Forsythoside A and Forsythoside B Contribute to Shuanghuanglian Injection-Induced Pseudoallergic Reactions through the RhoA/ROCK Signaling Pathway

 , ,
, , 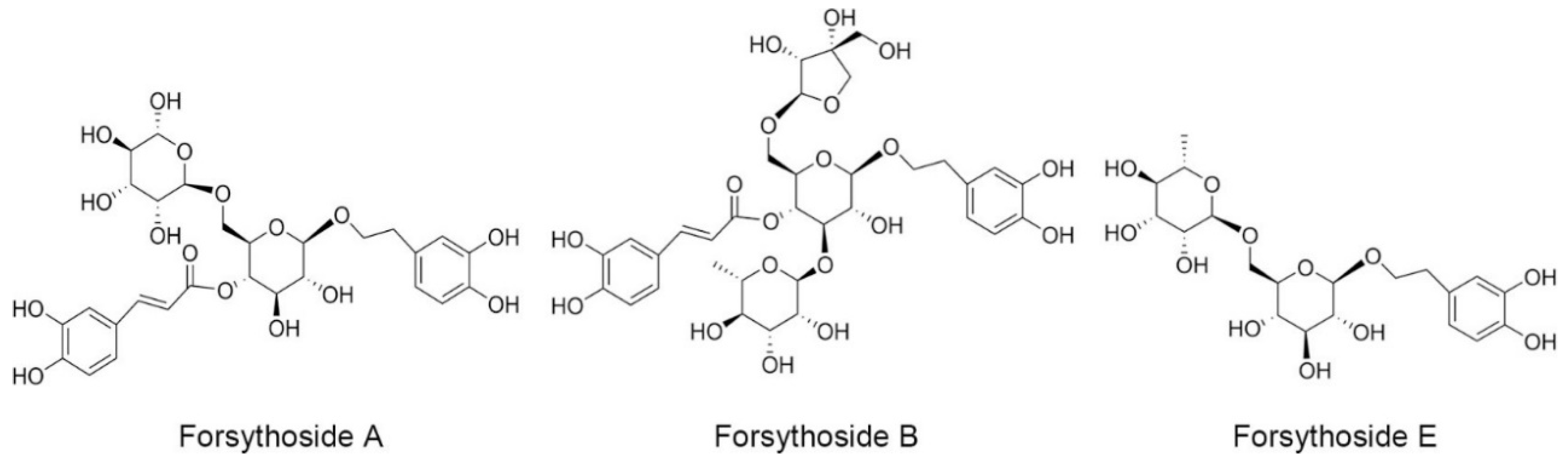{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
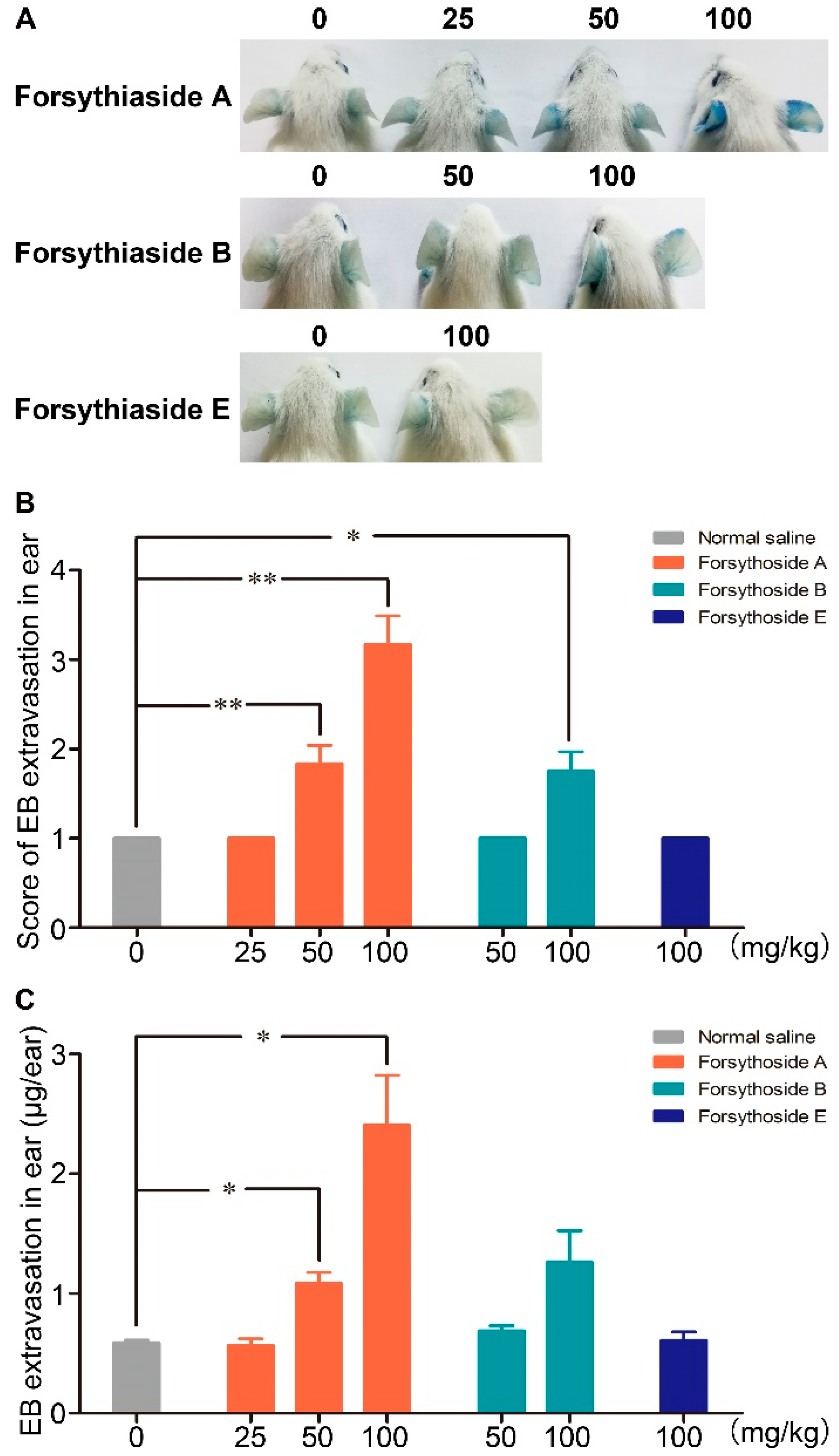
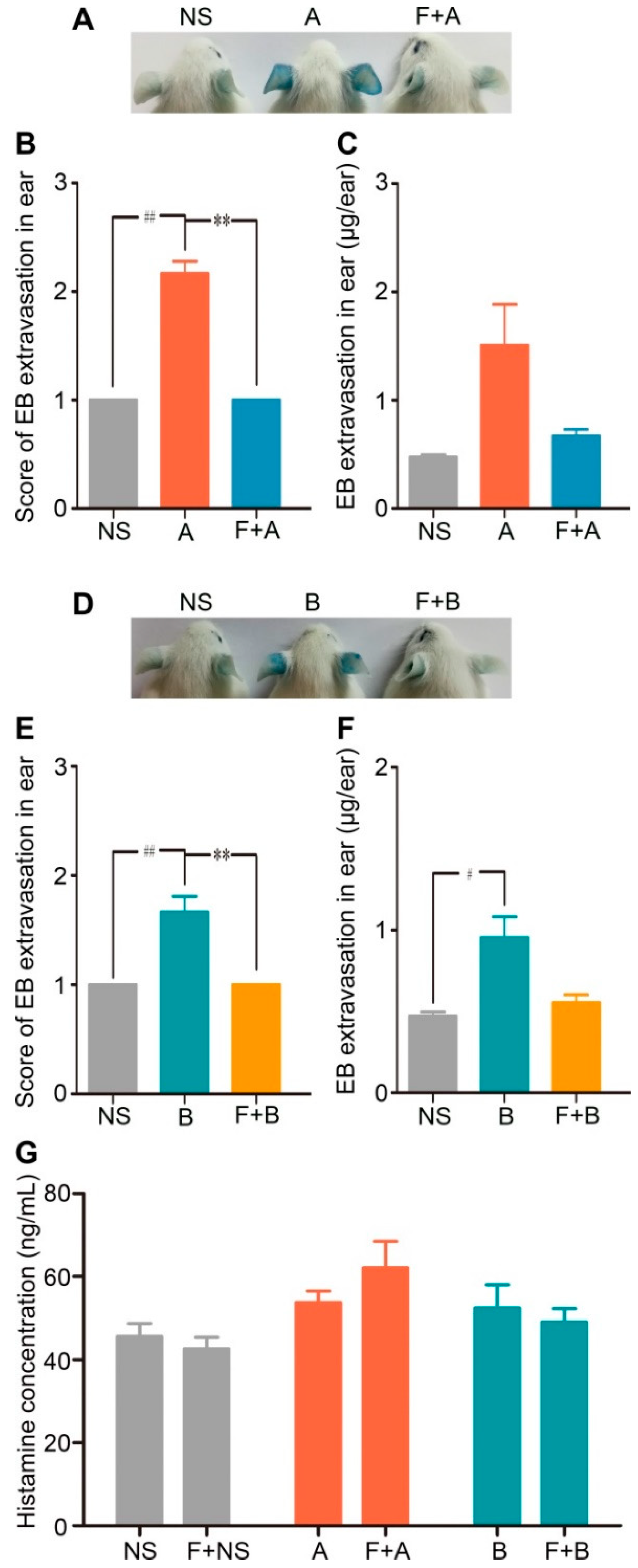
2.1. Forsythoside A and Forsythoside B Induce Vascular Leakage in Mice
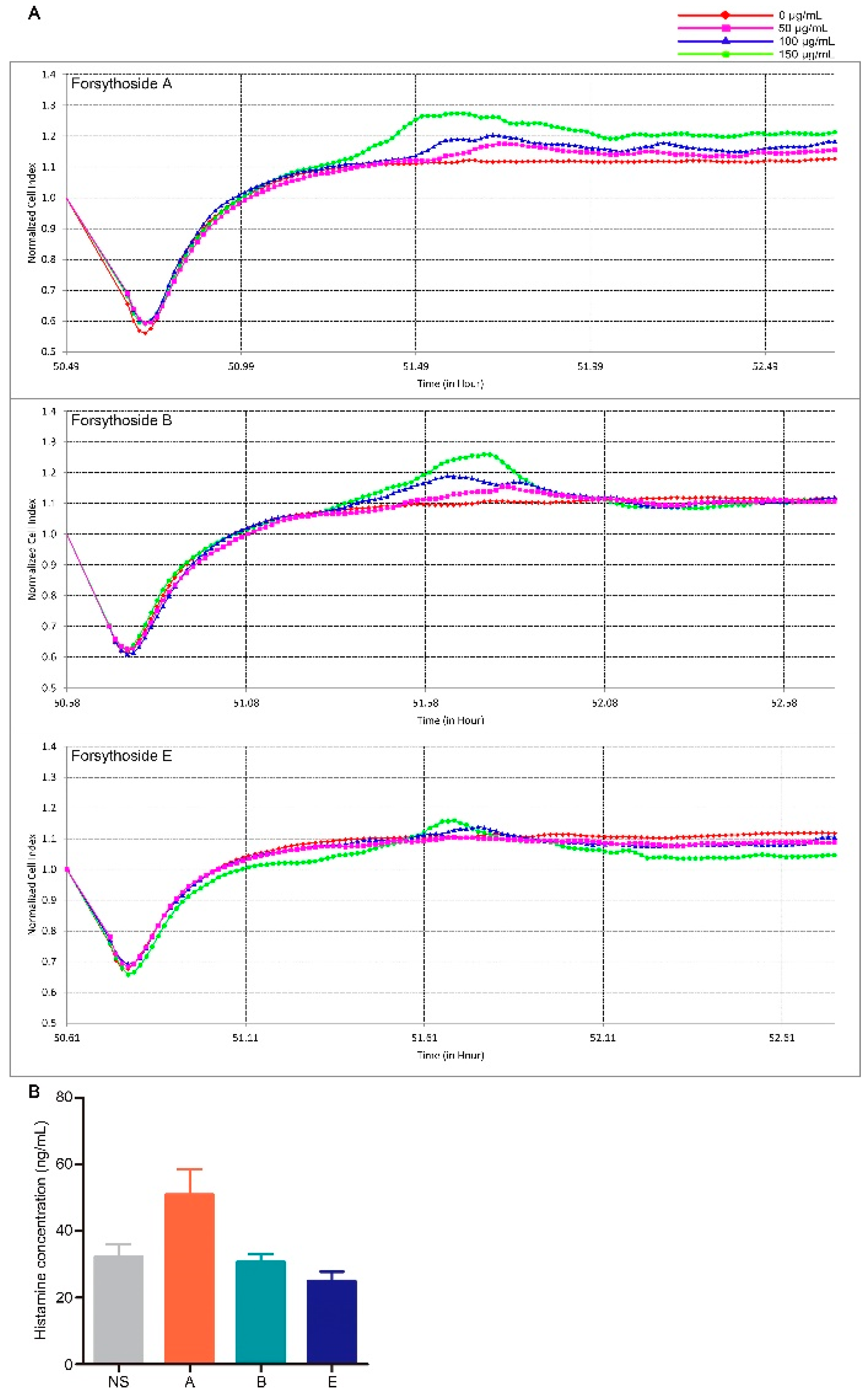
2.2. Forsythoside A and Forsythoside B Slightly Evoke Mast Cell Degranulation
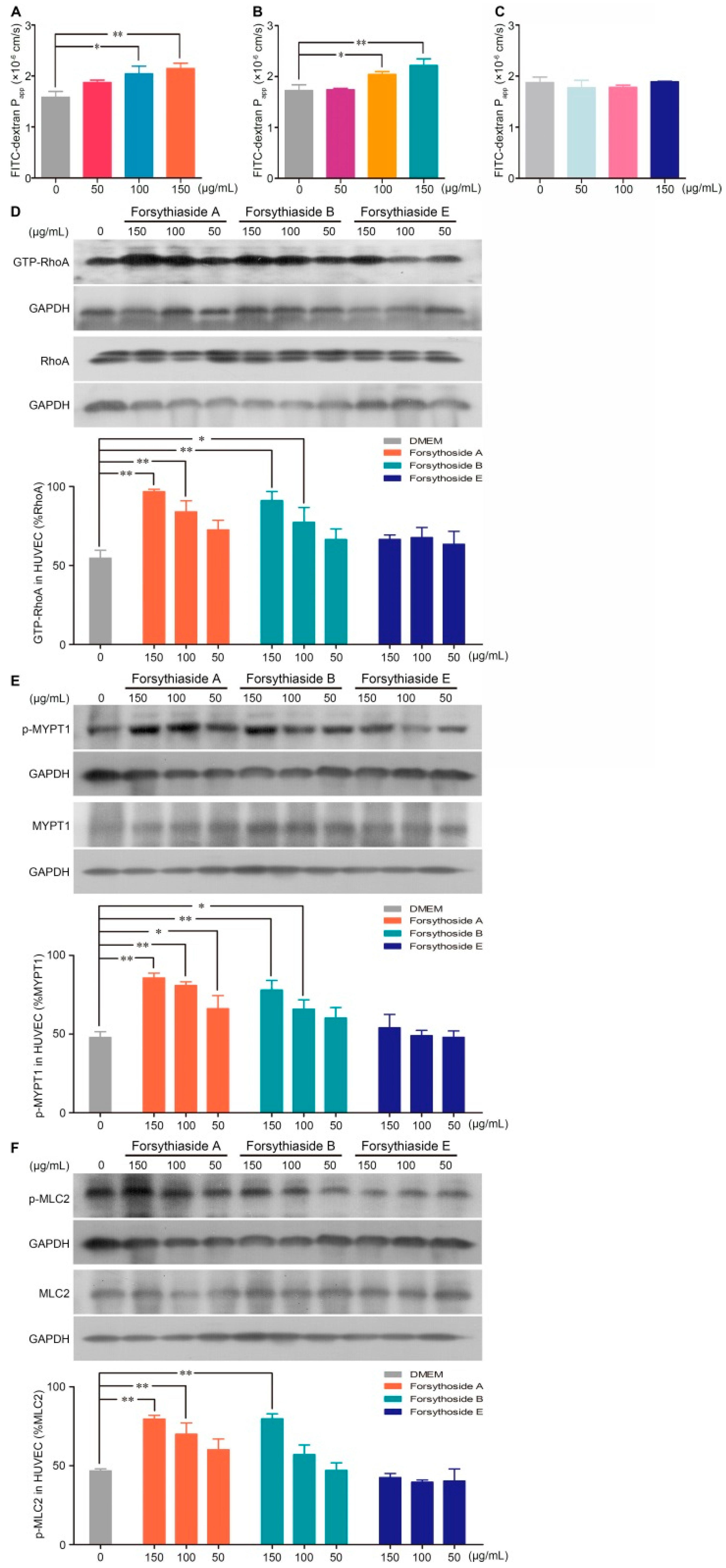
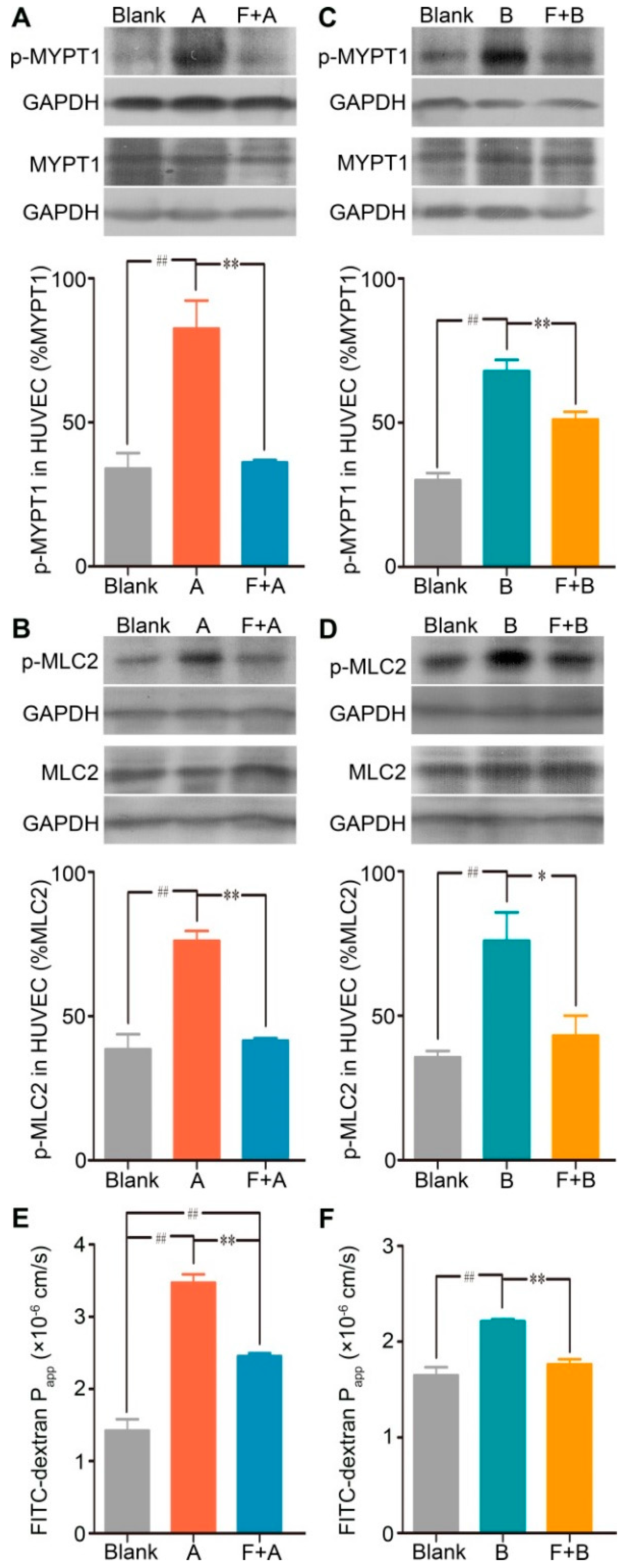
2.3. Forsythoside A and Forsythoside B Enhance Endothelial Permeability via Activation of the RhoA/ROCK Signaling Pathway
2.4. Fasudil Attenuates the Forsythoside A/Forsythoside B-induced Endothelial Monolayer and Vascular Hyperpermeability
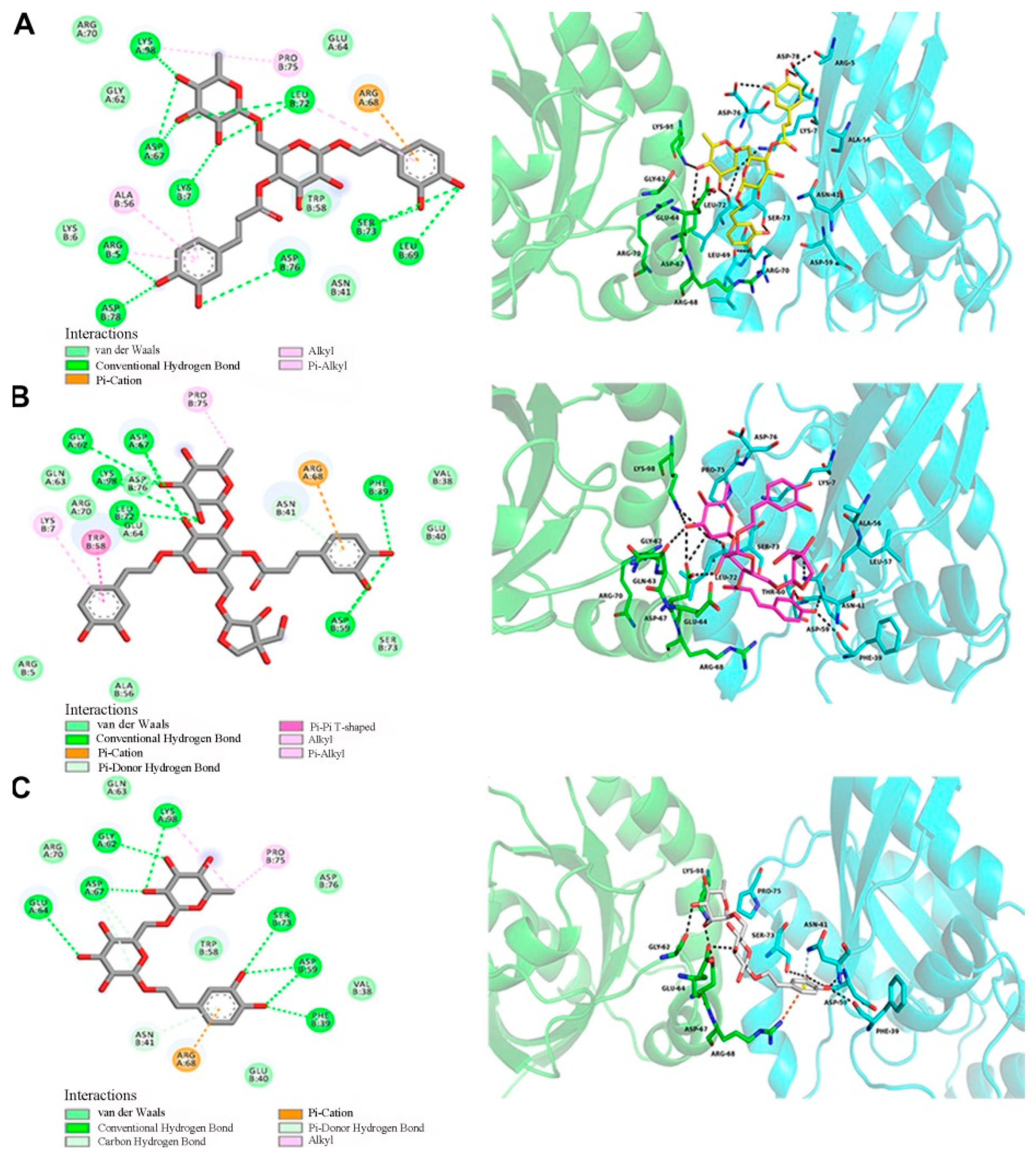
2.5. Docking Analysis of GTP-RhoA with Forsythoside A, Forsythoside B, and Forsythoside E
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Reagents
4.3. Animals
4.4. Cell Culture
4.5. Assessment of Vascular Leakage in Mice
4.6. Histamine Assay
4.7. Assessment of Mast Cell Degranulation
4.8. Endothelial Permeability Assay
4.9. Preparation of Cell Lysates
4.10. Immunoblot Analysis
4.11. Molecular Docking
