Environmental Enrichment: Disentangling the Influence of Novelty, Social, and Physical Activity on Cerebral Amyloid Angiopathy in a Transgenic Mouse Model

 , , and
, , and 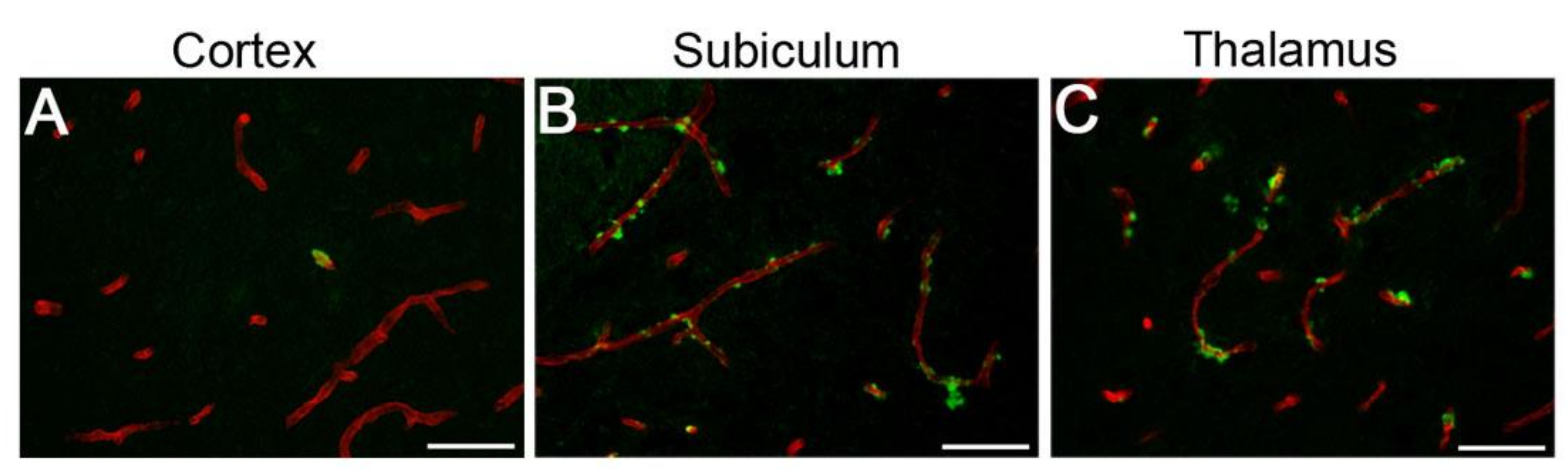{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Physiological Measures
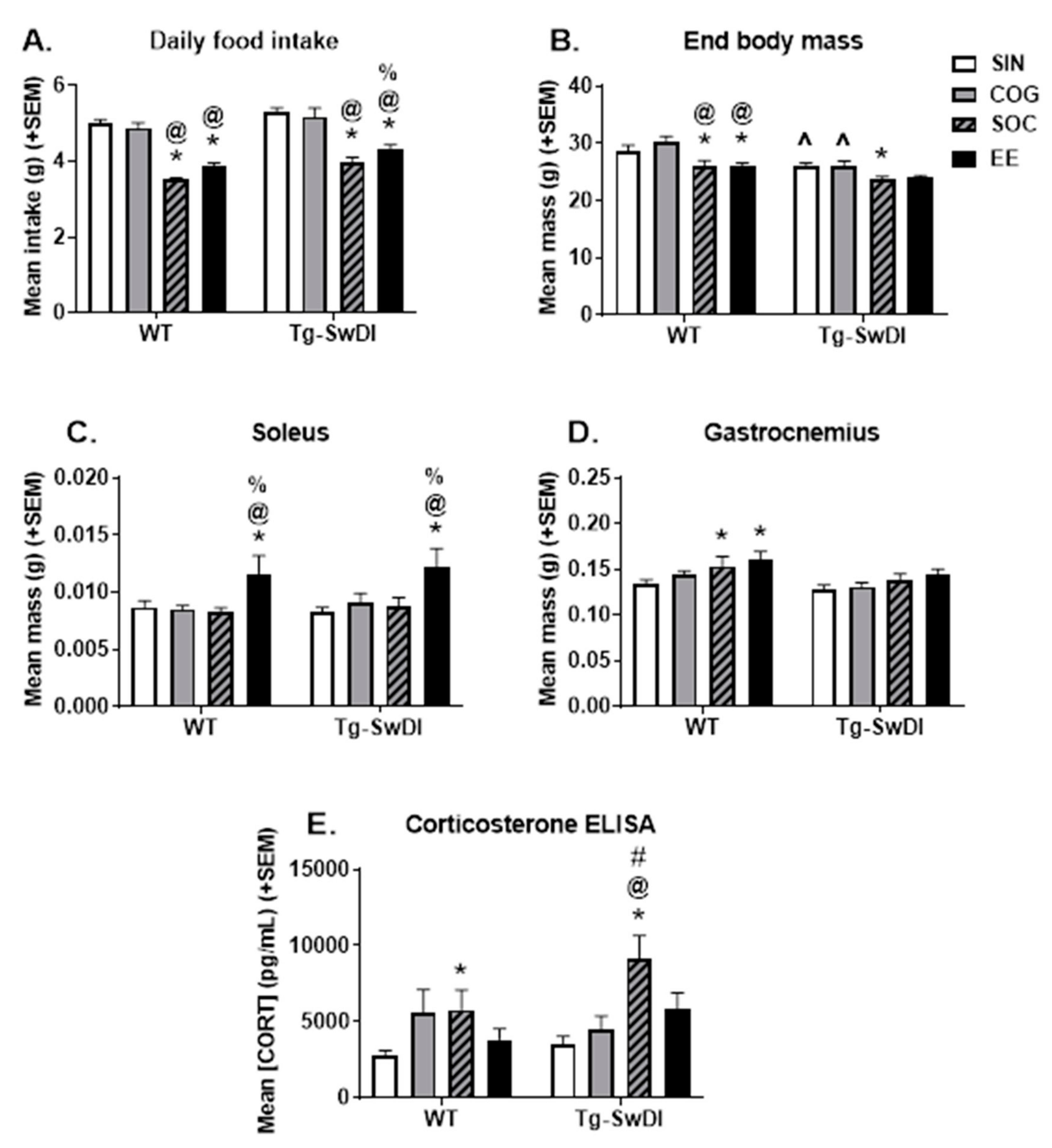
2.1.1. Food Intake and Body Weight
2.1.2. Muscle Mass
2.1.3. Corticosterone ELISA
2.2. Behavioral Tasks of Motor Function and Temperament
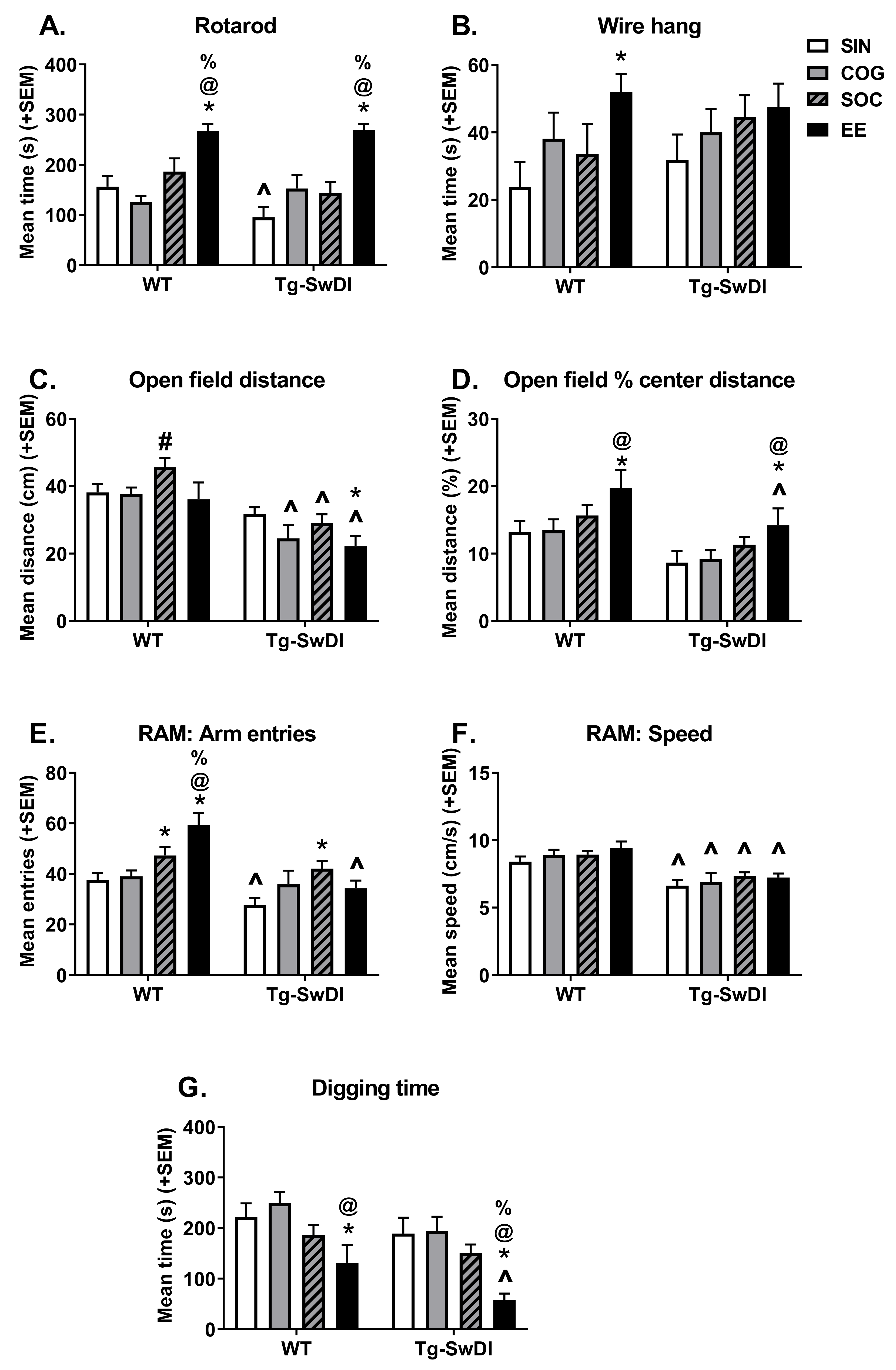
2.2.1. Rotarod
2.2.2. Wire Hang
2.2.3. Open Field
2.2.4. Unreinforced Radial Arm Maze
2.2.5. Digging
2.3. Cognitive Behavioral Performance
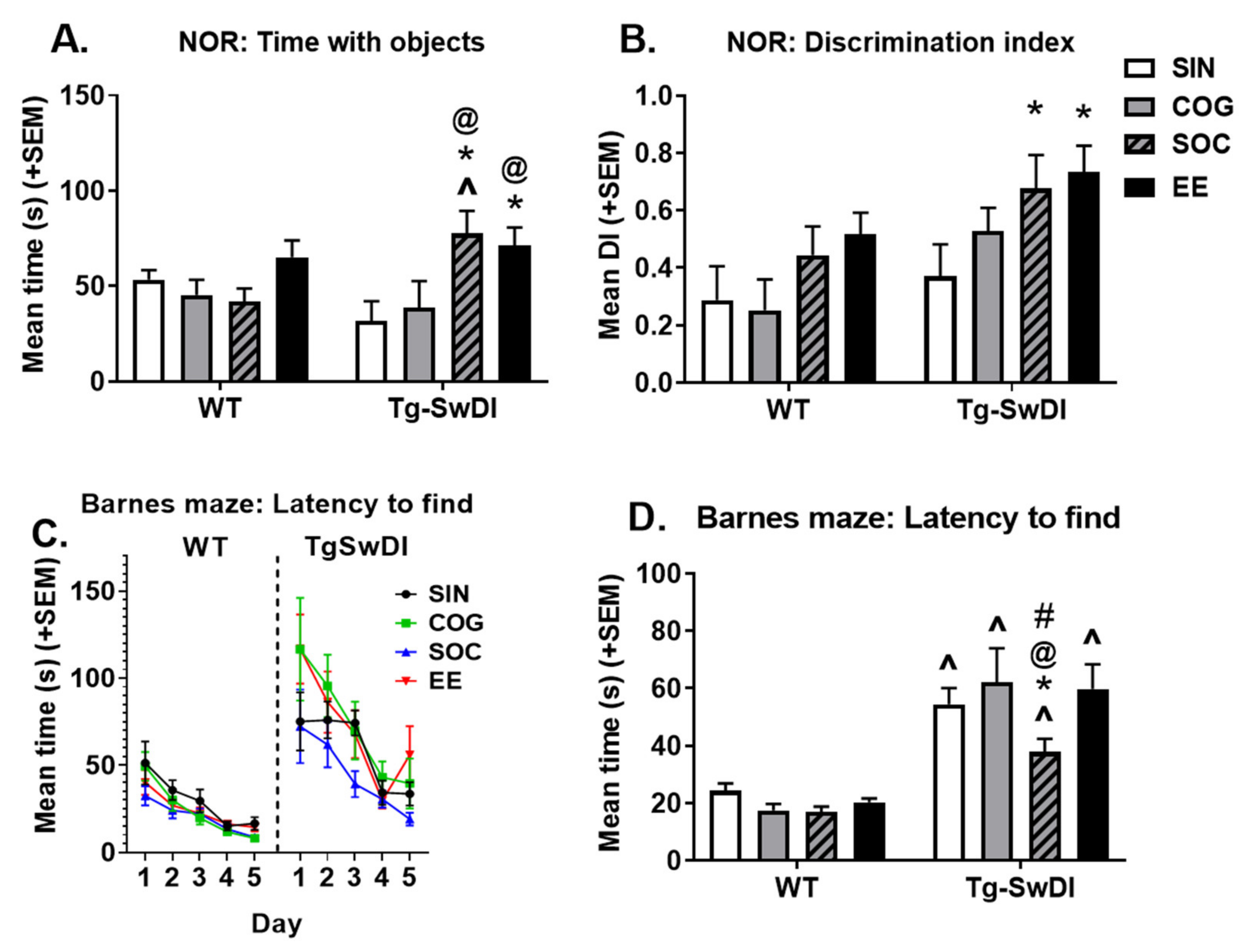
2.3.1. Novel Object Recognition
2.3.2. Barnes Maze
2.4. Pathology
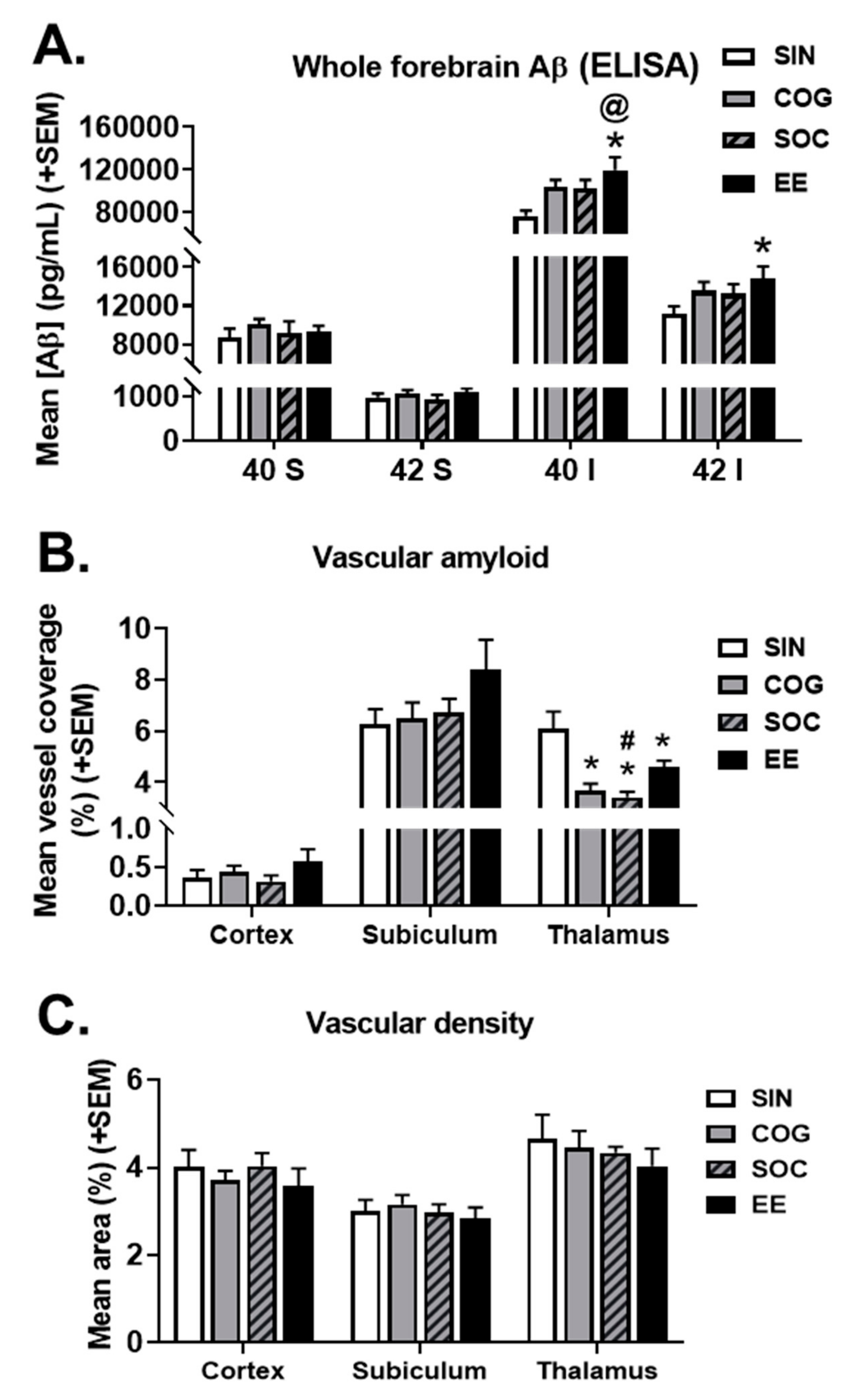
2.4.1. ELISAs for Aβ Species
2.4.2. Microvascular Amyloid Deposition
2.4.3. Vascular Density
3. Discussion
4. Materials and Methods
4.1. Animals
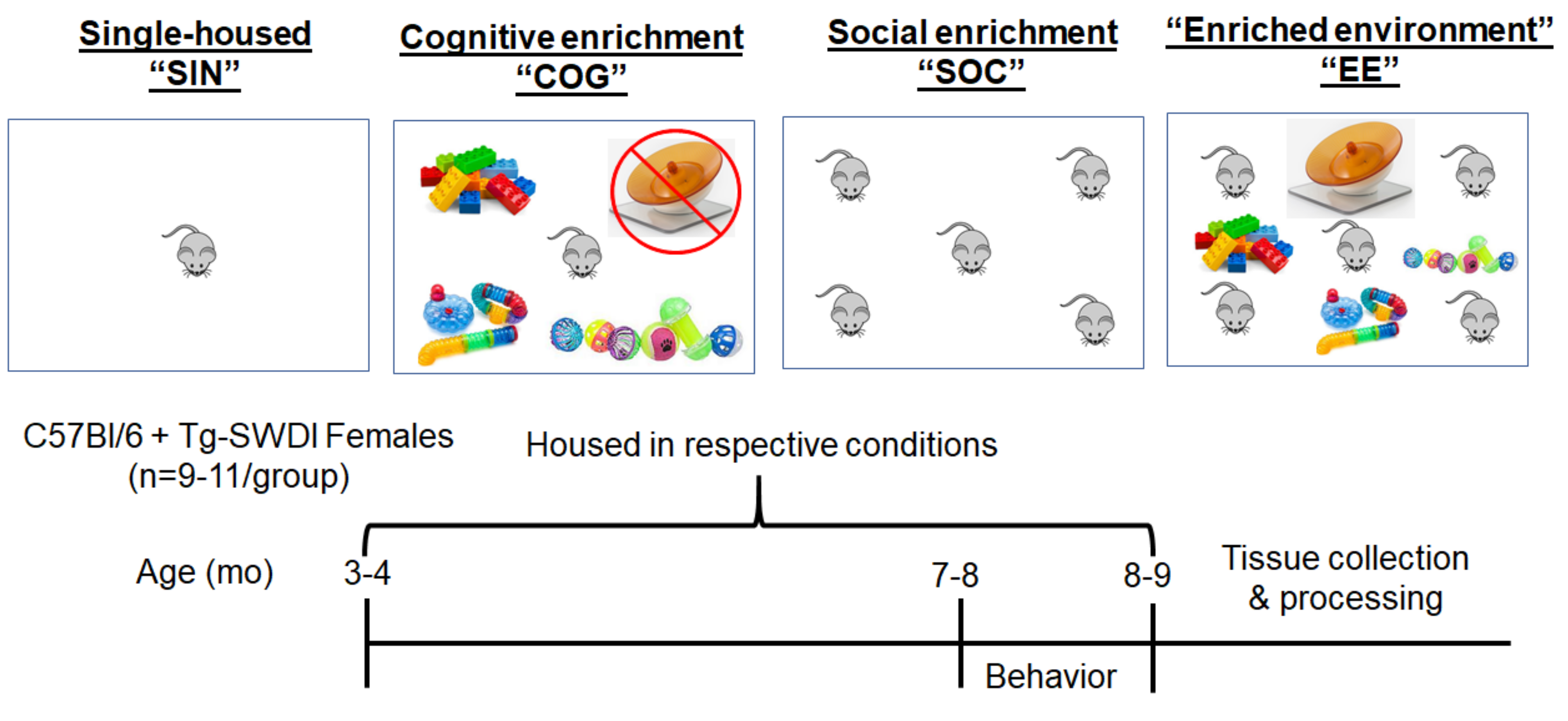
4.2. Housing Conditions
4.3. Cognitive/Behavioral Assessments
4.3.1. Rotarod
4.3.2. Wire Hang
4.3.3. Open Field
4.3.4. Unreinforced Radial Arm Maze
4.3.5. Marble Burying
4.3.6. Novel Object Recognition
4.3.7. Barnes Circular Maze
4.4. Physiological Measures
4.4.1. Blood, Organ, and Muscle Collection
4.4.2. Enzyme Linked Immunosorbent Assay (ELISA) For Serum Corticosterone
4.5. Pathological Aβ Measures
4.5.1. Enzyme-Linked Immunosorbent Assay (ELISA) For Aβ Species
4.5.2. Immunostaining and Analysis
4.6. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Aβ | Beta amyloid |
| AD | Alzheimer’s disease |
| APP | Amyloid precursor protein |
| BDNF | Brain-derived neurotrophic factor |
| CAA | Cerebral amyloid angiopathy |
| COG | Cognitively enriched treatment group |
| DI | Discrimination index |
| EE | Enriched environment |
| ELISA | Enzyme-linked immunosorbent assay |
| NOR | Novel object recognition |
| PBS | Phosphate buffered saline |
| SIN | Single-housed treatment group |
| SOC | Socially (group)-housed treatment group |
| TBS | Tris Buffered Saline |
| WT | Wild-type mice |
References
- Yamada, M. Risk factors for cerebral amyloid angiopathy in the elderly. Ann. New York Acad. Sci. 2002, 977, 37–44. [Google Scholar] [CrossRef]
- Kinnecom, C.; Lev, M.; Wendell, L.; Smith, E.; Rosand, J.; Frosch, M.; Greenberg, S. Course of cerebral amyloid angiopathy–related inflammation. Neurology 2007, 68, 1411–1416. [Google Scholar] [CrossRef] [PubMed]
- Suter, O.-C.; Sunthorn, T.; Kraftsik, R.; Straubel, J.; Darekar, P.; Khalili, K.; Miklossy, J. Cerebral hypoperfusion generates cortical watershed microinfarcts in Alzheimer disease. Stroke 2002, 33, 1986–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, C.M.; Smith, E.E.; Csapo, I.; Gurol, M.E.; Brylka, D.A.; Killiany, R.J.; Blacker, D.; Albert, M.S.; Guttmann, C.R.; Greenberg, S.M. Spatial distribution of white-matter hyperintensities in Alzheimer disease, cerebral amyloid angiopathy, and healthy aging. Stroke 2008, 39, 1127–1133. [Google Scholar] [CrossRef]
