Interruption of Jasmonic Acid Biosynthesis Causes Differential Responses in the Roots and Shoots of Maize Seedlings against Salt Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
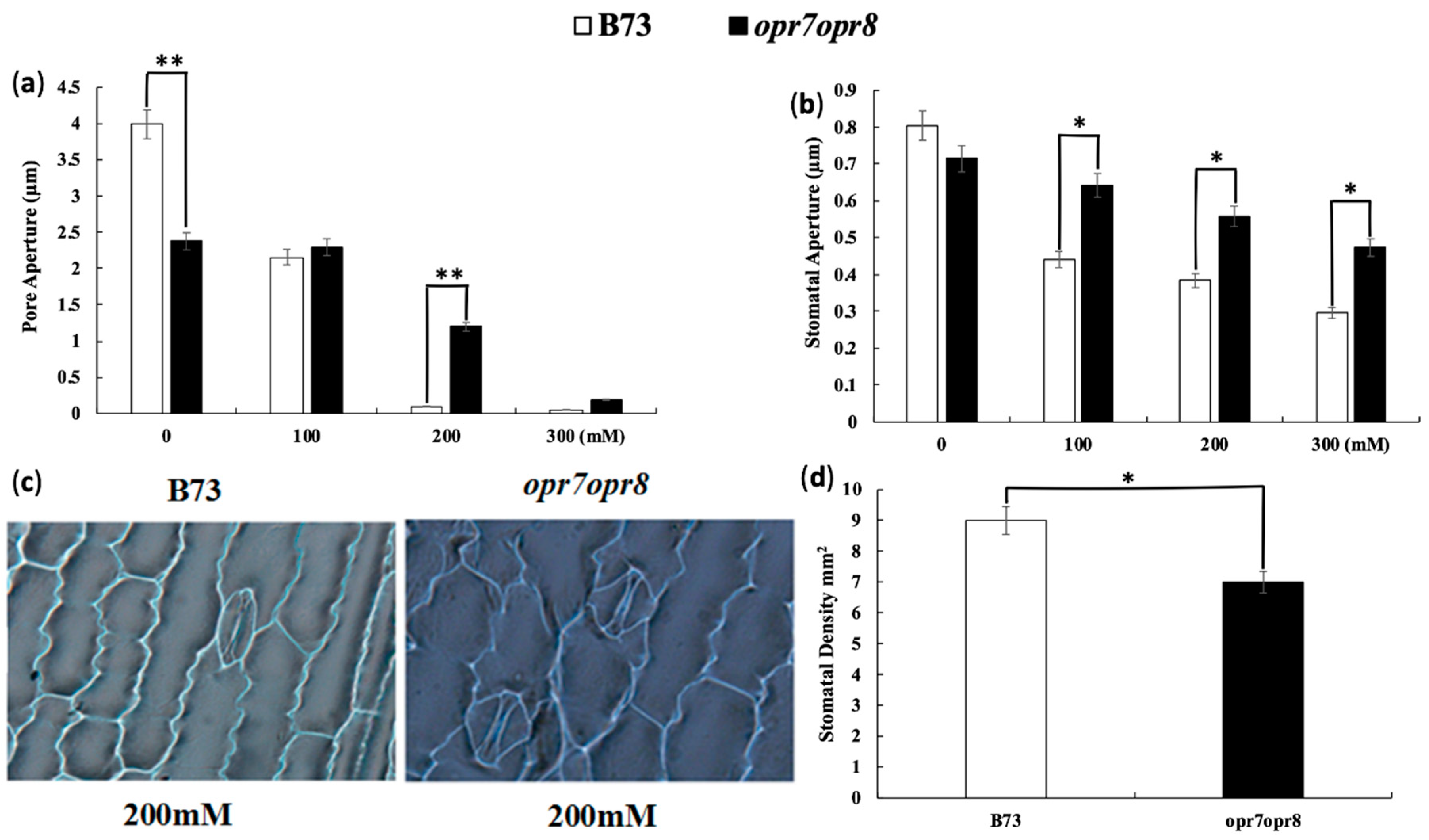
2.1. Jasmonate Is a Required Signal for Stomata Closure under Salt Stress
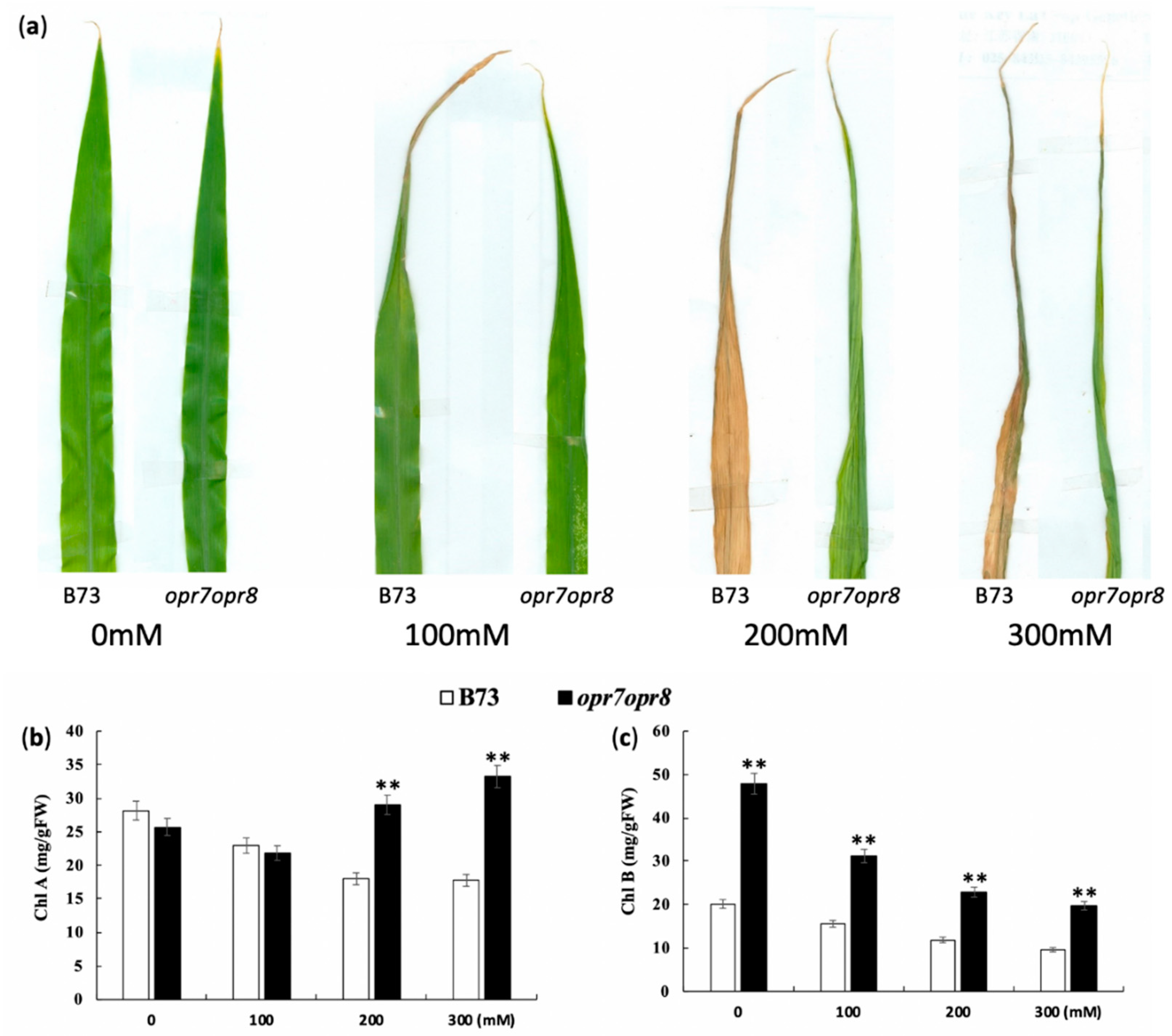
2.2. opr7opr8 Mutant Showed Delayed Leaf Senescence under Salt Stress
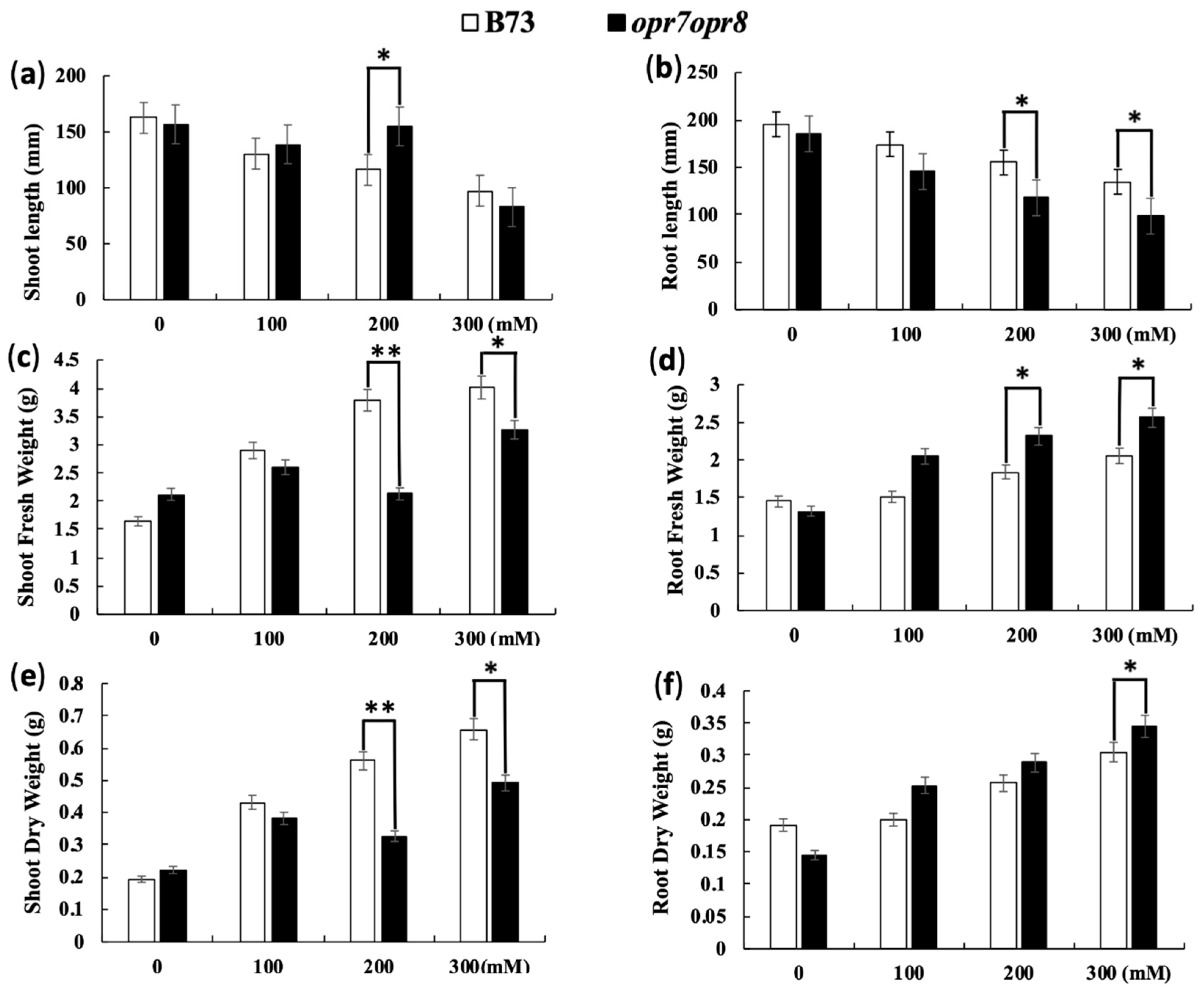
2.3. opr7op8 Displayed Better Growth in the Shoots but Worse Growth in the Roots Than B73 under Salt Stress
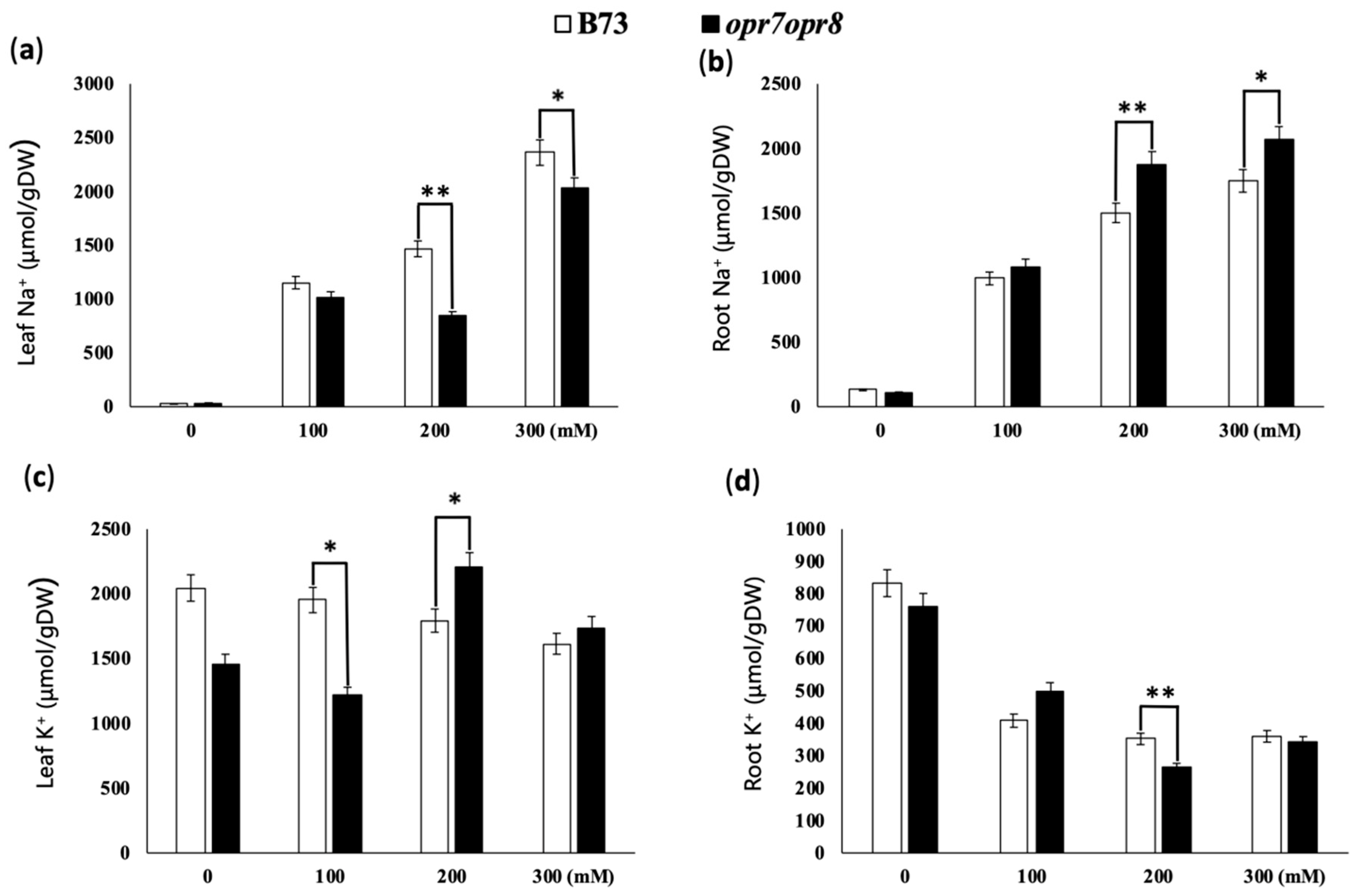
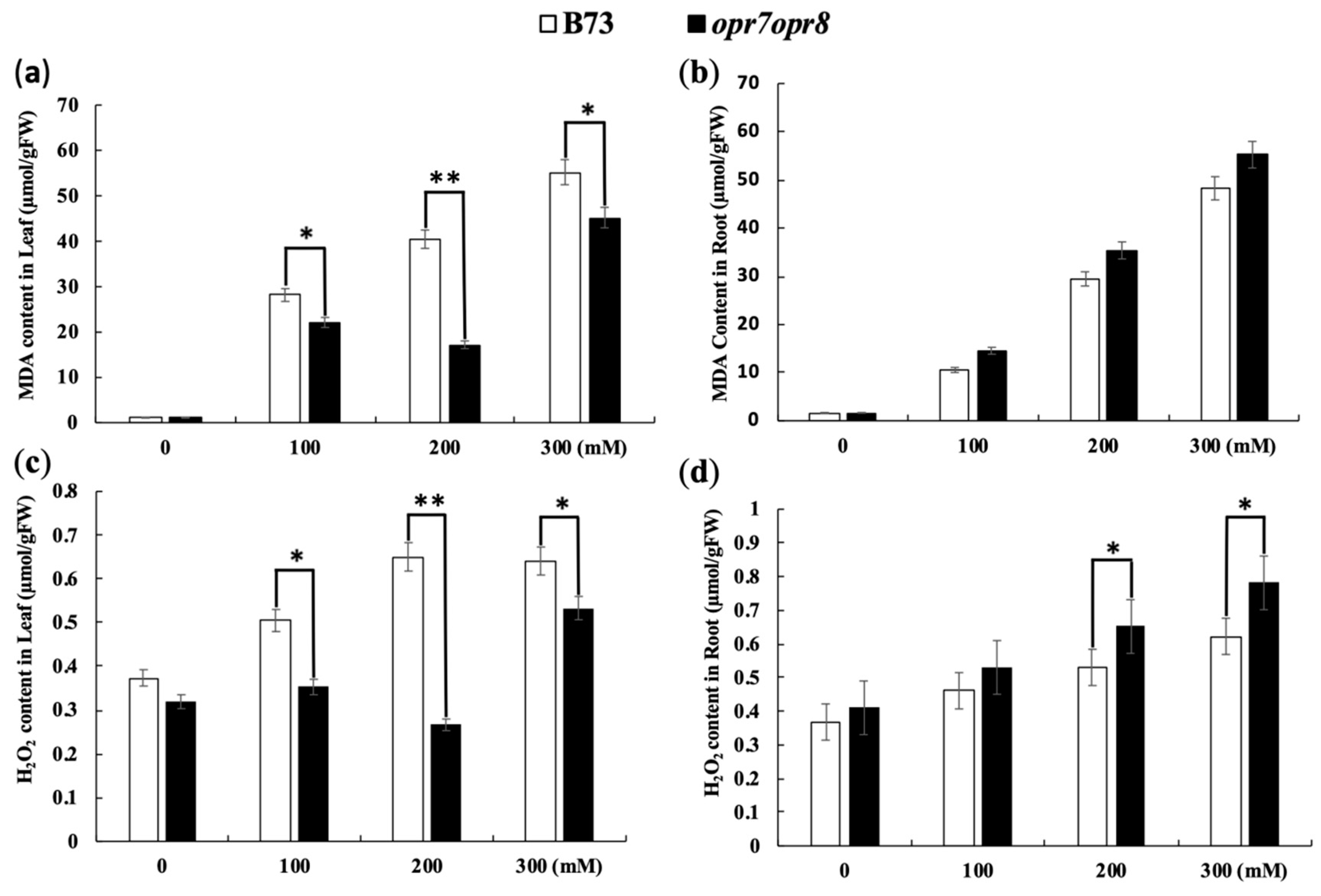
2.4. opr7opr8 Accumulated Less Sodium in the Leaves but More Sodium in the Roots Than WT under Salt Stress
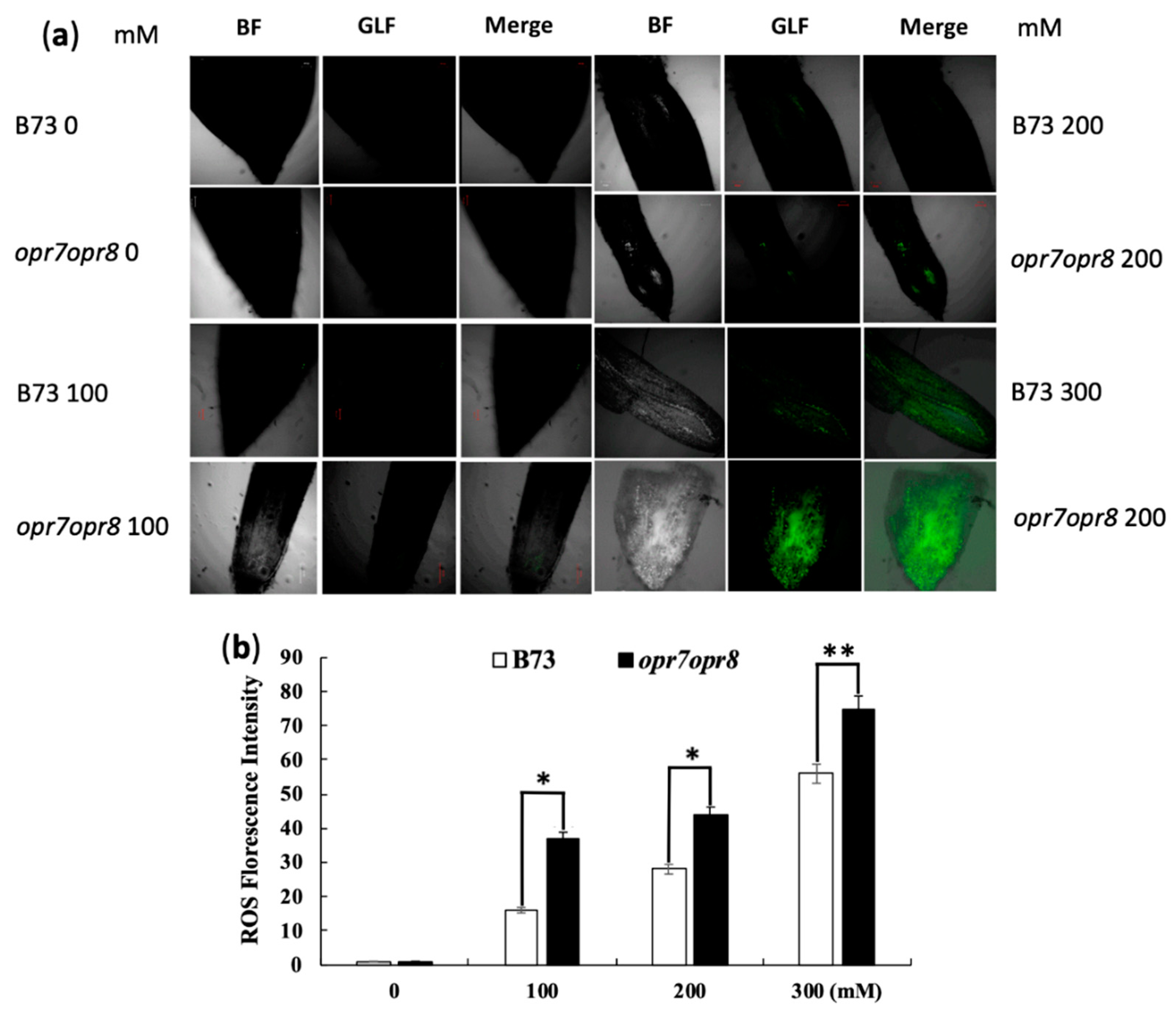
2.5. opr7opr8 and WT Accumulated a Different Level of ROS under Salt Stress
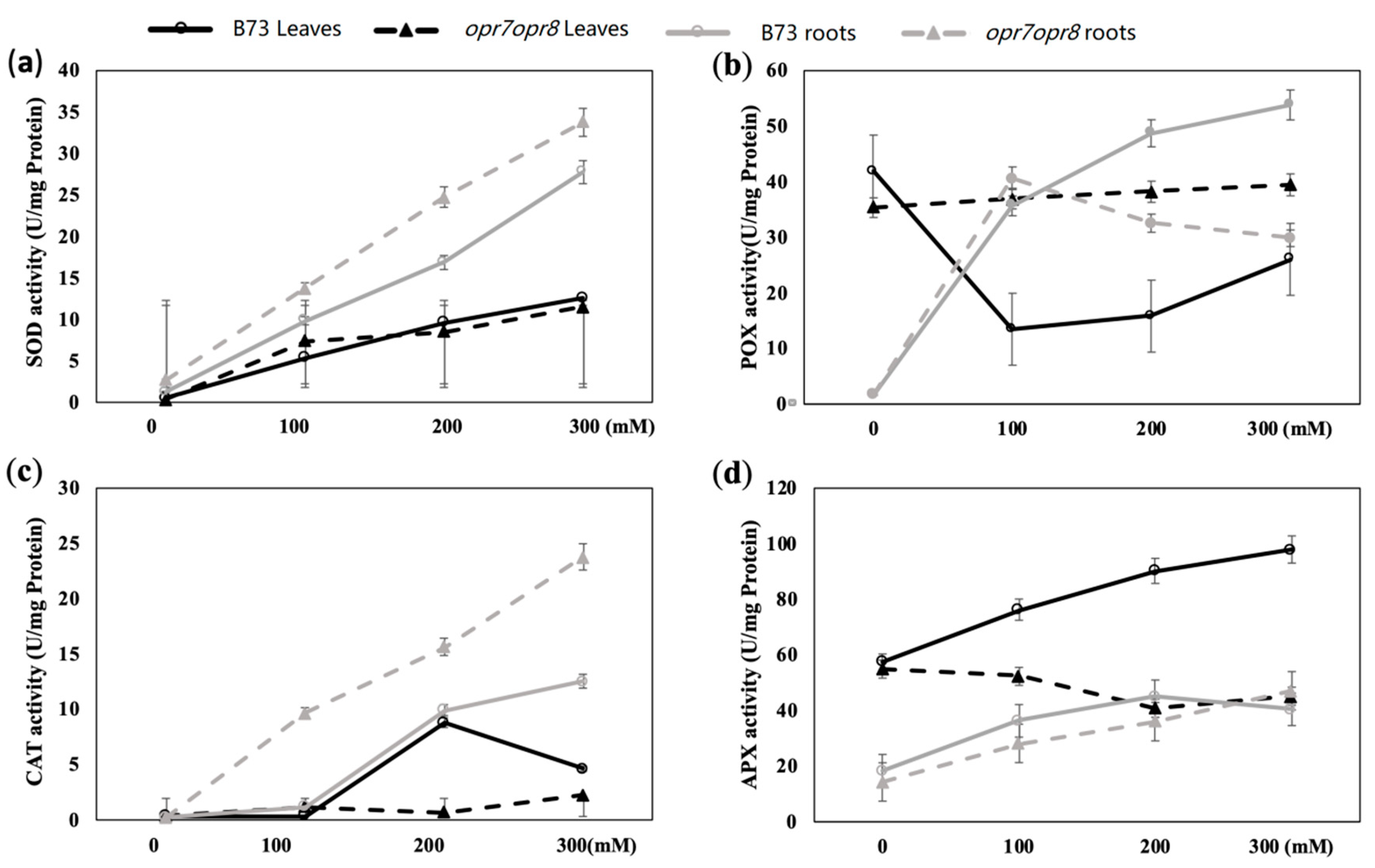
2.6. opr7opr8 Displayed Different Antioxidant Enzyme Activities from WT under Salt Treatment
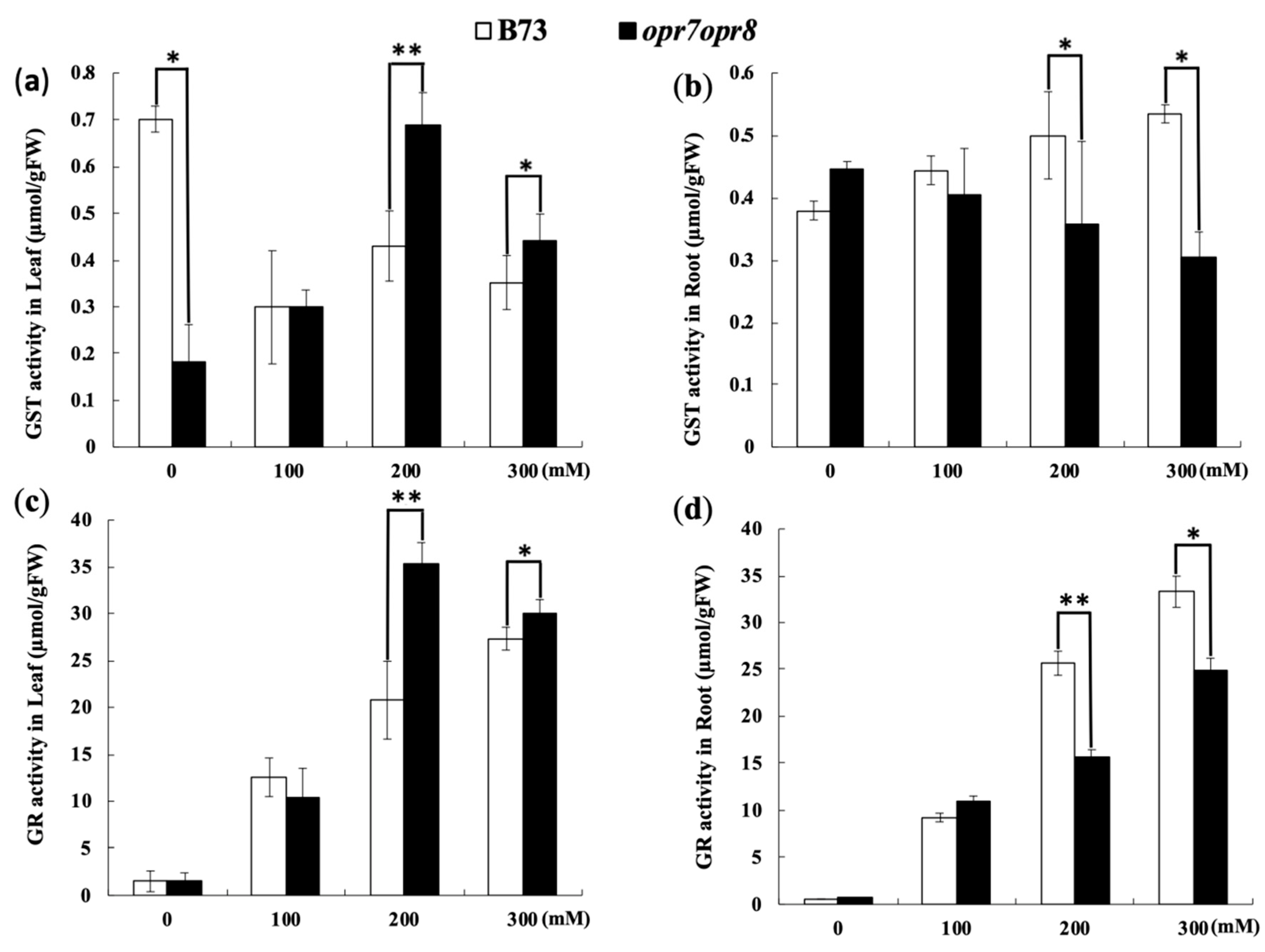
2.7. opr7opr8 Exhibited Different Levels of Glutathione Reductase (GR) and Glutathione-S-Transferase (GST) Activities from WT under Salt Treatment
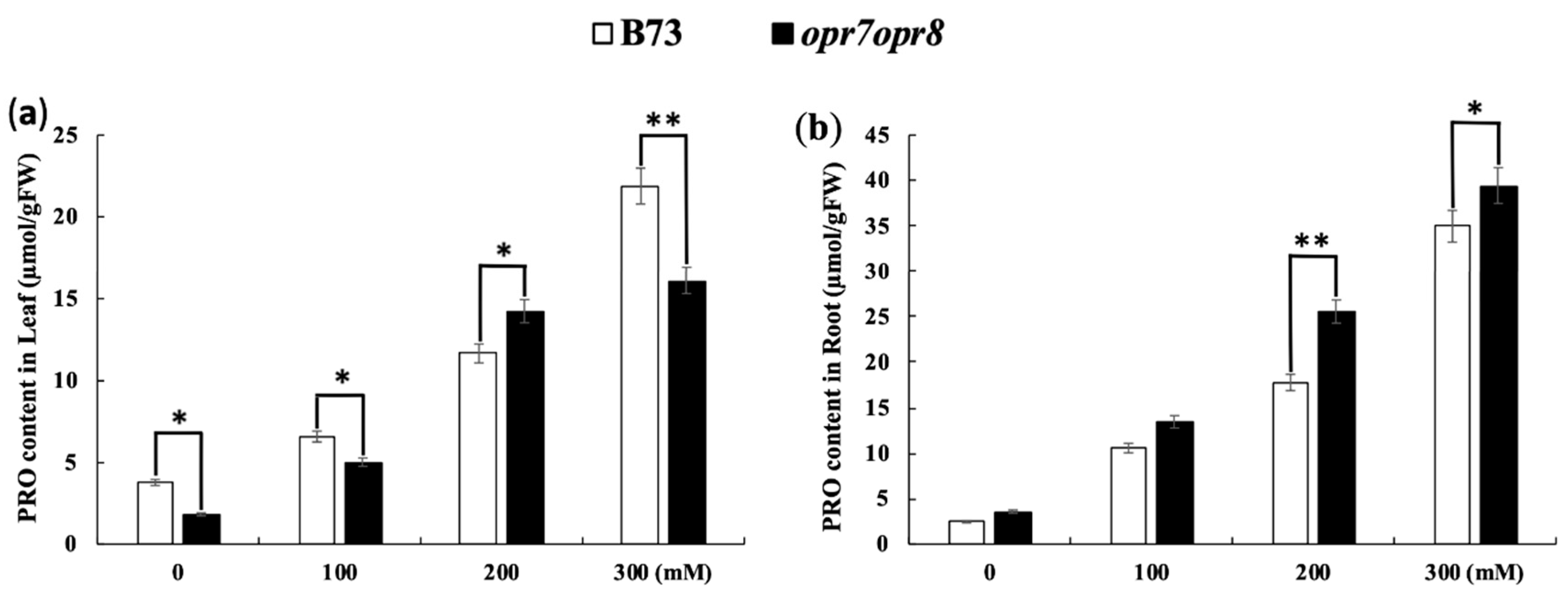
2.8. opr7opr8 Accumulated More Proline in the Roots but Less in the Leaves Than WT under Salt Stress
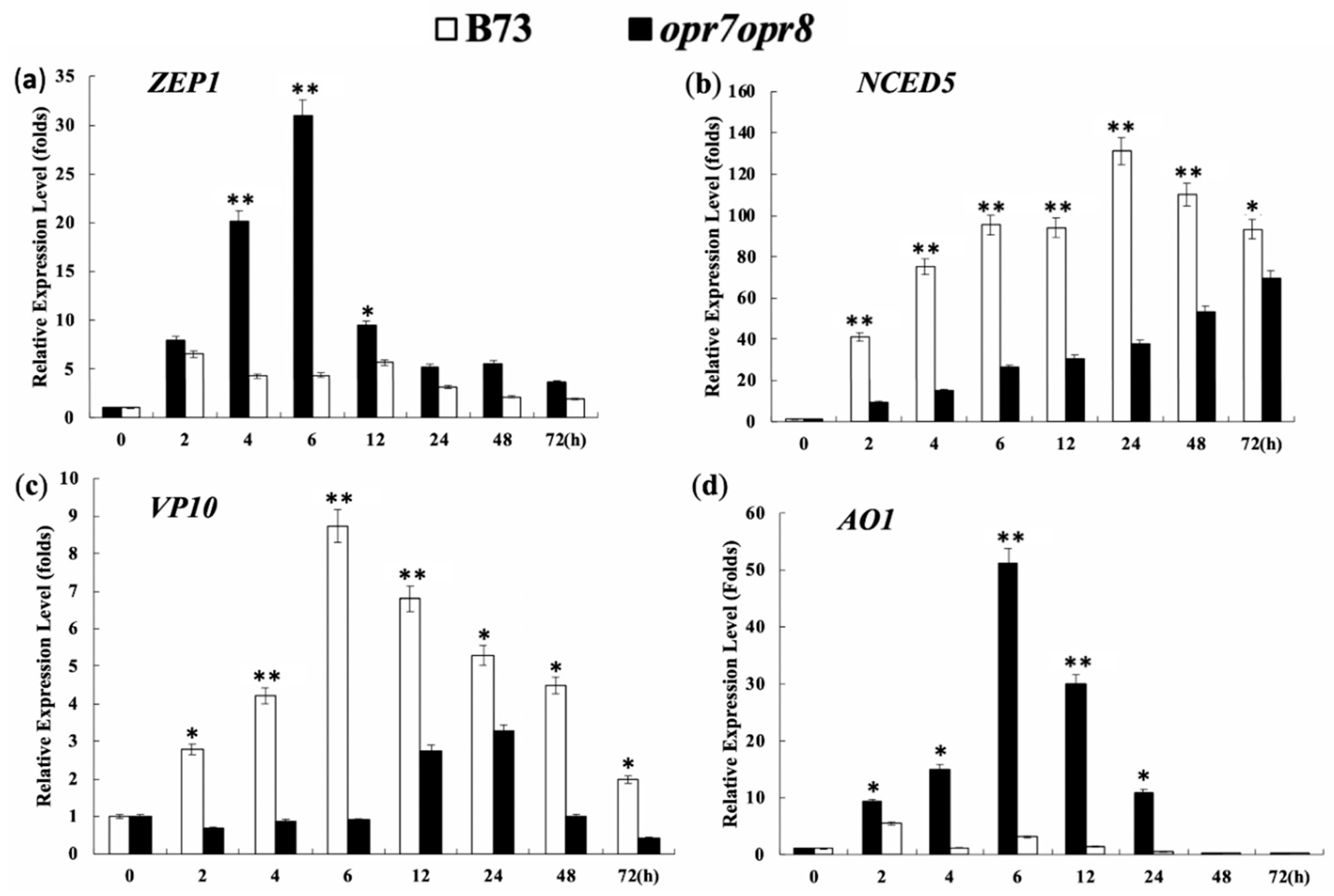
2.9. Endogenous JA Production Is Required for Transcriptional Activation of ABA Biosynthesis Genes under Salt Stress
3. Discussion
4. Materials and Methods
4.1. Experimental Material, Planting, and Salt Treatments
4.2. Analysis of Root and Shoot Elongation
4.3. Ion Content Profiling
4.4. Determination of Enzymatic Antioxidants
4.5. Photosynthetic Pigments
4.6. Stomatal Imaging and Quantification
4.7. Reactive Oxygen Species Visualization
4.8. Gene Expression Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FRW | Fresh root weight |
| FSW | Fresh shoot weight |
| DRW | Dry root weight |
| DSW | Dry shoot weight |
| FSL | Fresh shoot length |
| FRL | Fresh shoot length |
| Pro | Proline |
| MDA | Malondialdehyde |
| Chl | Chlorophyll |
| CAT | Catalase |
| SOD | Superoxide dismutase |
| APX | Ascorbate peroxidase |
References
- Setia, R.; Gottschalk, P.; Smith, P.; Marschner, P.; Baldock, J.; Setia, D.; Smith, J. Soil salinity decreases global soil organic carbon stocks. Sci. Total Environ. 2013, 465, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flowers, T.J. Improving crop salt tolerance. J. Exp. Bot. 2004, 55, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant. Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop. J. 2016, 4, 162–176. [Google Scholar] [CrossRef] [Green Version]
