1. Introduction
Lignocellulosic biomass is an abundant bio-renewable, carbon-neutral resource. It is the basis for alternative energy platforms with the potential to decrease CO
2 emissions and atmospheric pollution by reducing the usage of fossil fuels [
1]. Due to the abundance and availability of lignocellulosic biomass, lignocellulosic-derived cellulosic ethanol represents a significant and inexpensive petroleum fuel alternative [
2]. Lignocellulosic derived glucose is a potential feed stock for a wide variety of biologically produced chemicals and bio fuels [
3]. Cellulase has been widely used in various industrial applications such as the food, animal feed, beer and wine, textile, laundry industry pulp, paper, agriculture, bio-fuel, and pharmaceutical industries amongst others [
4].
Lignocellulosic biomass comprises of three major components: cellulose, hemicellulose, and lignin. Cellulose can be hydrolyzed by a group of enzymes referred to as cellulases, which themselves are classified within carbohydrate-active enzymes (CAZymes), including exoglucanase, endoglucanase, and β glucosidase [
5]. Endoglucanases, or Endo-1, 4-β-
d-glucanase (EC 3.2.1.4), act on amorphous sites. cleave randomly at internal glycoside bonds to generate both reducing and non-reducing ends of cellulose. Exoglucanase, or 1,4-β-
d-cellobiohydrolase (EC 3.2.1.91), hydrolyzes the reducing or non-reducing ends of cellulose, liberating either cellobiose or glucose as the major products. β-glucosidase (EC 3.2.1.21) is a key enzyme for complete hydrolyzing of cellobiose to liberate free glucose molecules [
5].
Increases in cellulase production have been achieved through a number of different approaches such as classical strain improvement (CSI), metabolic engineering, and synthetic biology [
6]. While CSI is robust, it is also time consuming, laborious, and depends on either mutagenesis followed by phenotypic screening for improved characteristics, or manipulation of desired genes known to play a significant role in the desired phenotype [
7].
Genome shuffling is an efficient tool to generate improved strains, whereby the genome of multiple parent strains is shuffled through recursive recombination. This can be achieved by multi-parental crossing though DNA shuffling with a homologous recombination of entire genomes, analogous to conventional breeding [
8]. Through this process, microbial genome shuffling efficiently generates combinatorial libraries of new strains with diverse genetic potential. Hence, this approach is able to produce strains with improved characteristics and efficiencies in respect to desirable phenotypes. For example, genome shuffling improved the production of Tylosin by
Streptomycin fradie, which has been estimated to reduce an approximately twenty year and a one million strain screen classical approach to just a year and 24,000 screens [
9].
Over the past decade, applications of genome shuffling have extended to various sectors, such as the health care, pharmaceutical, food, agriculture, textile and chemical industries. It has been used to enhance microbial stress and acid tolerance, as well as antibiotic production. The greatest advantage of genome shuffling is that many genes and their regulatory units can be randomly changed throughout the entire genome without prior knowledge of genome sequence information [
10]. Initially this method was developed for bacteria and was later extended to yeast and fungi. For example, reports using genome shuffling include increased antifungal activity in
Lactobacillus plantarum [
11], increased acetic acid tolerance of ethanologenic yeast
Candida krusei GL560 [
12], and increased production of cellulase production in
Trichoderma viride [
13].
Although fungi are the common sources of cellulases, cellulases from other microorganisms are being explored [
14,
15]. This is important because of the varied applications of cellulases in diverse backgrounds (paper, textiles, food, etc.). The diversity of microorganisms can cater to the varied conditions prevailing in different industrial applications. Further, the efforts to reduce cost and improve efficiency require better enzymes. Members of
Bacillus sp. are known to produce cellulases [
14,
16,
17]. They may be advantageous because of their fast growth. Further, the genomes are simple and convenient to manipulate.
B. subtilis is known to possess numerous CAZymes (
www.cazy.org;
File S3), with the
B. subtilis subspecies spizizenii str.W23,
B. subtilis B. subsp. natto BEST195, and
B. subtilis subsp subtilis str.168 having between 125 and 145 [
18,
19].
B. subtilis has also been utilized as a source for endogluconases, and thermostable β-glucosidases, with potential for improving bioethanol production [
20,
21]. Any mutations in the structural genes for cellulase activity or other CAZymes may influence the cellulase activity.
In this study, .genome shuffling was chosen to enhance the cellulase production in Bacillus subtilis VS15. Subsequently, the genetic variation between the wild VS15 and the mutant Gd7 strains was identified by using whole genome sequencing. We have sequenced assembly and annotated the whole genomes of both wild and mutant strains (VS15 and Gd7), and performed the comparative genome analysis with reference strain Bacillus subtilis subsp. subtilis str. 168 as a model cellulase producer.
The present study is a first attempt to improve the cellulase production in Bacillus subtilis strains through genome shuffling and comparative analysis between improved and wild strain by using whole genome resequencing.
2. Results
2.1. Screening for Cellulase Overproducing Mutant Strains
In order to produce mutants, mutagens EMS and NTG with UV were utilized. Various doses of the mutagens EMS (0.620–1.48 mM), NTG (2–5 µg) grains and UV (1–10 min) were tested to find out the optimum dose. Optimal rates were observed at 0.931 mM of EMS, 2 µg grains of NTG, and exposure to UV for 3 min, respectively.
After mutagenesis by the combination of EMS and UV, a total of 400 mutants were evaluated for increased cellulase production. Three mutants with increased cellulase production: EUA9, EUB8, and EUD6, were selected for further study. Similarly, after mutagenesis by NTG and UV mutagens, 298 colonies were evaluated, and NUA7, NUB2, and NUD8 were selected for recursive protoplast fusion. All six mutants had a slight increase in cellulase production in the range of 3.77–5.9 IU/mL compared to the wild type (3.77 IU/mL) referred to in
Table 1.
2.2. Genome Shuffling by Recursive Protoplast Fusion
Protoplast preparation efficiency was enhanced to 70% by the addition of lysozyme in the protoplast buffer. Inclusions of Mutanolysin in the protoplast buffer further increased the frequency of protoplast formation to 92%.