- Yoshita, M.; Fletcher, E.; Harvey, D.; Ortega, M.; Martinez, O.; Mungas, D.; Reed, B.; DeCarli, C. Extent and distribution of white matter hyperintensities in normal aging, MCI, and AD. Neurology 2006, 67, 2192–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viswanathan, A.; Patel, P.; Rahman, R.; Nandigam, R.K.; Kinnecom, C.; Bracoud, L.; Rosand, J.; Chabriat, H.; Greenberg, S.M.; Smith, E.E. Tissue microstructural changes are independently associated with cognitive impairment in cerebral amyloid angiopathy. Stroke 2008, 39, 1988–1992. [Google Scholar] [CrossRef] [PubMed]
- Attems, J.; Lauda, F.; Jellinger, K.A. Unexpectedly low prevalence of intracerebral hemorrhages in sporadic cerebral amyloid angiopathy. J. Neurol. 2008, 255, 70–76. [Google Scholar] [CrossRef]
- Attems, J.; Lintner, F.; Jellinger, K.A. Amyloid β peptide 1–42 highly correlates with capillary cerebral amyloid angiopathy and Alzheimer disease pathology. Acta Neuropathol. 2004, 107, 283–291. [Google Scholar] [CrossRef]
- Arvanitakis, Z.; Leurgans, S.E.; Wang, Z.; Wilson, R.S.; Bennett, D.A.; Schneider, J.A. Cerebral amyloid angiopathy pathology and cognitive domains in older persons. Ann. Neurol. 2011, 69, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Xu, F.; Anderson, M.E.; Kotarba, A.; Davis, J.; Robinson, J.K.; Van Nostrand, W.E. Cerebral microvascular rather than parenchymal amyloid-β protein pathology promotes early cognitive impairment in transgenic mice. J. Alzheimers Dis. 2013, 38, 621–632. [Google Scholar] [CrossRef] [Green Version]
- Pope, S.K.; Shue, V.M.; Beck, C. Will a healthy lifestyle help prevent Alzheimer’s disease? Annu. Rev. Public Health 2003, 24, 111–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange-Asschenfeldt, C.; Kojda, G. Alzheimer’s disease, cerebrovascular dysfunction and the benefits of exercise: From vessels to neurons. Exp. Gerontol. 2008, 43, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Cotman, C.W.; Berchtold, N.C. Exercise: A behavioral intervention to enhance brain health and plasticity. Trends Neurosci. 2002, 25, 295–301. [Google Scholar] [CrossRef]
- Radak, Z.; Hart, N.; Sarga, L.; Koltai, E.; Atalay, M.; Ohno, H.; Boldogh, I. Exercise plays a preventive role against Alzheimer’s disease. J. Alzheimers Dis. 2010, 20, 777–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzman, R. Education and the prevalence of dementia and Alzheimer’s disease. Neurology 1993, 43, 13–20. [Google Scholar] [CrossRef]
- Wilson, R.S.; De Leon, C.F.M.; Barnes, L.L.; Schneider, J.A.; Bienias, J.L.; Evans, D.A.; Bennett, D.A. Participation in cognitively stimulating activities and risk of incident Alzheimer disease. JAMA 2002, 287, 742–748. [Google Scholar] [CrossRef]