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, H.; Chen, Q. Chinese medicine injection Shuanghuanglian for treatment of acute upper respiratory tract infection: A systematic review of randomized controlled trials. Evid. Based Complement. Altern. Med. 2013, 2013, 987326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Douglas, R.M. Chinese herbal medicines in the treatment of acute respiratory infections: A review of randomised and controlled clinical trials. Med. J. Aust. 1998, 169, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Kua, K.P.; Lee, S.W. Complementary and alternative medicine for the treatment of bronchiolitis in infants: A systematic review. PLoS ONE 2017, 12, e0172289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.T.; Fang, H.T. Treatment of acute bronchiolitis with Chinese herbs. Arch. Dis. Child. 1993, 68, 468–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Cheng, L. Adverse drug reactions of Shuanghuanglian Injection: A systematic review of public literatures. J. Evid. Based Med. 2010, 3, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Li, J. Analysis of 56 cases of adverse drug reactions/events caused by Shuanghuanglian Injection. China Foreign Med. Treat. 2010, 7, 125–126. [Google Scholar]
- Shang, Y. Analysis of adverse drug reactions caused by Shuanghuanglian Injection. Chin. Med. Mod. Distance Educ. China 2014, 12, 109. [Google Scholar]
- Kumar, P.; Shen, Q. Molecular mechanisms of endothelial hyperpermeability: Implications in inflammation. Expert Rev. Mol. Med. 2009, 11, e19. [Google Scholar] [CrossRef] [Green Version]
- Spindler, V.; Schlegel, N. Role of GTPases in control of microvascular permeability. Cardiovasc. Res. 2010, 87, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Wojciak-Stothard, B.; Ridley, A.J. Rho GTPases and the regulation of endothelial permeability. Vasc. Pharmacol. 2002, 39, 187–199. [Google Scholar] [CrossRef]
- Narumiya, S.; Thumkeo, D. Rho signaling research: History, current status and future directions. FEBS Lett. 2018, 592, 1763–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Loredo, A.Y.; Lopez, E. Thrombin promotes actin stress fiber formation in RPE through Rho/ROCK-mediated MLC phosphorylation. J. Cell Physiol. 2011, 226, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Mikelis, C.M.; Simaan, M. RhoA and ROCK mediate histamine-induced vascular leakage and anaphylactic shock. Nat. Commun. 2015, 6, 6725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Qin, J. Role of Rho kinase in lysophosphatidic acid-induced altering of blood-brain barrier permeability. Int. J. Mol. Med. 2014, 33, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Yi, Y. Involvement of histamine and RhoA/ROCK in penicillin immediate hypersensitivity reactions. Sci. Rep. 2016, 6, 33192. