RNA-Seq-Based Comparative Transcriptome Analysis Highlights New Features of the Heat-Stress Response in the Extremophilic Bacterium Deinococcus radiodurans

Abstract
:1. Introduction
2. Results
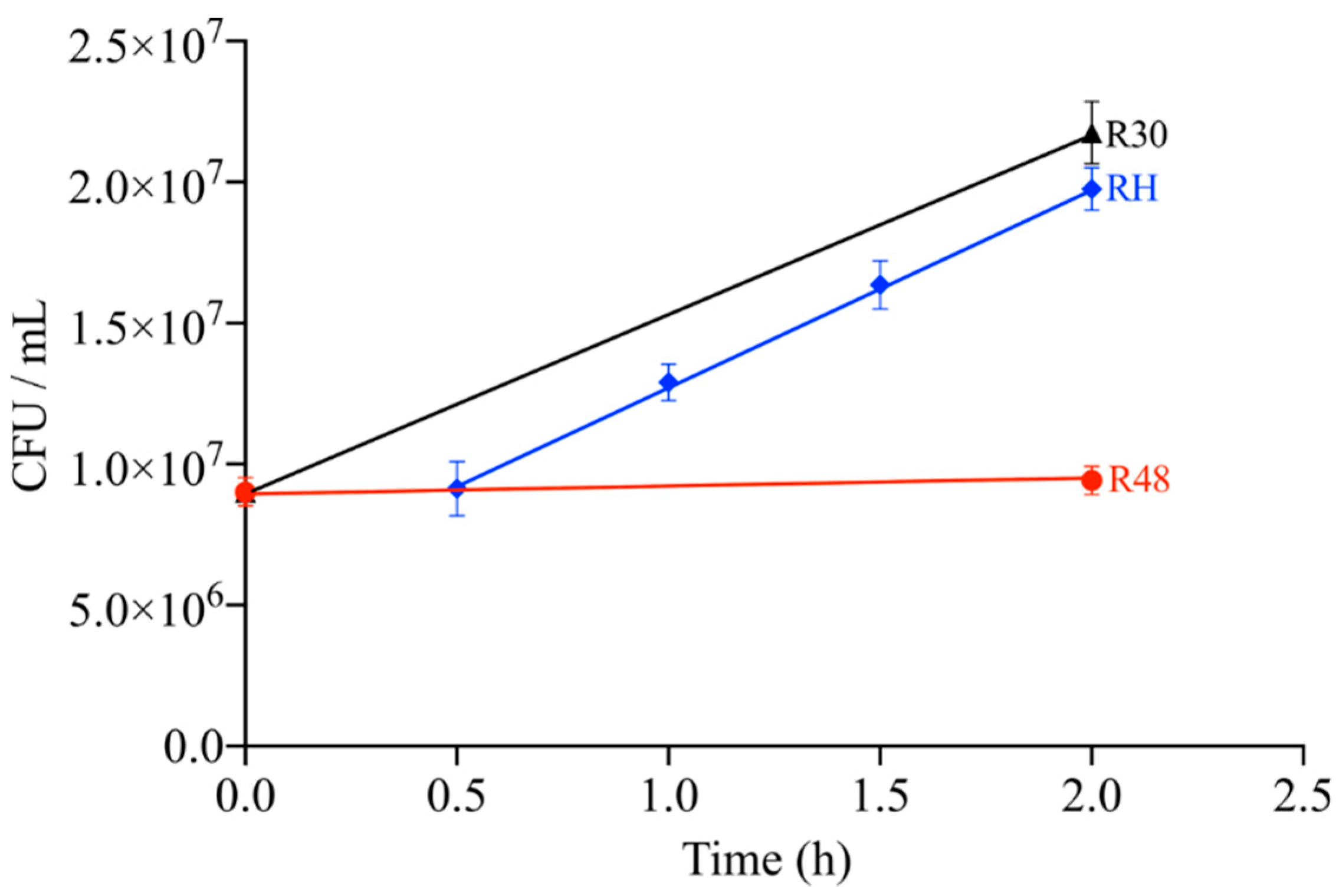
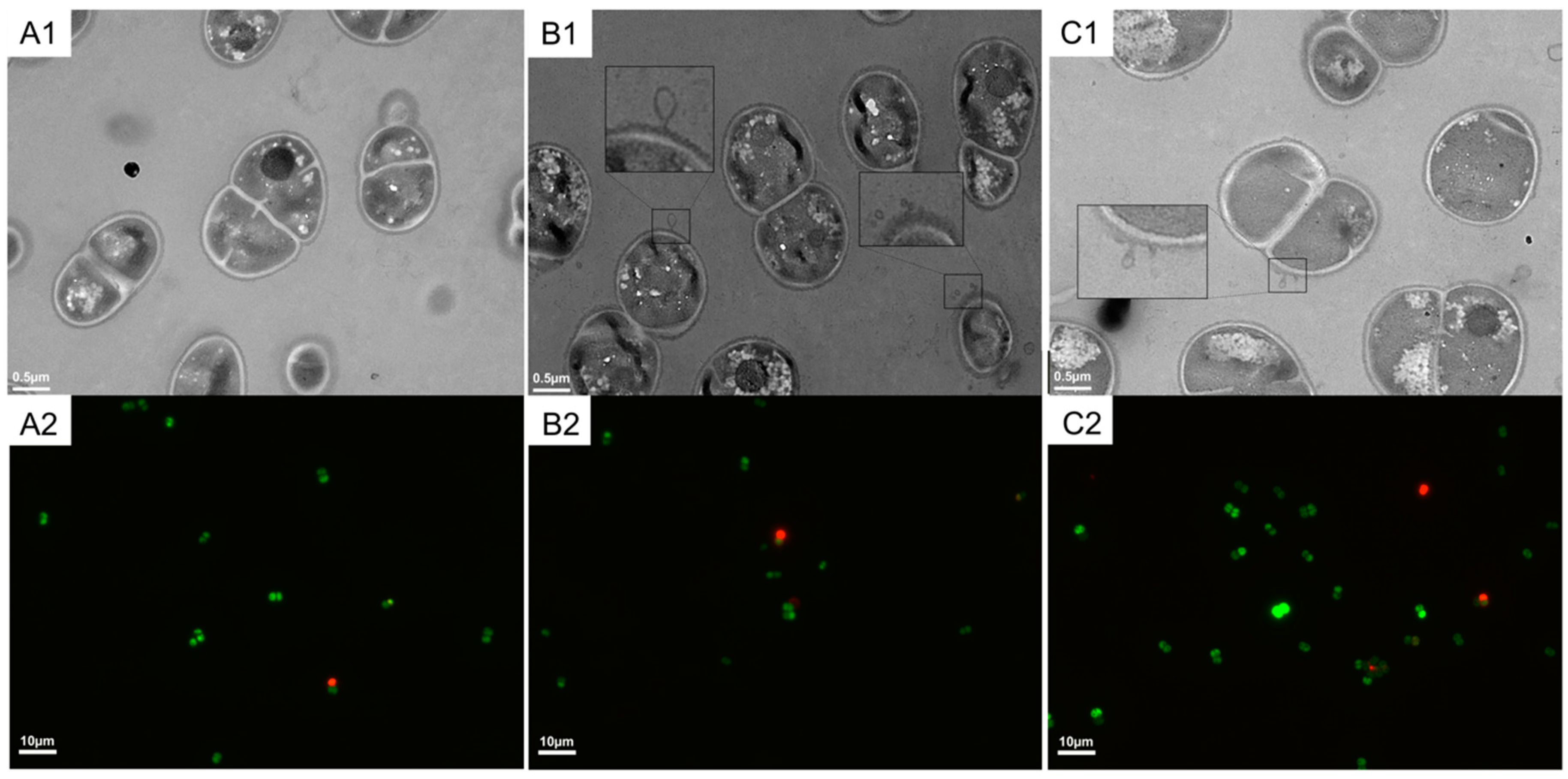
2.1. Cell Growth State and Viability of D. radiodurans under Heat Stress
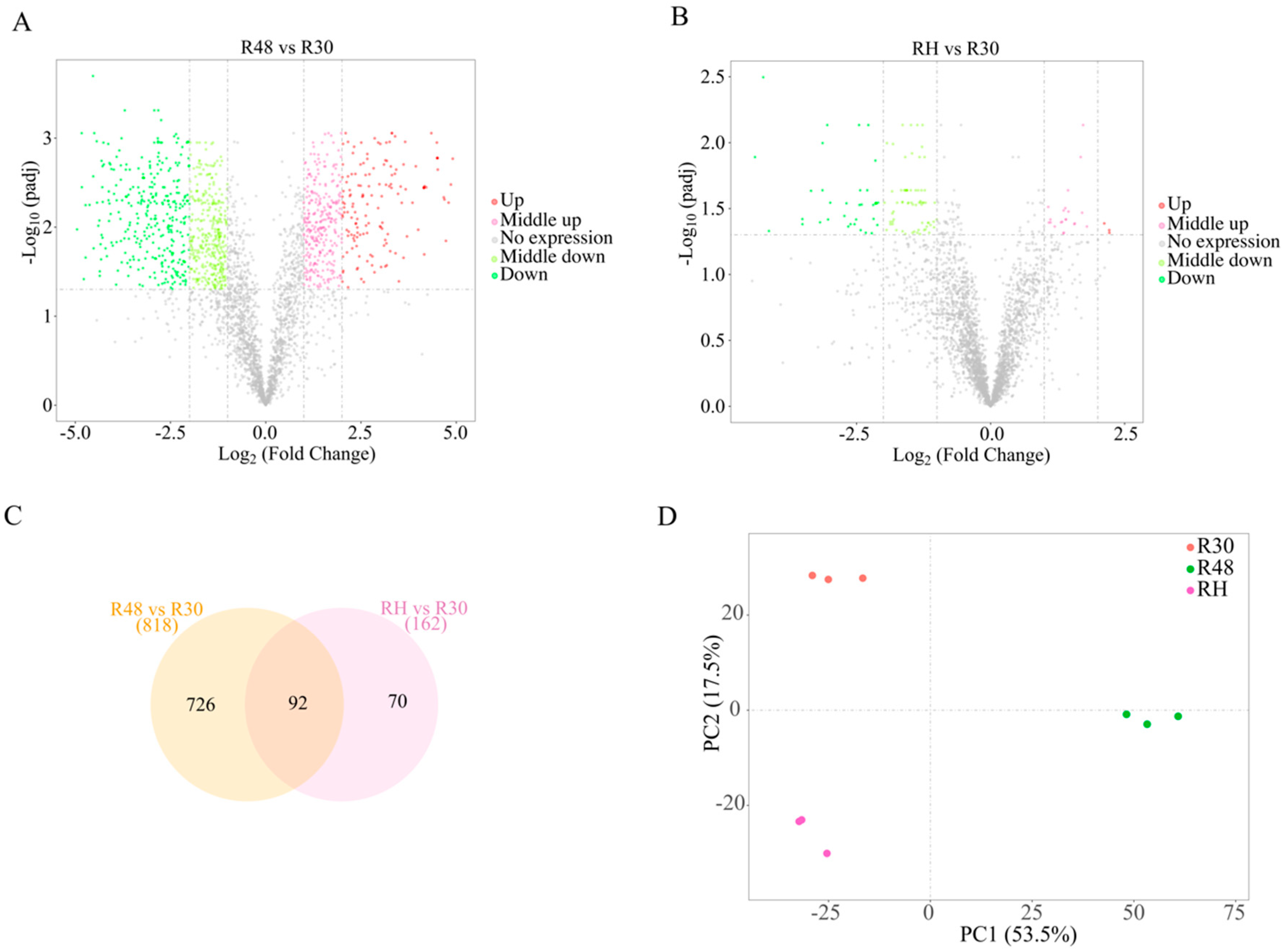
