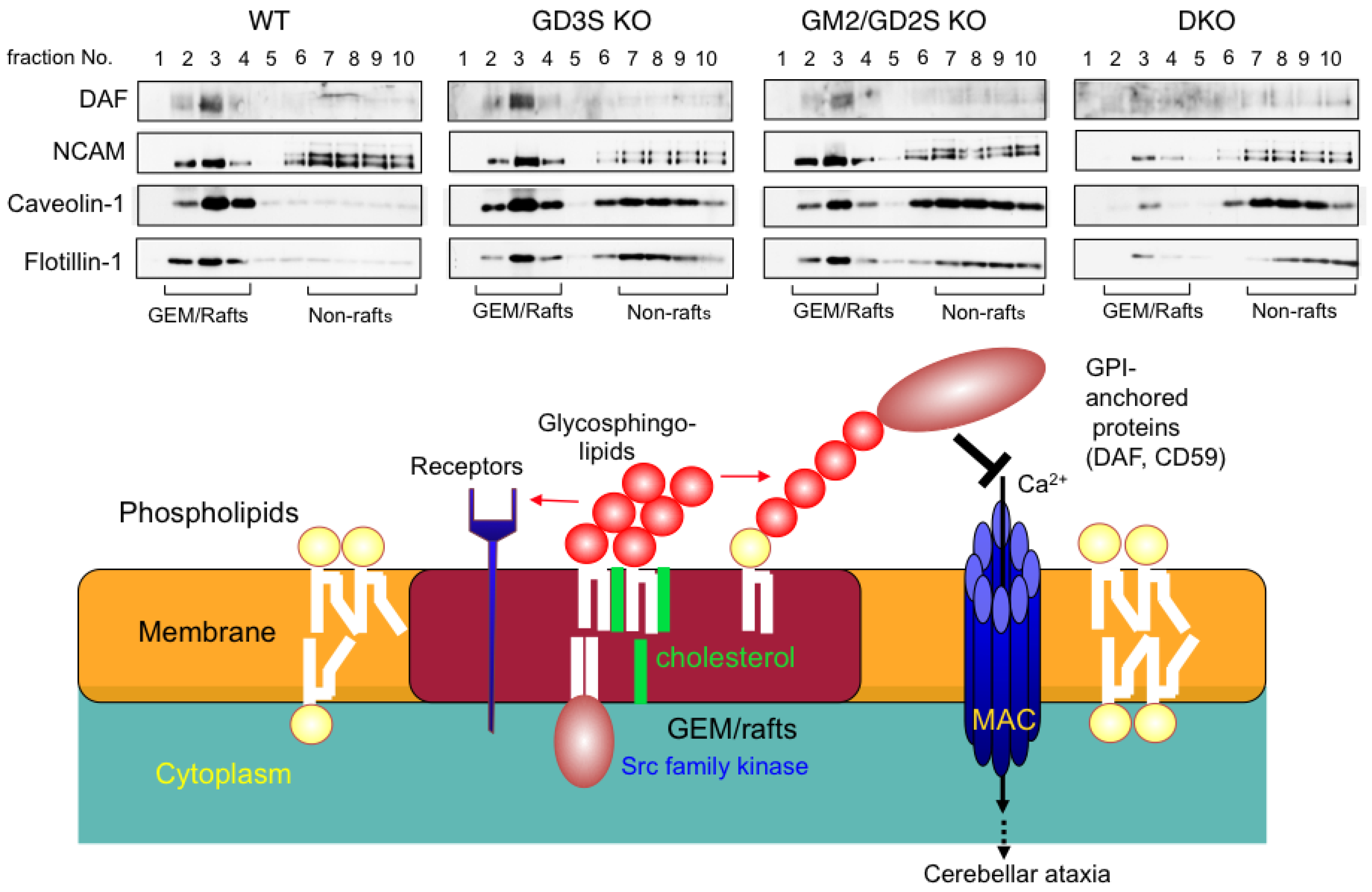
5. Microdomains on Cell Membrane and Gangliosides
5.1. Gangliosides Regulate Cell Signaling in Microdomains
5.2. Microdomain on the Cell Membrane of Nervous Systems and Gangliosides
5.3. Complement Activation and Destruction of Lipid Rafts
6. Human Diseases Caused by Congenital Deficiency of Gangliosides
6.1. GM3 Synthase Deficiency Causes Severe Clinical Features
6.2. GM2/GD2 Synthase Gene Deficiency Causes Hereditary Spastic Paraplegia
7. Future Scope of Ganglioside Research
8. Conclusions
Funding
Conflicts of Interest
References
- Batool, S.; Hussain Raza, H.; Zaidi, J.; Riaz, S.; Hasan, S.; Syed, N.I. Synapse Formation: From Cellular and Molecular Mechanisms to Neurodevelopmental and Neurodegenerative Disorders. J. Neurophysiol. 2019, 121, 1381–1397. [Google Scholar] [CrossRef]
- Ando, S. Neuronal dysfunction with aging and its amelioration. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2012, 88, 266–282. [Google Scholar] [CrossRef] [Green Version]
- Moll, T.; Shaw, P.J.; Cooper-Knock, J. Disrupted glycosylation of lipids and proteins is a cause of neurodegeneration. Brain 2019. [Google Scholar] [CrossRef]
- Yu, R.K.; Tsai, Y.T.; Ariga, T. Functional roles of gangliosides in neurodevelopment: An overview of recent advances. Neurochem. Res. 2012, 37, 1230–1244. [Google Scholar] [CrossRef] [Green Version]
- Itokazu, Y.; Wang, J.; Yu, R.K. Gangliosides in nerve cell specification. Prog. Mol. Biol. Transl. Sci. 2018, 156, 241–263. [Google Scholar]
- Maccioni, H.J.; Quiroga, R.; Ferrari, M.L. Cellular and molecular biology of glycosphingolipid glycosylation. J. Neurochem. 2011, 117, 589–602. [Google Scholar] [CrossRef]
- Schnaar, R.L. The Biology of gangliosides. Adv. Carbohydr. Chem. Biochem. 2019, 76, 113–148. [Google Scholar]
- Sonnino, S.; Chiricozzi, E.; Grassi, S.; Mauri, L.; Prioni, S.; Prinetti, A. Gangliosides in membrane organization. Prog. Mol. Biol. Transl. Sci. 2018, 156, 83–120. [Google Scholar] [PubMed]
- Furukawa, K.; Ohkawa, Y.; Yamauchi, Y.; Hamamura, K.; Ohmi, Y.; Furukawa, K. Fine tuning of cell signals by glycosylation. J. Biochem. 2012, 151, 573–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svennerholm, L. Chromatographic separateion of human brain gangliosides. J. Neurochem. 1963, 10, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Schengrund, C.L. Gangliosides: Glycosphingolipids essential for normal neural development and function. Trends Biochem. Sci. 2015, 40, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.K.; Macala, L.J.; Taki, T.; Weinfield, H.M.; Yu, F.S. Developmental changes in ganglioside composition and synthesis in embryonic rat brain. J. Neurochem. 1988, 50, 1825–1829. [Google Scholar] [CrossRef] [PubMed]
- Suzaki, K. The pattern of mammalian brain gangliosides. 3. Regional and developmental differences. J. Neurochem. 1965, 12, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Vrbaski, S.R. Cerebellum gangliosides in postnatal rats. J. Neurochem. 1980, 34, 1032–1033. [Google Scholar] [CrossRef] [PubMed]
- Greene, L.A.; Tischler, A.S. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Natl. Acad. Sci. USA 1976, 73, 2424–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutoh, T.; Tokuda, A.; Miyadai, T.; Hamaguchi, M.; Fujiki, N. Ganglioside GM1 binds to the Trk protein and regulates receptor function. Proc. Natl. Acad. Sci. USA 1995, 92, 5087–15091. [Google Scholar] [CrossRef] [Green Version]
- Nishio, M.; Fukumoto, S.; Furukawa, K.; Ichimura, A.; Miyazaki, H.; Kusunoki, S.; Urano, T.; Furukawa, K. Overexpressed GM1 suppresses nerve growth factor (NGF) signals by modulating the intracellular localization of NGF receptors and membrane fluidity in PC12 cells. J. Biol. Chem. 2004, 279, 33368–33378. [Google Scholar] [CrossRef] [Green Version]
- Fukumoto, S.; Mutoh, T.; Hasegawa, T.; Miyazaki, H.; Okada, M.; Goto, G.; Furukawa, K.; Urano, T.; Furukawa, K. GD3 synthase gene expression in PC12 cells results in the continuous activation of TrkA and ERK1/2 and enhanced proliferation. J. Biol. Chem. 2000, 275, 5832–5838. [Google Scholar] [CrossRef] [Green Version]