- Ku, Y.S.; Sintaha, M.; Cheung, M.Y.; Lam, H.M. Plant Hormone Signaling Crosstalks between Biotic and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [Green Version]
- Popko, J.; Hansch, R.; Mendel, R.R.; Polle, A.; Teichmann, T. The role of abscisic acid and auxin in the response of poplar to abiotic stress. Plant. Biol. 2010, 12, 242–258. [Google Scholar] [CrossRef]
- Shu, K.; Zhou, W.; Chen, F.; Luo, X.; Yang, W. Abscisic Acid and Gibberellins Antagonistically Mediate Plant Development and Abiotic Stress Responses. Front. Plant. Sci. 2018, 9, 416. [Google Scholar] [CrossRef] [Green Version]
- Zwack, P.J.; Rashotte, A.M. Interactions between cytokinin signalling and abiotic stress responses. J. Exp. Bot. 2015, 66, 4863–4871. [Google Scholar] [CrossRef]
- Tohidi, B.; Rahimmalek, M.; Trindade, H. Review on essential oil, extracts composition, molecular and phytochemical properties of Thymus species in Iran. Ind. Crop. Prod. 2019, 134, 89–99. [Google Scholar] [CrossRef]
- Ghasemi Pirbalouti, A.; Sajjadi, S.E.; Parang, K. ChemInform Abstract: A Review (Research and Patents) on Jasmonic Acid and Its Derivatives. Arch. Der Pharm. 2014, 347. [Google Scholar] [CrossRef]
- Farmer, E.E.; Almeras, E.; Krishnamurthy, V. Jasmonates and related oxylipins in plant responses to pathogenesis and herbivory. Curr. Opin. Plant. Biol. 2003, 6, 372–378. [Google Scholar] [CrossRef]
- Browse, J. Jasmonate passes muster: A receptor and targets for the defense hormone. Annu. Rev. Plant. Biol. 2009, 60, 183–205. [Google Scholar] [CrossRef]
- Wasternack, C.; Song, S. Jasmonates: Biosynthesis, metabolism, and signaling by proteins activating and repressing transcription. J. Exp. Bot. 2016, 68, 1303–1321. [Google Scholar] [CrossRef]
- McConn, M.; Browse, J. The Critical Requirement for Linolenic Acid Is Pollen Development, Not Photosynthesis, in an Arabidopsis Mutant. Plant. Cell 1996, 8, 403–416. [Google Scholar] [CrossRef]
- Sanders, P.M.; Lee, P.Y.; Biesgen, C.; Boone, J.D.; Beals, T.P.; Weiler, E.W.; Goldberg, R.B. The Arabidopsis DELAYED DEHISCENCE1 Gene Encodes an Enzyme in the Jasmonic Acid Synthesis Pathway. Plant. Cell 2000, 12, 1041–1061. [Google Scholar] [CrossRef] [Green Version]
