BAK1 Mediates Light Intensity to Phosphorylate and Activate Catalases to Regulate Plant Growth and Development

Abstract
:1. Introduction
2. Results
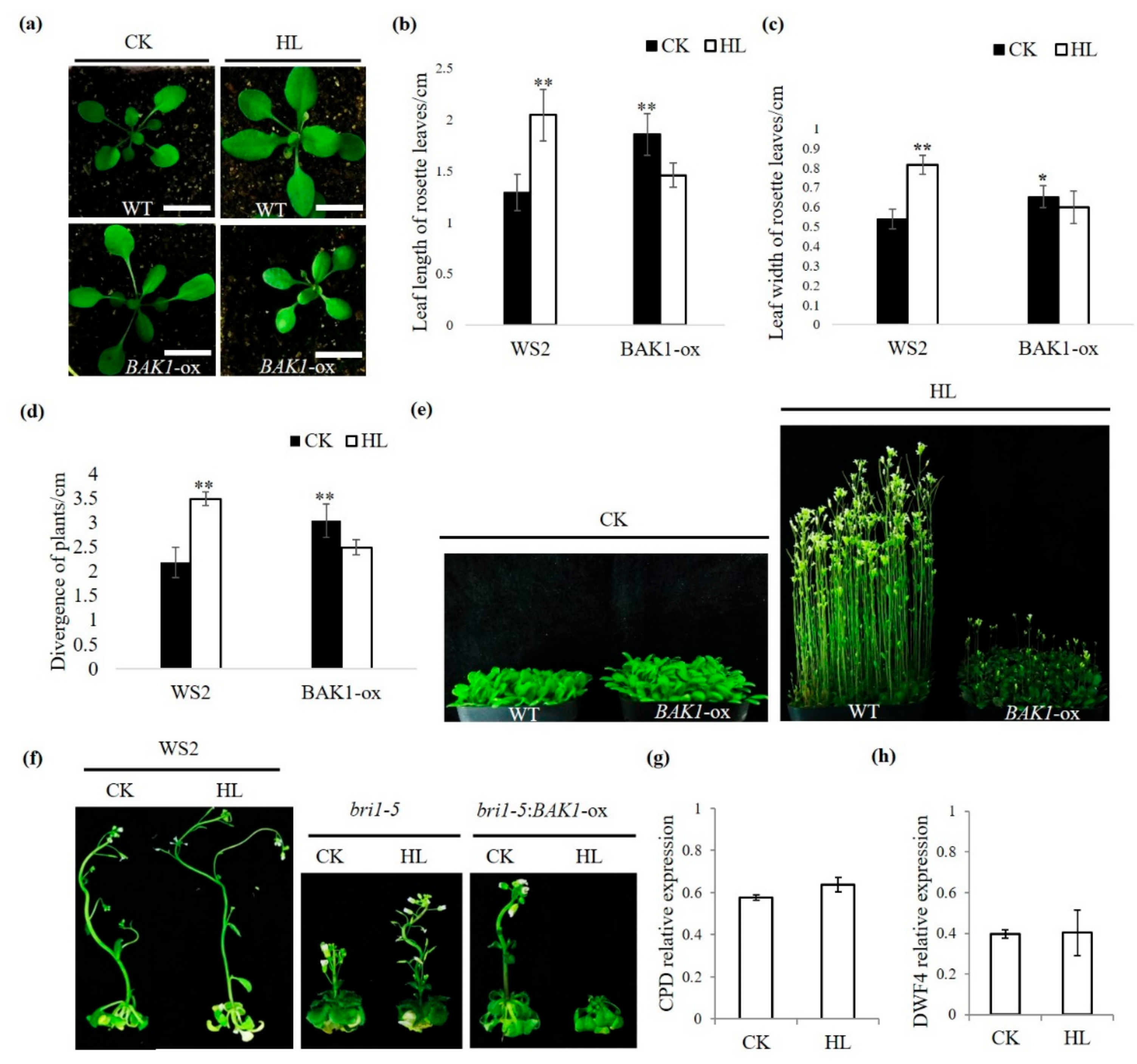
2.1. Overexpression of BAK1 Suppresses the High Light-Promoted Plant Growth
2.2. The Suppression of BAK1 Overexpression on High Light-Promoted Plant Growth is Independent of the BR Signaling
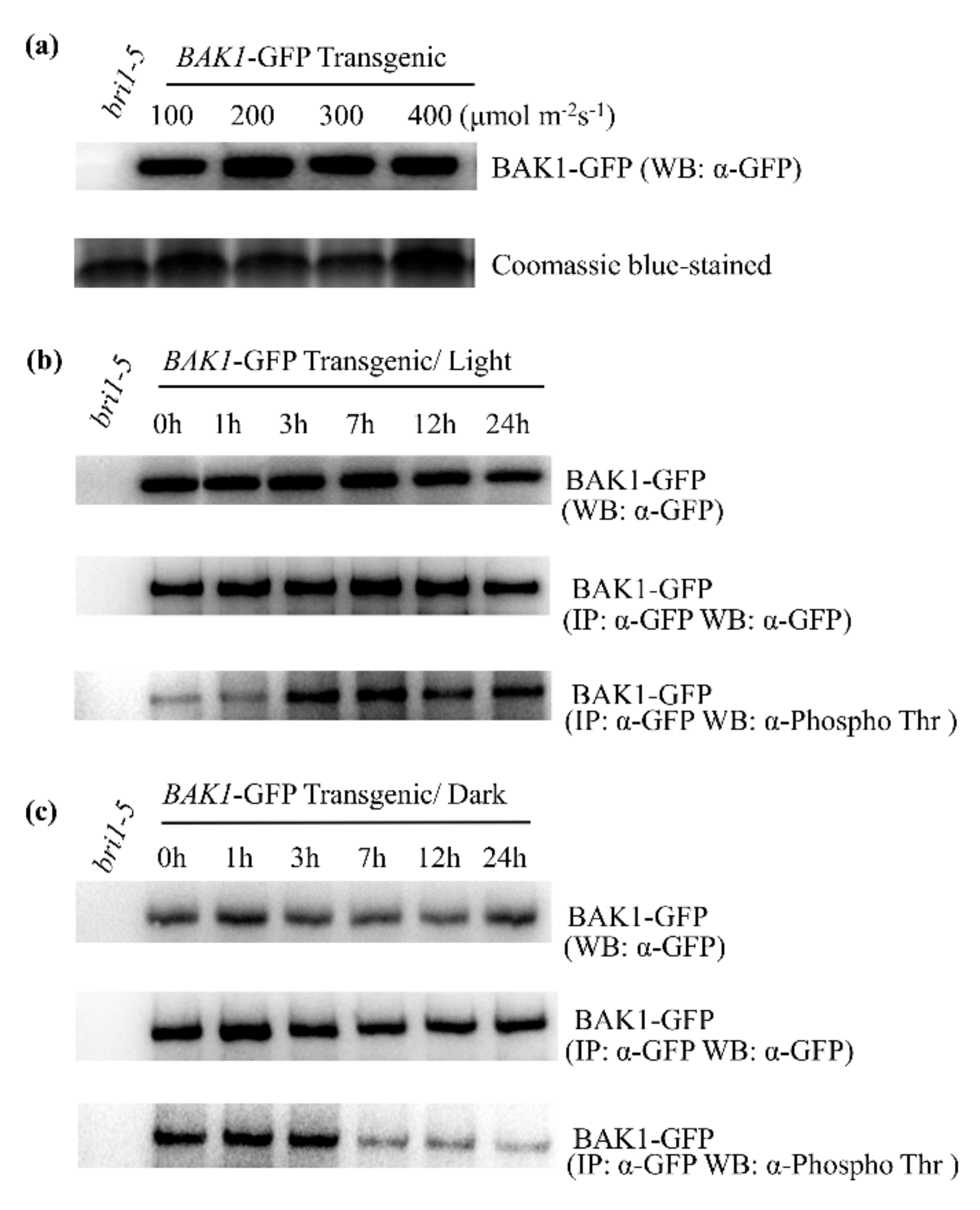
2.3. High Light Enhances the BAK1 Phosphorylation Level in Plant Cells
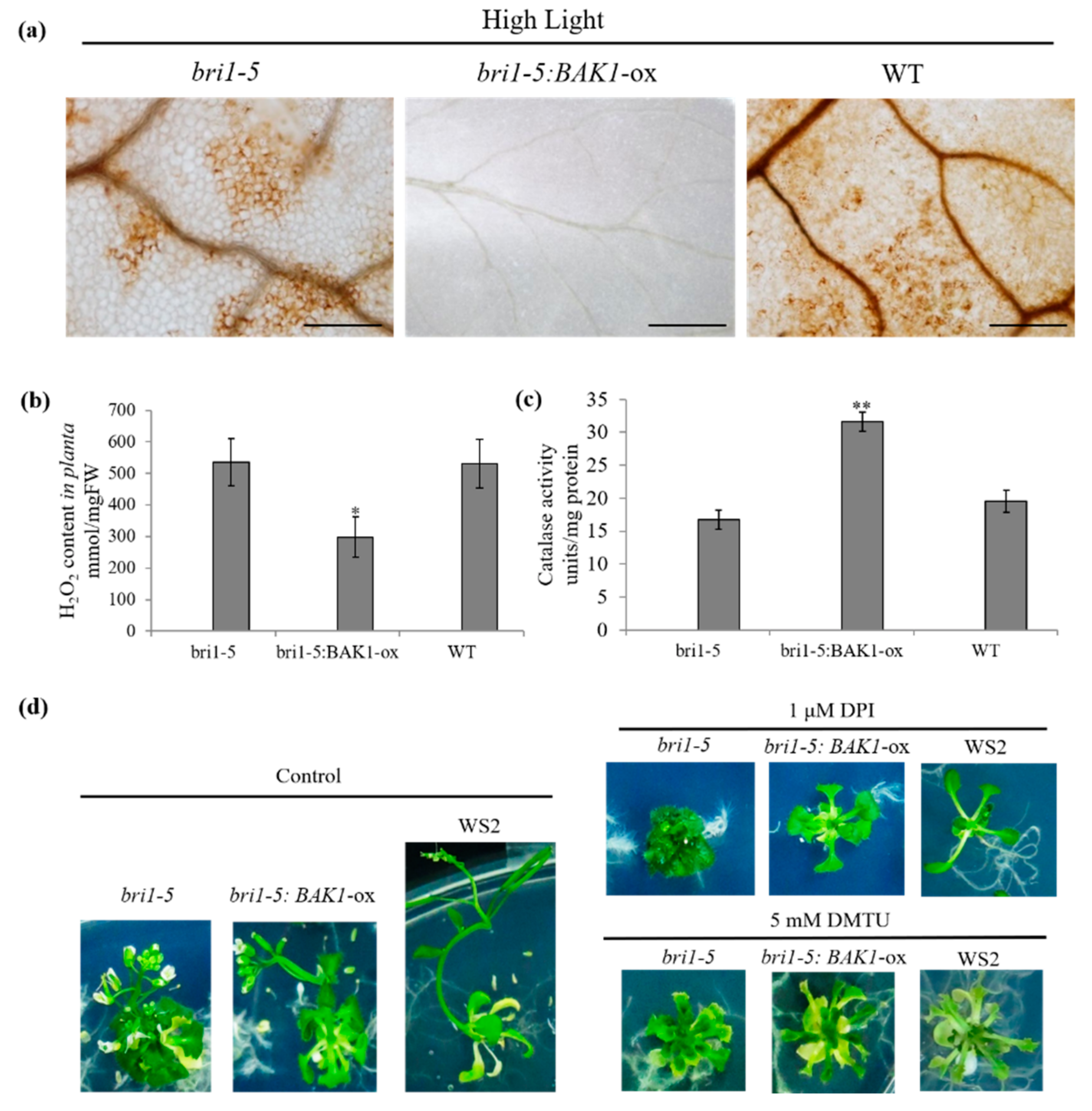
2.4. Overexpression of BAK1 Enhances the Catalase Activity, Resulting in a Low Level of H2O2 in Plant Cells
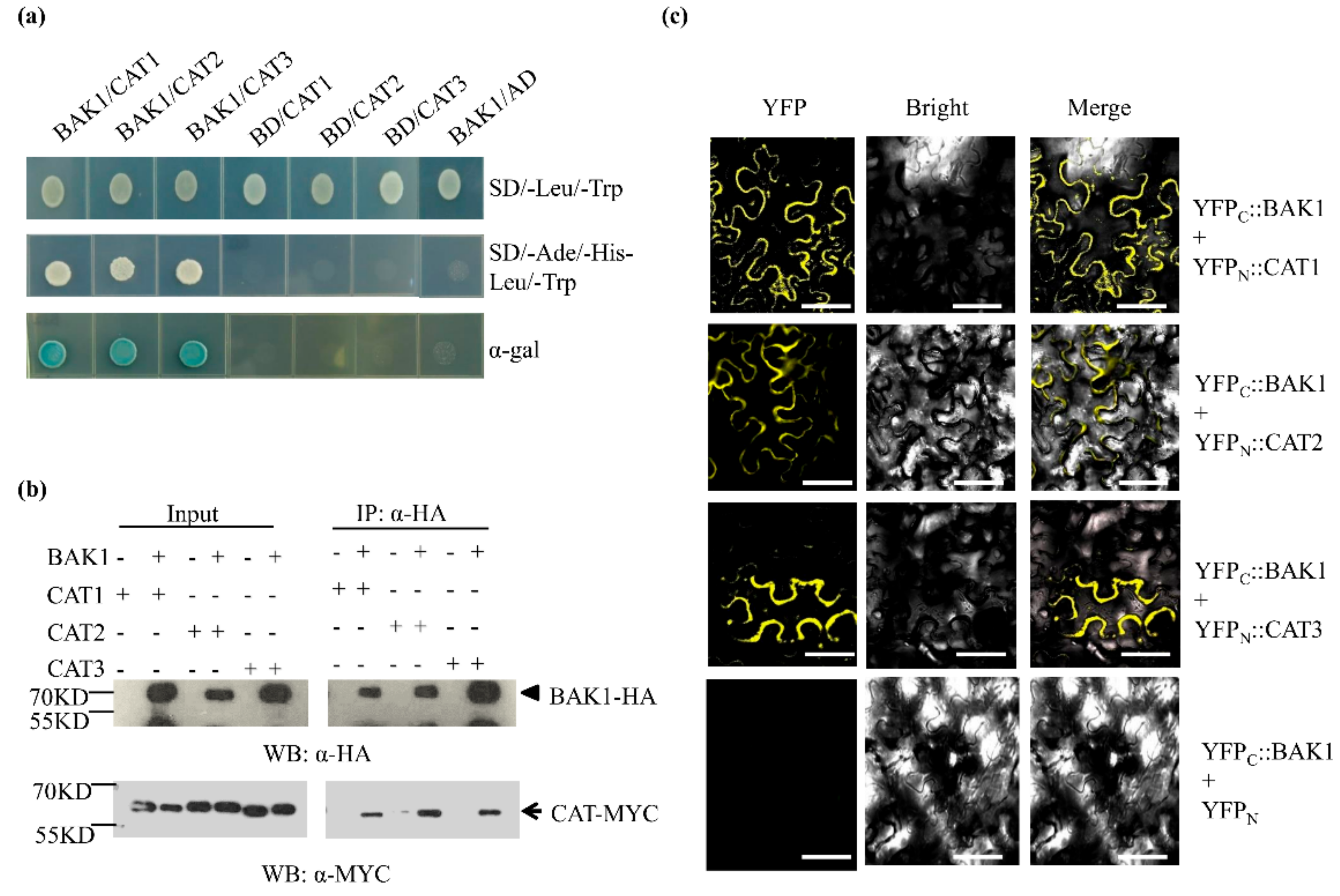
2.5. BAK1 Interacts with Three Catalases
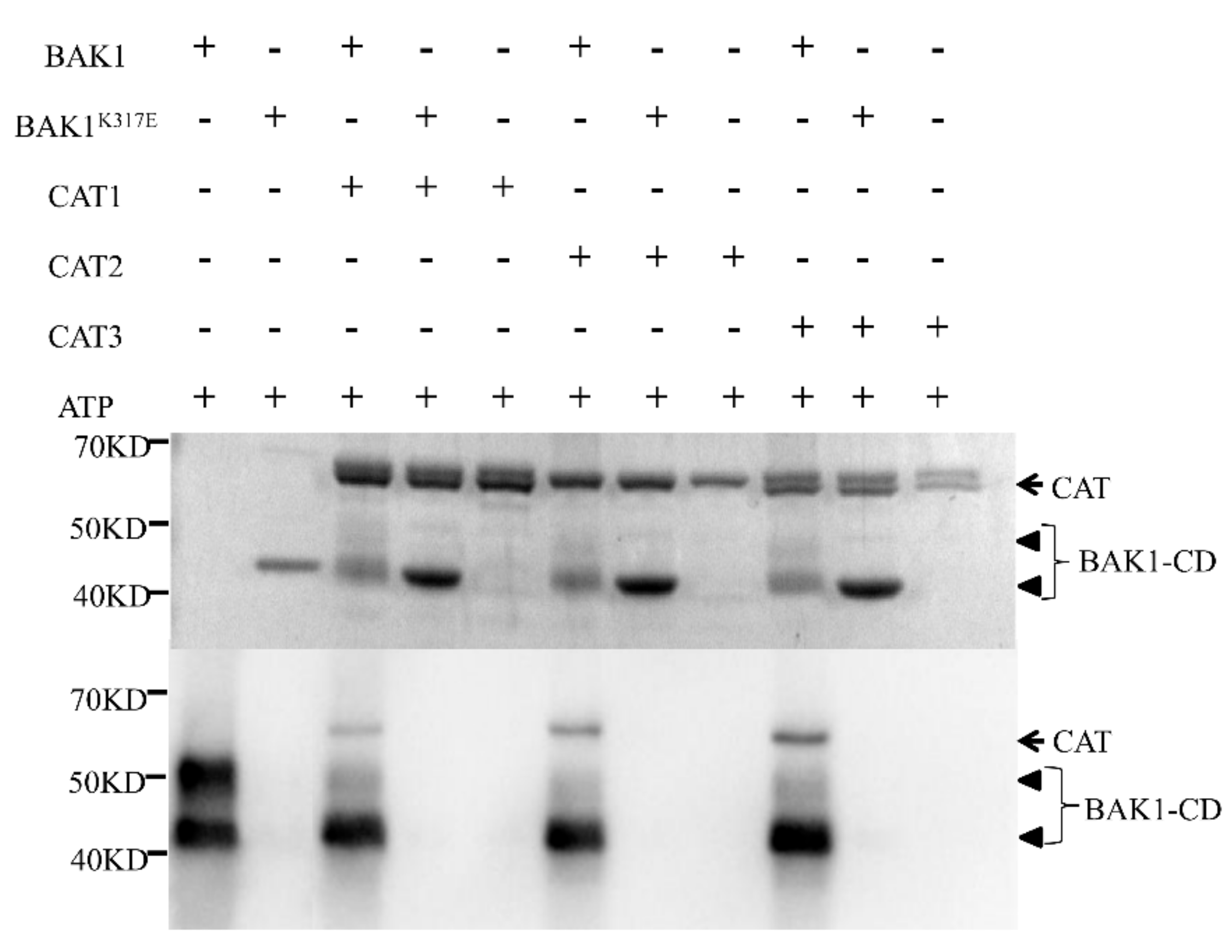
2.6. BAK1 Phosphorylates Three Catalases
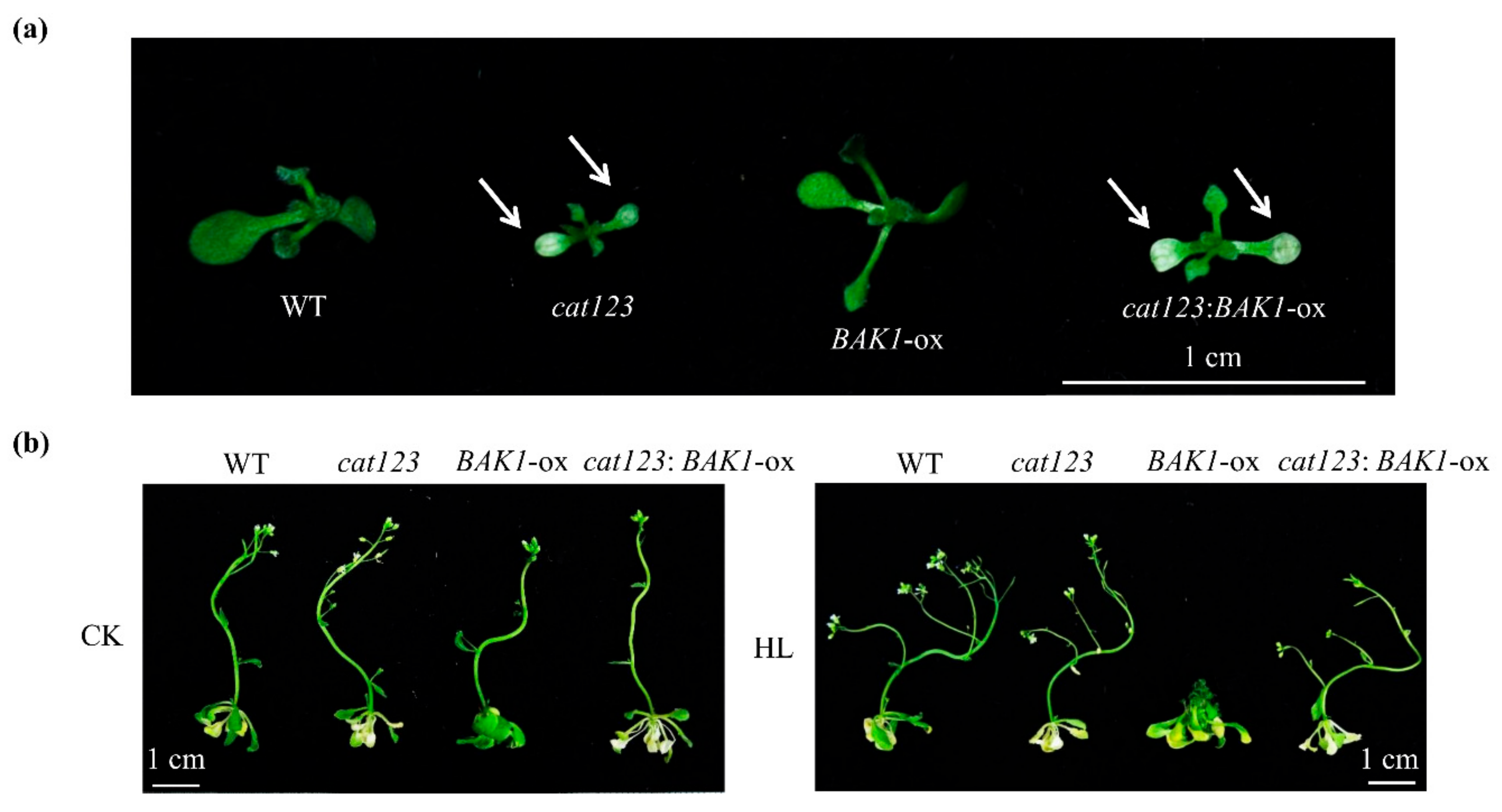
2.7. BAK1 Overexpression Effect on Suppressing High Light-Promoted Growth is Abolished by Knockout of Catalase Genes
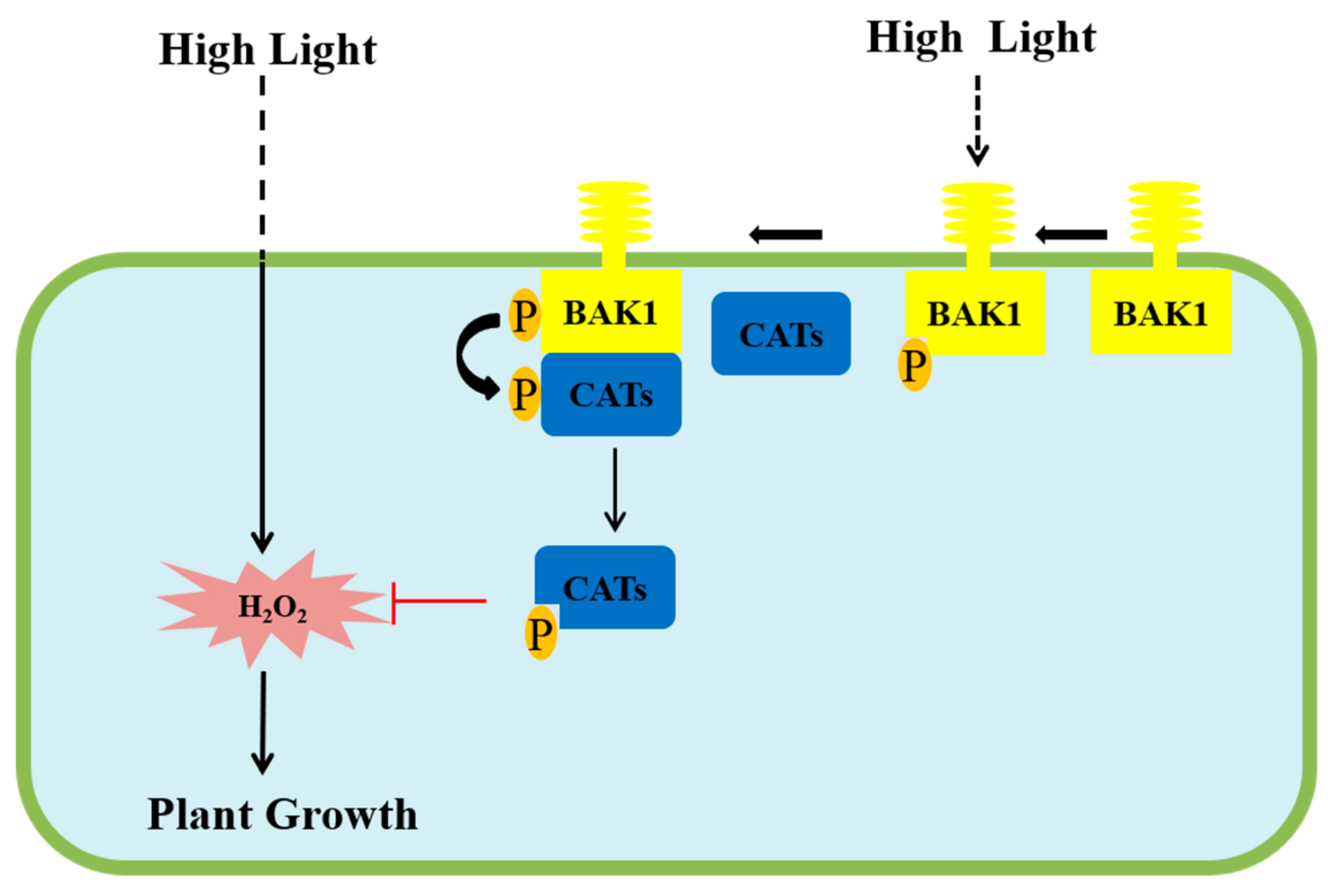
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Light Treatment Conditions
4.3. Statistical Analysis
4.4. Protein Phosphorylation Level Assay In Vivo
4.5. Western Blot Analysis
4.6. Mass Spectrometry
4.7. Physiological Biochemical Index Measurement in Planta
4.8. RNA Extraction and qRT-PCR
4.9. Yeast Two-Hybrid Assays
4.10. Co-IP Assay
4.11. Subcellular Localizations
4.12. BiFC assays
4.13. In Vitro Kinase Assay
4.14. Generation of cat123 Triple Mutants
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kami, C.; Lorrain, S.; Hornitschek, P.; Fankhauser, C. Light-regulated plant growth and development. Curr. Top. Dev. Biol. 2010, 91, 29–66. [Google Scholar]
