The Floral Repressor GmFLC-like Is Involved in Regulating Flowering Time Mediated by Low Temperature in Soybean

 , and
, and
Abstract
:1. Introduction
2. Results
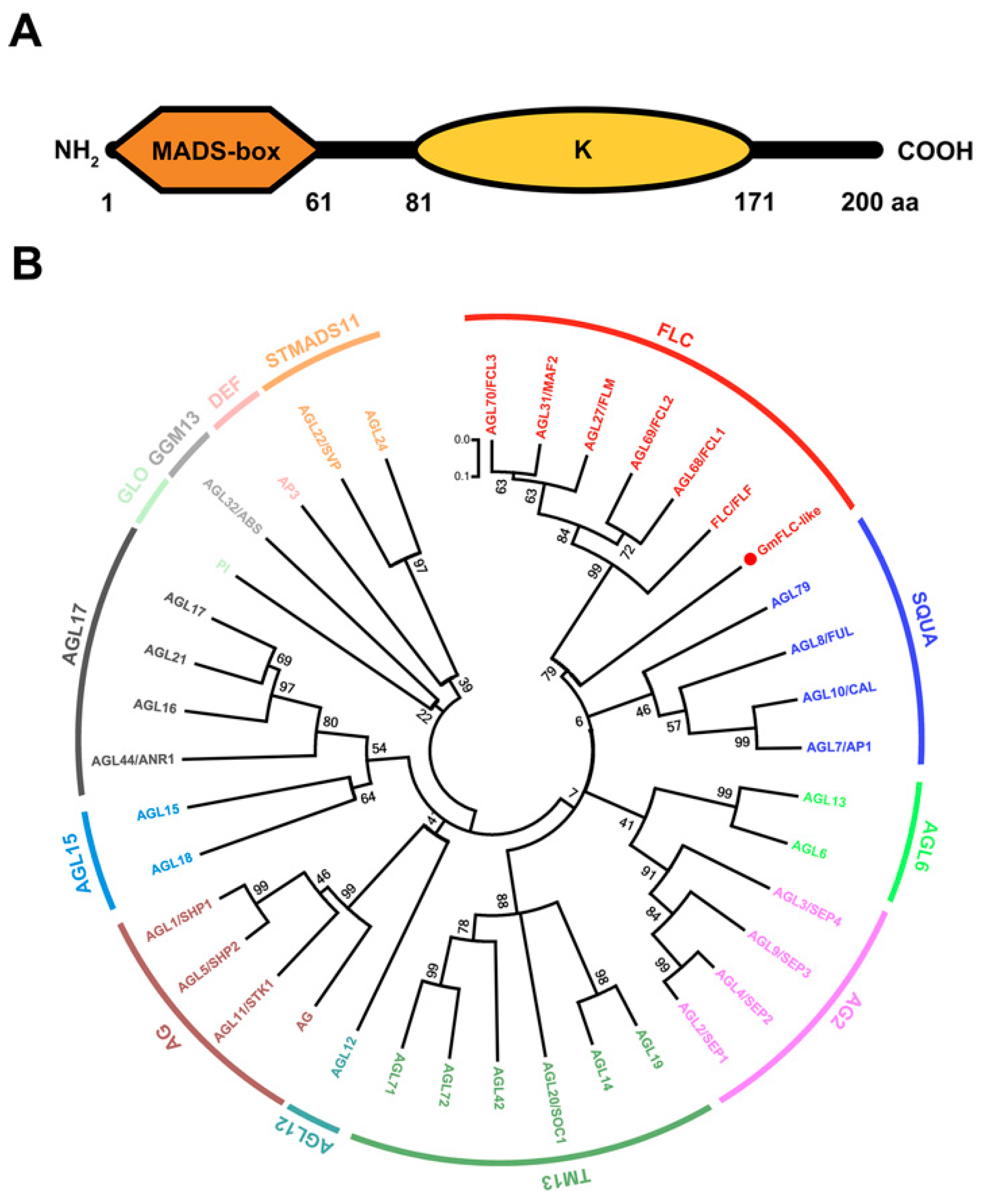
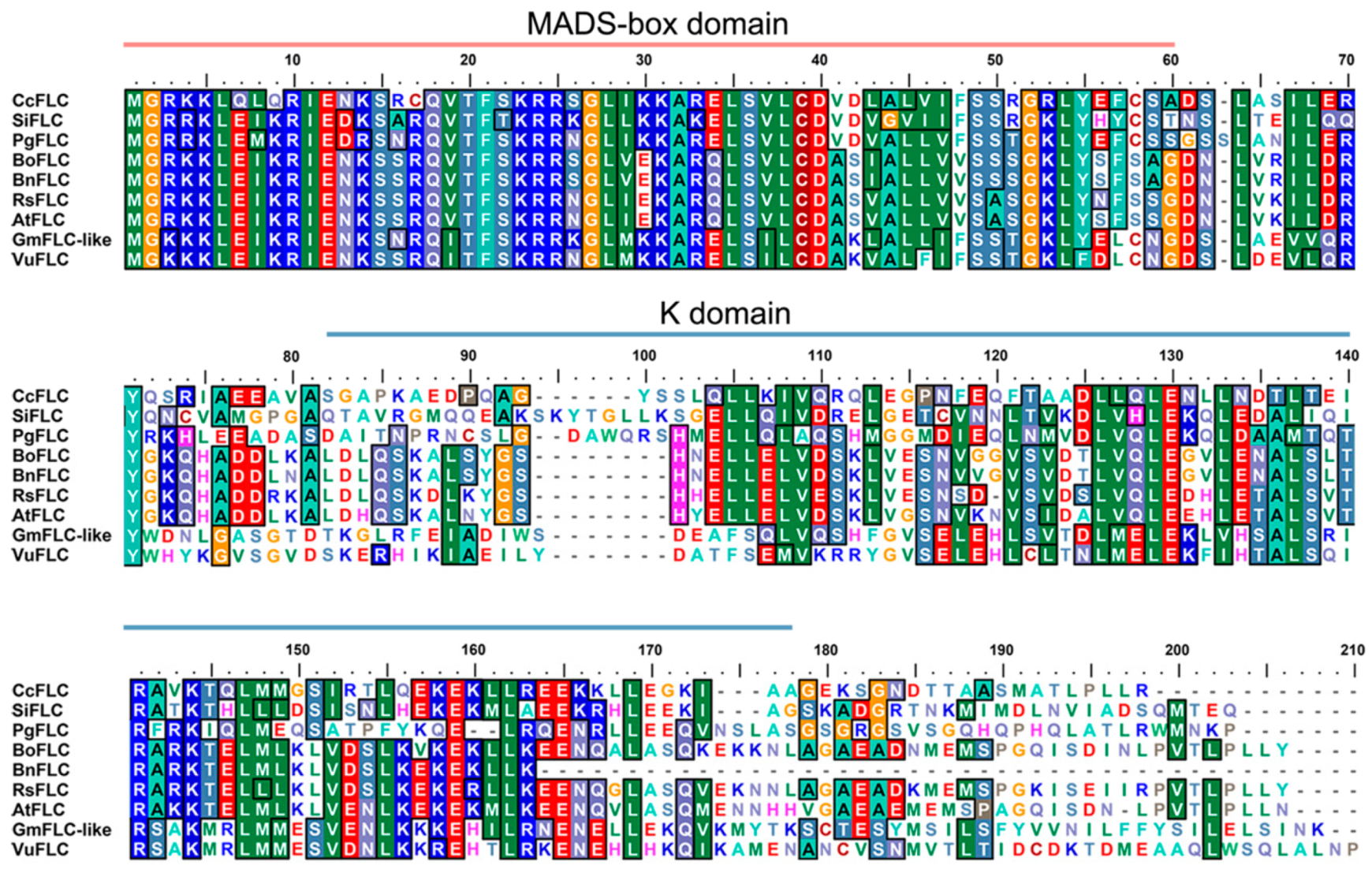
2.1. Glyma05g28130 Is a Homologue of AtFLC
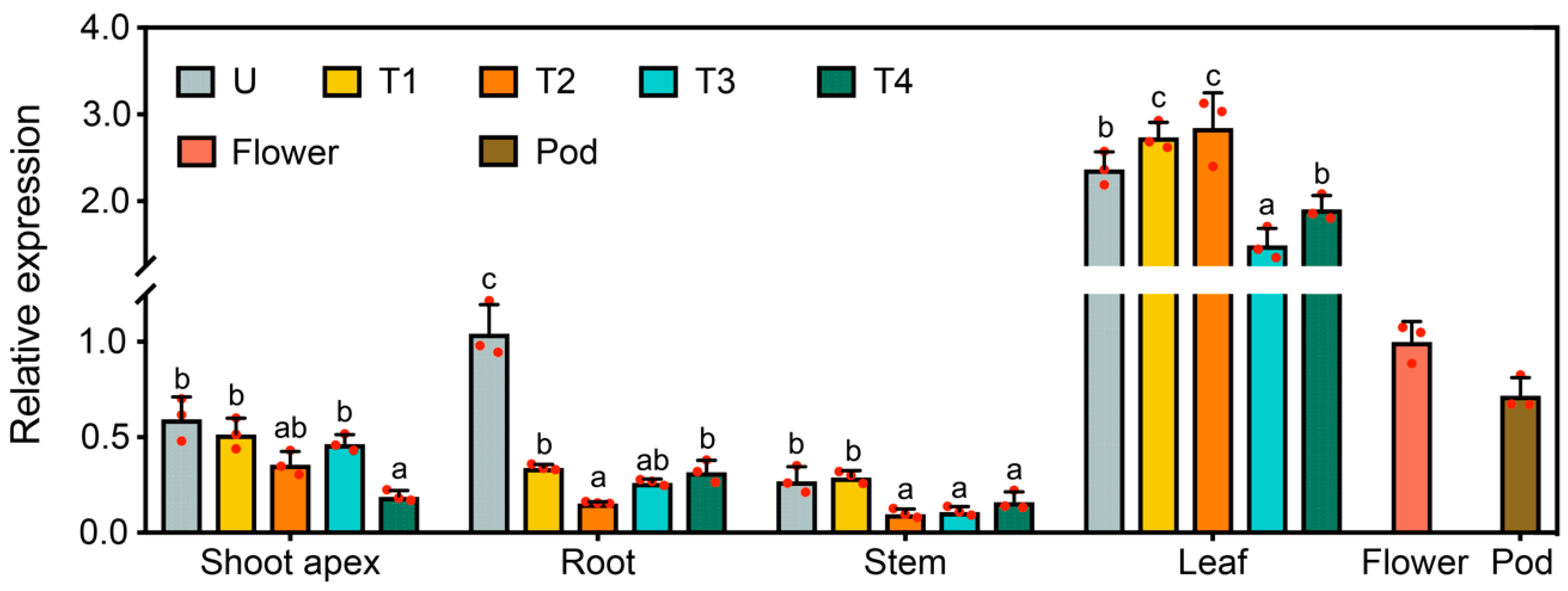
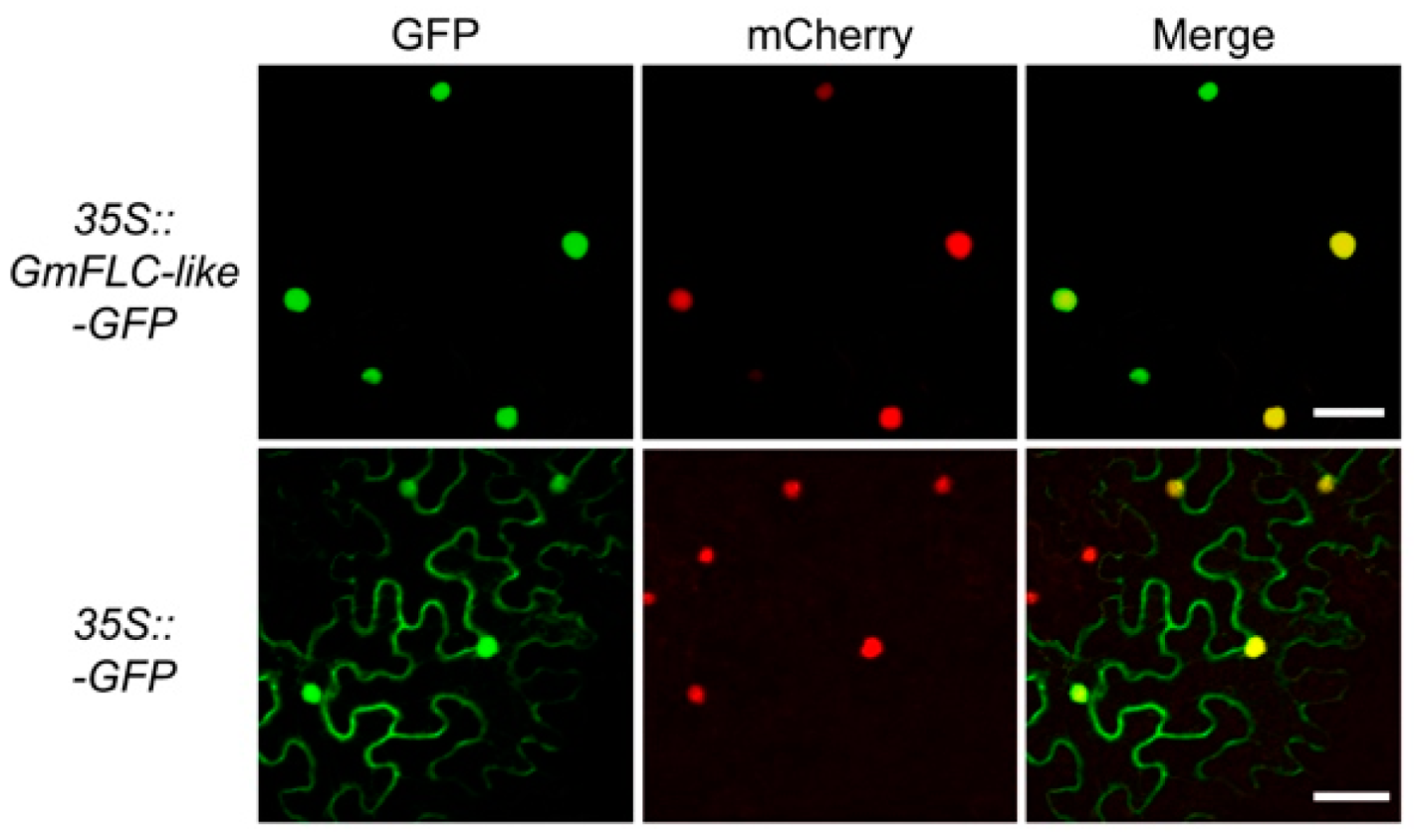
2.2. Expression Profile and Biochemical Properties of GmFLC-like
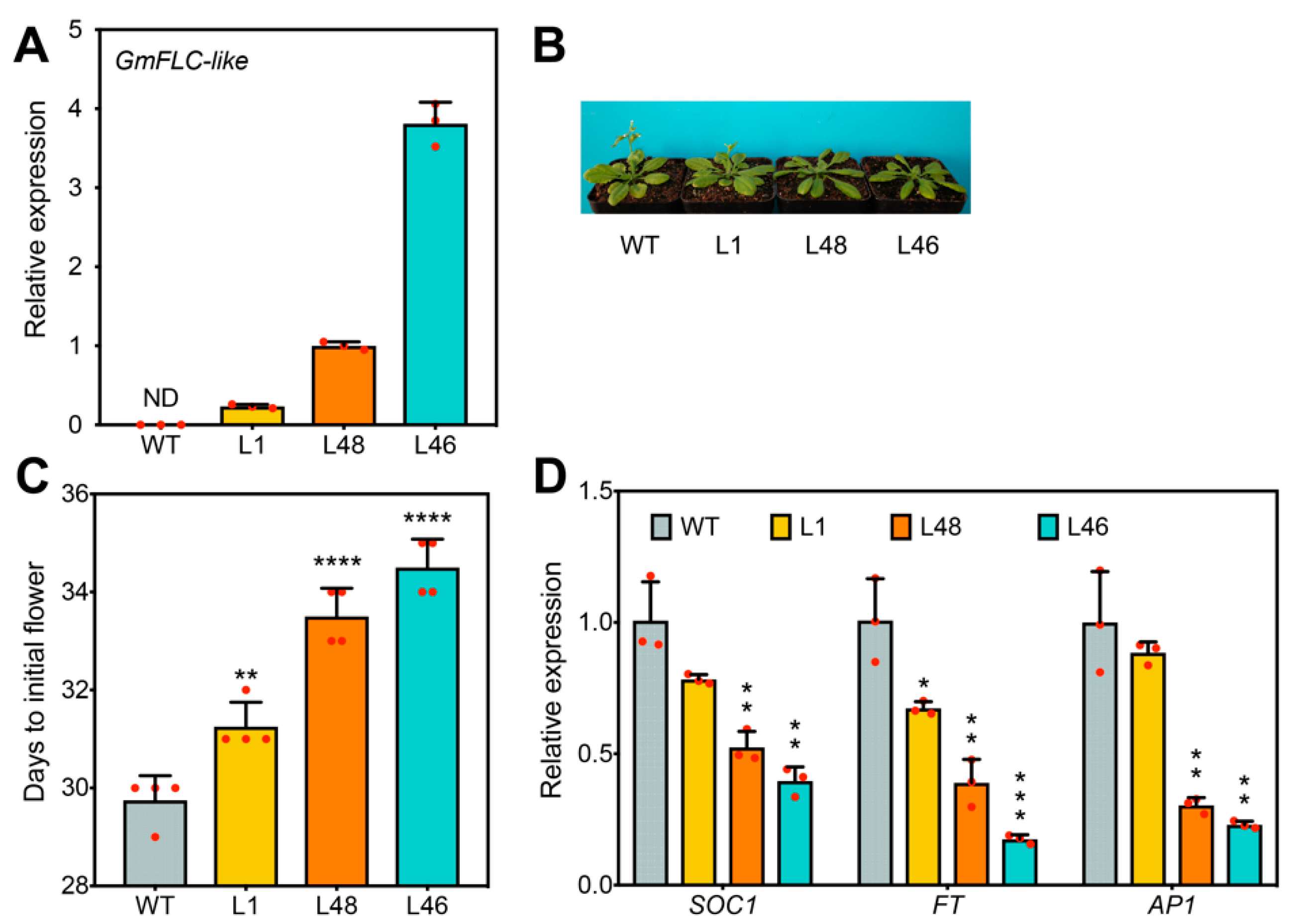
2.3. Overexpression of GmFLC-like Caused Late Flowering in Arabidopsis
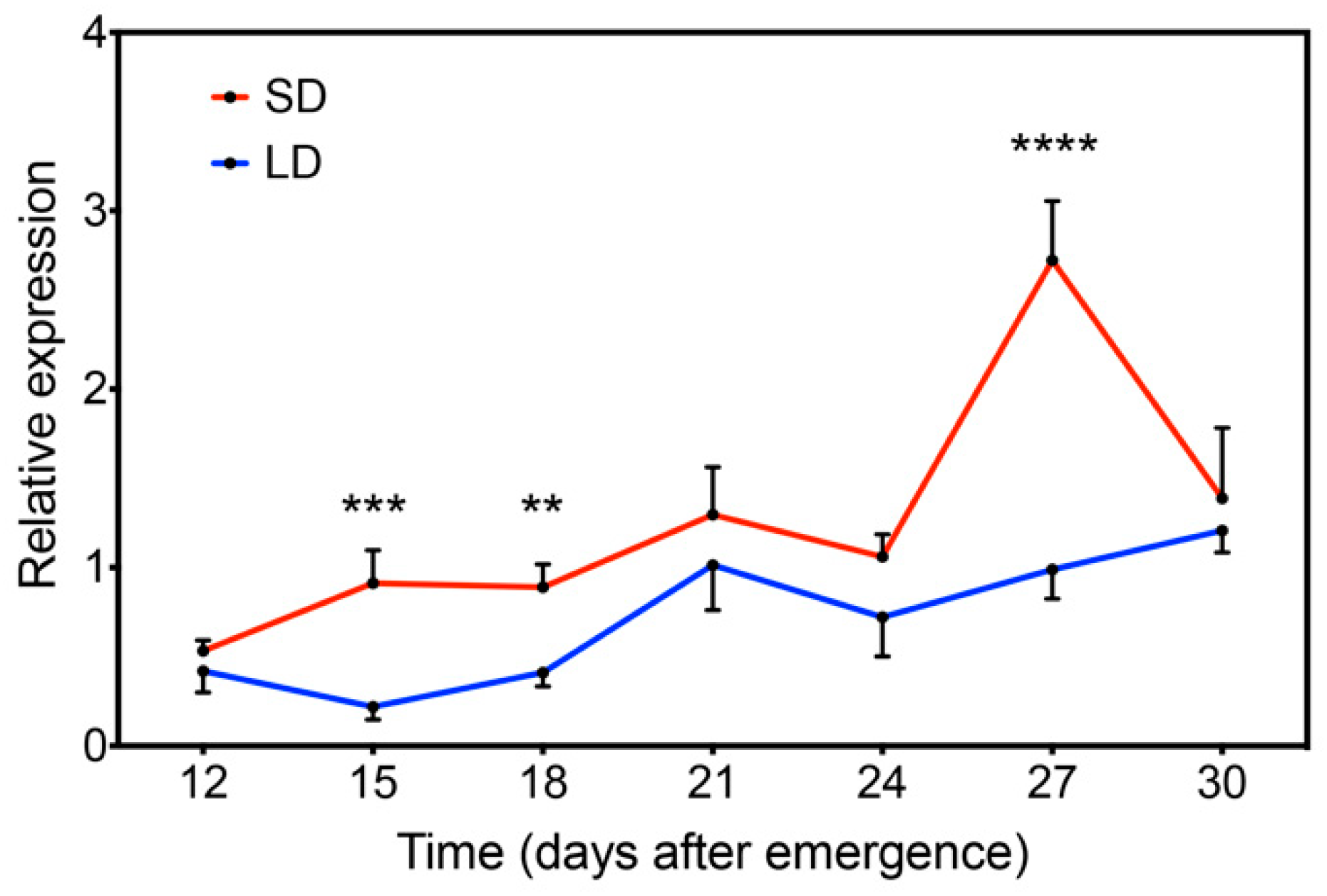
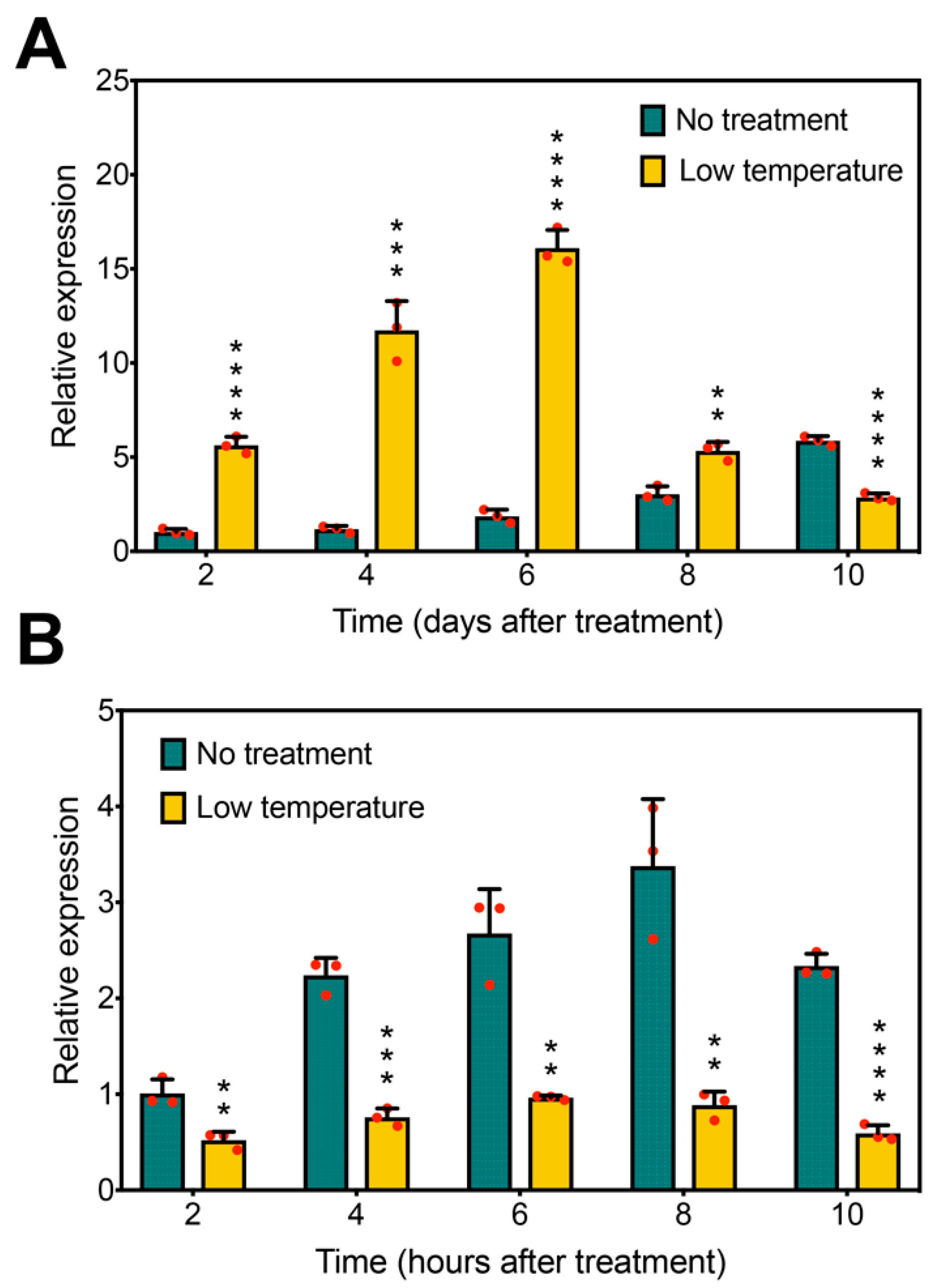
2.4. GmFLC-like Is Responsive to the Photoperiod and Low Temperature
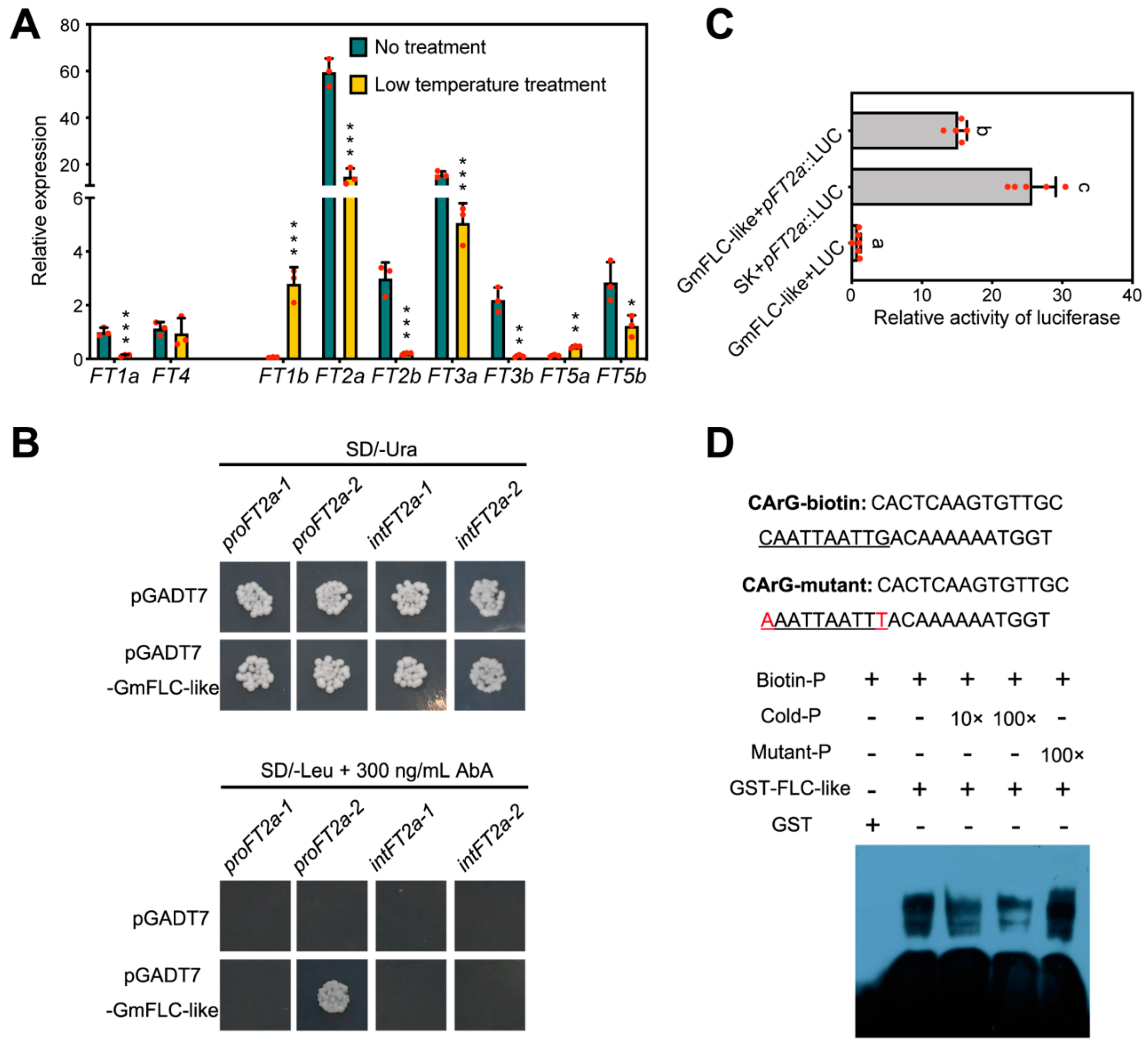
2.5. Identification of GmFT2a as a Downstream Target of GmFLC-like
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Total RNA Isolation and qRT-PCR Analysis
4.3. Plasmid Construction
4.4. Sequence Analysis
4.5. Subcellular Localization of GmFLC-like Protein
4.6. Ectopic Expression of GmFLC-like in Arabidopsis
4.7. Photoperiod Treatment
4.8. Low-Temperature Treatment
4.9. Yeast One-Hybrid Assay
4.10. Dual-Luciferase Reporter Assay
4.11. Electrophoretic Mobility Shift Assay
4.12. Data Analysis
4.13. Data Availability
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| LD | long-day |
| SD | short-day |
| FLC | flowering locus C |
| Y1H | yeast one-hybrid |
| EMSA | electrophoretic mobility shift assay |
| FT | flowering locus T |
| FRI | frigida |
| VIN3 | vernalization insensitive 3 |
| COOLAIR | cold induced long antisense intragenic RNA |
| COLDAIR | cold assisted intronic noncolding RNA |
| SOC1 | suppressor of overexpression of constants 1 |
| VRN1 | vernalization 1 |
| VRN2 | vernalization 2 |
| AP1 | apetala 1 |
| MS | murashige and skoog |
| TUB2 | tubulin beta chain 2 |
| AbA | aureobasidin A |
| IPTG | β-d-1-thiogalactopyranoside |
References
- Srikanth, A.; Schmid, M. Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef]
- Wei, Q.; Ma, C.; Xu, Y.; Wang, T.; Chen, Y.; Lü, J.; Zhang, L.; Jiang, C.-Z.; Hong, B.; Gao, J. Control of chrysanthemum flowering through integration with an aging pathway. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willmann, M.R.; Poethig, R.S. Time to grow up: The temporal role of smallRNAs in plants. Curr. Opin. Plant Biol. 2005, 8, 548–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bäurle, I.; Dean, C. The Timing of Developmental Transitions in Plants. Cell 2006, 125, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, P.; Guo, H.; Chi, W.; Chai, X.; Sun, X.; Xu, X.; Ma, J.; Rochaix, J.