- Hamamura, K.; Furukawa, K.; Hayashi, T.; Hattori, T.; Nakano, J.; Nakashima, H.; Okuda, T.; Mizutani, H.; Hattori, H.; Ueda, M.; et al. Ganglioside GD3 promotes cell growth and invasion through p130Cas and paxillin in malignant melanoma cells. Proc. Natl. Acad. Sci. USA 2005, 102, 11041–11046. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Ikeda, K.; Hamamura, K.; Zhang, Q.; Kondo, Y.; Matsumoto, Y.; Ohmi, Y.; Yamauchi, Y.; Furukawa, K.; Taguchi, R.; et al. GM1/GD1b/GA1 synthase expression results in the reduced cancer phenotypes with modulation of composition and raft-localization of gangliosides in a melanoma cell line. Cancer Sci. 2010, 101, 2039–2047. [Google Scholar] [CrossRef]
- Handa, K.; Hakomori, S.I. Changes of glycoconjugate expression profiles during early development. Glycoconj. J. 2017, 34, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Ohmi, Y.; Ji, S.; Zhang, P.; Bhuiyan, R.H.; Ohkawa, Y.; Tajima, O.; Hashimoto, N.; Furukawa, K. Glycolipids: Essential regulator of neuro- inflammation, metabolism and gliomagenesis. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2479–2484. [Google Scholar] [CrossRef]
- Ohsawa, T. Changes of mouse brain gangliosides during aging from young adult until senescence. Mech. Ageing Dev. 1989, 50, 169–177. [Google Scholar] [CrossRef]
- Hilbig, R.; Lauke, G.; Rahmann, H. Brain gangliosides during the life span (embryogenesis to senescence) of the rat. Dev. Neurosci. 1983, 6, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.K.; Nakatani, Y.; Yanagisawa, M. The role of glycosphingolipid metabolism in the developing brain. J. Lipid Res. 2009, 50, S440–S445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segler-Stahl, K.; Webster, J.C.; Brunngraber, E.G. Changes in the concentration and composition of human brain gangliosides with aging. Gerontology 1983, 29, 161–168. [Google Scholar] [CrossRef]
- Svennerholm, L.; Boström, K.; Helander, C.G.; Jungbjer, B. Membrane lipids in the aging human brain. J. Neurochem. 1991, 56, 2051–2059. [Google Scholar] [CrossRef]
- Svennerholm, L.; Boström, K.; Jungbjer, B.; Olsson, L. Membrane lipids of adult human brain: Lipid composition of frontal and temporal lobe in subjects of age 20 to 100 years. J. Neurochem. 1994, 63, 1802–1811. [Google Scholar] [CrossRef]
- Sarbu, M.; Dehelean, L.; Munteanu, C.V.A.; Vukelić, Ž.; Zamfir, A.D. Assessment of ganglioside age-related and topographic specificity in human brain by Orbitrap mass spectrometry. Anal. Biochem. 2017, 521, 40–54. [Google Scholar] [CrossRef]
- Rogers, J.; Cooper, N.R.; Webster, S.; Schultz, J.; McGeer, P.L.; Styren, S.D.; Civin, W.H.; Brachova, L.; Bradt, B.; Ward, P. Complement activation by beta-amyloid in Alzheimer disease. Proc. Natl. Acad. Sci. USA 1992, 89, 10016–10020. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Lue, L.; Yang, L.; Loher, A.; Kuo, Y.; Strohmeyer, R.; Goux, W.J.; Lee, V.; Johnson, G.V.; Webster, S.D.; et al. Complement activation by neurofibrillary tangles in Alzheimer’s disease. Neurosci. Lett. 2001, 305, 165–168. [Google Scholar] [CrossRef]
- Hoarau, J.J.; Krejbich-Trotot, P.; Jaffar-Bandjee, M.C.; Das, T.; Thon-Hon, G.V.; Kumar, S.; Neal, J.W.; Gasque, P. Activation and control of CNS innate immune responses in health and diseases: A balancing act finely tuned by neuroimmune regulators (NIReg). CNS Neurol. Disord. Drug Targets 2011, 10, 25–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Halperin, J.A.; Benzaquen, L.; Lee, C.M. Characterization of neuronal cell death induced by complement activation. Brain Res. Brain Res. Protoc. 1997, 1, 186–194. [Google Scholar] [CrossRef]
- Terai, K.; Walker, D.G.; McGeer, E.G.; McGeer, P.L. Neurons express proteins of the classical complement pathway in Alzheimer disease. Brain Res. 1997, 769, 385–390. [Google Scholar] [CrossRef]
- Yasojima, K.; Schwab, C.; McGeer, E.G.; McGeer, P.L. Up-regulated production and activation of the complement system in Alzheimer’s disease brain. Am. J. Pathol. 1999, 154, 927–936. [Google Scholar] [CrossRef]
- Jennemann, R.; Sandhoff, R.; Wang, S.; Kiss, E.; Gretz, N.; Zuliani, C.; Martin-Villalba, A.; Jäger, R.; Schorle, H.; Kenzelmann, N.; et al. Cell-specific deletion of glucosyl- ceramide synthase in brain leads to severe neural defects after birth. Proc. Natl. Acad. Sci. USA 2005, 102, 12459–12464. [Google Scholar] [CrossRef] [Green Version]
- Tenner, A.J. Complement in Alzheimer’s disease: Opportunities for modulating protective and pathogenic events. Neurobiol. Aging 2001, 22, 849–861. [Google Scholar] [CrossRef]