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Zhao, Y. RhoA/ROCK signaling pathway mediates Shuanghuanglian Injection-induced pseudo-allergic reactions. Front. Pharmacol. 2018, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.; Zhang, Y.S. Study of screening pseudoallergenic substances of Shuanghuanglian Injection. Zhongguo Zhong Yao Za Zhi 2015, 40, 2727–2731. [Google Scholar]
- Wang, Z.; Xia, Q. Phytochemistry, pharmacology, quality control and future research of Forsythia suspensa (Thunb.) Vahl: A review. J. Ethnopharmacol. 2018, 210, 318–339. [Google Scholar] [CrossRef]
- Dong, Z.; Lu, X. Forsythiae Fructus: A review on its phytochemistry, quality control, pharmacology and pharmacokinetics. Molecules 2017, 22, 1466. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Liu, M. Comprehensive identification of 125 multifarious constituents in Shuang-huang-lian powder injection by HPLC-DAD-ESI-IT-TOF-MS. J. Pharm. Biomed. Anal. 2015, 115, 86–106. [Google Scholar] [CrossRef]
- Guo, M.; Zhao, B. A metabolomic strategy to screen the prototype components and metabolites of Shuang-Huang-Lian Injection in human serum by ultra performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. J. Anal. Methods Chem. 2014, 