-D.; Leister, D.; Wang, H.; et al. Chloroplast retrograde signal regulates flowering. Proc. Natl. Acad. Sci. USA 2016, 113, 10708–10713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izawa, T. Daylength Measurements by Rice Plants in Photoperiodic Short-Day Flowering. In International Review of Cytology; Academic Press: Cambridge, MA, USA, 2007; Volume 256, pp. 191–222. [Google Scholar]
- Cerdán, P.D.; Chory, J. Regulation of flowering time by light quality. Nature 2003, 423, 881. [Google Scholar] [CrossRef]
- Capovilla, G.; Schmid, M.; Posé, D. Control of flowering by ambient temperature. J. Exp. Bot. 2015, 66, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Amasino, R. Vernalization, Competence, and the Epigenetic Memory of Winter. Plant Cell 2004, 16, 2553–2559. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, R.J.; Amasino, R.M. Vernalization: A model for investigating epigenetics and eukaryotic gene regulation in plants. Biochim. Biophys. Acta (BBA)-Gene Struct. Exp. 2007, 1769, 269–275. [Google Scholar] [CrossRef]
- Samach, A.; Wigge, P.A. Ambient temperature perception in plants. Curr. Opin. Plant Biol. 2005, 8, 483–486. [Google Scholar] [CrossRef]
- Michaels, S.D.; Amasino, R.M. FLOWERING LOCUS C Encodes a Novel MADS Domain Protein That Acts as a Repressor of Flowering. Plant Cell 1999, 11, 949–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Suh, S.-S.; Park, E.; Cho, E.; Ahn, J.H.; Kim, S.-G.; Lee, J.S.; Kwon, Y.M.; Lee, I. The AGAMOUS-LIKE 20 MADS domain protein integrates floral inductive pathways in Arabidopsis. Genes Dev. 2000, 14, 2366–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johanson, U.; West, J.; Lister, C.; Michaels, S.; Amasino, R.; Dean, C. Molecular Analysis of FRIGIDA, a Major Determinant of Natural Variation in Arabidopsis Flowering Time. Science 2000, 290, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Le Corre, V.; Roux, F.; Reboud, X. DNA Polymorphism at the FRIGIDA Gene in Arabidopsis thaliana: Extensive Nonsynonymous Variation Is Consistent with Local Selection for Flowering Time. Mol. Biol. Evol. 2002, 19, 1261–1271. [Google Scholar] [CrossRef] [Green Version]
- Gazzani, S.; Gendall, A.R.; Lister, C.; Dean, C. Analysis of the Molecular Basis of Flowering Time Variation in Arabidopsis Accessions. Plant Physiol. 2003, 132, 1107–1114. [Google Scholar] [CrossRef] [Green Version]
- Shindo, C.; Aranzana, M.J.; Lister, C.; Baxter, C.; Nicholls, C.; Nordborg, M.; Dean, C. Role of FRIGIDA and FLOWERING LOCUS C in Determining Variation in Flowering Time of Arabidopsis. Plant Physiol. 2005, 138, 1163–1173. [Google Scholar] [CrossRef] [Green Version]