- Feng, P.Q.; Guo, H.L.; Chi, W.; Chai, X.; Sun, X.W.; Xu, X.M.; Ma, J.F.; Rochaix, J.D.; Leister, D.; Wang, H.Y.; et al. Chloroplast retrograde signal regulates flowering. Proc. Natl. Acad. Sci. USA 2016, 113, 10708–10713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, M.G.; Beggs, C.J.; Jabben, M.; Schafer, E. Hypocotyl growth in Sinapis alba L: The roles of light quality and quantity. Plant Cell Environ. 1982, 5, 45–51. [Google Scholar]
- Bailey, S.; Walters, R.G.; Jansson, S.; Horton, P. Acclimation of Arabidopsis thaliana to the light environment: The existence of separate low light and high light responses. Planta 2001, 213, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Nagel, K.A.; Schurr, U.; Walter, A. Dynamics of root growth stimulation in Nicotiana tabacum in increasing light intensity. Plant Cell Environ. 2006, 29, 1936–1945. [Google Scholar] [CrossRef]
- Marin, I.C.; Loef, I.; Bartetzko, L.; Searle, I.; Coupland, G.; Stitt, M.; Osuna, D. Nitrate regulates floral induction in Arabidopsis, acting independently of light, gibberellin and autonomous pathways. Planta 2011, 233, 539–552. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef]
- Chen, Z.P.; Gu, Q.; Yu, X.L.; Huang, L.Q.; Xu, S.; Wang, R.; Shen, W.; Shen, W.B. Hydrogen peroxide acts downstream of melatonin to induce lateral root formation. Ann. Bot. 2018, 121, 1127–1136. [Google Scholar] [CrossRef]
- Bieker, S.; Riester, L.; Stahl, M.; Franzaring, J.; Zentgraf, U. Senescence-specific alteration of hydrogen peroxide levels in Arabidopsis thaliana and oilseed rape spring variety Brassica napus L. cv. Mozart. J. Integr. Plant Biol. 2012, 54, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Betsuyaku, S.; Takahashi, F.; Kinoshita, A.; Miwa, H.; Shinozaki, K.; Fukuda, H.; Sawa, S. Mitogen-activated protein kinase regulated by the CLAVATA receptors contributes to shoot apical meristem homeostasis. Plant Cell Physiol. 