The six mutants with increased cellulase production (EUA9, EUB8, EUD6, NUA7, NUB2, and NUD8) were employed for the first round of fusion. After the first round of shuffling, ninety colonies were screened for faster growth and bigger zone of hydrolysis. The shuffled strains G1, G21, G24, and G56 had CMCase activity of 7.27 IU/mL, 7.69 IU/mL, 7.91 IU/mL, and 8.06 IU/mL with an enhancement of 93%, 104%, 110%, and 114%, respectively, over the wild type (3.77 IU/mL). The increase in the zone of hydrolysis ranged from 2.2–2.5cm. Four isolates were selected for a second round of genome shuffling. After the second round of genome shuffling, 50 isolates were screened. The colonies Gb2, Gc8, and Gd7 showed CMCase activity of 8.9 IU/mL, 9.5 IU/mL, and 10.08 IU/mL corresponding to a 136%, 151%, and 167% enhancement over the wild type, respectively, with a zone of hydrolysis ranging from 3.15 cm to 3.5 cm (
Figure 1).
The efficiency of improved strains utilization of other carbon sources was analyzed using filter paper, carboxy methyl cellulose, cellobiose, and cotton gin waste. Supernatants from cultures grown for 54 h on an 8% wheat bran medium in a shake flask had 53–75%, 136–167%, 60–96% and 20–40% enhancement in FPase, CMCase, cotton gin waste, and Cellobiose assays compared to the wild type strain. Activities of FPase, CMCase, cotton gin waste, Cellobiose, and extracellular protein concentrations of the shuffled strains were higher than that of the wild types, as shown in
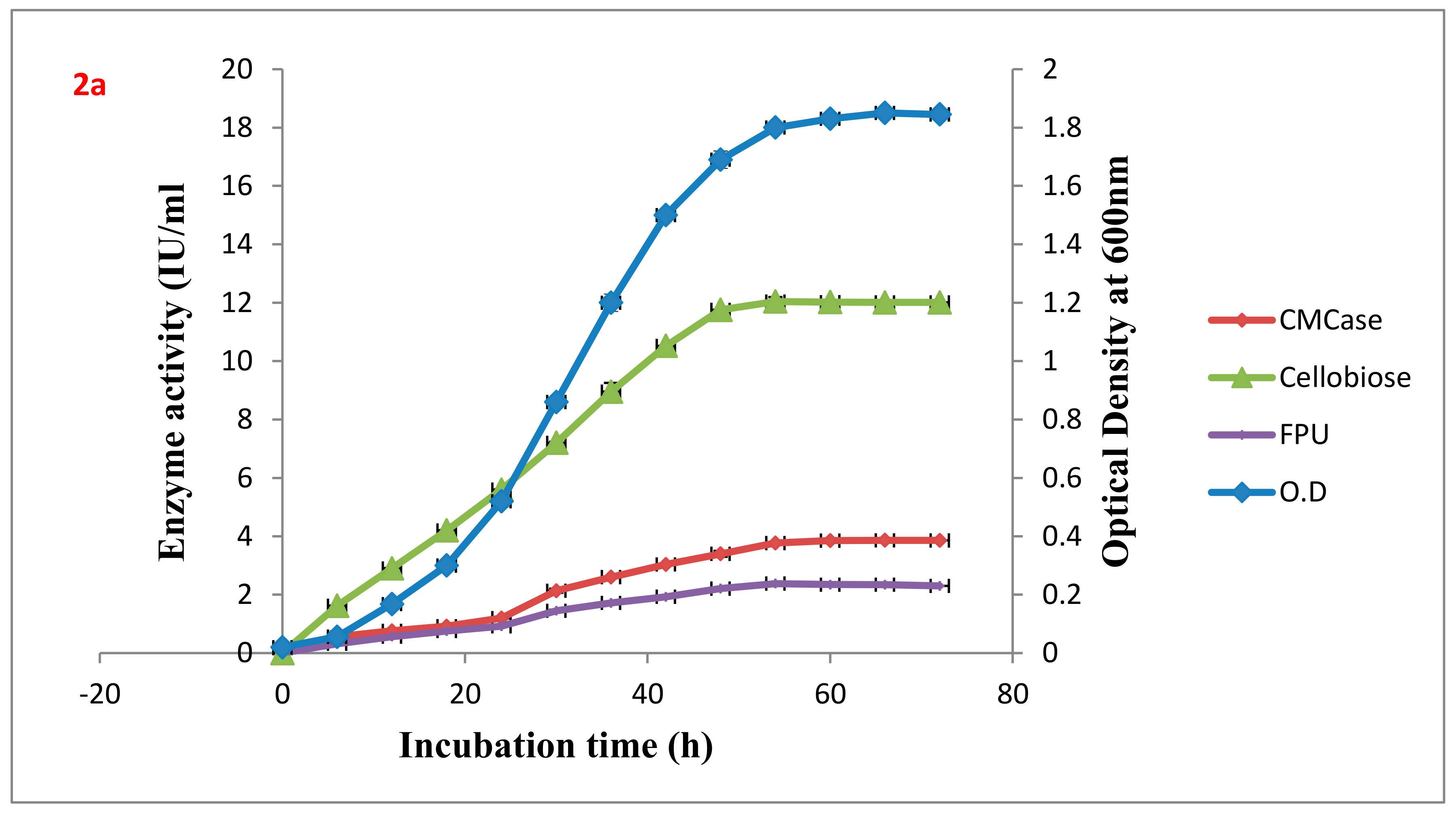
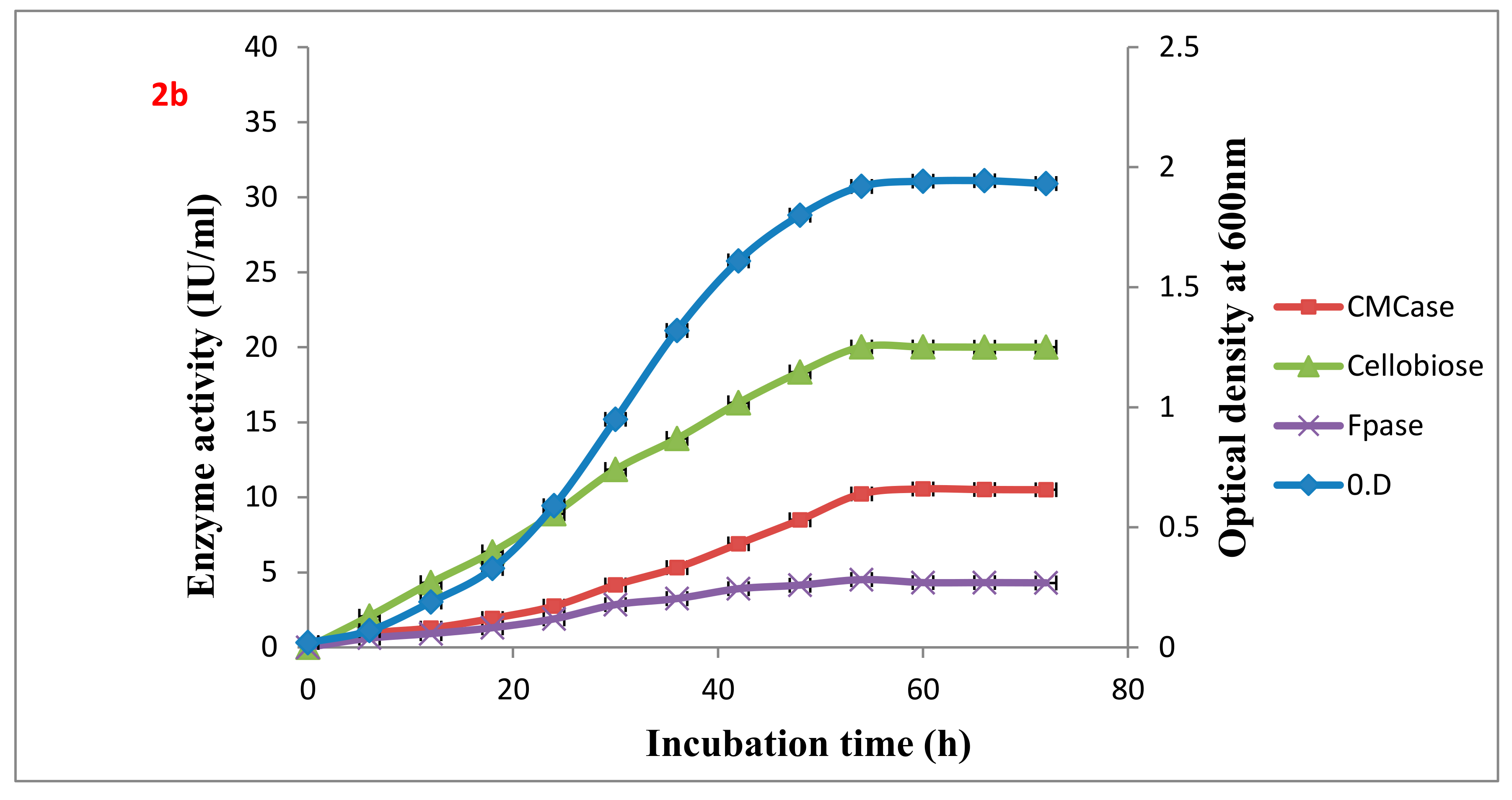
Table 2. The wild type VS15 and the highest producer of all the fusants Gd7 were analyzed for CMCase, FPase, and cellobiase production with respect to the growth curve in an 8% Wheat bran medium at 37 °C (
Figure 2a,b).
2.3. Utilization of Cotton Gin Waste as a Substrate
Acid pre-treatment increased enzymatic hydrolysis and sugar release of the cotton ginning waste. Initially, an increase in acidic concentration resulted in higher sugar releases from 8.93 IU/mL at 1% to 11.08 IU/mL at 3%. A further increase in the acid concentration past 3% caused decreases in the release of sugars to 7.56 IU/mL at 5% and 7.43 IU/mL at 7%. The optimum acid concentration for the pre-treatment was set at 3%. Gb-2, Gc-8 and Gd-7 isolates were used. At 3% concentration, acid showed 9.05 IU/mL, 9.78 IU/mL, and 11.08 IU/mL activity with an enhancement of 60%, 73%, and 96%, respectively, when compared to that of wild types (5.654 IU/mL) on cotton gin waste (
