Full-Length Transcriptome Sequencing and Different Chemotype Expression Profile Analysis of Genes Related to Monoterpenoid Biosynthesis in Cinnamomum porrectum

 ,
,
Abstract
:1. Introduction
2. Results
2.1. The Composition of Leaf Extracts from Different Chemotypes of C. porrectum
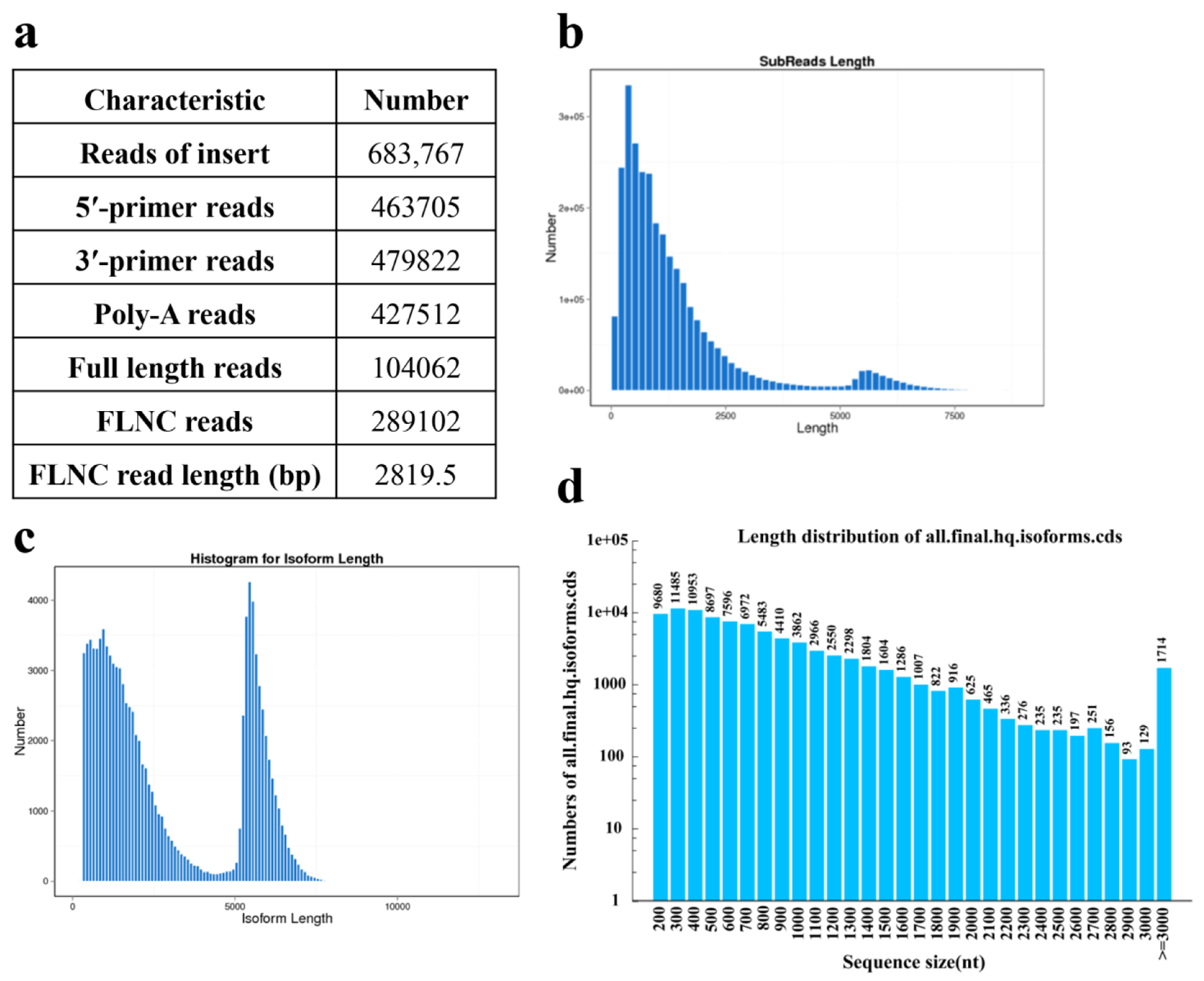
2.2. RNA Sequencing and Transcriptomic Assembly
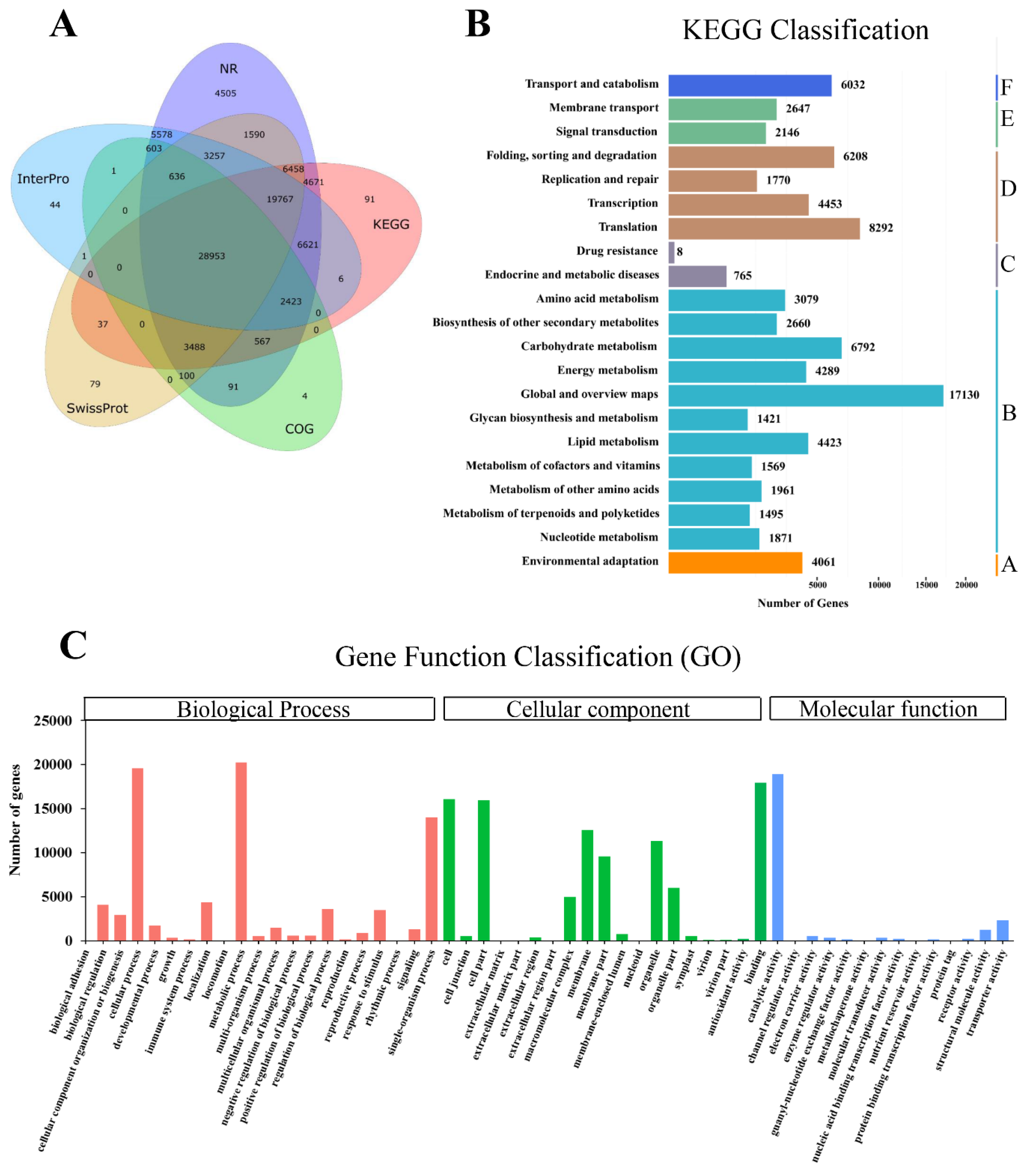
2.3. Gene Annotation and Functional Classification
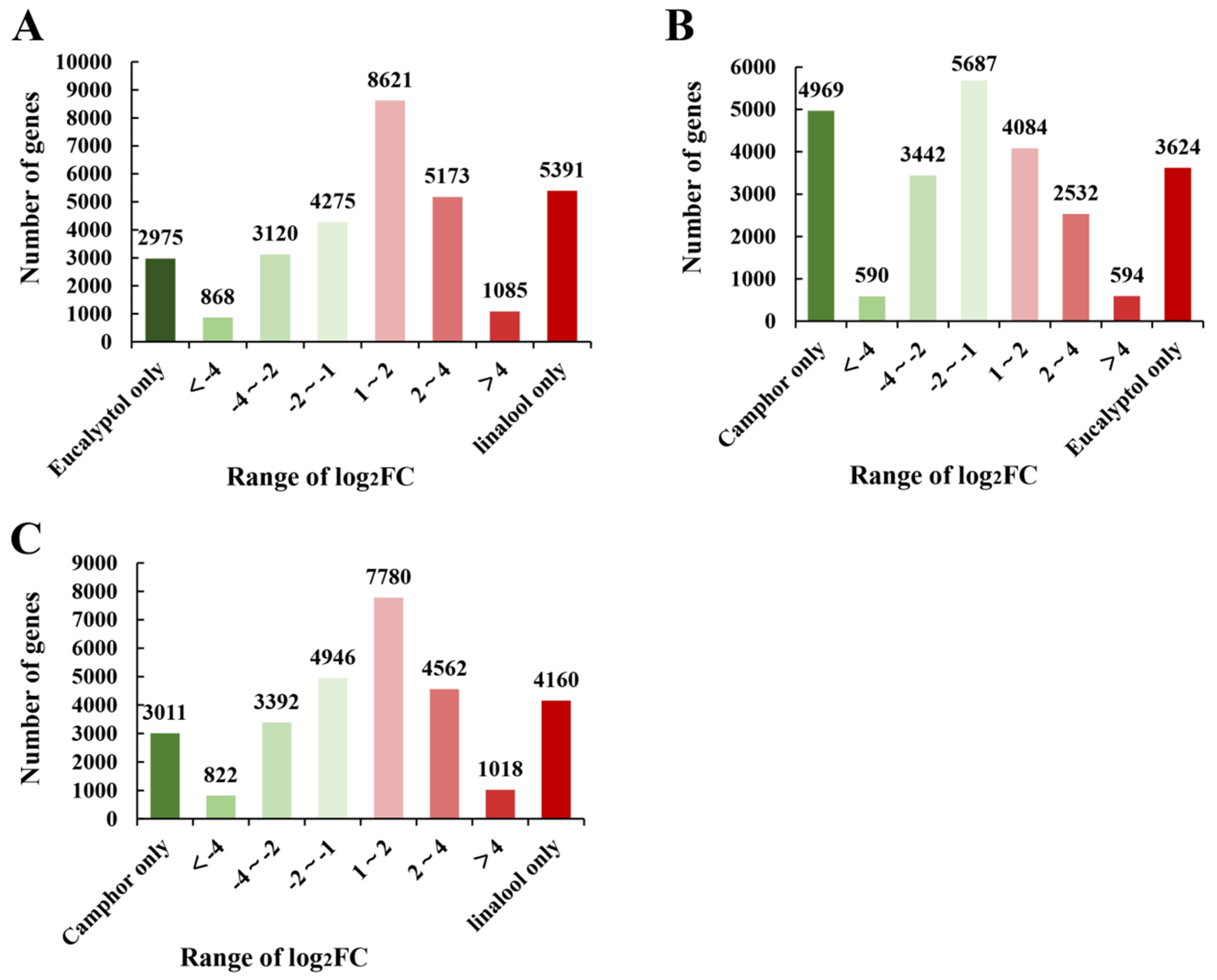
2.4. The Identification of Relative DEGs in C. porrectum Chemotypes and Enrichment Analysis of Transcripts
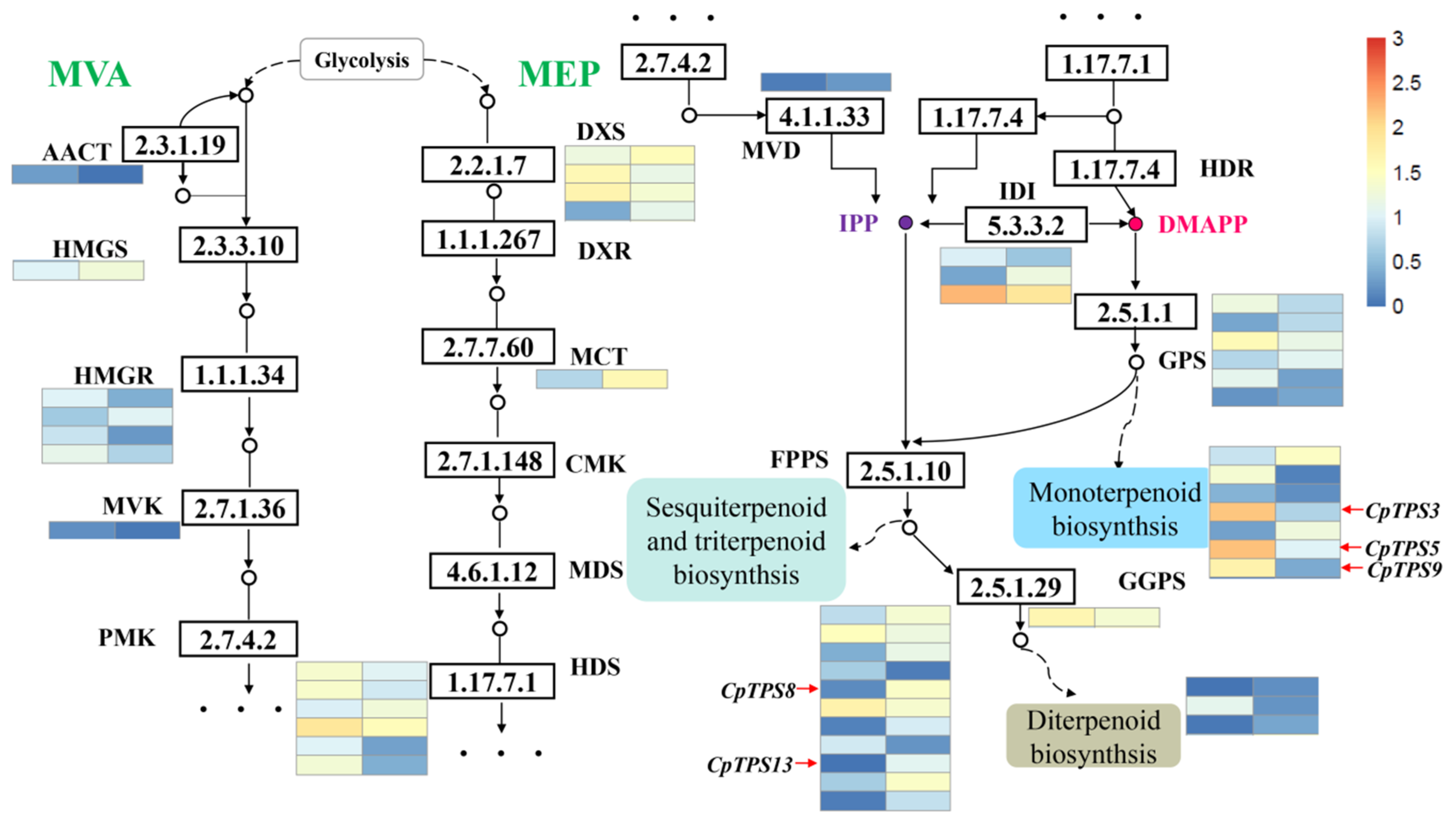
2.5. Candidate Genes Involved in Terpenoid Biosynthesis
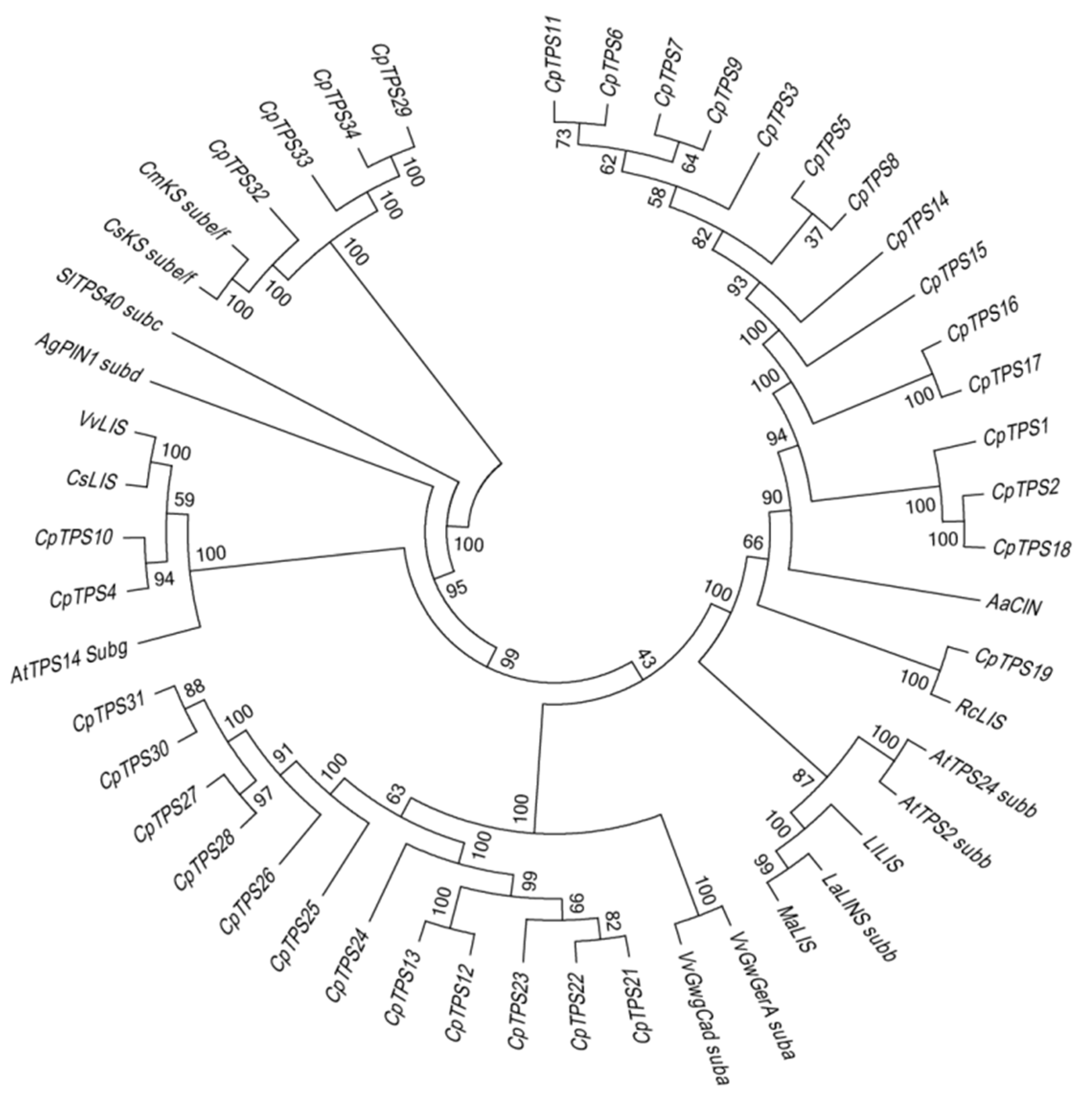
2.6. Construction of Phylogenetic Tree of TPS
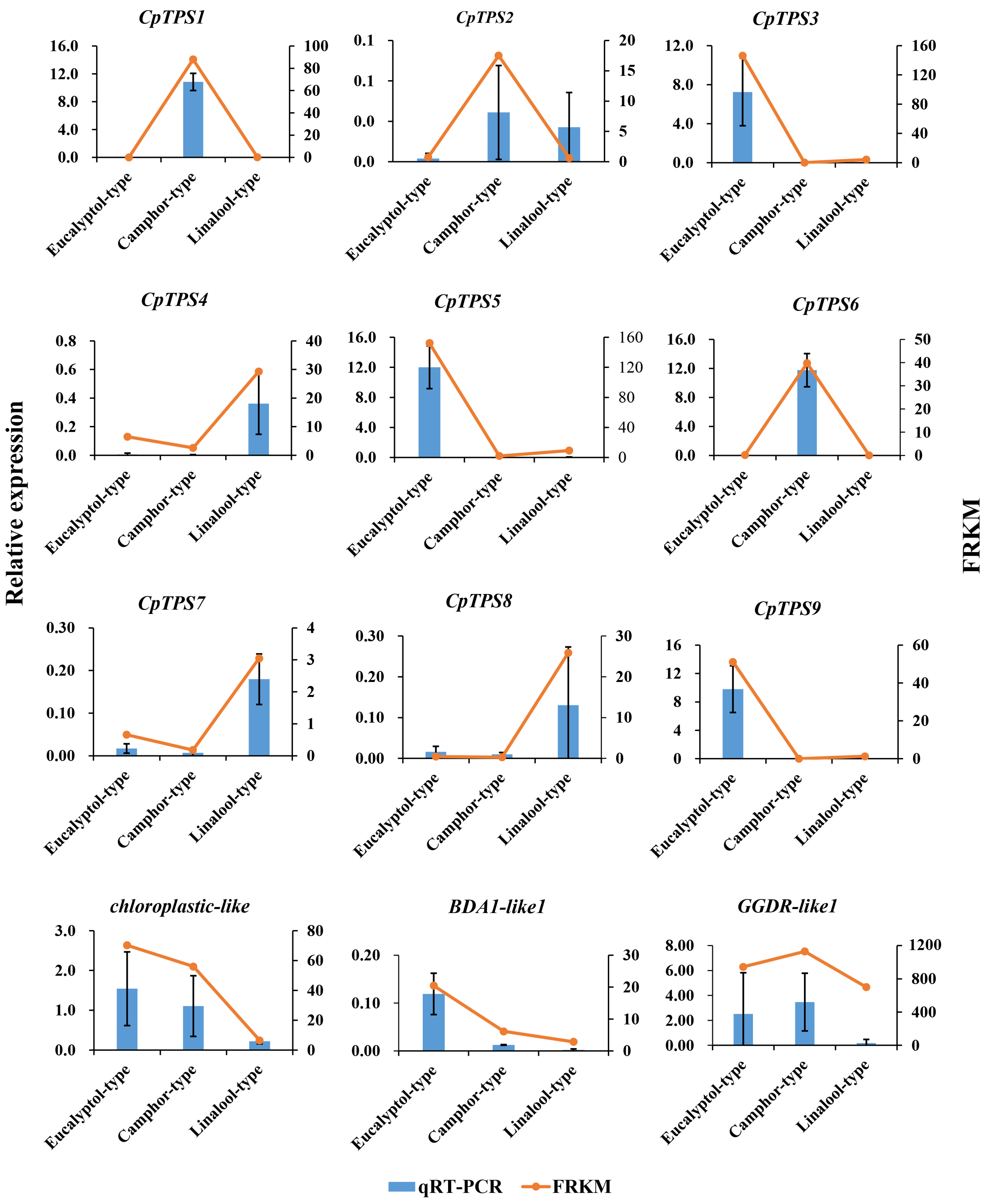
2.7. qRT-PCR Validation of DEGs from the RNA-Seq Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Determination of Chemical Composition
4.3. RNA Extraction and Library Construction
4.4. Transcriptome Sequencing and Assembly
4.5. Determination of the Expression Profiles of Three Chemotypes
4.6. Transcriptome Annotation and Differential Expression Analysis
4.7. Construction of Phylogenetic Tree of TPS
4.8. qRT-PCR Validation of DEGs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| bp | Base pairs (measuring unit) |
| CDS | Coding sequence |
| DEGs | Differentially expressed genes |
| DESeq | Differentially expressed sequence |
| DMAPP | Dimethylallyl diphosphate |
| FDR | False discovery rate |
| FPKM | Fragments per kilobase of exon per million fragments mapped |
| FPPS | Farnesyl diphosphate synthase |
| GC-MS | Gas chromatography-mass spectrometry |
| GGPS | Geranylgeranyl diphosphate synthase |
| GO | Gene ontology |
| GPS | Geranyl diphosphate synthase |
| IPP | Isopentenyl diphosphate |
| KEGG | Kyoto encyclopedia of genes and genomes |
| KO | KEGG ortholog database |
| KOG | EuKaryotic Orthologous Groups |
| MEP | 2-C-methyl-D-erythritol 4phosphate |
| MVA | Mevalonate acid |
| NIST | National institute of standards and technology |
| Nr | NCBI non-redundant protein sequences |
| Nt | NCBI non-redundant nucleotide sequences |
| Pfam | Protein family |
| qRT-PCR | Quantitative reverse transcription PCR |
| ROI | Reads of insert |
| RNA-Seq | RNA sequencing |
| SMRT | Single molecule real-time |
| TP | Terpene synthases |
References
- Li, S.; Li, X.; Li, J.; Huang, P.; Wei, F.; Cui, H.; Werff, H. Flora of China; Science Press and St. Louis: Missouri Botanical Garden Press: Beijing, China, 2008; Volume 7, p. 102. [Google Scholar]