- Sung, S.; Amasino, R.M. Vernalization in Arabidopsis thaliana is mediated by the PHD finger protein VIN3. Nature 2004, 427, 159. [Google Scholar] [CrossRef]
- Kim, D.-H.; Sung, S. Coordination of the Vernalization Response through a VIN3 and FLC Gene Family Regulatory Network in Arabidopsis. Plant Cell 2013, 25, 454–469. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H.; Doyle, M.R.; Sung, S.; Amasino, R.M. Vernalization: Winter and the Timing of Flowering in Plants. Annu. Rev. Cell Dev. Biol. 2009, 25, 277–299. [Google Scholar] [CrossRef] [Green Version]
- Swiezewski, S.; Liu, F.; Magusin, A.; Dean, C. Cold-induced silencing by long antisense transcripts of an Arabidopsis Polycomb target. Nature 2009, 462, 799–802. [Google Scholar] [CrossRef]
- Heo, J.B.; Sung, S. Vernalization-Mediated Epigenetic Silencing by a Long Intronic Noncoding RNA. Science 2011, 331, 76–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-H.; Xi, Y.; Sung, S. Modular function of long noncoding RNA, COLDAIR, in the vernalization response. PLoS Genet. 2017, 13, e1006939. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Penfield, S. Feedback regulation of COOLAIR expression controls seed dormancy and flowering time. Science 2018, 360, 1014–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepworth, S.R.; Valverde, F.; Ravenscroft, D.; Mouradov, A.; Coupland, G. Antagonistic regulation of flowering-time gene SOC1 by CONSTANS and FLC via separate promoter motifs. EMBO J. 2002, 21, 4327–4337. [Google Scholar] [CrossRef] [Green Version]
- Helliwell, C.A.; Wood, C.C.; Robertson, M.; Peacock, W.J.; Dennis, E.S. The Arabidopsis FLC protein interacts directly in vivo with SOC1 and FT chromatin and is part of a high-molecular-weight protein complex. Plant J. 2006, 46, 183–192. [Google Scholar] [CrossRef]
- Searle, I.; He, Y.; Turck, F.; Vincent, C.; Fornara, F.; Kröber, S.; Amasino, R.A.; Coupland, G. The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis. Genes Dev. 2006, 20, 898–912. [Google Scholar] [CrossRef] [Green Version]
- Lai, Z.; Schluttenhofer, C.M.; Bhide, K.; Shreve, J.; Thimmapuram, J.; Lee, S.Y.; Yun, D.-J.; Mengiste, T. MED18 interaction with distinct transcription factors regulates multiple plant functions. Nat. Commun. 2014, 5, 3064. [Google Scholar] [CrossRef] [Green Version]
- Tadege, M.; Sheldon, C.C.; Helliwell, C.A.; Stoutjesdijk, P.; Dennis, E.S.; Peacock, W.J. Control of flowering time by FLC orthologues in Brassica napus. Plant J. 2001, 28, 545–553. [Google Scholar] [CrossRef]