- Stevens, B.; Allen, N.J.; Vazquez, L.E.; Howell, G.R.; Christopherson, K.R.; Nouri, N.; Micheva, K.D.; Mehalow, A.K.; Huberman, A.D.; Stafford, B.; et al. The classical complement cascade mediates CNS synapse elimination. Cell 2007, 131, 1164–1178. [Google Scholar] [CrossRef] [Green Version]
- Wyss-Coray, T.; Yan, F.; Lin, A.H.; Lambris, J.D.; Alexander, J.J.; Quigg, R.J.; Masliah, E. Prominent neurodegeneration and increased plaque formation in complement-inhibited Alzheimer’s mice. Proc. Natl. Acad. Sci. USA 2002, 99, 10837–10842. [Google Scholar] [CrossRef] [Green Version]
- Yanagisawa, K. GM1 ganglioside and Alzheimer’s disease. Glycoconj. J. 2015, 32, 87–91. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Kato, K.; Yanagisawa, K. A beta polymerization through interaction with membrane gangliosides. Biochim. Biophys. Acta 2010, 1801, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, A.; Harrison, F.E.; McCord, M.; Zhao, J.; Bruchey, A.; Davies, S.S.; Roberts, L.J., 2nd; Mathews, P.M.; Matsuoka, Y.; Ariga, T.; et al. Elimination of GD3 synthase improves memory and reduces amyloid-beta plaque load in transgenic mice. Neurobiol. Aging 2009, 30, 1777–1791. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, N.; Yamaguchi, H.; Ogino, K.; Taki, T.; Yuyama, K.; Yamamoto, N.; Shin, R.W.; Furukawa, K.; Yanagisawa, K. Gangliosides determine the amyloid pathology of Alzheimer’s disease. Neuroreport 2009, 20, 1043–1046. [Google Scholar]
- Yamaguchi, T.; Yamauchi, Y.; Furukawa, K.; Ohmi, Y.; Ohkawa, Y.; Zhang, Q.; Okajima, T.; Furukawa, K. Expression of B4GALNT1, an essential glycosyltransferase for the synthesis of complex gangliosides, suppresses BACE1 degradation and modulates APP processing. Sci. Rep. 2016, 6, 34505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrier, L.; Ingrand, S.; Damjanac, M.; Bilan, A.R.; Hugon, J.; Page, G. Genotype-related changes of ganglioside composition in brain regions of transgenic mouse models of Alzheimer’s disease. Neurobiol. Aging 2007, 28, 1863–1872. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.S. Gangliosides and glycolipids in neurodegenerative disorders. Adv. Neurobiol. 2014, 9, 449–461. [Google Scholar] [CrossRef]
- Takamiya, K.; Yamamoto, A.; Furukawa, K.; Yamashiro, S.; Shin, M.; Okada, M.; Fukumoto, S.; Haraguchi, M.; Takeda, N.; Fujimura, K.; et al. Mice with disrupted GM2/GD2 synthase gene lack complex gangliosides but exhibit only subtle defects in their nervous system. Proc. Natl. Acad. Sci. USA 1996, 93, 10662–10667. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Lu, Z.H.; Kulkarni, N.; Ledeen, R.W. Deficiency of ganglioside GM1 correlates with Parkinson’s disease in mice and humans. J. Neurosci. Res. 2012, 90, 1997–2008. [Google Scholar] [CrossRef]
- Schneider, J.S.; Pope, A.; Simpson, K.; Taggart, J.; Smith, M.G.; DiStefano, L. Recovery from experimental parkinsonism in primates with GM1 ganglioside treatment. Science 1992, 256, 843–846. [Google Scholar] [CrossRef]
- Schneider, J.S.; Gollomp, S.M.; Sendek, S.; Colcher, A.; Cambi, F.; Du, W. A randomized, controlled, delayed start trial of GM1 ganglioside in treated Parkinson’s disease patients. J. Neurol. Sci. 2013, 324, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Itoh, M.; Haraguchi, M.; Okajima, T.; Inoue, M.; Oishi, H.; Matsuda, Y.; Iwamoto, T.; Kawano, T.; Fukumoto, S.; et al. b-series Ganglioside deficiency exhibits no definite changes in the neurogenesis and the sensitivity to Fas-mediated apoptosis but impairs regeneration of the lesioned hypoglossal nerve. J. Biol. Chem. 2002, 277, 1633–1636. [Google Scholar] [CrossRef] [Green Version]
- Akkhawattanangkul, Y.; Maiti, P.; Xue, Y.; Aryal, D.; Wetsel, W.C.; Hamilton, D.; Fowler, S.C.; McDonald, M.P. Targeted deletion of GD3 synthase protects against MPTP-induced neurodegeneration. Genes Brain Behav. 2017, 16, 522–536. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Ohmi, Y.; Kondo, Y.; Ohkawa, Y.; Tajima, O.; Furukawa, K. Regulatory function of glycosphingolipids in the inflammation and degeneration. Arch. Biochem. Biophys. 2015, 571, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Sela, B.A.; Offner, H.; Konat, G.; Lev-Ram, J.; Cohen, O.; Cohen, I.R. Immunological expression of gangliosides in multiple sclerosis and in a demyelinating model disease in rabbits. Adv. Exp. Med. Biol. 1984, 174, 441–453. [Google Scholar] [PubMed]
- Pilkington, G.J.; Dunan, J.R.; Rogers, J.P.; Clarke, T.M.; Knott, J.C. Growth factor modulation of surface ganglioside expression in cloned neoplastic glia. Neurosci. Lett. 1993, 149, 1–5. [Google Scholar] [CrossRef]