- Wu, H.; Wang, J.J.; Liu, C.; Zhu, L.F. A study on the chemotypes of Cinnamomum parthenoxylen(Jack) Nees. J. Plant Resour. Environ. 1992, 1, 45–49. (In Chinese) [Google Scholar]
- Kawamura, F.; Ramle, S.F.M.; Sulaiman, O.; Hashim, R.; Ohara, S. Antioxidant and antifungal activities of extracts from 15 selected hardwood species of Malaysian timber. Eur. J. Wood Prod. 2010, 69, 207–212. [Google Scholar] [CrossRef]
- Buru, A.S.; Pichika, M.R.; Neela, V.; Mohandas, K. In vitro antibacterial effects of Cinnamomum extracts on common bacteria found in wound infections with emphasis on methicillin-resistant Staphylococcus aureus. J. Ethnopharmacol. 2014, 153, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Phongpaichit, S.; Kummee, S.; Nilrat, L.; Itarat, A. Antimicrobial activity of oil from the root of Cinnamomum porrectum. Songklanakarin J. Sci. Technol. 2006, 29, 11–16. [Google Scholar]
- Palanuvej, C.; Werawatganone, P.; Lipipun, V.; Ruangrungsi, N. Chemical composition and antimicrobial activity against Candida albicans of essential oil from leaves of Cinnamomum porrectum. J. Health Res. 2006, 20, 69–76. [Google Scholar]
- Subki, S.Y.M.; Jamal, J.A.; Husain, K.; Manshoor, N. Characterisation of leaf essential oils of three Cinnamomum species from Malaysia by gas chromatography and multivariate data analysis. Pharmacogn. J. 2013, 5, 22–29. [Google Scholar] [CrossRef]
- Batista, P.A.; Werner, M.F.; Oliveira, E.C.; Burgos, L.; Pereira, P.; Brum, L.F.; Santos, A.R. Evidence for the involvement of ionotropic glutamatergic receptors on the antinociceptive effect of (-)-linalool in mice. Neurosci. Lett. 2008, 440, 299–303. [Google Scholar] [CrossRef]
- Huo, M.X.; Cui, X.R.; Xue, J.D.; Chi, G.F.; Gao, R.J.; Deng, X.M.; Guan, S.; Wei, J.Y.; Soromou, L.W.; Feng, H.H.; et al. Anti-inflammatory effects of linalool in RAW 264.7 macrophages and lipopolysaccharide-induced lung injury model. J. Surg. Res. 2013, 180, e47–e54. [Google Scholar] [CrossRef]
- Miyashita, M.; Sadzuka, Y. Effect of linalool as a component of Humulus lupulus on doxorubicin-induced antitumor activity. Food Chem. Toxicol. 2013, 53, 174–179. [Google Scholar] [CrossRef]
- Kladniew, R.B.; Polo, M.; Villegas, S.M.; Galle, M.; Crespo, R.; Garcia de Bravo, M. Synergistic antiproliferative and anticholesterogenic effects of linalool, 1,8-cineole, and simvastatin on human cell lines. Chem. Biol. Interact. 2014, 214, 57–68. [Google Scholar] [CrossRef]