- Schranz, M.E.; Quijada, P.; Sung, S.-B.; Lukens, L.; Amasino, R.; Osborn, T.C. Characterization and Effects of the Replicated Flowering Time Gene FLC in Brassica rapa. Genetics 2002, 162, 1457–1468. [Google Scholar]
- Reeves, P.A.; He, Y.; Schmitz, R.J.; Amasino, R.M.; Panella, L.W.; Richards, C.M. Evolutionary Conservation of the FLOWERING LOCUS C-Mediated Vernalization Response: Evidence from the Sugar Beet (Beta vulgaris). Genetics 2007, 176, 295–307. [Google Scholar] [CrossRef] [Green Version]
- D’Aloia, M.; Tocquin, P.; Périlleux, C. Vernalization-induced repression of FLOWERING LOCUS C stimulates flowering in Sinapis alba and enhances plant responsiveness to photoperiod. New Phytol. 2008, 178, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-L.; Todesco, M.; Hagmann, J.; Das, S.; Weigel, D. Independent FLC Mutations as Causes of Flowering-Time Variation in Arabidopsis thaliana and Capsella rubella. Genetics 2012, 192, 729–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Périlleux, C.; Pieltain, A.; Jacquemin, G.; Bouché, F.; Detry, N.; D’Aloia, M.; Thiry, L.; Aljochim, P.; Delansnay, M.; Mathieu, A.-S.; et al. A root chicory MADS box sequence and the Arabidopsis flowering repressor FLC share common features that suggest conserved function in vernalization and de-vernalization responses. Plant J. 2013, 75, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Ream, T.S.; Woods, D.P.; Amasino, R.M. The Molecular Basis of Vernalization in Different Plant Groups. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, S.; Sureshkumar, S.; Lempe, J.; Weigel, D. Potent Induction of Arabidopsis thaliana Flowering by Elevated Growth Temperature. PLoS Genet. 2006, 2, e106. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.T.; Lee, C.-R.; Mitchell-Olds, T. LIFE-HISTORY QTLS AND NATURAL SELECTION ON FLOWERING TIME IN BOECHERA STRICTA, A PERENNIAL RELATIVE OF ARABIDOPSIS. Evolution 2011, 65, 771–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, Y.; Higuchi, Y.; Sumitomo, K.; Hisamatsu, T. Flowering retardation by high temperature in chrysanthemums: Involvement of FLOWERING LOCUS T-like 3 gene repression. J. Exp. Bot. 2013, 64, 909–920. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Wu, P.; Wang, Q.; Wang, W.; Zhang, C.; Sun, F.; Liu, Z.; Li, Y.; Hou, X. Comparative transcriptome discovery and elucidation of the mechanism of long noncoding RNAs during vernalization in Brassica rapa. Plant Growth Regul. 2018, 85, 27–39. [Google Scholar] [CrossRef]
- Jiang, B.; Yue, Y.; Gao, Y.; Ma, L.; Sun, S.; Wu, C.; Hou, W.; Lam, H.-M.; Han, T. GmFT2a Polymorphism and Maturity Diversity in Soybeans. PLoS ONE 2013, 8, e77474. [Google Scholar] [CrossRef]