- Stintzi, A.; Browse, J. The Arabidopsis male-sterile mutant, opr3, lacks the 12-oxophytodienoic acid reductase required for jasmonate synthesis. Proc. Natl Acad Sci. USA 2000, 97, 10625–10630. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Halitschke, R.; Kim, H.B.; Baldwin, I.T.; Feldmann, K.A.; Feyereisen, R. A knock-out mutation in allene oxide synthase results in male sterility and defective wound signal transduction in Arabidopsis due to a block in jasmonic acid biosynthesis. Plant. J. Cell Mol. Biol. 2002, 31, 1–12. [Google Scholar] [CrossRef]
- Feys, B.; Benedetti, C.E.; Penfold, C.N.; Turner, J.G. Arabidopsis Mutants Selected for Resistance to the Phytotoxin Coronatine Are Male Sterile, Insensitive to Methyl Jasmonate, and Resistant to a Bacterial Pathogen. Plant. Cell 1994, 6, 751–759. [Google Scholar] [CrossRef] [Green Version]
- Browse, J. The power of mutants for investigating jasmonate biosynthesis and signaling. Phytochemistry 2009, 70, 1539–1546. [Google Scholar] [CrossRef]
- Staswick, P.E.; Su, W.; Howell, S.H. Methyl jasmonate inhibition of root growth and induction of a leaf protein are decreased in an Arabidopsis thaliana mutant. Proc. Natl. Acad Sci. USA 1992, 89, 6837–6840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, O.; Chico, J.M.; Sanchez-Serrano, J.J.; Solano, R. JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant. Cell 2004, 16, 1938–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.I.; Howe, G.A. The tomato mutant spr1 is defective in systemin perception and the production of a systemic wound signal for defense gene expression. Plant. J.: Cell Mol. Biol. 2003, 33, 567–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Li, C.; Lee, G.I.; Howe, G.A. Distinct roles for jasmonate synthesis and action in the systemic wound response of tomato. Proc. Natl. Acad Sci. USA 2002, 99, 6416–6421. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhao, Y.; McCaig, B.C.; Wingerd, B.A.; Wang, J.; Whalon, M.E.; Pichersky, E.; Howe, G.A. The tomato homolog of CORONATINE-INSENSITIVE1 is required for the maternal control of seed maturation, jasmonate-signaled defense responses, and glandular trichome development. Plant. Cell 2004, 16, 126–143. [Google Scholar] [CrossRef] [Green Version]
- Maksymiec, W.; Wianowska, D.; Dawidowicz, A.L.; Radkiewicz, S.; Mardarowicz, M.; Krupa, Z. The level of jasmonic acid in Arabidopsis thaliana and Phaseolus coccineus plants under heavy metal stress. J. Plant. Physiol. 2005, 162, 1338–1346. [Google Scholar] [CrossRef]