Table 2).
2.4. Hydrolysis Activity of VS 15 and the Shuffled Strains on the CMC Agar Plates
Zone of clearance and colony diameter was measured by using Image J software after 16 h of incubation. Shuffled strains Gb2, Gc8, and Gd7 showed a maximum zone of hydrolysis in the range of 232–244 cm2 around the colony, with a diameter in the range 79–89 cm2, an approximately 1.5 fold increase over the wild type zone of hydrolysis of 121 cm2 and colony diameter of 52 cm2.
2.5. Bioethanol Production
Previous studies reported that through the action of cellulase enzyme, bioethanol production was achieved involving simultaneous mechanism of fermentation and saccharification with saccharomyces cerevisiae [
22]. In this study, the fermentation of VS15 and Gd7 in 8% wheat bran medium and saccharification with 1%
saccharomyces cerevisae, converts the cellulose into ethanol. Further, the ethanol production was confirmed by the K
2Cr
2O
7 method. Through the reaction of K
2Cr
2O
7 with these samples, it was observed that a color change occurred from yellow-orange to green blue. These significant results showed that
saccharomyces cerevisae utilized the liberated sugars from cellulose degradation and show that
Bacillus subtilis VS15 and Gd7 strains are capable of fermenting wheat bran to facilitate the production of biofuel.
2.6. Whole Genome Sequence
Whole Genome sequence data is deposited at DDBJ/ENA/GenBank for the strains VS15 and Gd7 under the accession numbers QFZO00000000 and MSEJ00000000, respectively. The versions described in this paper are QFZO01000000 and MSEJ01000000.
2.7. Assembly and Annotation
No putative misassembles were detected with this genome assembly. The genome size of
Bacillus subtilis VS15 strain is 4,163,202 bp, and the mean GC content is 43.71%, with a final number of contigs of 67 and an N50 value of 293,644 bp (
Table 3). The genome assembly size of Gd7 is 4,146,024 bp, and the mean GC content is 43.65%, with a final number of contigs of 63 and an N50 value of 1,023,553 bp.