- Stern, Y.; Gurland, B.; Tatemichi, T.K.; Tang, M.X.; Wilder, D.; Mayeux, R. Influence of education and occupation on the incidence of Alzheimer’s disease. JAMA 1994, 271, 1004–1010. [Google Scholar] [CrossRef]
- Marioni, R.E.; Proust-Lima, C.; Amieva, H.; Brayne, C.; Matthews, F.E.; Dartigues, J.-F.; Jacqmin-Gadda, H. Cognitive lifestyle jointly predicts longitudinal cognitive decline and mortality risk. Eur. J. Epidemiol. 2014, 29, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Olson, A.K.; Eadie, B.D.; Ernst, C.; Christie, B.R. Environmental enrichment and voluntary exercise massively increase neurogenesis in the adult hippocampus via dissociable pathways. Hippocampus 2006, 16, 250–260. [Google Scholar] [CrossRef]
- Kempermann, G.; Gast, D.; Gage, F.H. Neuroplasticity in old age: Sustained fivefold induction of hippocampal neurogenesis by long-term environmental enrichment. Ann. Neurol. 2002, 52, 135–143. [Google Scholar] [CrossRef]
- Birch, A.M.; McGarry, N.B.; Kelly, Á.M. Short-term environmental enrichment, in the absence of exercise, improves memory, and increases NGF concentration, early neuronal survival, and synaptogenesis in the dentate gyrus in a time-dependent manner. Hippocampus 2013, 23, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Ekstrand, J.; Hellsten, J.; Tingström, A. Environmental enrichment, exercise and corticosterone affect endothelial cell proliferation in adult rat hippocampus and prefrontal cortex. Neurosci. Lett. 2008, 442, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Ickes, B.R.; Pham, T.M.; Sanders, L.A.; Albeck, D.S.; Mohammed, A.H.; Granholm, A.-C. Long-term environmental enrichment leads to regional increases in neurotrophin levels in rat brain. Exp. Neurol. 2000, 164, 45–52. [Google Scholar] [CrossRef]
- Rampon, C.; Jiang, C.H.; Dong, H.; Tang, Y.-P.; Lockhart, D.J.; Schultz, P.G.; Tsien, J.Z.; Hu, Y. Effects of environmental enrichment on gene expression in the brain. Proc. Natl. Acad. Sci. USA 2000, 97, 12880–12884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ip, E.Y.-Y.; Giza, C.C.; Griesbach, G.S.; Hovda, D.A. Effects of enriched environment and fluid percussion injury on dendritic arborization within the cerebral cortex of the developing rat. J. Neurotrauma 2002, 19, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.A.; Lewis, M.H.; King, M.A. Environmental enrichment: Effects on stereotyped behavior and dendritic morphology. Dev. Psychobiol. 2003, 43, 20–27. [Google Scholar] [CrossRef]
- Leggio, M.G.; Mandolesi, L.; Federico, F.; Spirito, F.; Ricci, B.; Gelfo, F.; Petrosini, L. Environmental enrichment promotes improved spatial abilities and enhanced dendritic growth in the rat. Behav. Brain Res. 2005, 163, 78–90. [Google Scholar] [CrossRef]