- Dong, W.; Wang, M.; Xu, F.; Quan, T.; Peng, K.; Xiao, L.; Xia, G. Wheat oxophytodienoate reductase gene TaOPR1 confers salinity tolerance via enhancement of abscisic acid signaling and reactive oxygen species scavenging. Plant. Physiol. 2013, 161, 1217–1228. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Guo, J.; Zhu, A.; Zhang, L.; Zhang, M. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 2014, 104, 202–208. [Google Scholar] [CrossRef]
- Zhao, Y.; Dong, W.; Zhang, N.; Ai, X.; Wang, M.; Huang, Z.; Xiao, L.; Xia, G. A wheat allene oxide cyclase gene enhances salinity tolerance via jasmonate signaling. Plant. Physiol. 2014, 164, 1068–1076. [Google Scholar] [CrossRef] [Green Version]
- Brossa, R.; López-Carbonell, M.; Jubany-Marí, T.; Alegre, L. Interplay Between Abscisic Acid and Jasmonic Acid and its Role in Water-oxidative Stress in Wild-type, ABA-deficient, JA-deficient, and Ascorbate-deficient Arabidopsis Plants. J. Plant. Growth Regul. 2011, 30, 322–333. [Google Scholar] [CrossRef]
- Clarke, S.M.; Cristescu, S.M.; Miersch, O.; Harren, F.J.; Wasternack, C.; Mur, L.A. Jasmonates act with salicylic acid to confer basal thermotolerance in Arabidopsis thaliana. New Phytol. 2009, 182, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Laxmi, A. Jasmonates: Emerging Players in Controlling Temperature Stress Tolerance. Front. Plant. Sci. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant. Sci. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, A.; Riemann, M.; Nick, P. The jasmonate pathway mediates salt tolerance in grapevines. J. Exp. Bot. 2012, 63, 2127–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.Y.; Hamayun, M.; Lee, S.-K.; Lee, I.-J. Methyl jasmonate alleviated salinity stress in soybean. J. Crop. Sci. Biotechnol. 2009, 12, 63–68. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Hosseinzadeh-Mahootchi, A. Improving physiological performance of safflower under salt stress by application of salicylic acid and jasmonic acid. WALIA J. 2015, 31, 104–109. [Google Scholar]
- Faghih, S.; Ghobadi, C.; Zarei, A. Response of Strawberry Plant cv. ‘Camarosa’ to Salicylic Acid and Methyl Jasmonate Application Under Salt Stress Condition. J. Plant. Growth Regul. 2017, 36. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. How can salicylic acid and jasmonic acid mitigate salt toxicity in soybean plants? Ecotoxicol. Environ. Saf. 2018, 147, 1010–1016. [Google Scholar] [CrossRef]