2011, 52, 14–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohm, H.; Albert, I.; Fan, L.; Reinhard, A.; Nurnberger, T. Immune receptor complexes at the plant cell surface. Curr. Opin. Plant Biol. 2014, 20, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.R.; Liang, Y.; Tanaka, K.; Nguyen, C.T.; Jedrzejczak, R.P.; Joachimiak, A.; Stacey, G. The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin-induced complex with related kinase CERK1. eLife 2014, 3, e03766. [Google Scholar] [CrossRef] [PubMed]
- Lehti-Shiu, M.D.; Zou, C.; Hanada, K.; Shiu, S.H. Evolutionary history and stress regulation of plant receptor-like kinase/pelle genes. Plant Physiol. 2009, 150, 12–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saijo, Y. ER quality control of immune receptors and regulators in plants. Cell Microbiol. 2010, 12, 716–724. [Google Scholar] [CrossRef]
- Li, J.; Wen, J.; Lease, K.A.; Doke, J.T.; Tax, F.E.; Walker, J.C. BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling. Cell 2002, 110, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Nam, K.H.; Li, J. BRI1/BAK1, a receptor kinase pair mediating brassinosteroid signaling. Cell 2002, 110, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Belkhadir, Y.; Yang, L.; Hetzel, J.; Dangl, J.L.; Chory, J. The growth–defense pivot: Crisis management in plants mediated by LRR-RK surface receptors. Trends Biochem. Sci. 2014, 39, 447–456. [Google Scholar] [CrossRef] [Green Version]
- Chinchilla, D.; Shan, L.; He, P.; de Vries, S.; Kemmerling, B. One for all: The receptor-associated kinase BAK1. Trends Plant Sci. 2009, 14, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.F.; Kota, U.; He, K.; Blackburn, K.; Li, J.; Goshe, M.B.; Huber, S.C.; Clouse, S.D. Sequential transphosphorylation of the BRI1/BAK1 receptor kinase complex impacts early events in brassinosteroid signaling. Dev. Cell 2008, 15, 220–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, K.; Xu, S.B.; Li, J. BAK 1 Directly Regulates Brassinosteroid Perception and BRI 1 Activation. J. Integr. Plant Biol. 2013, 55, 1264–1270. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.Z.; Chen, X.; Mang, H.; Liu, C.L.; Yu, X.; Gao, X.Q.; Torii, K.U.; He, P.; Shan, L.B. Differential Function of Arabidopsis SERK Family Receptor-like Kinases in Stomatal Patterning. Curr. Biol. 2015, 25, 2361–2372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, J.; Henzler, C.; Hothorn, M. Molecular mechanism for plant steroid receptor activation by somatic embryogenesis co-receptor kinases. Science 2013, 341, 889–892. [Google Scholar] [CrossRef]
- Kim, B.H.; Kim, S.Y.; Nam, K.H. Assessing the diverse functions of BAK1 and its homologs in arabidopsis, beyond BR signaling and PTI responses. Mol. Cells 2013, 35, 7–16. [Google Scholar] [CrossRef] [Green Version]
- He, K.; Gou, X.P.; Yuan, T.; Lin, H.H.; Asami, T.; Yoshida, S.; Russell, S.D.; Li, J. BAK1 and BKK1 regulate brassinosteroid-dependent growth and brassinosteroid-independent cell-death pathways. Curr. Biol. 2007, 17, 1109–1115. [Google Scholar] [CrossRef] [Green Version]
- Du, J.B.; Gao, Y.; Zhan, Y.Y.; Zhang, S.S.; Wu, Y.J.; Xiao, Y.; Zou, B.; He, K.; Gou, X.P.; Li, G.J.; et al. Nucleocytoplasmic trafficking is essential for BAK 1-and BKK 1-mediated cell-death control. Plant J. 2016, 85, 520–531. [Google Scholar] [CrossRef] [Green Version]
- Rudolf, M.; Machettira, A.B.; Groß, L.E.; Weber, K.L.; Bolte, K.; Bionda, T.; Sommer, M.S.; Maier, U.G.; Weber, A.P.M.; Schleiff, E.; et al. In vivo function of Tic22, a protein import component of the intermembrane space of chloroplasts. Mol. Plant 2013, 6, 817–829. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, T.; Fujioka, S.; Choe, S.; Takatsuto, S.; Yoshida, S.; Yuan, H.; Feldmann, K.A.; Tax, F.E. Brassinosteroid-insensitive dwarf mutants of Arabidopsis accumulate brassinosteroids. Plant Physiol. 1999, 121, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Yin, Y.H.; Fei, S.Z. Down-regulation of BdBRI1, a putative brassinosteroid receptor gene produces a dwarf phenotype with enhanced drought tolerance in Brachypodium distachyon. Plant Sci. 2015, 234, 163–173. [Google Scholar] [CrossRef]