- Nan, H.; Cao, D.; Zhang, D.; Li, Y.; Lu, S.; Tang, L.; Yuan, X.; Liu, B.; Kong, F. GmFT2a and GmFT5a Redundantly and Differentially Regulate Flowering through Interaction with and Upregulation of the bZIP Transcription Factor GmFDL19 in Soybean. PLoS ONE 2014, 9, e97669. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Price, B.W.; Haider, W.; Seufferheld, G.; Nelson, R.; Hanzawa, Y. Functional and Evolutionary Characterization of the CONSTANS Gene Family in Short-Day Photoperiodic Flowering in Soybean. PLoS ONE 2014, 9, e85754. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Lü, S.; Liang, S.; Wu, H.; Zhang, X.; Liu, B.; Kong, F.; Yuan, X.; Li, J.; Xia, Z. GmFT4, a Homolog of FLOWERING LOCUS T, Is Positively Regulated by E1 and Functions as a Flowering Repressor in Soybean. PLoS ONE 2014, 9, e89030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suo, H.; Ma, Q.; Ye, K.; Yang, C.; Tang, Y.; Hao, J.; Zhang, Z.J.; Chen, M.; Feng, Y.; Nian, H. Overexpression of AtDREB1A Causes a Severe Dwarf Phenotype by Decreasing Endogenous Gibberellin Levels in Soybean [Glycine max (L.) Merr.]. PLoS ONE 2012, 7, e45568. [Google Scholar] [CrossRef] [PubMed]
- Suo, H.; Lü, J.; Ma, Q.; Yang, C.; Zhang, X.; Meng, X.; Huang, S.; Nian, H. The AtDREB1A transcription factor up-regulates expression of a vernalization pathway gene, GmVRN1-like, delaying flowering in soybean. Acta Physiol. Plant. 2016, 38, 137. [Google Scholar] [CrossRef] [Green Version]
- Lü, J.; Suo, H.; Yi, R.; Ma, Q.; Nian, H. Glyma11g13220, a homolog of the vernalization pathway gene VERNALIZATION 1 from soybean [Glycine max (L.) Merr.], promotes flowering in Arabidopsis thaliana. BMC Plant Biol. 2015, 15, 232. [Google Scholar]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Buylla, E.R.; Liljegren, S.J.; Pelaz, S.; Gold, S.E.; Burgeff, C.; Ditta, G.S.; Vergara-Silva, F.; Yanofsky, M.F. MADS-box gene evolution beyond flowers: Expression in pollen, endosperm, guard cells, roots and trichomes. Plant J. 2000, 24, 457–466. [Google Scholar] [CrossRef]
- Becker, A.; Winter, K.-U.; Meyer, B.; Saedler, H.; Theißen, G. MADS-Box Gene Diversity in Seed Plants 300 Million Years Ago. Mol. Biol. Evol. 2000, 17, 1425–1434. [Google Scholar] [CrossRef] [Green Version]
- Theißen, G. Shattering developments. Nature 2000, 404, 711. [Google Scholar] [CrossRef]
