Applicability of Anti-Human Estrogen Receptor β Antibody PPZ0506 for the Immunodetection of Rodent Estrogen Receptor β Proteins

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Specificity and Cross-Reactivity of PPZ0506 Antibody Against Human, Mouse, and Rat ERβ Proteins
2.1.1. Construction of Expression Vectors Encoding Human, Mouse, and Rat ERβ
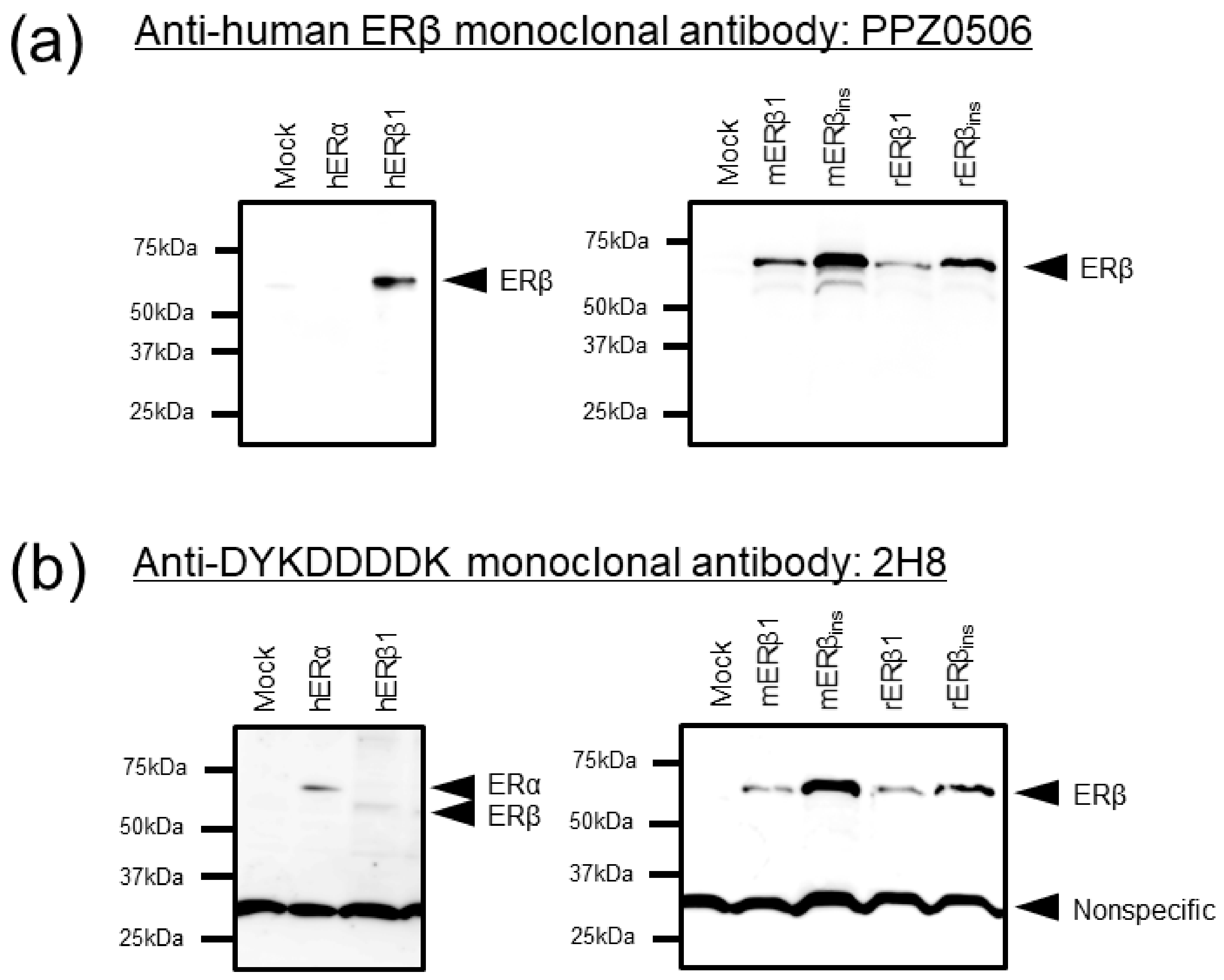
2.1.2. Immunoblot Analyses of Human, Mouse, and Rat ERβ in Transfected Cells
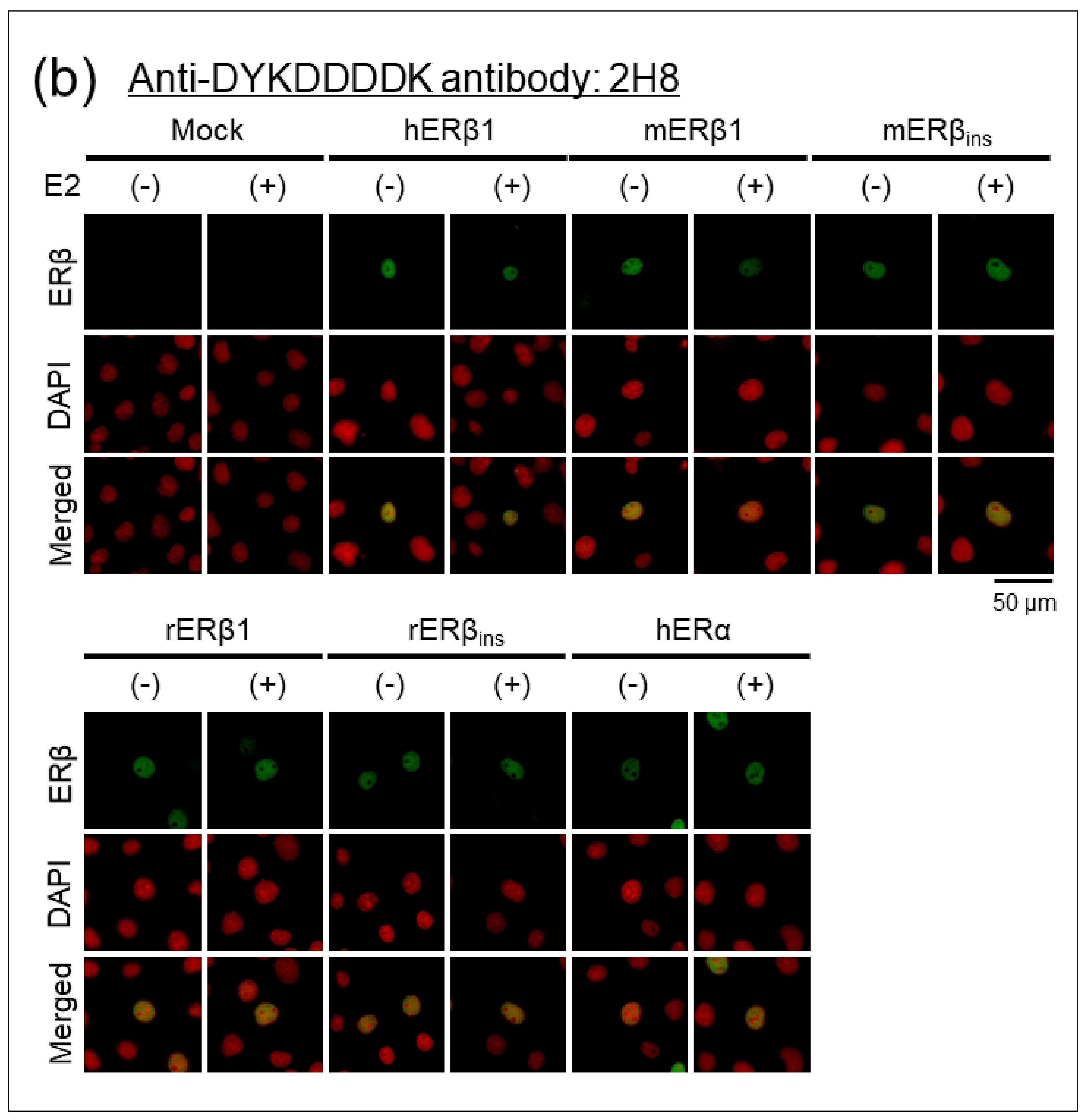
2.1.3. Immunocytochemical Analyses of Human, Mouse, and Rat ERβ in Transfected Cells
2.2. Immunohistochemical Analyses of ERβ Proteins in Rat Organs
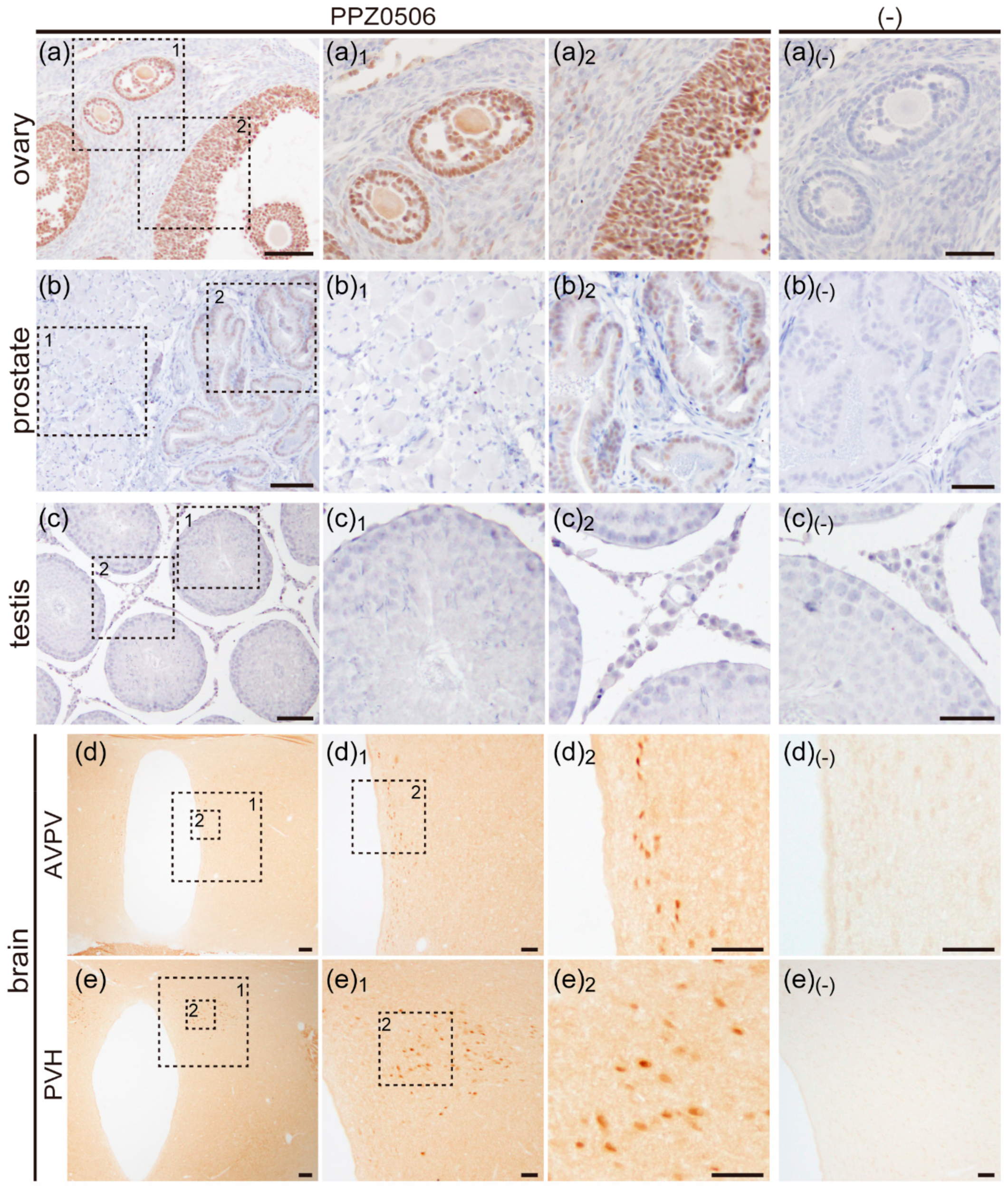
2.2.1. Expression of ERβ Proteins in Rat Ovary
2.2.2. Expression of ERβ Proteins in Rat Prostate
2.2.3. Expression of ERβ Proteins in Rat Testis
2.2.4. Expression of ERβ Proteins in Rat Brain
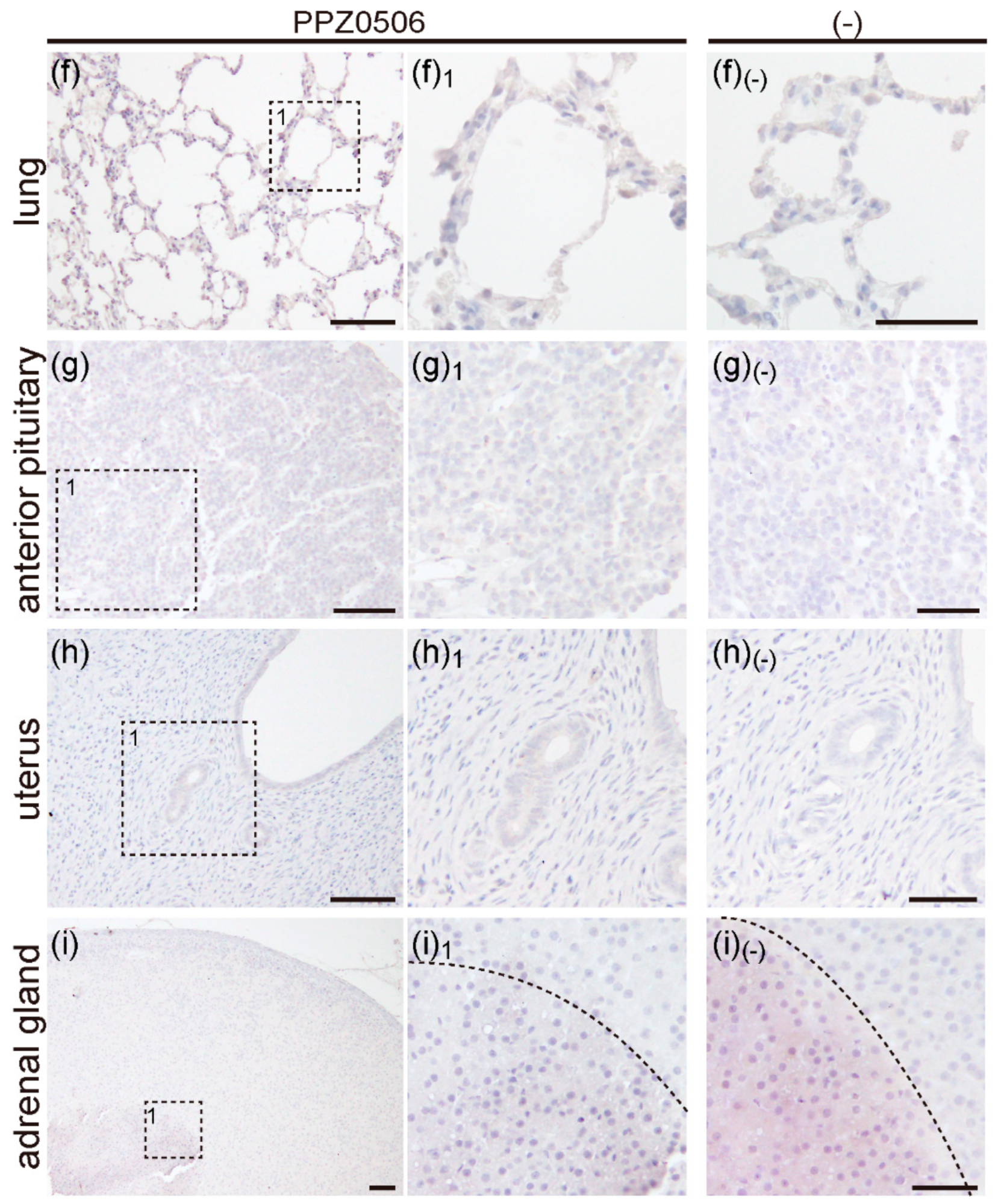
2.2.5. Expression of ERβ Proteins in Rat Lung, Anterior Pituitary, Uterus and, Adrenal Gland
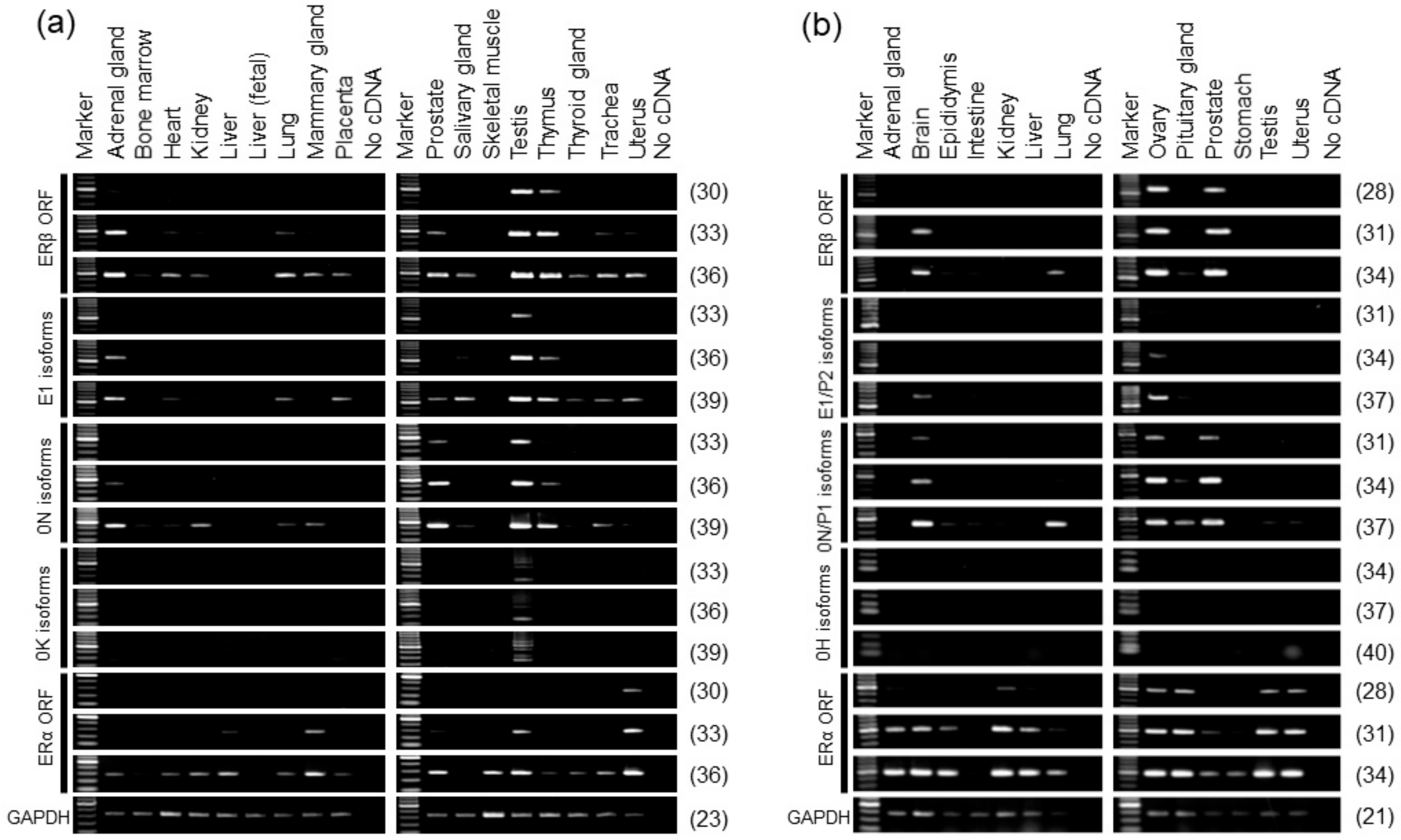
2.3. Alternative Promoter Usage Profiles of Human and Rat ERβ Genes
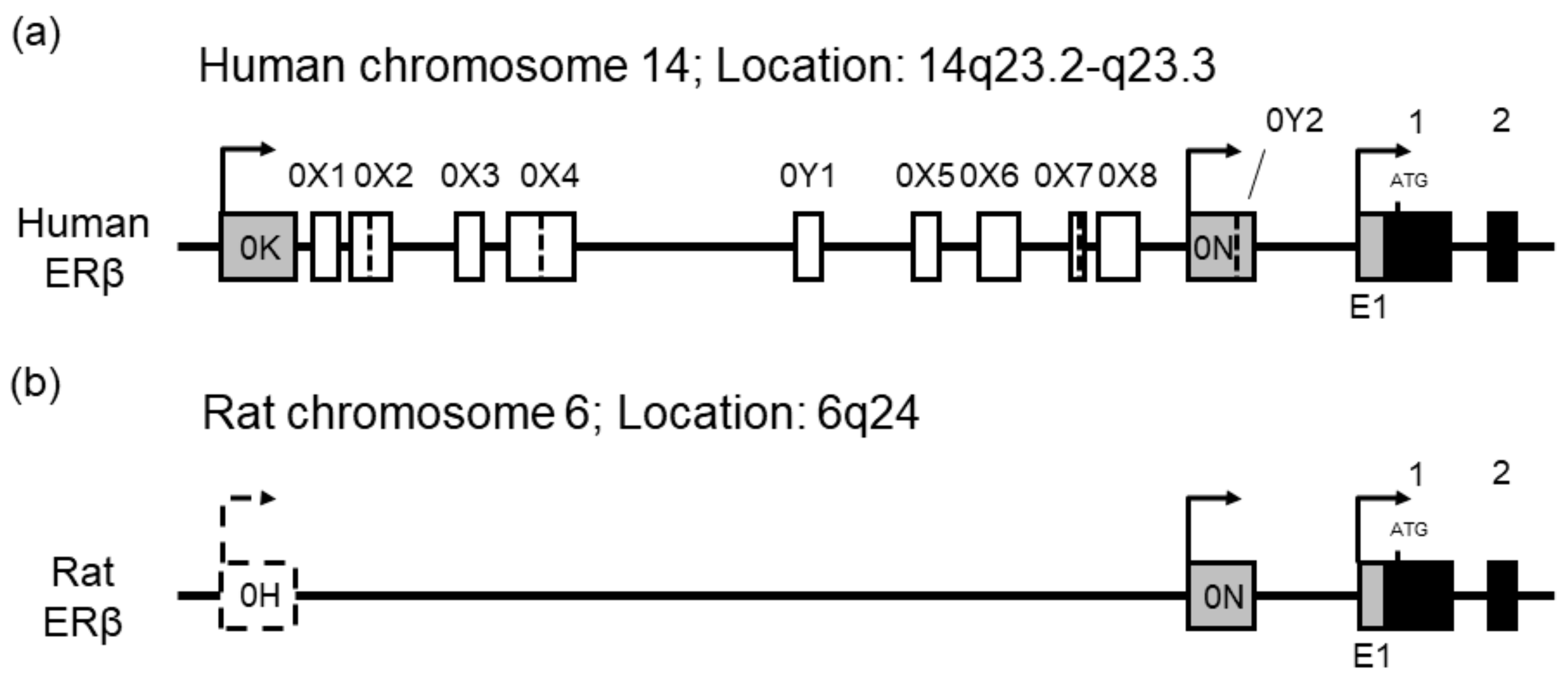
2.3.1. Genomic Organization of Human and Rat ERβ Genes
2.3.2. Alternative Promoter Usage of Human ERβ Gene
2.3.3. Alternative Promoter Usage of Rat ERβ Gene
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Total RNA Extraction, 5′-RACE, and RT-PCR
4.3. Vector Construction
4.4. Cell Culture and Transfection