- Frugoli, J.A.; Zhong, H.H.; Nuccio, M.L.; McCourt, P.; McPeek, M.A.; Thomas, T.L.; McClung, C.R. Catalase is encoded by a multigene family in Arabidopsis thaliana (L) Heynh. Plant Physiol. 1996, 112, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.Y.; Wang, P.C.; Chen, J.; Song, C.P. Comprehensive functional analysis of the catalase gene family in Arabidopsis thaliana. J. Integr. Plant Biol. 2008, 50, 1318–1326. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.Q.; Liu, S.; Yuan, H.M.; Li, J.; Yan, D.W.; Zhang, J.F.; Lu, Y.T. Functional comparison of catalase genes in the elimination of photorespiratory H2O2 using promoter-and 3’-untranslated region exchange experiments in the Arabidopsis cat2 photorespiratory mutant. Plant Cell Environ. 2010, 33, 1656–1670. [Google Scholar] [CrossRef]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [Green Version]
- Gou, X.P.; Yin, H.J.; He, K.; Du, J.B.; Yi, J.; Xu, S.B.; Lin, H.H.; Clouse, S.D.; Li, J. Genetic evidence for an indispensable role of somatic embryogenesis receptor kinases in brassinosteroid signaling. PLoS Genet. 2012, 8, e1002452. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.Z.; Wu, Y.J.; Gao, Y.; Li, M.Z.; Yin, H.J.; Lv, M.H.; Zhao, J.X.; Li, J.; He, K. Somatic embryogenesis receptor-like kinase 5 in the ecotype Landsberg erecta of Arabidopsis is a functional RD LRR-RLK in regulating brassinosteroid signaling and cell death control. Front. Plant Sci. 2015, 6, 852. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Ferreras, A.; Kiss-Papp, M.; Jehle, A.K.; Felix, G.; Chinchilla, D. An overdose of the Arabidopsis coreceptor BRASSINOSTEROID INSENSITIVE1-ASSOCIATED RECEPTOR KINASE1 or its ectodomain causes autoimmunity in a SUPPRESSOR OF BIR1-1-dependent manner. Plant Physiol. 