- Shi, C.; Sun, Y.; Liu, Z.Y.; Guo, D.; Sun, H.H.; Sun, Z.; Chen, S.; Zhang, W.T.; Wen, Q.W.; Peng, X.L.; et al. Inhibition of cronobacter sakazakii virulence factors by citral. Sci. Rep. 2017, 7, 43243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.H.; Zheng, Y.J.; Zhong, Y.D.; Wu, Y.F.; Li, Z.T.; Xu, L.A.; Xu, M. Transcriptome analysis and identification of genes related to terpenoid biosynthesis in Cinnamomum camphora. BMC Genom. 2018, 19, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63. [Google Scholar] [PubMed]
- Xu, Y.; Zhu, C.; Xu, C.; Sun, J.; Grierson, D. Integration of metabolite profiling and transcriptome analysis reveals genes related to volatile terpenoid metabolism in finger citron (C. medica var. sarcodactylis). Molecules 2019, 24, 2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordoba, E.; Salmi, M.; Leon, P. Unravelling the regulatory mechanisms that modulate the MEP pathway in higher plants. J. Exp. Bot. 2009, 60, 2933–2943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler; Hartmut, K. The 1-Deoxy-D-Xylulose-5-Phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Biol. 1999, 50, 47–65. [Google Scholar] [CrossRef] [PubMed]
- Sapir-Mir, M.; Mett, A.; Belausov, E.; Tal-Meshulam, S.; Frydman, A.; Gidoni, D.; Eyal, Y. Peroxisomal localization of arabidopsis isopentenyl diphosphate isomerases suggests that part of the plant isoprenoid mevalonic acid pathway is compartmentalized to peroxisomes. Plant Physiol. 2008, 148, 1219–1228. [Google Scholar] [CrossRef] [Green Version]
- Rohmer, M. The discovery of a mevalonate-independent pathway for isoprenoid biosynthesis in bacteria, algae and higher plants. Nat. Prod. Rep. 1999, 16, 565–574. [Google Scholar] [CrossRef]
- Kazunori, O.; Hiroyuki, K.; Shinjiro, Y.; Hiroshi, K.; Yuji, K.; Hideaki, N.; Hisakazu, Y. Genetic evidence for the role of isopentenyl diphosphate isomerases in the mevalonate pathway and plant development in Arabidopsis. Plant Cell Physiol. 2008, 49, 604. [Google Scholar]
- Moses, T.; Pollier, J.; Thevelein, J.M.; Goossens, A. Bioengineering of plant (tri)terpenoids: From metabolic engineering of plants to synthetic biology in vivo and in vitro. New Phytol. 2013, 200, 27–43. [Google Scholar] [CrossRef]
- Wise, M.L.; Savage, T.J.; Katahira, E.; Croteau, R. Monoterpene Synthases from Common Sage (Salvia officinalis). J. Biol. Chem. 1998, 273, 14891–14899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohlmann, J.; Phillips, M.; Ramachandiran, V.; Katoh, S.; Croteau, R. cDNA Cloning, Characterization, and Functional Expression of Four New Monoterpene Synthase Members of the Tpsd Gene Family from Grand Fir (Abies grandis). Arch. Biochem. Biophys. 1999, 368, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Colby, S.M.; Alonso, W.R.; Katahira, E.J.; McGarvey, D.J.; Croteau, R. 4S-limonene synthase from the oil glands of spearmint (Mentha spicata). cDNA isolation, characterization, and bacterial expression of the catalytically active monoterpene cyclase. J. Biol. Chem. 1993, 268, 23016–23024. [Google Scholar] [PubMed]