- Hazman, M.; Hause, B.; Eiche, E.; Nick, P.; Riemann, M. Increased tolerance to salt stress in OPDA-deficient rice ALLENE OXIDE CYCLASE mutants is linked to an increased ROS-scavenging activity. J. Exp. Bot. 2015, 66, 3339–3352. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Cai, H.; Luo, X.; Bai, X.; Deyholos, M.K.; Chen, Q.; Chen, C.; Ji, W.; Zhu, Y. Over-expression of a novel JAZ family gene from Glycine soja, increases salt and alkali stress tolerance. Biochem. Biophys. Res. Commun. 2012, 426, 273–279. [Google Scholar] [CrossRef]
- Yan, Y.; Christensen, S.; Isakeit, T.; Engelberth, J.; Meeley, R.; Hayward, A.; Emery, R.J.; Kolomiets, M.V. Disruption of OPR7 and OPR8 reveals the versatile functions of jasmonic acid in maize development and defense. Plant. Cell 2012, 24, 1420–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakeel, A. Potassium–sodium interactions in soil and plant under saline-sodic conditions. J. Plant. Nutr. Soil Sci. 2013, 176, 344–354. [Google Scholar] [CrossRef]
- Saxena, I.; Srikanth, S.; Chen, Z. Cross Talk between H2O2 and Interacting Signal Molecules under Plant Stress Response. Front. Plant. Sci. 2016, 7, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, L.; Zhu, J.K. Regulation of abscisic acid biosynthesis. Plant. Physiol. 2003, 133, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Porch, T.G.; Tseung, C.W.; Schmelz, E.A.; Settles, A.M. The maize Viviparous10/Viviparous13 locus encodes the Cnx1 gene required for molybdenum cofactor biosynthesis. Plant. J. Cell Mol. Biol. 2006, 45, 250–263. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant. Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant. Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.; Cho, Y.-G. Plant hormones in salt stress tolerance. J. Plant. Biol. 2015, 58, 147–155. [Google Scholar] [CrossRef]
- Raghavendra, A.S.; Gonugunta, V.K.; Christmann, A.; Grill, E. ABA perception and signalling. Trends Plant. Sci. 2010, 15, 395–401. [Google Scholar] [CrossRef]
- Kazan, K. Diverse roles of jasmonates and ethylene in abiotic stress tolerance. Trends Plant. Sci. 2015, 20, 219–229. [Google Scholar] [CrossRef]