- Segovia, G.; Del Arco, A.; de Blas, M.; Garrido, P.; Mora, F. Effects of an enriched environment on the release of dopamine in the prefrontal cortex produced by stress and on working memory during aging in the awake rat. Behav. Brain Res. 2008, 187, 304–311. [Google Scholar] [CrossRef]
- Benaroya-Milshtein, N.; Hollander, N.; Apter, A.; Kukulansky, T.; Raz, N.; Wilf, A.; Yaniv, I.; Pick, C. Environmental enrichment in mice decreases anxiety, attenuates stress responses and enhances natural killer cell activity. Eur. J. Neurosci. 2004, 20, 1341–1347. [Google Scholar] [CrossRef]
- Roy, V.; Belzung, C.; Delarue, C.; Chapillon, P. Environmental enrichment in BALB/c mice: Effects in classical tests of anxiety and exposure to a predatory odor. Physiol. Behav. 2001, 74, 313–320. [Google Scholar] [CrossRef]
- Llorens-Martin, M.; Rueda, N.; Martínez-Cué, C.; Torres-Alemán, I.; Florez, J.; Trejo, J.L. Both increases in immature dentate neuron number and decreases of immobility time in the forced swim test occurred in parallel after environmental enrichment of mice. Neuroscience 2007, 147, 631–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarov, O.; Robinson, J.; Tang, Y.-P.; Hairston, I.S.; Korade-Mirnics, Z.; Lee, V.M.-Y.; Hersh, L.B.; Sapolsky, R.M.; Mirnics, K.; Sisodia, S.S. Environmental enrichment reduces Aβ levels and amyloid deposition in transgenic mice. Cell 2005, 120, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.-S.; Xu, P.; Pigino, G.; Brady, S.T.; Larson, J.; Lazarov, O. Complex environment experience rescues impaired neurogenesis, enhances synaptic plasticity, and attenuates neuropathology in familial Alzheimer’s disease-linked APPswe/PS1ΔE9 mice. FASEB J. 2010, 24, 1667–1681. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.A.; Cracchiolo, J.R.; Bachstetter, A.D.; Hughes, T.F.; Bales, K.R.; Paul, S.M.; Mervis, R.F.; Arendash, G.W.; Potter, H. Enrichment improves cognition in AD mice by amyloid-related and unrelated mechanisms. Neurobiol. Aging 2007, 28, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Arendash, G.W.; Garcia, M.F.; Costa, D.A.; Cracchiolo, J.R.; Wefes, I.M.; Potter, H. Environmental enrichment improves cognition in aged Alzheimer’s transgenic mice despite stable β-amyloid deposition. Neuroreport 2004, 15, 1751–1754. [Google Scholar] [CrossRef] [PubMed]
- Jankowsky, J.L.; Xu, G.; Fromholt, D.; Gonzales, V.; Borchelt, D.R. Environmental enrichment exacerbates amyloid plaque formation in a transgenic mouse model of Alzheimer disease. J. Neuropathol. Exp. Neurol. 2003, 62, 1220–1227. [Google Scholar] [CrossRef]
- Jankowsky, J.L.; Melnikova, T.; Fadale, D.J.; Xu, G.M.; Slunt, H.H.; Gonzales, V.; Younkin, L.H.; Younkin, S.G.; Borchelt, D.R.; Savonenko, A.V. Environmental Enrichment Mitigates Cognitive Deficits in a Mouse Model of Alzheimer’s Disease. J. Neurosci. 2005, 25, 5217–5224. [Google Scholar] [CrossRef]