- Williams, D.C.; McGarvey, D.J.; Katahira, E.J.; Croteau, R. Truncation of limonene synthase preprotein provides a fully active ‘Pseudomature’ form of this monoterpene cyclase and reveals the function of the amino-terminal arginine pair. Biochemistry 1998, 37, 12213–12220. [Google Scholar] [CrossRef]
- Lin, Y.L.; Lee, Y.R.; Huang, W.K.; Chang, S.T.; Chu, F.H. Characterization of S-(+)-linalool synthase from several provenances of Cinnamomum osmophloeum. Tree Genet. Genomes 2014, 10, 75–86. [Google Scholar] [CrossRef]
- Despinasse, Y.; Fiorucci, S.; Antonczak, S.; Moja, S.; Bony, A.; Nicole, F.; Baudino, S.; Maqnard, J.L.; Jullien, F. Bornyl-diphosphate synthase from Lavandula angustifolia: A major monoterpene synthase involved in essential oil quality. Phytochemistry 2017, 137, 24–33. [Google Scholar] [CrossRef]
- Yahyaa, M.; Matsuba, Y.; Brandt, W.; Doron-Faigenboim, A.; Bar, E.; McClain, A.; Davidovich-Rikanati, R.; Lewinsohn, E.; Pichersky, E.; Ibdah, M.; et al. Identification, Functional Characterization, and Evolution of Terpene Synthases from a Basal Dicot. Plant Physiol. 2015, 169, 1683–1697. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Li, J.; Wang, H.X.; Zeng, Y.A. Geraniol-synthase gene from Cinnamomum tenuipilum. Phytochemistry 2005, 66, 285–293. [Google Scholar] [CrossRef]
- Hittinger, C.T.; Johnston, M.; Tossberg, J.T.; Rokas, A. Leveraging skewed transcript abundance by RNA-Seq to increase the genomic depth of the tree of life. Proc. Natl. Acad. Sci. USA 2010, 107, 1476–1481. [Google Scholar] [CrossRef] [Green Version]
- Morozova, O.; Hirst, M.; Marra, M.A. Applications of new sequencing technologies for transcriptome analysis. Annu. Rev. Genom. Hum. Genet. 2009, 10, 135–151. [Google Scholar] [CrossRef] [Green Version]
- Rhoads, A.; Au, K.F. PacBio sequencing and its applications. Genom. Proteom. Bioinf. 2015, 13, 278–289. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Ghany, S.E.; Hamilton, M.; Jacobi, J.L.; Ngam, P.; Devitt, N.; Schilkey, F.; Ben-Hur, A.; Reddy, A.S. A survey of the sorghum transcriptome using single-molecule long reads. Nat. Commun. 2016, 7, 11706. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Tseng, E.; Regulski, M.; Clark, T.A.; Hon, T.; Jiao, Y.; Lu, Z.; Olson, A.; Stein, J.C.; Ware, D. Unveiling the complexity of the maize transcriptome by single-molecule long-read sequencing. Nat. Commun. 