- Riemann, M.; Dhakarey, R.; Hazman, M.; Miro, B.; Kohli, A.; Nick, P. Exploring Jasmonates in the Hormonal Network of Drought and Salinity Responses. Front. Plant. Sci. 2015, 6, 1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela, C.E.; Acevedo-Acevedo, O.; Miranda, G.S.; Vergara-Barros, P.; Holuigue, L.; Figueroa, C.R.; Figueroa, P.M. Salt stress response triggers activation of the jasmonate signaling pathway leading to inhibition of cell elongation in Arabidopsis primary root. J. Exp. Bot. 2016, 67, 4209–4220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdala, G. Jasmonate and octadecanoid occurrence in tomato hairy roots. Endogenous level changes in response to NaCl. Plant. Growth Regul. 2003, 40, 21–27. [Google Scholar] [CrossRef]
- Pedranzani, H.; Racagni, G.; Alemano, S.; Miersch, O.; Ramírez, I.; Pena-Cortes, H.; Taleisnik, E.; Machado, E.; Abdala, G. Salt tolerant tomato plants show increased levels of jasmonic acid. Plant. Growth Regul. 2003, 41, 149–158. [Google Scholar] [CrossRef]
- Moons, A.; Prinsen, E.; Bauw, G.; Van Montagu, M. Antagonistic Effects of Abscisic Acid and Jasmonates on Salt Stress-Inducible Transcripts in Rice Roots. Plant. Cell 1997, 9, 2243–2259. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.-J.; Seo, Y.J.; Lee, J.D.; Ishii, R.; Kim, K.U.; Shin, D.H.; Park, S.K.; Jang, S.W.; Lee, I.J. Jasmonic Acid Differentially Affects Growth, Ion Uptake and Abscisic Acid Concentration in Salt-tolerant and Salt-sensitive Rice Cultivars. J. Agron. Crop. Sci. 2005, 191, 273–282. [Google Scholar] [CrossRef]
- Shahzad, A.N.; Pitann, B.; Ali, H.; Qayyum, M.F.; Fatima, A.; Bakhat, H.F. Maize Genotypes Differing in Salt Resistance Vary in Jasmonic Acid Accumulation During the First Phase of Salt Stress. J. Agron. Crop. Sci. 2015, 201, 443–451. [Google Scholar] [CrossRef]
- Pavlović, I.; Pencík, A.; Novak, O.; Vujčić, V.; Radić, S.; Lepeduš, H.; Strnad, M.; Salopek-Sondi, B. Short-term salt stress in Brassica rapa seedlings causes alterations in auxin metabolism. Plant. Physiol. Biochem. 2018, 125. [Google Scholar] [CrossRef]
- Mahmud, S.; Sharmin, S.; Chowdhury, B.; Hossain, M. Effect of Salinity and Alleviating Role of Methyl Jasmonate in Some Rice Varieties. Asian J. Plant. Sci. 2017, 16, 87–93. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Farhangi-Abriz, S. Foliar sprays of salicylic acid and jasmonic acid stimulate H(+)-ATPase activity of tonoplast, nutrient uptake and salt tolerance of soybean. Ecotoxicol. Environ. Saf. 2018, 166, 18–25. [Google Scholar] [CrossRef]
- Ahmadi, F.I.; Karimi, K.; Struik, P.C. Effect of exogenous application of methyl jasmonate on physiological and biochemical characteristics of Brassica napus L. cv. Talaye under salinity stress. South. Afr. J. Bot. 2018, 115, 5–11. [Google Scholar] [CrossRef]