- Whitelaw, C.A.; Lyssenko, N.N.; Chen, L.; Zhou, D.; Mattoo, A.K.; Tucker, M.L. Delayed abscission and shorter Internodes correlate with a reduction in the ethylene receptor LeETR1 transcript in transgenic tomato. Plant Physiol. 2002, 128, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Bastow, R.; Mylne, J.S.; Lister, C.; Lippman, Z.; Martienssen, R.A.; Dean, C. Vernalization requires epigenetic silencing of FLC by histone methylation. Nature 2004, 427, 164. [Google Scholar] [CrossRef]
- Kong, F.; Liu, B.; Xia, Z.; Sato, S.; Kim, B.M.; Watanabe, S.; Yamada, T.; Tabata, S.; Kanazawa, A.; Harada, K.; et al. Two Coordinately Regulated Homologs of FLOWERING LOCUS T Are Involved in the Control of Photoperiodic Flowering in Soybean. Plant Physiol. 2010, 154, 1220–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Choi, C.W.; Kim, M.C. Identification of diversified functions of soybean FT homologs in photoperiod-dependent flowering time control. In Proceedings of the Korean Society of Crop Science Conference; The Korean Society of Crop Science: Gyeonggi-do, Korea, 2017; p. 100. [Google Scholar]
- Lutz, U.; Posé, D.; Pfeifer, M.; Gundlach, H.; Hagmann, J.; Wang, C.; Weigel, D.; Mayer, K.F.X.; Schmid, M.; Schwechheimer, C. Modulation of Ambient Temperature-Dependent Flowering in Arabidopsis thaliana by Natural Variation of FLOWERING LOCUS M. PLoS Genet. 2015, 11, e1005588. [Google Scholar] [CrossRef] [PubMed]
- Wurr, D.C.E.; Fellows, J.R.; Phelps, K.; Reader, R.J. Vernalization in calabrese (Brassica oleracea var. italica)-a model for apex development. J. Exp. Bot. 1995, 46, 1487–1496. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.P.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Duan, W.; Zhang, H.; Zhang, B.; Wu, X.; Shao, S.; Li, Y.; Hou, X.; Liu, T. Role of vernalization-mediated demethylation in the floral transition of Brassica rapa. Planta 2017, 245, 227–233. [Google Scholar] [CrossRef]
- Blümel, M.; Dally, N.; Jung, C. Flowering time regulation in crops—What did we learn from Arabidopsis? Curr. Opin. Biotechnol. 2015, 32, 121–129. [Google Scholar] [CrossRef]
- Milec, Z.; Valárik, M.; Bartoš, J.; Šafář, J. Can a late bloomer become an early bird? Tools for flowering time adjustment. Biotechnol. Adv. 2014, 32, 200–214. [Google Scholar] [CrossRef]
- Arora, R.; Agarwal, P.; Ray, S.; Singh, A.K.; Singh, V.P.; Tyagi, A.K.; Kapoor, S. MADS-box gene family in rice: Genome-wide identification, organization and expression profiling during reproductive development and stress. BMC Genom. 2007, 8, 242. [Google Scholar] [CrossRef] [Green Version]