- Kawai, K.; Kuroda, S.; Watarai, S.; Takahashi, H.; Ikuta, F. Occurrence of GD3 ganglioside in reactive astrocytes--an immunocytochemical study in the ratbrain. Neurosci. Lett. 1994, 174, 225–227. [Google Scholar] [CrossRef]
- Welte, K.; Miller, G.; Chapman, P.B.; Yuasa, H.; Natoli, E.; Kunicka, J.E.; Cordon-Cardo, C.; Buhrer, C.; Old, L.J.; Houghton, A.N. Stimulation of T lymphocyte proliferation by monoclonal antibodies against GD3ganglioside. J. Immunol. 1987, 139, 1763–1771. [Google Scholar]
- Yamashiro, S.; Okada, M.; Haraguchi, M.; Furukawa, K.; Lloyd, K.O.; Shiku, H.; Furukawa, K. Expression of alpha 2,8-sialyltransferase (GD3 synthase) gene in human cancer cell lines: High level expression in melanomas and up-regulation in activated T lymphocytes. Glycoconj. J. 1995, 12, 894–900. [Google Scholar] [CrossRef]
- Yoshimura, A.; Takamiya, K.; Kato, I.; Nakaya, E.; Shiku, H.; Furukawa, K. GD2 ganglioside-specific monoclonal antibody reacts with murine cytotoxic T lymphocytes reactive with FBL-3N erythroleukaemia. Scand. J. Immunol. 1994, 40, 557–563. [Google Scholar] [CrossRef]
- Furukawa, K.; Akagi, T.; Nagata, Y.; Yamada, Y.; Shimotohno, K.; Cheung, N.K.; Shiku, H.; Furukawa, K. GD2 ganglioside on human T-lymphotropic virus type I-infected T cells: Possible activation of beta-1,4-N-acetylgalactosaminyl- transferase gene by p40tax. Proc. Natl. Acad. Sci. USA 1993, 90, 1972–1976. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Furukawa, K.; Yamashiro, S.; Yamada, Y.; Haraguchi, M.; Horibe, K.; Kato, K.; Tsuji, Y.; Furukawa, K. High expression of ganglioside alpha-2,8-sialyl- transferase (GD3 synthase) gene in adult T-cell leukemia cells unrelated to the gene expression of human T-lymphotropic virus type I. Cancer Res. 1996, 56, 2844–2848. [Google Scholar] [PubMed]
- Villanueva-Cabello, T.M.; Mollicone, R.; Cruz-Muñoz, M.E.; López-Guerrero, D.V.; Martínez-Duncker, I. Activation of human naïve Th cells increases surface expression of GD3 and induces neoexpression of GD2 that colocalize with TCR clusters. Glycobiology 2015, 25, 1454–1464. [Google Scholar] [CrossRef] [PubMed]
- Nagata, Y.; Yamashiro, S.; Yodoi, J.; Loyd, K.O.; Shiku, H.; Furukawa, K. Expression cloning of beta 1,4 N-acetylgalactosaminyltransferase cDNAs that determine the expression of GM2 and GD2 gangliosides. J. Biol. Chem. 1992, 267, 12082–12089. [Google Scholar] [PubMed]
- Haraguchi, M.; Yamashiro, S.; Yamamoto, A.; Furukawa, K.; Takamiya, K.; Lloyd, K.O.; Shiku, H.; Furukawa, K. Isolation of GD3 synthase gene by expression cloning of GM3 alpha-2,8-sialyltransferase cDNA using anti-GD2 monoclonal antibody. Proc. Natl. Acad. Sci. USA 1994, 91, 10455–10459. [Google Scholar] [CrossRef] [Green Version]
- Nara, K.; Watanabe, Y.; Maruyama, K.; Kasahara, K.; Nagai, Y.; Sanai, Y. Expression cloning of a CMP-NeuAc:NeuAc alpha 2-3Gal beta 1-4Glc beta 1-1′Cer alpha 2,8-sialyltransferase (GD3 synthase) from human melanoma cells. Proc. Natl. Acad. Sci. USA 1994, 91, 7952–7956. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, K.; Kurata, K.; Kojima, N.; Kurosawa, N.; Ohta, S.; Hanai, N.; Tsuji, S.; Nishi, T. Expression cloning of a GM3-specific alpha-2,8-sialyltransferase (GD3synthase). J. Biol. Chem. 1994, 269, 15950–15956. [Google Scholar] [PubMed]
- Miyazaki, H.; Fukumoto, S.; Okada, M.; Hasegawa, T.; Furukawa, K. Expression cloning of rat cDNA encoding UDP-galactose: GD2 beta1,3-galactosyltransferase that determines the expression of GD1b/GM1/GA1. J. Biol. Chem. 1997, 272, 24794–24799. [Google Scholar] [CrossRef] [Green Version]
- Fukumoto, S.; Miyazaki, H.; Goto, G.; Urano, T.; Furukawa, K.; Furukawa, K. Expression cloning of mouse cDNA of CMP-NeuAc: Lactosylceramide alpha2,3-sialyltransferase, an enzyme that initiates the synthesis of gangliosides. J. Biol. Chem. 1999, 274, 9271–9276. [Google Scholar] [CrossRef] [Green Version]
- Kono, M.; Takashima, S.; Liu, H.; Inoue, M.; Kojima, N.; Lee, Y.C.; Hamamoto, T.; Tsuji, S. Molecular cloning and functional expression of a fifth-type alpha2,3-sialyltransferase (mST3Gal V: GM3 synthase). Biochem. Biophys. Res. Commun. 1998, 253, 170–175. [Google Scholar] [CrossRef]