- Stern, Y. Cognitive reserve and Alzheimer disease. Alzheimer Dis. Assoc. Disord. 2006, 20, 112–117. [Google Scholar] [CrossRef]
- Wolf, S.A.; Kronenberg, G.; Lehmann, K.; Blankenship, A.; Overall, R.; Staufenbiel, M.; Kempermann, G. Cognitive and Physical Activity Differently Modulate Disease Progression in the Amyloid Precursor Protein (APP)-23 Model of Alzheimer’s Disease. Biol. Psychiatry 2006, 60, 1314–1323. [Google Scholar] [CrossRef]
- Cracchiolo, J.R.; Mori, T.; Nazian, S.J.; Tan, J.; Potter, H.; Arendash, G.W. Enhanced cognitive activity—Over and above social or physical activity—Is required to protect Alzheimer’s mice against cognitive impairment, reduce Aβ deposition, and increase synaptic immunoreactivity. Neurobiol. Learn. Mem. 2007, 88, 277–294. [Google Scholar] [CrossRef] [Green Version]
- Herring, A.; Yasin, H.; Ambrée, O.; Sachser, N.; Paulus, W.; Keyvani, K. Environmental Enrichment Counteracts Alzheimer’s Neurovascular Dysfunction in TgCRND8 Mice. Brain Pathol. 2008, 18, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Ambrée, O.; Leimer, U.; Herring, A.; Görtz, N.; Sachser, N.; Heneka, M.T.; Paulus, W.; Keyvani, K. Reduction of Amyloid Angiopathy and Aβ Plaque Burden after Enriched Housing in TgCRND8 Mice: Involvement of Multiple Pathways. Am. J. Pathol. 2006, 169, 544–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herring, A.; Lewejohann, L.; Panzer, A.-L.; Donath, A.; Kröll, O.; Sachser, N.; Paulus, W.; Keyvani, K. Preventive and therapeutic types of environmental enrichment counteract beta amyloid pathology by different molecular mechanisms. Neurobiol. Dis. 2011, 42, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.; Xu, F.; Ou-Yang, M.H.; Davis, J.; Van Nostrand, W.E.; Robinson, J.K. Intensive ‘Brain Training’ Intervention Fails to Reduce Amyloid Pathologies or Cognitive Deficits in Transgenic Mouse Models of Alzheimer’s Disease. J. Alzheimers Dis. 2017, 55, 1109–1121. [Google Scholar] [CrossRef]
- Ballard, C.; Khan, Z.; Clack, H.; Corbett, A. Nonpharmacological treatment of Alzheimer disease. Can. J. Psychiatry Rev. Can. Psychiatr. 2011, 56, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Redick, T.S.; Shipstead, Z.; Harrison, T.L.; Hicks, K.L.; Fried, D.E.; Hambrick, D.Z.; Kane, M.J.; Engle, R.W. No evidence of intelligence improvement after working memory training: A randomized, placebo-controlled study. J. Exp. Psychol. 2013, 142, 359–379. [Google Scholar] [CrossRef]
- Noack, H.; Lovden, M.; Schmiedek, F. On the validity and generality of transfer effects in cognitive training research. Psychol. Res. 2014, 78, 773–789. [Google Scholar] [CrossRef]
- Shipstead, Z.; Redick, T.S.; Engle, R.W. Is working memory training effective? Psychol. Bull. 2012, 138, 628–654. [Google Scholar] [CrossRef] [Green Version]