- Dennis, E.S.; Peacock, W.J. Vernalization in cereals. J. Biol. 2009, 8, 57. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, C.-H.; Wong, C.E.; Singh, M.B.; Bhalla, P.L. Comparative Genomic Analysis of Soybean Flowering Genes. PLoS ONE 2012, 7, e38250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locascio, A.; Lucchin, M.; Varotto, S. Characterization of a MADS FLOWERING LOCUS C-LIKE (MFL) sequence in Cichorium intybus: A comparative study of CiMFL and AtFLC reveals homologies and divergences in gene function. New Phytol. 2009, 182, 630–643. [Google Scholar] [CrossRef] [PubMed]
- Vogt, S.H.; Weyens, G.; Lefèbvre, M.; Bork, B.; Schechert, A.; Müller, A.E. The FLC-like gene BvFL1 is not a major regulator of vernalization response in biennial beets. Front. Plant Sci. 2014, 5, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheldon, C.C.; Burn, J.E.; Perez, P.P.; Metzger, J.; Edwards, J.A.; Peacock, W.J.; Dennis, E.S. The FLF MADS Box Gene: A Repressor of Flowering in Arabidopsis Regulated by Vernalization and Methylation. Plant Cell 1999, 11, 445–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Ge, X.; Sun, M. Modified CTAB Protocol Using a Silica Matrix for Isolation of Plant Genomic DNA. BioTechniques 2000, 28, 432–434. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]
- HALL, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A Sequence Logo Generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Feng, M.; Chen, W.; Zhou, X.; Lu, J.; Wang, Y.; Li, Y.; Jiang, C.-Z.; Gan, S.-S.; Ma, N.; et al. In rose, transcription factor PTM balances growth and drought survival via PIP2;1 aquaporin. Nat. Plants 2019, 5, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium -mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Qin, R.-Y.; Li, H.; Xu, R.-F.; Yang, Y.-C.; Ni, D.-H.; Ma, H.; Li, L.; Wei, P.-C.; Yang, J.-B. Low-temperature-induced expression of rice ureidoglycolate amidohydrolase is mediated by a C-repeat/dehydration-responsive element that specifically interacts with rice C-repeat-binding factor 3. Front. Plant Sci. 2015, 6, 1011. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Feng, H.; Meng, X.; Li, D.; Yang, D.; Wu, C.; Meng, Q. Overexpression of tomato SlNAC1 transcription factor alters fruit pigmentation and softening. BMC Plant Biol. 2014, 14, 351. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cis-Element | Sequence (5′-3′) | Position (From ATG) | Description |
|---|---|---|---|
| Light regulation elements | |||
| AE-box | AGAAACTT | -1089(-) | Part of a module for light response |
| CATT-motif | GCATTC | -1467(-),-517(-) | Part of a light responsive element |
| G-box | TACGTG | -134(-) | cis-acting regulatory element involved in light responsiveness |
| TCT-motif | TCTTAC | -425(-) | Part of a light responsive element |