4.5. Immunoblotting
4.6. Immunocytochemistry
4.7. Immunohistochemistry
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ER | Estrogen receptor |
| ERα | Estrogen receptor α |
| ERβ | Estrogen receptor β |
| ERE | Estrogen response element |
| E2 | 17β-estradiol |
| EtOH | Ethanol |
| DAPI | 4,6-diamino-2-phenylindole |
| AVPV | Anteroventral paraventricular nucleus |
| PVH | Paraventricular nucleus of hypothalamus |
| UTR | Untranslated region |
| RACE | Rapid amplification of cDNA end |
| ORF | Open reading frame |
| FBS | Fetal bovine serum |
| DMEM | Dulbecco’s modified Eagle medium |
| PBS | Phosphate-buffed saline |
| PB | Phosphate buffer |
| TBS | Tris-buffered saline |
| BSA | Bovine serum albumin |
References
- Nilsson, S.; Gustafsson, J.A. Estrogen receptors: Their actions and functional roles in health and disease. In Nuclear Receptors, Proteins and Cell Regulation; Bunce, C., Campbell, M., Eds.; Springer: Berlin, Germany, 2010; Volume 8, pp. 91–141. [Google Scholar]
- Nuclear Receptors Nomenclature Committee. A unified nomenclature system for the nuclear receptor superfamily. Cell 1999, 97, 161–163. [Google Scholar] [CrossRef] [Green Version]
- Hirata, S.; Shoda, T.; Kato, J.; Hoshi, K. Isoform/variant mRNAs for sex steroid hormone receptors in humans. Trends Endocrinol. Metab. 2003, 14, 124–129. [Google Scholar] [CrossRef]