2015, 168, 1106–1121. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Yang, H.; Mockler, T.C.; Lin, C.J.S. Regulation of flowering time by Arabidopsis photoreceptors. Science 1998, 279, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Zoratti, L.; Karppinen, K.; Luengo Escobar, A.; Häggman, H.; Jaakola, L. Light-controlled flavonoid biosynthesis in fruits. Front. Plant Sci. 2014, 5, 534. [Google Scholar] [CrossRef]
- Xia, X.J.; Gao, C.J.; Song, L.X.; Zhou, Y.H.; Shi, K.; Yu, J.Q. Role of H2O2 dynamics in brassinosteroid-induced stomatal closure and opening in S olanum lycopersicum. Plant Cell Environ. 2014, 37, 2036–2050. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.J.; Wang, Y.J.; Zhou, Y.H.; Tao, Y.; Mao, W.H.; Shi, K.; Asami, T.; Chen, Z.; Yu, J.Q. Reactive oxygen species are involved in brassinosteroid-induced stress tolerance in cucumber. Plant Physiol. 2009, 150, 801–814. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wang, J.; Li, X.; Xia, X.J.; Zhou, Y.H.; Shi, K.; Chen, Z.X.; Yu, J.Q. H2O2 mediates the crosstalk of brassinosteroid and abscisic acid in tomato responses to heat and oxidative stresses. J. Exp. Bot. 2014, 65, 4371–4383. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Heinlein, C.; Orendi, G.; Zentgraf, U. Senescence-specific regulation of catalases in Arabidopsis thaliana (L.) Heynh. Plant Cell Environ. 2006, 29, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Shiu, S.H.; Karlowski, W.M.; Pan, R.; Tzeng, Y.H.; Mayer, K.F.; Li, W.H. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell 2004, 16, 1220–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.D.; Li, L.; Macho, A.P.; Han, Z.F.; Hu, Z.H.; Zipfel, C.; Zhou, J.M.; Chai, J.J. Structural basis for flg22-induced activation of the Arabidopsis FLS2-BAK1 immune complex. Science 2013, 342, 624–628. [Google Scholar] [CrossRef]