2016, 24, 11708. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, P.; Liang, F.; Ye, Z.; Li, J.; Shen, C.; Pei, L.; Wang, F.; Hu, J.; Tu, L.; et al. A global survey of alternative splicing in allopolyploid cotton: Landscape, complexity and regulation. New Phytol. 2018, 217, 163–178. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, S.; Shi, X.; Liu, D.; Zhao, P.; Lu, Y.; Cheng, Y.; Liu, Z.; Nie, X.; Song, W. Hybrid sequencing reveals insight into heat sensing and signaling of bread wheat. Plant J. 2019, 98, 10151032. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscope, 4th ed.; Allured Publishing Corporation: Carol Steam, IL, USA, 2007; pp. 1–804. [Google Scholar]
- Bordiga, M.; Rinaldi, M.; Locatelli, M.; Piana, G.; Travaglia, F.; CoÏsson, J.D.; Arlorio, M. Characterization of Muscat wines aroma evolution using comprehensive gas chromatography followed by a post-analytic approach to 2D contour plots comparison. Food Chem. 2013, 140, 57–67. [Google Scholar] [CrossRef]
- Ribeiro, V.L.S.; Rolim, V.; Bordignon, S.; Henriques, A.T.; Dorneles, G.G.; Limberger, R.P.; Poster, G.V. Chemical composition and larvicidal properties of the essential oils from Drimys brasiliensis Miers (Winteraceae) on the cattle tick Rhipicephalus (Boophilus) microplus and the brown dog tick Rhipicephalus sanguineus. Parasitol. Res. 2008, 102, 531–535. [Google Scholar] [CrossRef]
- Andrade, E.H.A.; Zoghbi, M.D.G.B.; Lima, M.D.P.; Botânica, C.D.B.; Goeldi, M.P.E. Chemical composition of the essential oils of Cymbopogon citratus (DC.) Stapf cultivated in north of Brazil. J. Essent. Oil Bear. Plants 2009, 12, 41–45. [Google Scholar] [CrossRef]
- Darbani, B.; Stewart, C.N., Jr.; Noeparvar, S.; Borg, S. Correction of gene expression data: Performance-dependency on inter-replicate and inter-treatment biases. J. Biotechnol. 2014, 188, 100–109. [Google Scholar] [CrossRef]
- He, Y.; Deng, C.; Xiong, L.; Qin, S.S.; Peng, C. Transcriptome sequencing provides insights into the metabolic pathways of patchouli alcohol and pogostone in Pogostemon cablin (Blanco) Benth. Genes Genom. 2016, 38, 1031–1039. [Google Scholar] [CrossRef]
- Tilgner, H.; Jahanbani, F.; Blauwkamp, T.; Moshrefi, A.; Jaeger, E.; Chen, F.; Harel, I.; Bustamante, C.D.; Rasmussen, M.; Snyder, M.P. Comprehensive transcriptome analysis using synthetic long-read sequencing reveals molecular co-association of distant splicing events. Nat. Biotechnol. 2015, 33, 736–742. [Google Scholar] [CrossRef]