- Ishii, A.; Ohta, M.; Watanabe, Y.; Matsuda, K.; Ishiyama, K.; Sakoe, K.; Nakamura, M.; Inokuchi, J.; Sanai, Y.; Saito, M. Expression cloning and functional characterization of human cDNA for ganglioside GM3 synthase. J. Biol. Chem. 1998, 273, 31652–31655. [Google Scholar] [CrossRef] [Green Version]
- Kojima, Y.; Fukumoto, S.; Furukawa, K.; Okajima, T.; Wiels, J.; Yokoyama, K.; Suzuki, Y.; Urano, T.; Ohta, M.; Furukawa, K. Molecular cloning of globotriaosyl- ceramide/CD77 synthase, a glycosyltransferase that initiates the synthesis of globo series glycosphingolipids. J. Biol. Chem. 2000, 275, 15152–15156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffensen, R.; Carlier, K.; Wiels, J.; Levery, S.B.; Stroud, M.; Cedergren, B.; Nilsson Sojka, B.; Bennett, E.P.; Jersild, C.; Clausen, H. Cloning and expression of the histo-blood group Pk UDP-galactose: Ga1beta-4G1cbeta1-cer alpha1,4-galactosyltransferase. Molecular genetic basis of the p phenotype. J. Biol. Chem. 2000, 275, 16723–16729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keusch, J.J.; Manzella, S.M.; Nyame, K.A.; Cummings, R.D.; Baenziger, J.U. Cloning of Gb3 synthase, the key enzyme in globo-series glycosphingolipid synthesis, predicts a family of alpha1,4-glycosyltransferases conserved in plants, insects, and mammals. J. Biol. Chem. 2000, 275, 25315–25321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuda, T.; Tokuda, N.; Numata, S.; Ito, M.; Ohta, M.; Kawamura, K.; Wiels, J.; Urano, T.; Tajima, O.; Furukawa, K.; et al. Targeted disruption of Gb3/CD77 synthase gene resulted in the complete deletion of globo-series glycosphingolipids and loss of sensitivity to verotoxins. J. Biol. Chem. 2006, 281, 10230–10235. [Google Scholar] [CrossRef] [Green Version]
- Nishie, T.; Hikimochi, Y.; Zama, K.; Fukusumi, Y.; Ito, M.; Yokoyama, H.; Naruse, C.; Ito, M.; Asano, M. Beta4-galactosyltransferase-5 is a lactosylceramide synthase essential for mouse extra-embryonic development. Glycobiology 2010, 20, 1311–1322. [Google Scholar] [CrossRef] [Green Version]
- Tokuda, N.; Numata, S.; Li, X.; Nomura, T.; Takizawa, M.; Kondo, Y.; Yamashita, Y.; Hashimoto, N.; Kiyono, T.; Urano, T. β4GalT6 is involved in the synthesis of lactosylceramide with less intensity than β4GalT5. Glycobiology 2013, 23, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- Inoue, M.; Fujii, Y.; Furukawa, K.; Okada, M.; Okumura, K.; Hayakawa, T.; Furukawa, K.; Sugiura, Y. Refractory skin injury in the complex knock-out mice expressing only GM3 ganglioside. J. Biol. Chem. 2002, 277, 29881–29888. [Google Scholar] [CrossRef] [Green Version]
- Tajima, O.; Egashira, N.; Ohmi, Y.; Fukue, Y.; Mishima, K.; Iwasaki, K.; Fujiwara, M.; Inokuchi, J.; Sugiura, Y.; Furukawa, K.; et al. Reduced motor and sensory functions and emotional response in GM3-only mice: Emergence from early stage of life and exacerbation with aging. Behav. Brain Res. 2009, 198, 74–82. [Google Scholar] [CrossRef]
- Yoshihara, T.; Satake, H.; Nishie, T.; Okino, N.; Hatta, T.; Otani, H.; Naruse, C.; Suzuki, H.; Sugihara, K.; Kamimura, E.; et al. Lactosylceramide synthases encoded by B4galt5 and 6 genes are pivotal for neuronal generation and myelin formation in mice. PLoS Genet. 2018, 14, e1007545. [Google Scholar] [CrossRef]
- Ohmi, Y.; Tajima, O.; Ohkawa, Y.; Mori, A.; Sugiura, Y.; Furukawa, K.; Furukawa, K. Gangliosides play pivotal roles in the regulation of complement systems and in the maintenance of integrity in nerve tissues. Proc. Natl. Acad. Sci. USA 2009, 106, 22405–22410. [Google Scholar] [CrossRef] [Green Version]
- Ohmi, Y.; Tajima, O.; Ohkawa, Y.; Yamauchi, Y.; Sugiura, Y.; Furukawa, K.; Furukawa, K. Gangliosides are essential in the protection of inflammation and neurodegeneration via maintenance of lipid rafts: Elucidation by a series of ganglioside-deficient mutant mice. J. Neurochem. 2011, 116, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Palmano, K.; Rowan, A.; Guillermo, R.; Guan, J.; McJarrow, P. The role of gangliosides in neurodevelopment. Nutrients 2015, 7, 3891–3913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, K.; Ohmi, Y.; Ohkawa, Y.; Tokuda, N.; Kondo, Y.; Tajima, O.; Furukawa, K. Regulatory mechanisms of nervous systems with glycosphingolipids. Neurochem. Res. 2011, 36, 1578–1586. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, Y.; Furukawa, K.; Tajima, O.; Mii, S.; Honda, T.; Furukawa, K. Sensory nerve-dominant nerve degeneration and remodeling in the mutant mice lacking complex gangliosides. Neuroscience 2005, 135, 1167–1178. [Google Scholar] [CrossRef]