- Kos, M.; Reid, G.; Denger, S.; Gannon, F. Minireview: Genomic organization of the human ERalpha gene promoter region. Mol. Endocrinol. 2001, 15, 2057–2063. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Ishii, H.; Morita, A.; Sakuma, Y.; Ozawa, H. Characterization of the fundamental properties of the N-terminal truncation (Δ exon 1) variant of estrogen receptor α in the rat. Gene 2015, 571, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Kobayashi, M.; Munetomo, A.; Miyamoto, T.; Sakuma, Y. Novel splicing events and post-transcriptional regulation of human estrogen receptor α E isoforms. J. Steroid Biochem. Mol. Biol. 2013, 133, 120–128. [Google Scholar] [CrossRef]
- Ishii, H.; Sakuma, Y. Complex organization of the 5′-untranslated region of the mouse estrogen receptor α gene: Identification of numerous mRNA transcripts with distinct 5′-ends. J. Steroid Biochem. Mol. Biol. 2011, 125, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ishii, H.; Sakuma, Y. Identification of novel splicing events and post-transcriptional regulation of human estrogen receptor α F isoforms. Mol. Cell. Endocrinol. 2011, 333, 55–61. [Google Scholar] [CrossRef]
- Ishii, H.; Kobayashi, M.; Sakuma, Y. Alternative promoter usage and alternative splicing of the rat estrogen receptor alpha gene generate numerous mRNA variants with distinct 5′-ends. J. Steroid Biochem. Mol. Biol. 2010, 118, 59–69. [Google Scholar] [CrossRef]