- Chang, Y.T.; Chu, F.H. Molecular cloning and characterization of monoterpene synthases from Litsea cubeba (Lour.) Persoon. Tree Genet. Genomes 2011, 7, 835–844. [Google Scholar] [CrossRef]
- Vranová, E.; Coman, D.; Gruissem, W. Network Analysis of the MVA and MEP Pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef] [PubMed]
- Friesen, J.A.; Rodwell, V.W. The 3-hydroxy-3-methylglutaryl coenzyme-A (HMG-CoA) reductases. Genome Biol. 2004, 5, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohdich, F.; Lauw, S.; Kaiser, J.; Feicht, R.P.; Bacher, A.; Eisenreich, W. Isoprenoid biosynthesis in plants-2 C-methyl-d-erythritol-4-phosphate synthase (IspC protein) of Arabidopsis thaliana. FEBS J. 2006, 273, 4446. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.S.; Lin, C.Y.; Yang, C.K.; Chen, Y.J.; Chung, M.J.; Chang, S.T. Chemical polymorphism and composition of leaf essential oils of Cinnamomum kanehirae using Gas Chromatography/Mass Spectrometry, cluster analysis, and principal component analysis. J. Wood Chem. Technol. 2015, 35, 207–219. [Google Scholar] [CrossRef]
- Zulak, K.G.; Lippert, D.N.; Kuzyk, M.A.; Domanski, D.; Chou, T.; Borchers, C.H.; Bohlmann, J. Targeted proteomics using selected reaction monitoring reveals the induction of specific terpene synthases in a multi-level study of methyl jasmonate-treated Norway spruce (Picea abies). Plant J. 2009, 60, 1015–1030. [Google Scholar] [CrossRef]
- Dudareva, N. (E)-beta-Ocimene and Myrcene synthase genes of floral scent biosynthesis in snapdragon: Function and expression of three terpene synthase genes of a new terpene synthase subfamily. Plant Cell. 2003, 15, 1227–1241. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.M.; Faldt, J.; Bohlmann, J. Functional characterization of nine Norway spruce TPS genes and evolution of gymnosperm terpene synthases of the TPS-d subfamily. Plant Physiol. 2004, 135, 1908–1927. [Google Scholar] [CrossRef] [Green Version]
- Naik, D.G.; Dandge, C.N.; Rupanar, S.V. Chemical examination and evaluation of antioxidant and antimicrobial activities of essential oil from Gymnema sylvestre R. Br. Leaves. J. Essent. Oil Res. 2011, 23, 12–19. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Cantrell, C.L.; Tekwani, B.; Khan, S.I. Content, composition, and bioactivity of the essential oils of three basil genotypes as a function of harvesting. J. Agric. Food Chem. 2008, 56, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; Mccue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götz, S.; Garcíagómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, SH.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RIa | RIb | Component | Y_L1 (%) | Y_L2 (%) | Y_L3 (%) | F_L1 (%) | F_L2 (%) | F_L3 (%) | N_L1 (%) | N_L2 (%) | N_L3 (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Monoterpenes | 92.98 | 93.65 | 90.04 | 89.46 | 88.04 | 88.33 | 99.01 | 92.47 | 98.32 | ||