- Schwessinger, B.; Roux, M.; Kadota, Y.; Ntoukakis, V.; Sklenar, J.; Jones, A.; Zipfel, C. Phosphorylation-dependent differential regulation of plant growth, cell death, and innate immunity by the regulatory receptor-like kinase BAK1. PLoS Genet. 2011, 7, e1002046. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.J.; Li, X.D.; Ratnasekera, D.; Wang, C.; Liu, W.X.; Song, L.F.; Zhang, W.Z.; Wu, W.H. Arabidopsis CALCIUM-DEPENDENT PROTEIN KINASE8 and CATALASE3 Function in Abscisic Acid-Mediated Signaling and H2O2 Homeostasis in Stomatal Guard Cells under Drought Stress. Plant Cell 2015, 27, 1445–1460. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.B.; Liu, C.; Tang, D.Y.; Yan, L.; Wang, D.; Yang, Y.Z.; Gui, J.S.; Zhao, X.Y.; Li, L.G.; Tang, X.D.; et al. The Receptor-Like Cytoplasmic Kinase STRK1 Phosphorylates and Activates CatC, Thereby Regulating H2O2 Homeostasis and Improving Salt Tolerance in Rice. Plant Cell 2018, 30, 1100–1118. [Google Scholar] [CrossRef] [Green Version]
- Miao, M.; Niu, X.L.; Kud, J.; Du, X.R.; Avila, J.; Devarenne, T.P.; Kuhl, J.C.; Liu, Y.S.; Xiao, F.M. The ubiquitin ligase SEVEN IN ABSENTIA (SINA) ubiquitinates a defense-related NAC transcription factor and is involved in defense signaling. New Phytol. 2016, 211, 138–148. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Li, C.; Li, Q.; Wang, Q.N.; Huang, S.H.; Zhang, Y.F.; Wang, X.F. Functional divergence of BAK1 genes from Brassica rapa in regulating plant architecture. Genet. Mol. Res. 2015, 14, 14587–14596. [Google Scholar] [CrossRef]
- Waadt, R.; Kudla, J. In planta visualization of protein interactions using bimolecular fluorescence complementation (BiFC). CSH Protoc. 2008, 2008, pdb.prot4995. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.L.; Dong, L.; Wang, Z.P.; Zhang, H.Y.; Han, C.Y.; Liu, B.; Wang, X.C.; Chen, Q.J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.X.; Ding, Y.L.; Shi, Y.T.; Zhang, X.Y.; Gong, Z.Z.; Yang, S.H. The cbfs triple mutants reveal the essential functions of CBFs in cold acclimation and allow the definition of CBF regulons in Arabidopsis. New Phytol. 2016, 212, 345–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Identified Site |
|---|---|
| BAK1-CD+CAT1 | |
| GPILLEDyHLLEK | Y45 |
| GFFEVtHDITQLtSADFLR | T78, T85 |
| GFFEVTHDITQLTsADFLR | S86 |
| IFSYADsQR | S351 |
| HAEKYPTtPIVCSGNR | T409 |
| FVEALSEPRVtHEIR | T460 |
| SIWISYWSQADKsLGQK | S477 |
| BAK1-CD+CAT2 | |
| YRPASSYNSPFFTTNSGAPVWNNNsSMTVGPR | S30 |
| HMDGsGVNTYMLINK | S205 |
| VFSYADtQR | T352 |
| DEEVNYFPsRYDQVR | S395 |
| HAEKYPtPPAVCSGK | T408 |
| BAK1-CD+CAT3 | |
| YRPSSAYNAPFYTTNGGAPVSNNISsLTIGER | S31 |
| GFFEVTHDIsNLTCADFLR | S82 |
| GFFEVTHDIsNLTCADFLRAPGVQTPVIVR | S82 |
| FYtREGNFDLVGNNTPVFFIR | T128 |
| LFIQtMDPADEDKFDFDPLDVTK | T273 |
| DEEINYYPsKFDPVR | S395 |
| VPTPTNsYTGIR | S412 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Li, C.; Ren, H.; Zhao, T.; Li, Q.; Wang, S.; Zhang, Y.; Xiao, F.; Wang, X. BAK1 Mediates Light Intensity to Phosphorylate and Activate Catalases to Regulate Plant Growth and Development. Int. J. Mol. Sci. 2020, 21, 1437. https://doi.org/10.3390/ijms21041437
Zhang S, Li C, Ren H, Zhao T, Li Q, Wang S, Zhang Y, Xiao F, Wang X. BAK1 Mediates Light Intensity to Phosphorylate and Activate Catalases to Regulate Plant Growth and Development. International Journal of Molecular Sciences. 2020; 21(4):1437. https://doi.org/10.3390/ijms21041437
Chicago/Turabian StyleZhang, Shan, Cheng Li, Haihua Ren, Tong Zhao, Qi Li, Shufen Wang, Yanfeng Zhang, Fangming Xiao, and Xiaofeng Wang. 2020. "BAK1 Mediates Light Intensity to Phosphorylate and Activate Catalases to Regulate Plant Growth and Development" International Journal of Molecular Sciences 21, no. 4: 1437. https://doi.org/10.3390/ijms21041437