2014, 241505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, H.S. Agents that induce pseudo-allergic reaction. Drug Discov. Ther. 2011, 5, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X. Identification of a specific agonist of human TAS2R14 from Radix Bupleuri through virtual screening, functional evaluation and binding studies. Sci. Rep. 2017, 7, 12174. [Google Scholar] [CrossRef] [PubMed]
- Roshan Moniri, M.; Young, A. Dynamic assessment of cell viability, proliferation and migration using real time cell analyzer system (RTCA). Cytotechnology 2015, 67, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, G.; Du, Q. Application of real-time cell electronic analysis system in modern pharmaceutical evaluation and analysis. Molecules 2018, 23, 3280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Zhang, E. Determination of Shuanghuanglian Injection of chlorogenic acid sensitized rabbit specific antibody in serum. Pharmacol. Clin. Chin. Mater. Med. 2016, 32, 70–73. [Google Scholar]
- Lin, M.; Gong, W. Evaluation of the potential sensitization of chlorogenic acid: A meta-analysis. Evid. Based Complement. Altern. Med. 2013, eCAM 2013, 208467. [Google Scholar] [CrossRef]
- Zeng, J.; He, F. Study of allergenicity of baicalin in Shuanghuanglian Injection. Chin. J. Exp. Tradit. Med Formulae 2012, 18, 102–106. [Google Scholar]