All assemblies were annotated using online RAST (Rapid Annotations using Subsystems Technology) Server to evaluate gene set completeness [
23]. A total of 4426 and 4375 genes were predicted for VS15 and Gd7, respectively (
Table 4). Out of 4426 genes, 4316 protein coding gene was reported for VS15, which includes 3317 genes with characterized protein annotation and 1038 hypothetical/putative proteins. Out of 110 predicted non-protein coding genes, 84 are rRNA encoding genes and the remaining 26 codes are for tRNA.
Whereas for Gd7 assembly, out of 4375 predicted genes, 4278 were annotated as protein coding genes and 97 are non-protein coding genes. Altogether, 3287 genes have a characterized protein and the remaining 1017 genes are hypothetical/putative proteins. Out of a total of 97 non-coding genes, 79 genes code for rRNA and 18 code for tRNA.
RAST identified 397 carbohydrate metabolism encoding enzymes. Among these enzymes, 25GHs, 4CEs, 5GTs, 2PLs, and 0AAs were identified as CAZymes using the CAZy database (
File S2).
2.7.1. SNP Calling
Using Bacillus subtilis subsp. subtilis str. 168 as reference strains, a total of 40166 SNPs and 758 indels were reported by VarScan for VS15, and 45797 SNPs and 1104 indels were reported for a Gd7 sample with a minimum coverage of 8, minimum variant frequency of 0.2, minimum average quality of 15, and a P-value threshold ≤ 0.01.
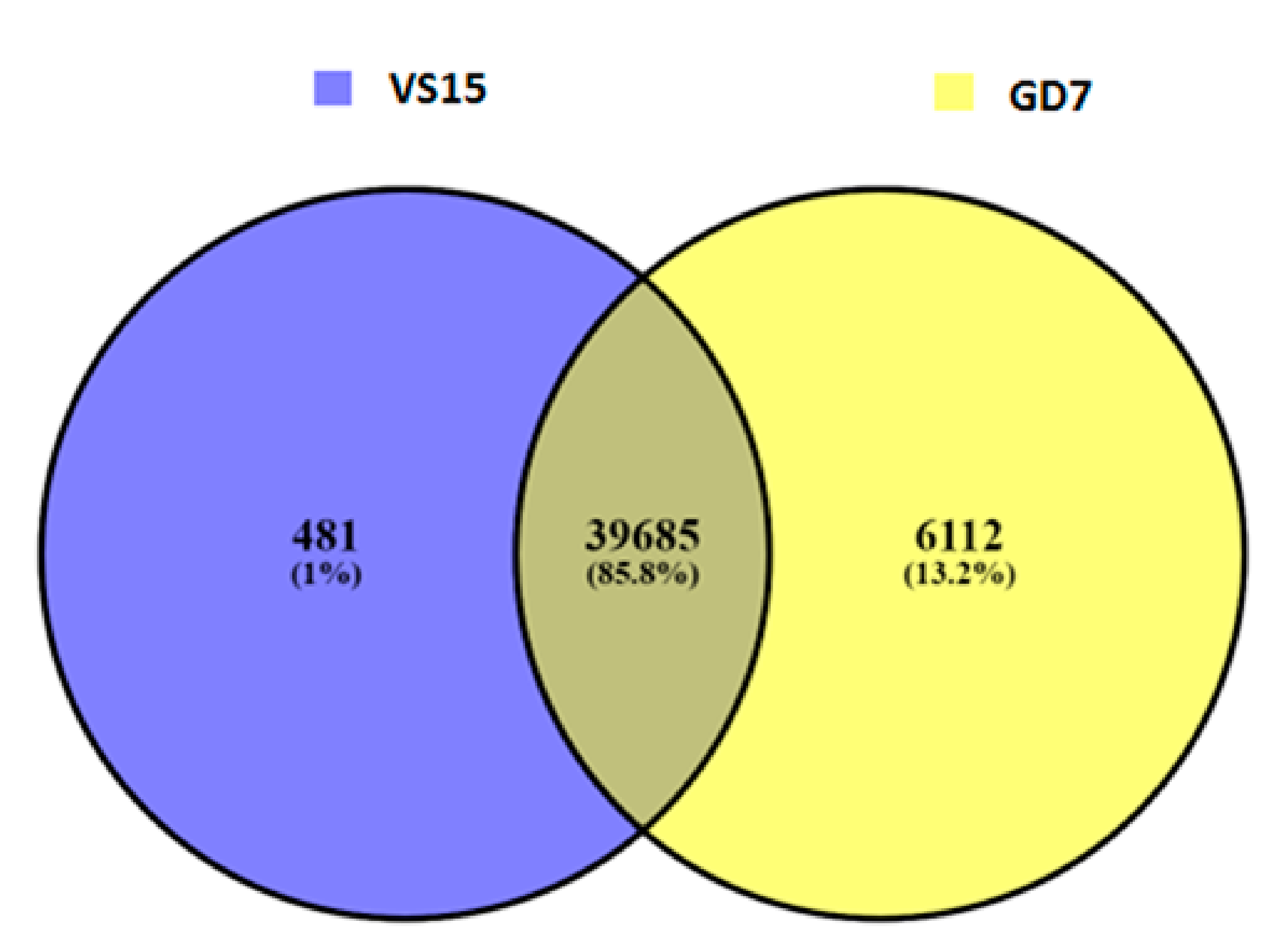
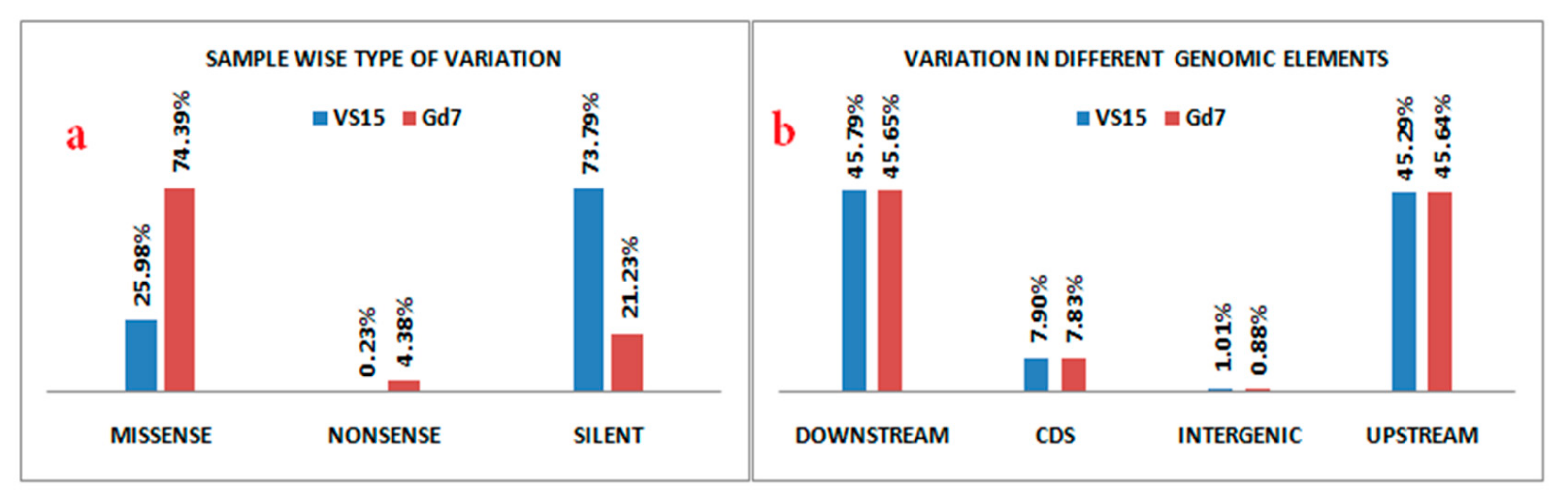
A total of 4375 and 4358 genes were affected by variation i.e., both SNPs and indels in VS15 and Gd7, respectively. Out of total 4375 affected genes, 4194 are protein coding genes, 80 rRNA, 66 pseudogenes, 27 sRNA, 6 lincRNA, 1 antisense, and 1 ribozyme genes. Out of total 4358 affected genes in Gd7, 4176 genes are protein coding, 80 rRNA, 67 pseudogenes, 27 sRNA, 6 lincRNA, 1 antisense, and 1 ribozyme gene. The effect of variation in both the samples is more abundant in RNA encoding genes. 481 unique SNPs were found in VS15, whereas in Gd7, 6112 unique SNPs were found, which were represented in a Venn diagram (
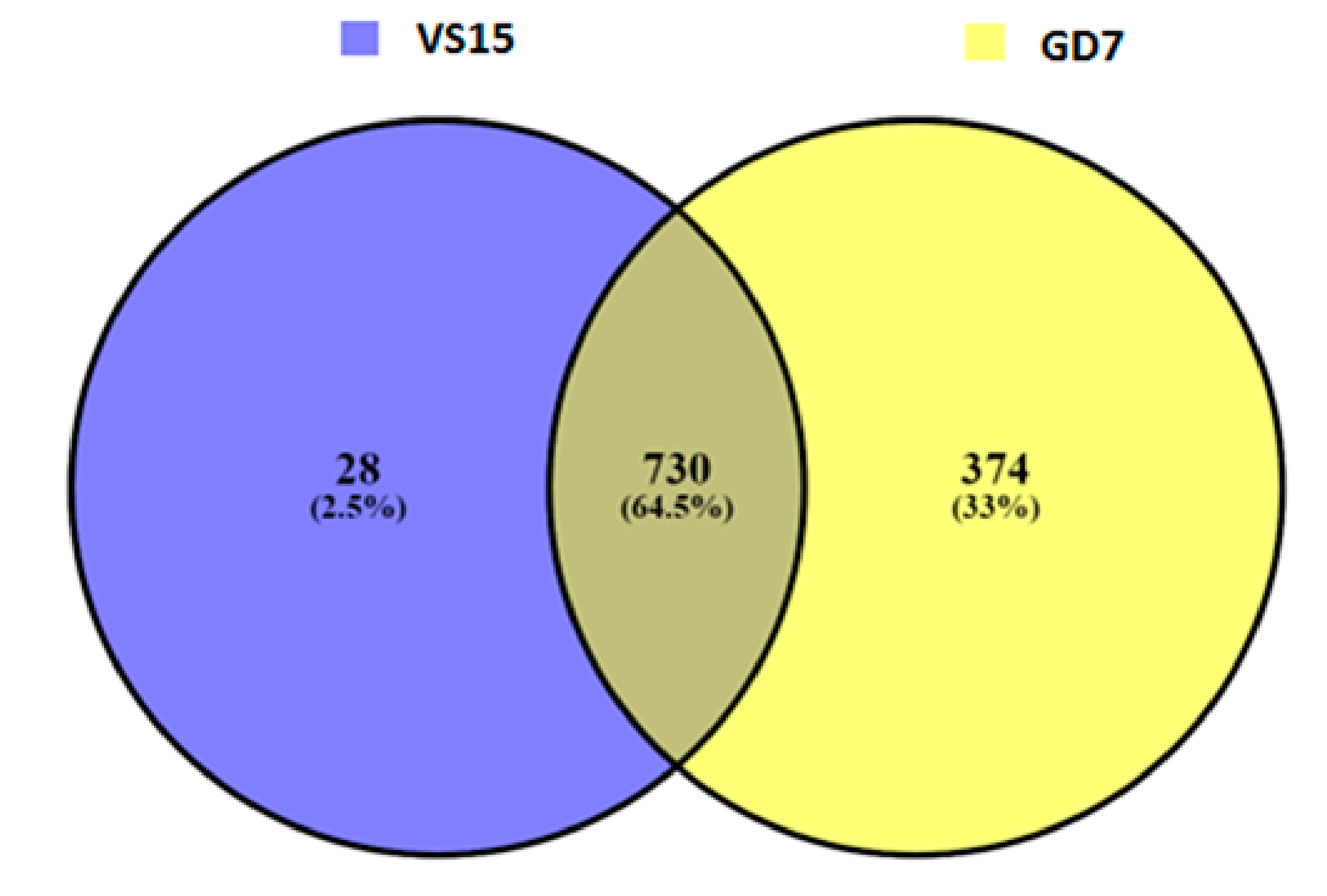
Figure 3). A total of 28 unique indels were predicted for sample VS15 and 374 unique indels were predicted by DELLY in Gd7 (
Figure 4). SNP variation in VS15 and Gd7 were performed on the basis of effect and its distribution in different genomic regions (
Figure 5a,b).