- Sheikh, K.A.; Sun, J.; Liu, Y.; Kawai, H.; Crawford, T.O.; Proia, R.L.; Griffin, J.W.; Schnaar, R.L. Mice lacking complex gangliosides develop Wallerian degeneration and myelination defects. Proc. Natl. Acad. Sci. USA 1999, 96, 7532–7537. [Google Scholar] [CrossRef] [Green Version]
- Kawai, H.; Allende, M.L.; Wada, R.; Kono, M.; Sango, K.; Deng, C.; Miyakawa, T.; Crawley, J.N.; Werth, N.; Bierfreund, U.; et al. Mice expressing only monosialoganglioside GM3 exhibit lethal audiogenic seizures. J. Biol. Chem. 2001, 276, 6885–6888. [Google Scholar] [CrossRef] [Green Version]
- Ichikawa, S.; Sakiyama, H.; Suzuki, G.; Hidari, K.I.; Hirabayashi, Y. Expression cloning of a cDNA for human ceramide glucoseyltransferase that catalyze the first glycosylation step of glycosphingolipid synthesis. Proc. Natl. Acad. Sci. USA 1996, 93, 4638–4643. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, T.; Wada, R.; Sasaki, T.; Deng, C.; Bierfreund, U.; Sandoff, K.; Proia, R.L. A vital role for glycosphingolipid synthesis during development and differentiation. Proc. Natl. Acad. Sci. USA 1999, 96, 9142–9147. [Google Scholar] [CrossRef] [Green Version]
- Ohmi, Y.; Ohkawa, Y.; Tajima, O.; Sugiura, Y.; Furukawa, K.; Furukawa, K. Ganglioside deficiency causes inflammation and neurodegeneration via the activation of complement system in the spinal cord. J. Neuroinflammation 2014, 11, 61. [Google Scholar] [CrossRef] [Green Version]
- McGonigal, R.; Rowan, E.G.; Greenshields, K.N.; Halstead, S.K.; Humphreys, P.D.; Rother, R.P.; Furukawa, K.; Willison, H.J. Anti-GD1a antibodies activate complement and calpain to injure distal motor nodes of Ranvier in mice. Brain 2010, 133, 1944–1960. [Google Scholar] [CrossRef] [Green Version]
- McGonigal, R.; Cunningham, M.E.; Yao, D.; Barrie, J.D.; Sankaranarayanan, S.; Fewou, S.N.; Furukawa, K.; Yednock, T.; Willison, H.J. C1q-targeted inhibition of the classical complement pathway prevents injury in a novel mouse model of acute motor axonal neuropathy. Acta Neuropathol. Commun. 2016, 4, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halstead, S.K.; Zitman, F.M.; Humphreys, P.D.; Greenshields, K.; Versschuuren, J.J.; Jacobs, B.C.; Rother, R.P.; Promp, J.J.; Willison, H.J. Eculizumab prevents anti-ganglioside antibody-mediated neuropathy in a murine model. Brain 2008, 131, 1197–1208. [Google Scholar] [CrossRef] [Green Version]
- Davidson, A.I.; Halstead, S.K.; Goodfellow, J.A.; Chavada, G.; Mallik, A.; Overell, J.; Lunn, M.P.; MaConnachie, A.; van Doorn, P.; Willison, H.J. Inhibition of complement in Guillain-Barré syndrome: The ICA-GBS study. J. Peripher. Nerv. Syst. 2017, 22, 4–12. [Google Scholar] [CrossRef]
- Furukawa, K.; Ohmi, Y.; Ohkawa, Y.; Hashimoto, N.; Kondo, Y.; Tajima, O.; Furukawa, K. Roles of glycosphingolipids in the regulation of the membrane organization and cell signaling in lipid rafts. In Membrane Organization and Lipid Rafts in the Cell and Artificial Membranes; Catala, A., Ed.; Nova Science Publishers: London, UK, 2016; pp. 129–146. [Google Scholar]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef]
- Hamamura, K.; Tsuji, M.; Hotta, H.; Ohkawa, Y.; Takahashi, M.; Shibuya, H.; Nakashima, H.; Yammauchi, Y.; Hashimoto, N.; Hattori, H.; et al. Functional activation of Src family kinase yes protein is essential for the enhanced malignant properties of human melanoma cells expressing ganglioside GD3. J. Biol. Chem. 2011, 286, 18526–18537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkawa, Y.; Miyazaki, S.; Hamamura, K.; Kambe, M.; Miyata, M.; Tajima, O.; Ohmi, Y.; Yamauchi, Y.; Furukawa, K.; Furukawa, K. Ganglioside GD3 enhances adhesion signals and augments malignant properties of melanoma cells by recruiting integrins to glycolipid-enriched microdomains. J. Biol. Chem. 2010, 285, 27213–27223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotani, N.; Gu, J.; Isaji, T.; Udaka, K.; Taniguchi, N.; Honke, K. Biochemical visualization of cell surface molecular clustering in living cells. Proc. Natl. Acad. Sci. USA 2008, 105, 7405–7409. [Google Scholar] [CrossRef] [Green Version]
- Todeschini, A.R.; Hakomori, S.I. Functional role of glycosphingolipids and gangliosides in control of cell adhesion, motility, and growth, through glycosynaptic microdomains. Biochim. Biophys. Acta 2008, 1780, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Simons, K.; Gerl, M.J. Revitalizing membrane rafts: New tools and insights. Nat. Rev. Mol. Cell Biol. 2010, 11, 688–699. [Google Scholar] [CrossRef]