- Robison, L.S.; Popescu, D.L.; Anderson, M.E.; Francis, N.; Hatfield, J.; Sullivan, J.K.; Xu, F.; Beigelman, S.I.; Anderson, B.J.; Van Nostrand, W.E.; et al. Long-term voluntary wheel running does not alter vascular amyloid burden but reduces neuroinflammation in the Tg-SwDI mouse model of cerebral amyloid angiopathy. J. Neuroinflamm. 2019, 16, 144. [Google Scholar] [CrossRef] [Green Version]
- Robison, L.S.; Popescu, D.L.; Anderson, M.E.; Beigelman, S.I.; Fitzgerald, S.M.; Kuzmina, A.E.; Lituma, D.A.; Subzwari, S.; Michaelos, M.; Anderson, B.J.; et al. The effects of volume versus intensity of long-term voluntary exercise on physiology and behavior in C57/Bl6 mice. Physiol. Behav. 2018, 194, 218–232. [Google Scholar] [CrossRef]
- Witte, A.V.; Fobker, M.; Gellner, R.; Knecht, S.; Flöel, A. Caloric restriction improves memory in elderly humans. Proc. Natl. Acad. Sci. USA 2009, 106, 1255–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luchsinger, J.A.; Tang, M.-X.; Shea, S.; Mayeux, R. Caloric Intake and the Risk of Alzheimer Disease. Arch. Neurol. 2002, 59, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.; Cole, G.; Head, E.; Ingram, D. Nutrition, Brain Aging, and Neurodegeneration. J. Neurosci. 2009, 29, 12795–12801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Cauwenberghe, C.; Vandendriessche, C.; Libert, C.; Vandenbroucke, R.E. Caloric restriction: Beneficial effects on brain aging and Alzheimer’s disease. Mamm. Genome 2016, 27, 300–319. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.; Mitchell, J.; Kalant, N. The effects of life-long food restriction on spatial memory in young and aged Fischer 344 rats measured in the eight-arm radial and the Morris water mazes. Neurobiol. Aging 1989, 10, 669–675. [Google Scholar] [CrossRef]
- Means, L.W.; Higgins, J.L.; Fernandez, T.J. Mid-life onset of dietary restriction extends life and prolongs cognitive functioning. Physiol. Behav. 1993, 54, 503–508. [Google Scholar] [CrossRef]
- Qin, W.; Yang, T.; Ho, L.; Zhao, Z.; Wang, J.; Chen, L.; Zhao, W.; Thiyagarajan, M.; MacGrogan, D.; Rodgers, J.T. Neuronal SIRT1 activation as a novel mechanism underlying the prevention of Alzheimer disease amyloid neuropathology by calorie restriction. J. Biol. Chem. 2006, 281, 21745–21754. [Google Scholar] [CrossRef] [Green Version]
- Halagappa, V.K.; Guo, Z.; Pearson, M.; Matsuoka, Y.; Cutler, R.G.; Laferla, F.M.; Mattson, M.P. Intermittent fasting and caloric restriction ameliorate age-related behavioral deficits in the triple-transgenic mouse model of Alzheimer’s disease. Neurobiol. Dis. 2007, 26, 212–220. [Google Scholar] [CrossRef]
- Bok, E.; Jo, M.; Lee, S.; Lee, B.R.; Kim, J.; Kim, H.J. Dietary Restriction and Neuroinflammation: A Potential Mechanistic Link. Int. J. Mol. Sci. 2019, 20, 464. [Google Scholar] [CrossRef] [Green Version]