2.7.2. Repeat Analysis
A total of 235 and 133 SSRs were predicted by FullSSR for VS15 and Gd7, respectively. VS15 was predicated to have 85 inverted repeats by using the inverted utility of the EMBOSS package. Out of the 85 inverted repeats, 26 showed inversion with 100% identity, 13 with 94%, and 9 with 95% identity. Gd7 had 25 inverted repeats showing 100% inversion identity, 12 with 94%, and 10 with 95% inversion identity out of a total of 81 inverted repeats. There were 32 and 35 tandem repeats predicted for the VS15 and Gd7 samples, respectively, as well as 1492 and 1483 palindromic sequences with zero mismatches for sample VS15 and sample Gd7. The average size of palindromic sequence for both samples is 11 bp. A total of 9066 total direct repeats were predicted by Red in sample VS15 with a 263 bp average repeat length. In sample Gd7, 8548 direct repeats have been predicted with the average length of 286 bp.
2.8. Specific Gene Set Analysis for Single Nucleotide Polymorphisms (SNPs)
A total of eight genes playing a key role in cellulase production were selected. Four genes (
bglA, bglC and
bglH and
EglS) cause hydrolysis of lignocellulosic biomass. BGL family genes are β-glucosidases, which hydrolyze the non-reducing end β-D-glucosyl residues to release glucose. Among the three BGL family genes (
bglA, bglC, and
bglH), one unique mutation was observed in the bglH gene region at position 4033186 (A substituted with G) compared to its wild type. However, no change in the translated message could be found (
Figure S3). EglS is a hydrolytic enzyme which hydrolyzes the β (1–4)
d-glucosidic linkage. A unique mutation in Gd7 was identified at position 1941335 (changing base A to G). This mutation does not affect the translated messenger RNA or repress the gene function.
Three genes are involved in the regulation of cellulase production. Regulation is mainly caused by carbon catabolite repression (CCR) and LicT anti-termination. The antiterminator LicT specifically binds to the genes involved in cellulase production. LicT positively regulates
bglC gene [
24]. The absolute binding position of
LicT on
bglC genes is located in the region starting from 4035760 to 4035792 with +25 bp upstream and +57 bp downstream, and the binding sequence/cis-element is GGATTGTTACTGCGAAAGCAGGCAAAACCTAAA. Four SNPs are associated with the binding range of LicT on bglC. Out of these four SNPs, three were located within additional +54bp downstream to the absolute binding position, whereas one SNP occurred at 4,035,780 bp, within the actual LicT-bglC binding site (changing base G to A). Although we did not find any variation in the gene expression of the active gene, these mutations may indirectly influence the enhanced cellulase production.
The CCR mechanism in Bacillus subtilis is controlled by CcpA and HprK phosphorylation. Sugar transportation is initiated by entry of glucose via the cytoplasmic membrane (CM) along with the EIICBAGlc component, which provides phosphorylation to the received glucose. The resultant glucose phosphate (Glc-P) activates Hpr that phosphorylates at two sites, serine and histidine. At the serine regulatory site, Hpr is phosphorylated as Hpr-S46-P, with the consumption of ATP. Later, Hpr-S46-P couples with regulatory protein CcpA. The Hpr-S46-P-CcpA complex binds to DNA at the cre (catabolite responsive element) site in the promoter region of the target gene that represses the transcription of the gene encoding hydrolytic enzymes (cellulase, xylanase etc.). In our study, within the CcpA region, nine SNPs were identified in Gd7, while six SNPs were identified in VS15, when the genome of those strains was compared to the reference strain Bacillus subtilis subsp. subtilis str. 168. Three unique mutations were identified in the mutant Gd7 at positions 3044276 (G to A), 3044348 (A to G), and 3044930 (C to T) compared to its wild type.
In Bacillus subtilis, Hpr is a signaling intermediate which is phosphorylated by HprK. Eight SNPs were found in VS15, whereas eleven SNPs were found in Gd7 compared to the reference strain Bacillus subtilis subsp. subtilis str. 168. Among these eleven SNPs, three unique mutations were observed in the mutant (Gd7) at positions 3594559 (C to T), 3595004 (T to C), and 3595028 (C to T) compared to its wild type (VS15). However, no change in the amino acid sequence was observed.
YxaL is a protein kinase enzyme, and repression or deletion of this enzyme results in various defects in glucose metabolism, which tends to force the cell under starvation condition and looking for the alternative source. This condition causes starved cells to switch to the production of hydrolytic enzymes and uptake of cellular storage . YxaL is important for the production of hydrolytic enzymes. Here we found three unique mutations at positions 4102465 (A to T), 4102474 (T to C) and 4102591 (A to C), although these mutations do not alter the translated message.
The detailed SNP distribution among the specific gene set is shown in
Table 5 and
File S1a.
2.9. Specific Gene Set Analysis for InDels
For the selected gene set, InDels for both VS15 and Gd7 samples were analyzed and insertions were found in all of the samples. Three InDels in sample VS15 (
Table 6 and
Table 7) and seven InDels in sample Gd7 (
Table 8 and
Table 9) were found compared to the reference strain
Bacillus subtilis subsp. subtilis str. 168. These InDels have been found upstream within 5 kb of the gene. Among these seven InDels in Gd7,
BglC, and
LicT genes were affected three times, whereas
EglS, gene were affected two times. In wild type (VS15)
BglC,
EglS, and
LicT genes had 1 InDel each. In Gd7, seven InDels were observed, with four being unique InDels when compared to the wild type (VS15). A nucleotide insertion was observed in BglC at a position 369225 (G is inserted at reference base T and the altered base composition was TG) −1.034 kb upstream to the gene.
The licT gene had two insertions, one at a position 4011724 (GAG is inserted in the position T and the altered bases are TGAG) −1.142 kb upstream to the gene, and the other at the position 4011712 with three nucleotide insertions (ACA is inserted in the reference position T and the altered bases are TACA) −1.154 kb upstream to the gene.
A single nucleotide insertion in EglS was observed at position 1938628 (T is inserted in the reference position G and the altered base pair is GT) −1.997 kb upstream to the gene region. All these InDels did not alter the translated messenger RNA.