| 929 | 928 | 3-Thujene | 0.51 | 0.51 | 0.28 | - | - | - | - | - | - |
| 939 | 935 | α-Pinene | 3.16 | 3.47 | 2 | - | - | - | - | - | - |
| 977 | 1006 | β-Phellandrene | 12.16 | 9.9 | 9.53 | - | - | - | - | - | - |
| 986 | 978 | β-Pinene | 3.47 | 3.41 | 2.55 | - | - | - | - | - | - |
| 989 | 988 | Myrcene | 1.78 | 1.99 | 1.76 | - | - | - | - | - | - |
| 1011 | 1002 | α-Phellandrene | 0.37 | 0.46 | 0.16 | - | - | - | - | - | - |
| 1021 | 1014 | α-Terpinene | 1.5 | 2.39 | 1.57 | - | - | - | - | - | - |
| 1033 | 1031 | D-Limonene | - | - | - | tr | 0.2 | 0.67 | - | - | - |
| 1037 | 1037 | Eucalyptol | 35.63 | 34.67 | 34.48 | 0.1 | 0.23 | 0.59 | tr | tr | tr |
| 1061 | 1058 | γ-Terpinene | 4.21 | 5.16 | 4.15 | tr | - | - | - | - | - |
| 1104 | 1102 | Linalool | 1.36 | 0.81 | 1.64 | 87.88 | 86.46 | 77.58 | tr | tr | tr |
| 1108 | 1108 | cis-β-Terpineol | 0.57 | 0.44 | 0.54 | - | - | - | - | - | - |
| 1148 | 1139 | Pinocarveol | 1.93 | 1.92 | 2.09 | - | - | - | - | - | - |
| 1154 | 1150 | Camphor | - | - | 0.1 | 1.13 | tr | 7.85 | 97.78 | 92.47 | 97.04 |
| 1178 | 1172 | Borneol | 0.25 | 0.71 | 0.57 | tr | 0.55 | 0.37 | 1.23 | - | 1.28 |
| 1185 | 1176 | 4-Terpineol | 6.18 | 8.53 | 7.44 | tr | tr | 0.23 | - | - | - |
| 1198 | 1193 | α-Terpineol | 19.75 | 19.15 | 20.96 | 0.35 | 0.6 | 0.89 | - | - | - |
| 1213 | 1207 | Piperitol | 0.16 | 0.15 | 0.16 | - | - | - | - | - | - |
| 1226 | 1223 | β-Citronellol | 0.32 | 0.35 | 0.34 | - | - | - | - | - | - |
| 1318 | 1318 | Pinanediol | 0.18 | 0.14 | - | - | tr | 0.15 | - | - | - |
| Sesquiterpenes | 5.64 | 4.83 | 9.27 | 8.41 | 10.45 | 8.44 | 0.00 | 3.64 | 0.00 | ||
| 1430 | 1418 | β-caryophyllene | 2.29 | 4.17 | 3.41 | 4.92 | 2.33 | 1.62 | - | - | - |
| 1466 | 1454 | α-caryophyllene | 0.35 | 0.5 | 0.4 | 0.69 | 0.92 | 0.63 | - | - | - |
| 1471 | 1461 | Alloaromadendrene | - | - | 0.19 | 0.13 | 0.21 | - | - | - | - |
| 1491 | 1484 | Germacrene D | 2.39 | - | 1.2 | 0.44 | 3.82 | 1.09 | - | - | - |
| 1505 | 1503 | β-Chamigrene | - | 0.16 | 0.25 | 0.11 | 2.78 | 1.06 | - | - | - |
| 1562 | 1563 | trans-Nerolidol | 0.61 | - | 3.82 | 1.58 | 0.29 | 2.25 | - | - | - |
| 1588 | 1577 | (-)-Spathulenol | - | - | - | tr | tr | 0.3 | - | - | - |
| 1594 | 1596 | Caryophyllene oxide | - | - | - | 0.17 | - | - | - | 1.91 | - |
| 1665 | 1653 | α-Eudesmol | - | - | - | - | 0.1 | 1.24 | - | - | - |
| 1786 | 1762 | Aristolone | - | - | - | 0.37 | 0.25 | - | - | - | |