- Irizarry, M.C.; Soriano, F.; McNamara, M.; Page, K.J.; Schenk, D.; Games, D.; Hyman, B.T. Aβ deposition is associated with neuropil changes, but not with overt neuronal loss in the human amyloid precursor protein V717F (PDAPP) transgenic mouse. J. Neurosci. 1997, 17, 7053–7059. [Google Scholar] [CrossRef]
- Westerman, M.A.; Cooper-Blacketer, D.; Mariash, A.; Kotilinek, L.; Kawarabayashi, T.; Younkin, L.H.; Carlson, G.A.; Younkin, S.G.; Ashe, K.H. The relationship between Aβ and memory in the Tg2576 mouse model of Alzheimer’s disease. J. Neurosci. 2002, 22, 1858–1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieschke, J.; Herbst, M.; Wiglenda, T.; Friedrich, R.P.; Boeddrich, A.; Schiele, F.; Kleckers, D.; del Amo, J.M.L.; Grüning, B.A.; Wang, Q. Small-molecule conversion of toxic oligomers to nontoxic β-sheet–rich amyloid fibrils. Nat. Chem. Biol. 2012, 8, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Tanei, Z.I.; Hashimoto, T.; Wakabayashi, T.; Okuno, H.; Naka, Y.; Yizhar, O.; Fenno, L.E.; Fukayama, M.; Bito, H.; et al. Chronic optogenetic activation augments abeta pathology in a mouse model of Alzheimer disease. Cell Rep. 2015, 11, 859–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamenetz, F.; Tomita, T.; Hsieh, H.; Seabrook, G.; Borchelt, D.; Iwatsubo, T.; Sisodia, S.; Malinow, R. APP processing and synaptic function. Neuron 2003, 37, 925–937. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, M.; Perfilieva, E.; Johansson, U.; Orwar, O.; Eriksson, P.S. Enriched environment increases neurogenesis in the adult rat dentate gyrus and improves spatial memory. J. Neurobiol. 1999, 39, 569–578. [Google Scholar] [CrossRef]
- Xu, H.; Gelyana, E.; Rajsombath, M.; Yang, T.; Li, S.; Selkoe, D. Environmental enrichment potently prevents microglia-mediated neuroinflammation by human amyloid β-protein oligomers. J. Neurosci. 2016, 36, 9041–9056. [Google Scholar] [CrossRef] [Green Version]
- Beauquis, J.; Pavía, P.; Pomilio, C.; Vinuesa, A.; Podlutskaya, N.; Galvan, V.; Saravia, F. Environmental enrichment prevents astroglial pathological changes in the hippocampus of APP transgenic mice, model of Alzheimer’s disease. Exp. Neurol. 2013, 239, 28–37. [Google Scholar] [CrossRef]
- Ngandu, T.; Lehtisalo, J.; Solomon, A.; Levalahti, E.; Ahtiluoto, S.; Antikainen, R.; Backman, L.; Hanninen, T.; Jula, A.; Laatikainen, T.; et al. A 2 year multidomain intervention of diet, exercise, cognitive training, and vascular risk monitoring versus control to prevent cognitive decline in at-risk elderly people (FINGER): A randomised controlled trial. Lancet 2015, 385, 2255–2263. [Google Scholar] [CrossRef]
- Dhana, K.; Evans, D.A.; Rajan, K.B.; Bennett, D.A.; Morris, M.C. Impact of healthy lifestyle factors on the risk of Alzheimer’s dementia: Findings from two prospective cohort studies. Alzheimers Dement. 2019, 15, P207. [Google Scholar] [CrossRef]