- Yin, J.; Yin, L. Enzyme-linked immunosorbent assay of hapten in Shuanghuanglian Injection. Chin. Tradit. Pat. Med. 2013, 35, 2678–2683. [Google Scholar]
- Wu, J.; Zhang, B. Epidemiological characteristic of anaphylactic shock caused by Shuanghuanglian Injection. Chin. Tradit. Pat. Med. 2007, 29, 638–641. [Google Scholar]
- Li, Q.; Zhao, Y. Chlorogenic acid alters the biological characteristics of basophil granulocytes by affecting the fluidity of the cell membrane and triggering pseudoallergic reactions. Int J. Mol. Med. 2013, 32, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.H.; Zhang, X.Y. Mast cell degranulation induced by chlorogenic acid. Acta Pharmacol. Sin. 2010, 31, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Zhang, T. New method of screening allergenic components from Shuanghuanglian Injection: With RBL-2H3/CMC model online HPLC/MS system. J. Pharm. Biomed. Anal. 2014, 88, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Jing, Z. Screening anaphylactoid components of Shuang Huang Lian Injection by analyzing spectrum-effect relationships coupled with UPLC-TOF-MS. Biomed. Chromatogr. 2019, 33, e4376. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, C. Study on the anaphylactoid of three phenolic acids in Honeysuckle. J. Ethnopharmacol. 2015, 170, 1–7. [Google Scholar] [CrossRef]