- Lee, M.T.; Ouyang, B.; Ho, S.M.; Leung, Y.K. Differential expression of estrogen receptor beta isoforms in prostate cancer through interplay between transcriptional and translational regulation. Mol. Cell. Endocrinol. 2013, 376, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.; Brannan, R.A.; Hanby, A.M.; Shaaban, A.M.; Verghese, E.T.; Peter, M.B.; Pollock, S.; Satheesha, S.; Szynkiewicz, M.; Speirs, V.; et al. Differential regulation of oestrogen receptor β isoforms by 5’ untranslated regions in cancer. J. Cell. Mol. Med. 2010, 14, 2172–2184. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.; Coleman, L.J.; Cummings, M.; Satheesha, S.; Shaw, S.O.; Speirs, V.; Hughes, T.A. Expression of oestrogen receptor beta isoforms is regulated by transcriptional and post-transcriptional mechanisms. Biochem. J. 2010, 429, 283–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamoto, H.; Hirata, S.; Shoda, T.; Kato, J.; Hoshi, K. The novel 5’-untranslated first exon, exon 0H, of the rat estrogen receptor beta gene. Endocr. Res. 2003, 29, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Hirata, S.; Shoda, T.; Kato, J.; Hoshi, K. The multiple untranslated first exons system of the human estrogen receptor beta (ER beta) gene. J. Steroid Biochem. Mol. Biol. 2001, 78, 33–40. [Google Scholar] [CrossRef]
- O’Brien, M.L.; Park, K.; In, Y.; Park-Sarge, O.K. Characterization of estrogen receptor-beta (ERbeta) messenger ribonucleic acid and protein expression in rat granulosa cells. Endocrinology 1999, 140, 4530–4541. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.A. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef] [Green Version]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Häggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [CrossRef]