- Kuzma, E.; Lourida, I.; Hannon, E.; Littlejohns, T.J.; Langa, K.M.; Hyppönen, E.; Llewellyn, D.J. Genetic risk, lifestyle, and dementia. Alzheimers Dement. 2019, 15, P890. [Google Scholar] [CrossRef]
- Davis, J.; Xu, F.; Miao, J.; Previti, M.L.; Romanov, G.; Ziegler, K.; Van Nostrand, W.E. Deficient cerebral clearance of vasculotropic mutant Dutch/Iowa Double Aß in human AßPP transgenic mice. Neurobiol. Aging 2006, 27, 946–954. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Xu, F.; Deane, R.; Romanov, G.; Previti, M.L.; Zeigler, K.; Zlokovic, B.V.; Van Nostrand, W.E. Early-onset and robust cerebral microvascular accumulation of amyloid β-protein in transgenic mice expressing low levels of a vasculotropic Dutch/Iowa mutant form of amyloid β-protein precursor. J. Biol. Chem. 2004, 279, 20296–20306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, J.; Xu, F.; Davis, J.; Otte-Höller, I.; Verbeek, M.M.; Van Nostrand, W.E. Cerebral microvascular amyloid β protein deposition induces vascular degeneration and neuroinflammation in transgenic mice expressing human vasculotropic mutant amyloid β precursor protein. Am. J. Pathol. 2005, 167, 505–515. [Google Scholar] [CrossRef]
- Xu, F.; Grande, A.M.; Robinson, J.K.; Previti, M.L.; Vasek, M.; Davis, J.; Van Nostrand, W.E. Early-Onset Subicular Microvascular Amyloid and Neuroinflammation Correlate With Behavioral Deficits in Vasculotropic Mutant APP Transgenic Mice. Neuroscience 2007, 146, 98–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharm. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Deacon, R.M.J. Digging and marble burying in mice: Simple methods for in vivo identification of biological impacts. Nat. Protoc. 2006, 1, 122–124. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robison, L.S.; Francis, N.; Popescu, D.L.; Anderson, M.E.; Hatfield, J.; Xu, F.; Anderson, B.J.; Van Nostrand, W.E.; Robinson, J.K. Environmental Enrichment: Disentangling the Influence of Novelty, Social, and Physical Activity on Cerebral Amyloid Angiopathy in a Transgenic Mouse Model. Int. J. Mol. Sci. 2020, 21, 843. https://doi.org/10.3390/ijms21030843
Robison LS, Francis N, Popescu DL, Anderson ME, Hatfield J, Xu F, Anderson BJ, Van Nostrand WE, Robinson JK. Environmental Enrichment: Disentangling the Influence of Novelty, Social, and Physical Activity on Cerebral Amyloid Angiopathy in a Transgenic Mouse Model. International Journal of Molecular Sciences. 2020; 21(3):843. https://doi.org/10.3390/ijms21030843
Chicago/Turabian StyleRobison, Lisa S., Nikita Francis, Dominique L. Popescu, Maria E. Anderson, Joshua Hatfield, Feng Xu, Brenda J. Anderson, William E. Van Nostrand, and John K. Robinson. 2020. "Environmental Enrichment: Disentangling the Influence of Novelty, Social, and Physical Activity on Cerebral Amyloid Angiopathy in a Transgenic Mouse Model" International Journal of Molecular Sciences 21, no. 3: 843. https://doi.org/10.3390/ijms21030843