- Gao, Y.; Hou, R. Shuang-Huang-Lian injection induces an immediate hypersensitivity reaction via C5a but not IgE. Sci. Rep. 2018, 8, 3572. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Liu, C. Evaluation of anaphylactoid constituents in vitro and in vivo. Int. Immunopharmacol. 2017, 43, 79–84. [Google Scholar] [CrossRef]
- Yi, Y.; Liang, A. Material and mechanisms for evaluation of Shuanghuanglian Injection induced pseudoanaphylactoid reactions. Zhong Guo Zhong Yao Za Zhi 2011, 36, 1865–1869. [Google Scholar]
- Feng, Y.; Jing, Z. Anaphylactoid reactions of three effective components in Shuanghuanglian Injection. Acta Chin. Med. Pharmacol. 2014, 42, 33–36. [Google Scholar]
- Zhang, M.; Jin, J. Analysis of the literature on adverse reactions of Tanreqing injection. China J. Clin. Ration. Drug Use 2019, 12, 13–15. [Google Scholar]
- Wang, L.; Zhang, F. Literature analysis of ADE induced by Tanreqing Injection. China Pharm. 2019, 30, 694–697. [Google Scholar]
- FDA. Guidance for Industry on Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers; Availability. 2005; Federal Register. Available online: https://www.federalregister.gov/documents/2005/07/22/05-14456/guidance-for-industry-on-estimating-the-maximum-safe-starting-dose-in-initial-clinical-trials-for (accessed on 12 December 2019).