- Nakashima, H.; Hamamura, K.; Houjou, T.; Taguchi, R.; Yamamoto, N.; Mitsudo, K.; Tohnai, I.; Ueda, M.; Urano, T.; Furukawa, K.; et al. Overexpression of caveolin-1 in a human melanoma cell line results in dispersion of ganglioside GD3 from lipid rafts and alteration of leading edges, leading to attenuation of malignant properties. Cancer Sci. 2007, 98, 512–520. [Google Scholar] [CrossRef]
- Fujita, A.; Cheng, J.; Hirakawa, M.; Furukawa, K.; Kusunoki, S.; Fujimoto, T. Gangliosides GM1 and GM3 in the living cell membrane form clusters susceptible to cholesterol depletion and chilling. Mol. Biol. Cell 2007, 18, 2112–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohmi, Y.; Ohkawa, Y.; Yamauchi, Y.; Tajima, O.; Furukawa, K.; Furukawa, K. Essential roles of gangliosides in the formation and maintenance of membrane microdomains in brain tissues. Neurochem. Res. 2012, 37, 1185–1191. [Google Scholar] [CrossRef]
- Yang, L.B.; Li, R.; Meri, S.; Rogers, J.; Shen, Y. Deficiency of complement defense protein CD59 may contribute to neurodegeneration in Alzheimer’s disease. J. Neurosci. 2000, 20, 7505–7509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, J.C.; Yarwood, H.; Sugars, K.; Morgan, B.P.; Davies, K.A.; Haskard, K.A. Induction of decay-accelerating factor by cytokines or the membrane-attack complex protects vascular endothelial cells against complement deposition. Blood 1999, 94, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Zurzolo, C.; Simons, K. Glycosylphosphatidylinositol-anchored proteins: Membrane organization and transport. Biochim. Biophys. Acta 2016, 1858, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.A.; Cross, H.; Proukakis, C.; Priestman, D.A.; Neville, D.C.; Reinkensmeier, G.; Wang, H.; Wiznitzer, M.; Gurtz, K.; Verganelaki, A.; et al. Infantile-onset symptomatic epilepsy syndrome caused by a homozygous loss-of-function mutation of GM3 synthase. Nat. Genet. 2004, 36, 1225–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccuto, L.; Aoki, K.; Flanagan-Steet, H.; Chen, C.F.; Fan, X.; Bartel, F.; Petukh, M.; Pittman, A.; Saul, R.; Chaubey, A.; et al. A mutation in a ganglioside biosynthetic enzyme, ST3GAL5, results in salt & pepper syndrome, a neurocutaneous disorder with altered glycolipid and glycoprotein glycosylation. Hum. Mol. Genet. 2014, 23, 418–433. [Google Scholar] [PubMed] [Green Version]
- Wang, H.; Bright, A.; Xin, B.; Bockoven, J.R.; Paller, A.S. Cutaneous dyspigmentation in patients with ganglioside GM3 synthase deficiency. Am. J. Med. Genet. A 2013, 161, 875–879. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Go, S.; Suzuki, S.; Suzuki, A.; Katori, Y.; Morlet, T.; Gottlieb, S.M.; Fujiwara, M.; Iwasaki, K.; Strauss, K.A.; et al. Ganglioside GM3 is essential for the structural integrity and function of cochlear hair cells. Hum. Mol. Genet. 2015, 24, 2796–2807. [Google Scholar] [CrossRef] [Green Version]
- Boukhris, A.; Schule, R.; Loureiro, J.L.; Lourenço, C.M.; Mundwiller, E.; Gonzalez, M.A.; Charles, P.; Gauthier, J.; Rekik, I.; Acosta Lebrigio, R.F.; et al. Alteration of ganglioside biosynthesis responsible for complex hereditary spastic paraplegia. Am. J. Hum. Genet. 2013, 93, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Harlalka, G.V.; Lehman, A.; Chioza, B.; Baple, E.L.; Maroofian, R.; Cross, H.; Sreekantan-Nair, A.; Priestman, D.A.; Al-Turki, S.; McEntagart, M.E.; et al. Mutations in B4GALNT1 (GM2 synthase) underlie a new disorder of ganglioside biosynthesis. Brain 2013, 136, 3618–3624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakil, S.M.; Monies, D.M.; Ramzan, K.; Hagos, S.; Bastaki, L.; Meyer, B.F.; Bohlega, S. Novel B4GALNT1 mutations in a complicated form of hereditary spastic paraplegia. Clin. Genet. 2014, 86, 500–501. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, R.H.; Ohmi, Y.; Ohkawa, Y.; Zhang, P.; Takano, M.; Hashimoto, N.; Okajima, T.; Furukawa, K.; Furukawa, K. Loss of enzyme activity in mutated B4GALNT1 gene products found in patients with hereditary spastic paraplegia resulted in relatively mild neurological disorders: Similarity with phenotypes of B4galnt1 knockout mice. Neuroscience 2018, 397, 94–106. [Google Scholar] [CrossRef]