| Phenols | 0.19 | 0.00 | 0.00 | 0.00 | 0.35 | 0.18 | 0.00 | 0.00 | 0.00 | ||
| 1398 | 1404 | Methyleugenol | 0.19 | - | - | tr | 0.2 | 0.18 | - | - | - |
| 1495 | 1491 | Methylisoeugenol | - | - | - | tr | 0.15 | - | - | - | - |
| Total % | 98.81 | 98.48 | 99.31 | 97.87 | 98.84 | 96.95 | 99.01 | 94.38 | 98.32 | ||
| Oil yield % | 1.50 | 1.65 | 1.15 | 1.90 | 2.14 | 1.86 | 0.61 | 0.78 | 0.96 | ||
| Sample | Total Isoforms | Total Base (bp) | Mean Length (bp) | N50 (bp) |
|---|---|---|---|---|
| Total | 104,062 | 306,337,921 | 2944 | 5449 |
| Software | Total Number | Total Length | Mean Length | N50 | N70 | N90 | GC (%) |
|---|---|---|---|---|---|---|---|
| Blast | 88763 | 66645204 | 750 | 1041 | 687 | 363 | 44.66 |
| ESTScan | 340 | 205098 | 603 | 729 | 501 | 336 | 43.75 |
| Overall | 89103 | 66850302 | 750 | 1038 | 687 | 363 | 44.66 |
| Sample | Total Raw Reads (M) | Total Clean Reads (M) | Total Clean Bases (Gb) | Clean Reads Q20 (%) | Clean Reads Q30 (%) | Total Mapping (%) |
|---|---|---|---|---|---|---|
| AYYS1 | 45.72 | 42.54 | 6.38 | 98.79 | 96.38 | 80.41 |
| AYYS2 | 45.72 | 42.45 | 6.37 | 98.82 | 96.46 | 79.94 |
| AYYS3 | 45.72 | 42.36 | 6.35 | 98.78 | 96.37 | 81.93 |
| NZ1 | 47.35 | 43.01 | 6.45 | 98.68 | 96.07 | 78.49 |
| NZ2 | 47.35 | 43.24 | 6.49 | 98.73 | 96.2 | 81.5 |
| NZ3 | 47.35 | 42.95 | 6.44 | 98.63 | 95.93 | 78.7 |
| YXFZC1 | 47.35 | 42.51 | 6.38 | 98.27 | 94.78 | 76.74 |
| YXFZC2 | 47.35 | 42.5 | 6.38 | 98.29 | 94.82 | 73.63 |
| YXFZC3 | 47.35 | 42.75 | 6.41 | 98.31 | 94.9 | 74.33 |
| summary | 421.26 | 384.31 | 57.65 | / | / | / |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, F.; Wang, X.; Zheng, Y.; Wang, H.; Liu, X.; Su, X. Full-Length Transcriptome Sequencing and Different Chemotype Expression Profile Analysis of Genes Related to Monoterpenoid Biosynthesis in Cinnamomum porrectum. Int. J. Mol. Sci. 2019, 20, 6230. https://doi.org/10.3390/ijms20246230
Qiu F, Wang X, Zheng Y, Wang H, Liu X, Su X. Full-Length Transcriptome Sequencing and Different Chemotype Expression Profile Analysis of Genes Related to Monoterpenoid Biosynthesis in Cinnamomum porrectum. International Journal of Molecular Sciences. 2019; 20(24):6230. https://doi.org/10.3390/ijms20246230
Chicago/Turabian StyleQiu, Fengying, Xindong Wang, Yongjie Zheng, Hongming Wang, Xinliang Liu, and Xiaohua Su. 2019. "Full-Length Transcriptome Sequencing and Different Chemotype Expression Profile Analysis of Genes Related to Monoterpenoid Biosynthesis in Cinnamomum porrectum" International Journal of Molecular Sciences 20, no. 24: 6230. https://doi.org/10.3390/ijms20246230