- Mir, M.A.; John, R.; Alyemeni, M.N.; Alam, P.; Ahmad, P. Jasmonic acid ameliorates alkaline stress by improving growth performance, ascorbate glutathione cycle and glyoxylase system in maize seedlings. Sci. Rep. 2018, 8, 2381. [Google Scholar] [CrossRef]
- Kurotani, K.; Hayashi, K.; Hatanaka, S.; Toda, Y.; Ogawa, D.; Ichikawa, H.; Ishimaru, Y.; Tashita, R.; Suzuki, T.; Ueda, M.; et al. Elevated levels of CYP94 family gene expression alleviate the jasmonate response and enhance salt tolerance in rice. Plant. Cell Physiol. 2015, 56, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Ye, H.; Yao, R.; Zhang, T.; Xiong, L. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling and modulates salt stress tolerance in rice. Plant. Sci. Int. J. Exp. Plant. Biol. 2015, 232, 1–12. [Google Scholar] [CrossRef]
- Peethambaran, P.K.; Glenz, R.; Honinger, S.; Shahinul Islam, S.M.; Hummel, S.; Harter, K.; Kolukisaoglu, U.; Meynard, D.; Guiderdoni, E.; Nick, P.; et al. Salt-inducible expression of OsJAZ8 improves resilience against salt-stress. Bmc Plant. Biol. 2018, 18, 311. [Google Scholar] [CrossRef]
- Yastreb, T.; Kolupaev, Y.; Kokorev, A.; Horielova, E.; Dmitriev, A. Methyl Jasmonate and Nitric Oxide in Regulation of the Stomatal Apparatus of Arabidopsis thaliana. Cytol. Genet. 2018, 52, 400–405. [Google Scholar] [CrossRef]
- Hossain, M.A.; Munemasa, S.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Murata, Y. Involvement of endogenous abscisic acid in methyl jasmonate-induced stomatal closure in Arabidopsis. Plant. Physiol. 2011, 156, 430–438. [Google Scholar] [CrossRef] [Green Version]
- Sanz, L.C.; Fernández-Maculet, J.C.; Gómez, E.; Vioque, B.; Olías, J.M. Effect of methyl jasmonate on ethylene biosynthesis and stomatal closure in olive leaves. Phytochemistry 1993, 33, 285–289. [Google Scholar] [CrossRef]
- Metodiev, M.; Tsonev, T.; Popova, L. Effect of jasmonic acid on the stomatal and nonstomatal limitation of leaf photosynthesis in barley leaves. J. Plant. Growth Regul. 1996, 15, 75–80. [Google Scholar] [CrossRef]
- Han, X.; Hu, Y.; Zhang, G.; Jiang, Y.; Chen, X. Jasmonate Negatively Regulates Stomatal Development in Arabidopsis Cotyledons. Plant Physiol. 2018, 176, 2871–2885. [Google Scholar] [CrossRef] [Green Version]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High Salinity Induces Different Oxidative Stress and Antioxidant Responses in Maize Seedlings Organs. Front. Plant. Sci. 2016, 7, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant. Physiol. Biochem. Ppb 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant. Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. Jasmonates: Mechanisms and functions in abiotic stress tolerance of plants. Biocatal. Agric. Biotechnol. 2019, 20, 101210. [Google Scholar] [CrossRef]