- Wang, Y.; Zhao, H. Forsythiaside A exhibits anti-inflammatory effects in LPS-stimulated BV2 microglia cells through activation of Nrf2/HO-1 signaling pathway. Neurochem. Res. 2016, 41, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Zhao, Y. Forsythiaside attenuates lipopolysaccharide-induced inflammatory responses in the bursa of Fabricius of chickens by downregulating the NF-kappaB signaling pathway. Exp. Ther. Med. 2014, 7, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Li, F. Forsythiaside inhibits cigarette smoke-induced lung inflammation by activation of Nrf2 and inhibition of NF-kappaB. Int. Immunopharmacol. 2015, 28, 494–499. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y. Forsythoside A inhibited S. aureus stimulated inflammatory response in primary bovine mammary epithelial cells. Microb. Pathog. 2018, 116, 158–163. [Google Scholar] [CrossRef]
- Qian, J.; Ma, X. Protective effect of forsythiaside A on OVA-induced asthma in mice. Eur. J. Pharmacol. 2017, 812, 250–255. [Google Scholar] [CrossRef]
- Li, H.; Tang, D. Forsythiaside inhibits bacterial adhesion on titanium alloy and attenuates Ti-induced activation of nuclear factor-kappaB signaling-mediated macrophage inflammation. J. Orthop. Surg. Res. 2018, 13, 139. [Google Scholar] [CrossRef]
- Zhang, X.T.; Ding, Y. Forsythoside A modulates zymosan-induced peritonitis in mice. Molecules 2018, 23, 593. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.W.; Zhou, G.Y. Protective effect of forsythiaside A on lipopolysaccharide/d-galactosamine-induced liver injury. Int. Immunopharmacol. 2015, 26, 80–85. [Google Scholar] [CrossRef]
- Zeng, X.Y.; Yuan, W. Forsythoside A exerts an anti-endotoxin effect by blocking the LPS/TLR4 signaling pathway and inhibiting Tregs in vitro. Int. J. Mol. Med. 2017, 40, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wu, J. Forsythoside A inhibits the avian infectious bronchitis virus in cell culture. Phytother. Res. 2011, 25, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Su, H. Forsythoside A exerts antipyretic effect on yeast-induced pyrexia mice via inhibiting transient receptor potential vanilloid 1 function. Int. J. Biol. Sci. 2017, 13, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Yan, Y. Forsythiaside prevents beta-amyloid-induced hippocampal slice injury by upregulating 2-arachidonoylglycerol via cannabinoid receptor 1-dependent NF-kappaB pathway. Neurochem. Int. 2019, 125, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chai, Q. Pharmacokinetics of forsythiaside in rats. World Sci. Technol./Mod. Tradit. Chin. Med. Mater. Med. 2008, 10, 53–56. [Google Scholar]
- Pan, C.; Meng, H. Homology modeling and 3D-QSAR study of benzhydrylpiperazine delta opioid receptor agonists. Comput. Biol. Chem. 2019, 83, 107109. [Google Scholar] [CrossRef] [PubMed]
- Jobichen, C.; Pal, K. Crystal structure of mouse RhoA:GTPγS complex in a centered lattice. J. Struct. Funct. Genom. 2012, 13, 241–245. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.; Zhang, Y.; Pan, C.; Xian, Z.; Pan, C.; Zhao, Y.; Li, C.; Yi, Y.; Wang, L.; Tian, J.; et al. Forsythoside A and Forsythoside B Contribute to Shuanghuanglian Injection-Induced Pseudoallergic Reactions through the RhoA/ROCK Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 6266. https://doi.org/10.3390/ijms20246266
Han J, Zhang Y, Pan C, Xian Z, Pan C, Zhao Y, Li C, Yi Y, Wang L, Tian J, et al. Forsythoside A and Forsythoside B Contribute to Shuanghuanglian Injection-Induced Pseudoallergic Reactions through the RhoA/ROCK Signaling Pathway. International Journal of Molecular Sciences. 2019; 20(24):6266. https://doi.org/10.3390/ijms20246266
Chicago/Turabian StyleHan, Jiayin, Yushi Zhang, Chen Pan, Zhong Xian, Chenling Pan, Yong Zhao, Chunying Li, Yan Yi, Lianmei Wang, Jingzhuo Tian, and et al. 2019. "Forsythoside A and Forsythoside B Contribute to Shuanghuanglian Injection-Induced Pseudoallergic Reactions through the RhoA/ROCK Signaling Pathway" International Journal of Molecular Sciences 20, no. 24: 6266. https://doi.org/10.3390/ijms20246266