- Mosselman, S.; Polman, J.; Dijkema, R. ER beta: Identification and characterization of a novel human estrogen receptor. FEBS Lett. 1996, 392, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.T.; McKee, D.D.; Slentz-Kesler, K.; Moore, L.B.; Jones, S.A.; Horne, E.L.; Su, J.L.; Kliewer, S.A.; Lehmann, J.M.; Willson, T.M. Cloning and characterization of human estrogen receptor beta isoforms. Biochem. Biophys. Res. Commun. 1998, 247, 75–78. [Google Scholar] [CrossRef]
- Van Pelt, A.M.; de Rooij, D.G.; van der Burg, B.; van der Saag, P.T.; Gustafsson, J.A.; Kuiper, G.G. Ontogeny of estrogen receptor-beta expression in rat testis. Endocrinology 1999, 140, 478–483. [Google Scholar] [CrossRef]
- Shughrue, P.J.; Lane, M.V.; Scrimo, P.J.; Merchenthaler, I. Comparative distribution of estrogen receptor-alpha (ER-alpha) and beta (ER-beta) mRNA in the rat pituitary, gonad, and reproductive tract. Steroids 1998, 63, 498–504. [Google Scholar] [CrossRef]
- Saunders, P.T.; Fisher, J.S.; Sharpe, R.M.; Millar, M.R. Expression of oestrogen receptor beta (ER beta) occurs in multiple cell types, including some germ cells, in the rat testis. J. Endocrinol. 1998, 156, R13–R17. [Google Scholar] [CrossRef]
- Saunders, P.T.; Maguire, S.M.; Gaughan, J.; Millar, M.R. Expression of oestrogen receptor beta (ER beta) in multiple rat tissues visualised by immunohistochemistry. J. Endocrinol. 1997, 154, R13–R16. [Google Scholar] [CrossRef]