Besides Indels away from the orfs, Indels could also be located within genes also (
Table 7 and
Table 9). These deletions/insertions lead to frame shift in the reading frame of the ribosome leading resulting in altered translated sequence(as in fatR and GpsA;
File S4) and can lead to altered expression of respective genes or loss of function. These altered expression of these genes possibly influence the cellulase expression.
3. Discussion
Genome shuffling enhanced cellulase production in Bacillus subtilis VS15 through the production of improved mutant strains. Mutagenesis using EMS with UV and NTG with UV produced starting strains for genome shuffling. Shuffled strains were then developed by pooling of desired traits through homologous recombination by protoplast fusion. In order to identify the mutations responsible for this enhanced production of cellulase, the whole genome of two strains (Bacillus subtilis wild type VS15 and mutant type Gd7) were sequenced by NGS and compared to Bacillus subtilis subsp. subtilis str. 168 as a reference genome.
After two rounds of genome shuffling, the CMCase, FPase and Cellobiose activity of Gd7 increased to 167%, 75%, and 40%, respectively, over that of wild type VS15 (
Table 2). Previous studies [
25,
26,
27] reported that the increased growth rate in mutants allows the cells to utilize nutrients more effectively than their respective wild type strains, suggesting that increased extracellular production of enzyme proteins may contribute to enhanced production of cellulase. Earlier genome shuffling studies reported improved FPase activity in
Penicillium decumbens JU-A10 by 117%, 142%, and 118% by UV, EMS, and N+ ion implantation mutagenesis, respectively, after two rounds of shuffling and screening over 2000 shuffled strains [
28]. Cellulase activity in
Trichoderma viride F161 was enhanced by 1.97 fold using UV irradiation, low-energy ion beam implantation, and Atmospheric Pressure Non-equilibrium Discharge plasma (APNEDP) [
13]. Improved FPase and CMCase activity was observed in
Aspergillus glaucus HGZ-2 by 1.95 fold and 1.72 fold, respectively, through UV irradiation after two rounds of shuffling [
29].
Cellulolytic enzymes were induced and repressed depending on the availability of easily utilizable sugars [
14]. Cellulase inducers included sepharose, cellulose, cellobiose, δcellobiose-1-5, lactone, lactose, and other oxidized products of cellulose [
30]. In
Trichoderma Reesei, two CBH genes (
cbh1, cbh2), five endoglucanases genes (
Eg1, Eg2,
Eg3,
Eg4, and
Eg5) [
31,
32] and two b-glucosidase (
bgl1/cel3a, bgl2/cel1a) are reported as cellulase producer genes, and five positive transcriptional activators (XYR1, ACE II, HAP 2/3/5, PacC, and AreA) have been identified to play a major role in gene regulation and expression of cellulase. In addition, three repressors (ACE I, Rce1, and CRE I [
33,
34] have been identified. In
Penicillium oxalicum, a novel transcription factor, ClrC, has been identified, which regulates the multiple stress responses and cellulase expression. The absence of ClrC in a mutant strain reduces the induction of cellulase expression [
35].
Transcriptional activators play an important role in the regulation of the production of cellulase in fungal species. Two transcription factors (clr-1 and clr-2) from
Neurospora crassa which were required for growth have been identified [
36]. Phylogenetic analyses also confirmed that clr-1 and clr-2 are conserved in the genomes of filamentous ascomycete fungi capable of degrading cellulose, but is not required for either growth or hemicellulase activity on xylan. A deletion of the clr-2 homolog (clrB) transcription factor in
Aspergillus nidulans failed to induce cellulase gene expression, and reduced cellulolytic activity on Avicel was observed [
36].
As DBTBS [
24] reported, the LicT will regulate the bgIP gene with positive regulation. In the Gd7, one SNP occurred at the actual binding site of bgl gene and three SNPs were observed between 4035735 bp and 4035849 bp compared to the reference strain. These mutations may influence Bgl expression.
The mutant GD7 was obtained by genome shuffling. In this regard, a number of mutations were observed (
Table 5,
Table 7 and
Table 9,
Figure 3,
Figure 4 and
Figure 5). The phenotype of enhanced cellulase production could be the result of these mutations. It may not be possible to attribute the activity to a single mutation and its effect. In this regard, the possible role of multiple mutations in a group of genes is proposed.
A mutant β-glucosidase (bgl) of
Spodoptera frugiperda, obtained by random mutagenesis, increased the rate of the cellulase production [
37]. In our study, a SNP was observed in bglH and eglS. However, these mutations may not lead to an affective expression of the cellulase gene since the translated messenger RNA was unchanged. Similarly, mutations in the yxaL gene do not seem to influence the cellulase production.
In bacteria, cellulase gene regulation is finely controlled by a regulatory mechanism through two key regulons: CcpA and LicT. Carbon Catabolite Repression (CCR) is referred as a glucose effect. The keys factors involved in the CCR are CcpA, Hpr protein, Hpr kinase/phosphorylase and glycolytic intermediates (G6p and FDP) [
38]. In
Bacillus subtilis, CCR is mediated by catabolite control protein A (CcpA). Bacteria can prefer the glucose as a primary carbon source, and it cannot utilize any secondary substrate by allowing the CcpA to block several catabolite genes and often also to repress the transcription of alternative sugar pathway operons. CcpA is regulated by its phosphorylated cofactors Hpr and Crh, and its phosphorylation is done by HprK. As such, expression of cellulase cannot happen in the presence of glucose in media. In our study, three mutations were found in the CcpA gene of the mutant compared to the wild type, but since these mutations did not affect the amino acid sequence, the function was not affected. Other studies reported that RL-P37 enhances its cellulase and endoglucanase activity by two-fold [
39]. Cre1, which is a CCR protein of
T.ressei, was found to be truncated in the hyper producer of cellulase in
T. reesei Rut-C30 and demonstrated that Cre1 regulates the cellulase expression [
40]. The disruption of
CreA gene enhanced the production of cellulase and hemicellulase in
A.cellulolyticus [
41].
In
Bacillus subtilis, CCR mechanism is regulated by the HprK catalyzed phosphorylation of Hpr (ser-46). Maximum levels of Hpr are phosphorylated on strongly repressing substrate, and minimum levels of Hpr phosphorylation are observed on weakly repressing substrates. Hence, the strength of the CCR is dependent upon the activity of the HprK/P –dependent de (phosphorylation) of Hpr. Consequently, low Hpr kinase activity limits CCR by weakly repressing sources [
42]. In our study, three unique mutations occurred in the mutant without changing the amino acid sequence, implying no loss of function of HprK gene compared to its wild type (VS15).