- Soares, A.M.d.S.; Souza, T.F.d.; Jacinto, T.; Machado, O.L.T. Effect of Methyl Jasmonate on antioxidative enzyme activities and on the contents of ROS and H2O2 in Ricinus communis leaves. Braz. J. Plant. Physiol. 2010, 22, 151–158. [Google Scholar] [CrossRef]
- Parra-Lobato, M.; Fernández-García, N.; Olmos, E.; Alvarez-Tinaut, M.; Gomez-Jimenez, M. Methyl jasmonate-induced antioxidant defence in root apoplast from sunflower seedlings. Environ. Exp. Bot. 2009, 66, 9–17. [Google Scholar] [CrossRef]
- Regni, L.; Del Pino, A.M.; Mousavi, S.; Palmerini, C.A.; Baldoni, L.; Mariotti, R.; Mairech, H.; Gardi, T.; D’Amato, R.; Proietti, P. Behavior of Four Olive Cultivars During Salt Stress. Front. Plant. Sci. 2019, 10. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant. Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Kundu, S.; Gantait, S. Abscisic acid signal crosstalk during abiotic stress response. Plant. Gene. 2017, 11, 61–69. [Google Scholar] [CrossRef]
- SestÁK, Z.; CatskÝ, J.; Jarvis, P.G. Plant Photosynthetic Production. Manual of Methods; Dr. W. Junk NV: The Hague, The Netherlands, 1971; p. 818. [Google Scholar]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, R.M.; Cheng, C.; Sheng, J.; Wang, W.; Ren, H.; Aslam, M.; Yan, Y. Interruption of Jasmonic Acid Biosynthesis Causes Differential Responses in the Roots and Shoots of Maize Seedlings against Salt Stress. Int. J. Mol. Sci. 2019, 20, 6202. https://doi.org/10.3390/ijms20246202
Ahmad RM, Cheng C, Sheng J, Wang W, Ren H, Aslam M, Yan Y. Interruption of Jasmonic Acid Biosynthesis Causes Differential Responses in the Roots and Shoots of Maize Seedlings against Salt Stress. International Journal of Molecular Sciences. 2019; 20(24):6202. https://doi.org/10.3390/ijms20246202
Chicago/Turabian StyleAhmad, Ramala Masood, Cheng Cheng, Jia Sheng, Wei Wang, Hong Ren, Muhammad Aslam, and Yuanxin Yan. 2019. "Interruption of Jasmonic Acid Biosynthesis Causes Differential Responses in the Roots and Shoots of Maize Seedlings against Salt Stress" International Journal of Molecular Sciences 20, no. 24: 6202. https://doi.org/10.3390/ijms20246202