- Snyder, M.A.; Smejkalova, T.; Forlano, P.M.; Woolley, C.S. Multiple ERbeta antisera label in ERbeta knockout and null mouse tissues. J. Neurosci. Methods 2010, 188, 226–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skliris, G.P.; Parkes, A.T.; Limer, J.L.; Burdall, S.E.; Carder, P.J.; Speirs, V. Evaluation of seven oestrogen receptor beta antibodies for immunohistochemistry, western blotting, and flow cytometry in human breast tissue. J. Pathol. 2002, 197, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Weitsman, G.E.; Skliris, G.; Ung, K.; Peng, B.; Younes, M.; Watson, P.H.; Murphy, L.C. Assessment of multiple different estrogen receptor-beta antibodies for their ability to immunoprecipitate under chromatin immunoprecipitation conditions. Breast Cancer Res. Treat. 2006, 100, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Andersson, S.; Sundberg, M.; Pristovsek, N.; Ibrahim, A.; Jonsson, P.; Katona, B.; Clausson, C.M.; Zieba, A.; Ramström, M.; Söderberg, O.; et al. Insufficient antibody validation challenges oestrogen receptor beta research. Nat. Commun. 2017, 8, 15840. [Google Scholar] [CrossRef] [PubMed]
- Hawse, J.R.; Carter, J.M.; Aspros, K.G.M.; Bruinsma, E.S.; Koepplin, J.W.; Negron, V.; Subramaniam, M.; Ingle, J.N.; Rech, K.L.; Goetz, M.P. Optimized immunohistochemical detection of estrogen receptor beta using two validated monoclonal antibodies confirms its expression in normal and malignant breast tissues. Breast Cancer Res. Treat. 2019. In press. [Google Scholar] [CrossRef]