There are several genes that can lead to inhibition of phospholipid synthesis like gpsA (Cronan et al. 1975 [
43]; Loewy 1990 [
44], MORBIDONI et al. 1995 [
45]). The inhibition of phosphoplipid synthesis can cause leaky surface allowing the enhanced transport of cellulases to the exterior. There could be inhibition of peptidoglycan synthesis without affecting macromolecular synthesis or growth as was noted in
E. coli (Rodionov et al. 1996 [
46]).
Also, there are mutations found in penicillin binding proteins(pbpA, pbpB) with role in cell wall synthesis and elongation( Murray et al. 1997 [
47]). The affect on peptidoglycan synthesis can cause the development of a leaky mutant. Cell wall endo-peptidases like YkfC (Xu et al. 2010 [
48]), cwlO (Hashimotyo et al. 2012 [
49]; Hashimotyo et al. 2018 [
50]) can create hurdles in cell wall synthesis.
There were mutations in spore coat components of cotT, cotB, spoIIQ or yeeK. Unfavourable conditions like starvation may induce sporulation. Lack of ready substrates like glucose may favour sporulation. However, in order to utilise and grow on alternative substrates like cellulose, the host inactivated sporulation genes. Other surface components included membrane protein, bacteriophage SPP1 receptor(yueB). However, their role cannot be related to cellulase activity.
There are mutations to be found in genes related to carbohydrate metabolism like endo-1,4-beta-xylanase(xylanaseD,xynD),beta-galacturonidase(yesZ), alpha;-galacturonidase (lplD), chitosanase(csn). Probably, these genes or pathways compete for the intermediates or metabolites of the pathway involving cellulases. Possibly the metabolites of cellulase activity can act as inducers for these genes or operons, and during selective pressure of the genome shuffling, these genes were silenced.
Genes of regulatory origin have also been identified as mutants. These include rsbX(serine phosphatase), gutR (transcriptional regulator of the glucitol operon), yhcZ(two-component response regulator ,YhcY), fatR(transcriptional regulator for cypB), yobD((transcriptional regulator; phage-related; Xre family)).
Also deletions/insertions were found in other genes hemN(coproporphyrinogen III oxidase), pKSM(polyketide synthase), rseP (inner membrane zinc metalloprotease required for the extracytoplasmic stress response mediated by sigma€, YaeL), rok (repressor of comK), plipastatin synthetase(PpsC), swrAA (motility and swarming), SwrC(surfactin self-resistance) tuaA(teichuronic acid synthesis).
Rok is a repressor of the transcriptional activator ComK and is therefore an important regulator of competence in Bacillus subtilis and is also proved to be a suppressor of membrane bound and secreted proteins. HtpX is an integral cytoplasmic membrane metalloprotease facillitating growth especially at high temperatures is repressed by rok(Lin et al. 2018 [
51]). In the event of mutation in rok, the host is able to grow well at high temperatures. Mutant of rok displayed altered colony morphology (Kovacs et al. 2011 [
52]).
Sigma (E) is required for expression of phospholipids, membrane proteins etc during stress conditions. Inactivation of rseP may cause lack of activity of sigma(E), affecting phospholipid synthesis or membrane synthesis resulting in leaky mutants.
The tuaA gene is involved in the incorporation of teichuronic acid into cell walls of B.subtilis (Qi et al. 1998 [
53]). The mutations in tuaA may cause decrease in level of teichuronic acid in cell wall, possibly leading to a leaky mutant.
The fatR protein represses the expression of the fatR-cyp102A3 operon involved in fatty acid detoxification.Unsaturated fatty acids have the capacity to interact with FatR and prevent repression (Gustafsson et al. 2001 [
54]). There is accumulation of fatty acids during inhibition of phospholipid synthesis (like during inactivation of gpsA). Mutation in fatR prevents its repression activity.This may cause the expression of fatR-cyp102A3 operon facilitatiing fatty acid detoxification or surviving fatty acid accumulation.
The gene YhcZ forms part of two component regulatory system (YfcZ-YfcY) and promotes glucsoe and carbon source utilisation (Jia et al. 2018). Mutations in the yhcZ may lead to conditions of carbon starvation,activation of the carbon catabolite repressor pathways, that cause induction of cellulase activity.
Bacillus subtilis can utilize glucitol as a sole carbon source via the gut operon (Watanabe et al. 2003 [
55]). The gene gutR can act as regulator of gut operon. Mutantion of gutR, could cause the host to utilise the xylitol(CHALUMEAU et al. 1978 [
56]). Probably, it may also help the host in utilisation of other alternative substrates like cellulose.
Sigma(B), stress response sigma factor of
Bacillus subtilis, is induced during environmental stress or at low cellular ATP levels.The gene, rsbX is a negative regulator of of sigma(B) (Smirnova et al. 1998 [
57]; Teh et al. 2015 [
58]). Mutation in rsbX, continues the stress response and possible utilisation of alternative substrates pathways (like cellulases) to replenish ATP levels.
The molecular membrane protein chaperon, prsA is also mutated. This chaperon has been implicated in increased secretion of amylase(Quesada-Ganuza et al. 2019 [
59]), lipases (Ma et al. 2018 [
60]) and several other proteins(Kakeshita et al. 2011 [
61]). However, the role of the frameshift mutation in prsA resulting in increased cellulase activity cannot be explained. Similar is the case with mutation in another chaperon, GroeL.
Mutation in SwrAA involved in motility and swarming and gamma- poly glutamic acid synthesis was observed(Osera et al. 2009 [
62]) . The gene (SwrC) conferring resistance to surfactin produced by the host was also mutated. Also, there were mutations to be found in a polyketide synthesis operon,and the plipastatin production operon. The gene hemN facilitating heme synthesis in anerobic conditions(Hippler et al. 1997 [
63]) was mutated. The gene(pucF; allantoate amidohydrolase) involved in purine degradation as nitrogen source(Schultz et al. 2001 [
64]) was mutated. However, their role in cellulase activity cannot be predicted.


 and
and 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}