- Takamiya, K.; Yamamoto, A.; Furukawa, K.; Zhao, J.; Fukumoto, S.; Yamashiro, S.; Okada, M.; Haraguchi, M.; Shin, M.; Kishikawa, M.; et al. Complex gangliosides are essential in spermatogenesis of mice: Possible roles in the transport of testosterone. Proc. Natl. Acad. Sci. USA 1998, 95, 12147–12152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitajima, K.; Varki, N.; Sato, C. Advanced Technologies in Sialic Acid and Sialoglycoconjugate Analysis. Top. Curr. Chem. 2015, 367, 75–103. [Google Scholar] [PubMed]
- Cavdarli, S.; Dewald, J.H.; Yamakawa, N.; Guérardel, Y.; Terme, M.; Le Doussal, J.M.; Delannoy, P.; Groux-Degroote, S. Identification of 9-O-acetyl-N-acetyl-neuraminic acid (Neu5,9Ac2) as main O-acetylated sialic acid species of GD2 in breast cancer cells. Glycoconj. J. 2019, 36, 79–90. [Google Scholar] [CrossRef]
- Nakayama, H.; Nagafuku, M.; Suzuki, A.; Iwabuchi, K.; Inokuchi, J.I. The regulatory roles of glycosphingolipid-enriched lipid rafts in immune systems. FEBS Lett. 2018, 592, 3921–3942. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Ikeda, K.; Tokuda, N.; Nishitani, C.; Ohto, U.; Akashi, T.S.; Ito, Y.; Uchikawa, M.; Kuroki, Y.; Taguchi, R.; et al. TLR4-MD-2 complex is negatively regulated by an endogenous ligand, globotetraosylceramide. Proc. Natl. Acad. Sci. USA 2013, 110, 4714–4719. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, N.; Ito, S.; Tsuchida, A.; Bhuiyan, R.H.; Okajima, T.; Yamamoto, A.; Furukawa, K.; Ohmi, Y.; Furukawa, K. The ceramide moiety of disialoganglioside (GD3) is essential for GD3 recognition by the sialic acid-binding lectin SIGLEC7 on the cell surface. J. Biol. Chem. 2019, 294, 10833–10845. [Google Scholar] [CrossRef]
- Yao, D.; McGonigal, R.; Barrie, J.A.; Cappell, J.; Cunningham, M.E.; Meehan, G.R.; Fewou, S.N.; Edgar, J.M.; Rowan, E.; Ohmi, Y.; et al. Neuronal expression of GalNAc transferase is sufficient to prevent the age-related neurodegenerative phenotype of complex ganglioside deficient mice. J. Neurosci. 2014, 34, 880–891. [Google Scholar] [CrossRef]
- Komura, N.; Suzuki, K.G.; Ando, H.; Konishi, M.; Koikeda, M.; Imamura, A.; Chadda, R.; Fujiwara, T.K.; Tsuboi, H.; Sheng, R.; et al. Ultra-high-resolution imaging of gangliosides in GEM/rafts Raft-based interactions of gangliosides with a GPI-anchored receptor. Nat. Chem. Biol. 2016, 12, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Lingwood, D.; Simons, K. Lipid rafts as a membrane-organizing principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KO Gene | Glc-Cer Syn | GM3 Syn | GD3 Syn | GM2/GD2 Syn | DKO 1) |
|---|---|---|---|---|---|
| Lost structures | all glyco- sphingolipids | ganglio-series (a-, b-, c-) | b-series (and c-series) | all complex gangliosides(inc. asialo-series) | all complex gangliosides(inc. asialo-series) |
| Remaining structures | asialo-series | a-series and asialo-series | GM3, GD3 (and GT3) | GM3 | |
| Gal-Cer and sulfatedes | Gal-Cer and sulfatedes neutral glycolipids | Gal-Cer and sulfatedes neutral glycolipids | Gal-Cer and sulfatedes neutral glycolipids | Gal-Cer and sulfatedes neutral glycolipids | |
| Phenotypes |  | | | | |
| Embryonal lethal | No apparent abnormalities | Mild abnormalities | Gradual abnormalities | Neurodegeneration from early phase | |
| Remarks | Auditory disorder | Poor repair Low serum leptin | Male infertileity Low serum testostelone Wallerian degen 2) | Refractory skin lesion Auditory shock |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furukawa, K.; Ohmi, Y.; Yesmin, F.; Tajima, O.; Kondo, Y.; Zhang, P.; Hashimoto, N.; Ohkawa, Y.; Bhuiyan, R.H.; Furukawa, K. Novel Molecular Mechanisms of Gangliosides in the Nervous System Elucidated by Genetic Engineering. Int. J. Mol. Sci. 2020, 21, 1906. https://doi.org/10.3390/ijms21061906
Furukawa K, Ohmi Y, Yesmin F, Tajima O, Kondo Y, Zhang P, Hashimoto N, Ohkawa Y, Bhuiyan RH, Furukawa K. Novel Molecular Mechanisms of Gangliosides in the Nervous System Elucidated by Genetic Engineering. International Journal of Molecular Sciences. 2020; 21(6):1906. https://doi.org/10.3390/ijms21061906
Chicago/Turabian StyleFurukawa, Koichi, Yuhsuke Ohmi, Farhana Yesmin, Orie Tajima, Yuji Kondo, Pu Zhang, Noboru Hashimoto, Yuki Ohkawa, Robiul H. Bhuiyan, and Keiko Furukawa. 2020. "Novel Molecular Mechanisms of Gangliosides in the Nervous System Elucidated by Genetic Engineering" International Journal of Molecular Sciences 21, no. 6: 1906. https://doi.org/10.3390/ijms21061906