| AT1-motif | AATAATTTTTTATT | -915(+) | Part of a light responsive module |
| Box 4 | ATTAAT,TA(C/A)TTA | -1259(+),-498(+),-169(+),-574(+),-285(+) | Part of a conserved DNA module involved in light responsiveness |
| Box I | TTTCAAA | -581(-),-506(-) | Light responsive element |
| G-Box | CACGT(T/A) | -790(+),-117(+) | cis-acting regulatory element involved in light responsiveness |
| as-2-box | GATAATGATT | -657(-) | Involved in shoot-specific expression and light responsiveness |
| rbcS-CMA7a | GGCTATAAGG | -104(+) | Part of a light responsive element |
| chs-CMA1a | TTACTTAA | -575(-) | Part of a light responsive element |
| Hormone and development-related elements | |||
| Circadian | CAANNNNATC | -86(-) | cis-acting regulatory element involved in circadian control |
| Skn-1_motif | GTCAT | -148(+),-1396(+) | cis-acting regulatory element required for endosperm expression |
| ABRE | (C/T)ACGTG | -139(+),-112(-) | cis-acting element involved in the abscisic acid responsiveness |
| GARE-motif | TCTGTTG | -1071(-) | gibberellin-responsive element |
| TCA | GAGAAGAATA,CCATTTTTTT | -1162(-),-751(-) | cis-acting element involved in salicylic acid responsiveness |
| AuxRR-core | GGTCCAT | -748(-) | cis-acting regulatory element involved in auxin responsiveness |
| Abiotic stress response elements | |||
| HSE | AAAAAATTTA | -423(+) | cis-acting element involved in heat shock responsiveness |
| MBS | (C/T)AACTG | -1081(-),-649(+) | MYB binding site involved in drought-inducibility |
| ARE | TGGTTT | -1131(+) | cis-acting regulatory element essential for the anaerobic induction |
| CE3 | AACGCGTGTC | -1330(+) | cis-acting element involved in ABA and VP1 responsiveness |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, J.; Cai, Z.; Li, Y.; Suo, H.; Yi, R.; Zhang, S.; Nian, H. The Floral Repressor GmFLC-like Is Involved in Regulating Flowering Time Mediated by Low Temperature in Soybean. Int. J. Mol. Sci. 2020, 21, 1322. https://doi.org/10.3390/ijms21041322
Lyu J, Cai Z, Li Y, Suo H, Yi R, Zhang S, Nian H. The Floral Repressor GmFLC-like Is Involved in Regulating Flowering Time Mediated by Low Temperature in Soybean. International Journal of Molecular Sciences. 2020; 21(4):1322. https://doi.org/10.3390/ijms21041322
Chicago/Turabian StyleLyu, Jing, Zhandong Cai, Yonghong Li, Haicui Suo, Rong Yi, Shuai Zhang, and Hai Nian. 2020. "The Floral Repressor GmFLC-like Is Involved in Regulating Flowering Time Mediated by Low Temperature in Soybean" International Journal of Molecular Sciences 21, no. 4: 1322. https://doi.org/10.3390/ijms21041322