- Leygue, E.; Dotzlaw, H.; Lu, B.; Glor, C.; Peter, H.; Watson, P.H.; Murphy, L.C. Estrogen receptor beta: Mine is longer than yours? J. Clin. Endocrinol. Metab. 1998, 83, 3754–3755. [Google Scholar]
- Petersen, D.N.; Tkalcevic, G.T.; Koza-Taylor, P.H.; Turi, T.G.; Brown, T.A. Identification of estrogen receptor beta2, a functional variant of estrogen receptor beta expressed in normal rat tissues. Endocrinology 1998, 139, 1082–1092. [Google Scholar] [CrossRef]
- Lu, B.; Leygue, E.; Dotzlaw, H.; Murphy, L.J.; Murphy, L.C.; Watson, P.H. Estrogen receptor-beta mRNA variants in human and murine tissues. Mol. Cell. Endocrinol. 1998, 138, 199–203. [Google Scholar] [CrossRef]
- Chu, S.; Fuller, P.J. Identification of a splice variant of the rat estrogen receptor beta gene. Mol. Cell. Endocrinol. 1997, 132, 195–199. [Google Scholar] [CrossRef]
- Yang, S.H.; Liu, R.; Perez, E.J.; Wen, Y.; Stevens, S.M., Jr.; Valencia, T.; Brun-Zinkernagel, A.M.; Prokai, L.; Will, Y.; Dykens, J.; et al. Mitochondrial localization of estrogen receptor beta. Proc. Natl. Acad. Sci. USA 2004, 101, 4130–4135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilad, L.A.; Schwartz, B. Association of estrogen receptor beta with plasma-membrane caveola components: Implication in control of vitamin D receptor. J. Mol. Endocrinol. 2007, 38, 603–618. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Ishii, H.; Munetomo, A.; Watanabe, H.; Morita, A.; Sakuma, Y.; Ozawa, H. Human C-terminally truncated ERα variants resulting from the use of alternative exons in the ligand-binding domain. Mol. Cell. Endocrinol. 2016, 425, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Hattori, Y.; Munetomo, A.; Watanabe, H.; Sakuma, Y.; Ozawa, H. Characterization of rodent constitutively active estrogen receptor α variants and their constitutive transactivation mechanisms. Gen. Comp. Endocrinol. 2017, 248, 16–26. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishii, H.; Otsuka, M.; Kanaya, M.; Higo, S.; Hattori, Y.; Ozawa, H. Applicability of Anti-Human Estrogen Receptor β Antibody PPZ0506 for the Immunodetection of Rodent Estrogen Receptor β Proteins. Int. J. Mol. Sci. 2019, 20, 6312. https://doi.org/10.3390/ijms20246312
Ishii H, Otsuka M, Kanaya M, Higo S, Hattori Y, Ozawa H. Applicability of Anti-Human Estrogen Receptor β Antibody PPZ0506 for the Immunodetection of Rodent Estrogen Receptor β Proteins. International Journal of Molecular Sciences. 2019; 20(24):6312. https://doi.org/10.3390/ijms20246312
Chicago/Turabian StyleIshii, Hirotaka, Mai Otsuka, Moeko Kanaya, Shimpei Higo, Yujiro Hattori, and Hitoshi Ozawa. 2019. "Applicability of Anti-Human Estrogen Receptor β Antibody PPZ0506 for the Immunodetection of Rodent Estrogen Receptor β Proteins" International Journal of Molecular Sciences 20, no. 24: 6312. https://doi.org/10.3390/ijms20246312