2.2. Qualitative and Quantitative Analyses of the Differentially Expressed Genes (DEGs) in the Three Groups under Heat Stress
2.3. Gene Ontology (GO) and Kyoto Encyclopedia of Gene and Genomes (KEGG) Analyses of the DEGs
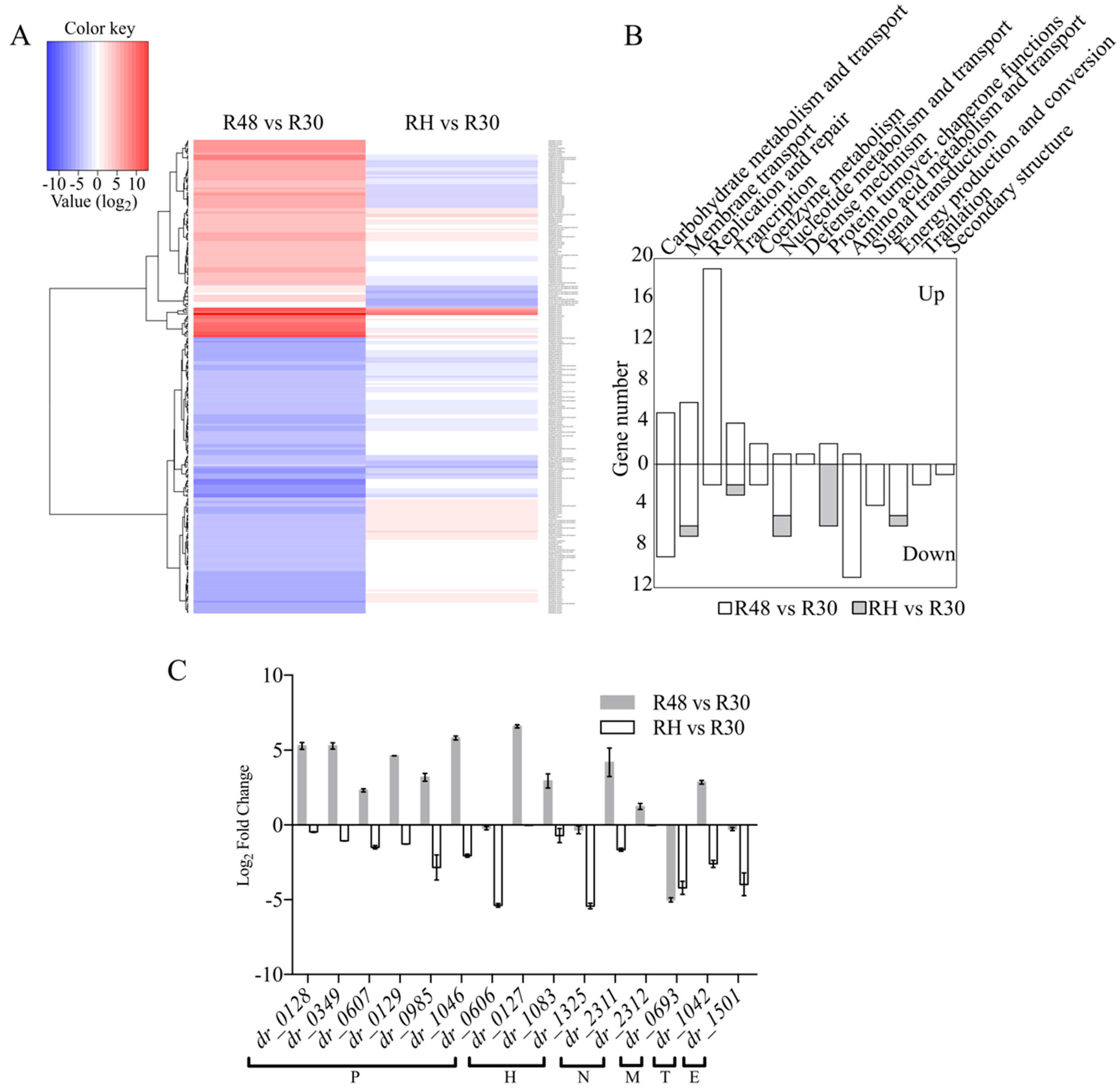
2.4. Analysis of Heat-Related Genes with over Eight-Fold Higher Expressions in Response to Heat Stress
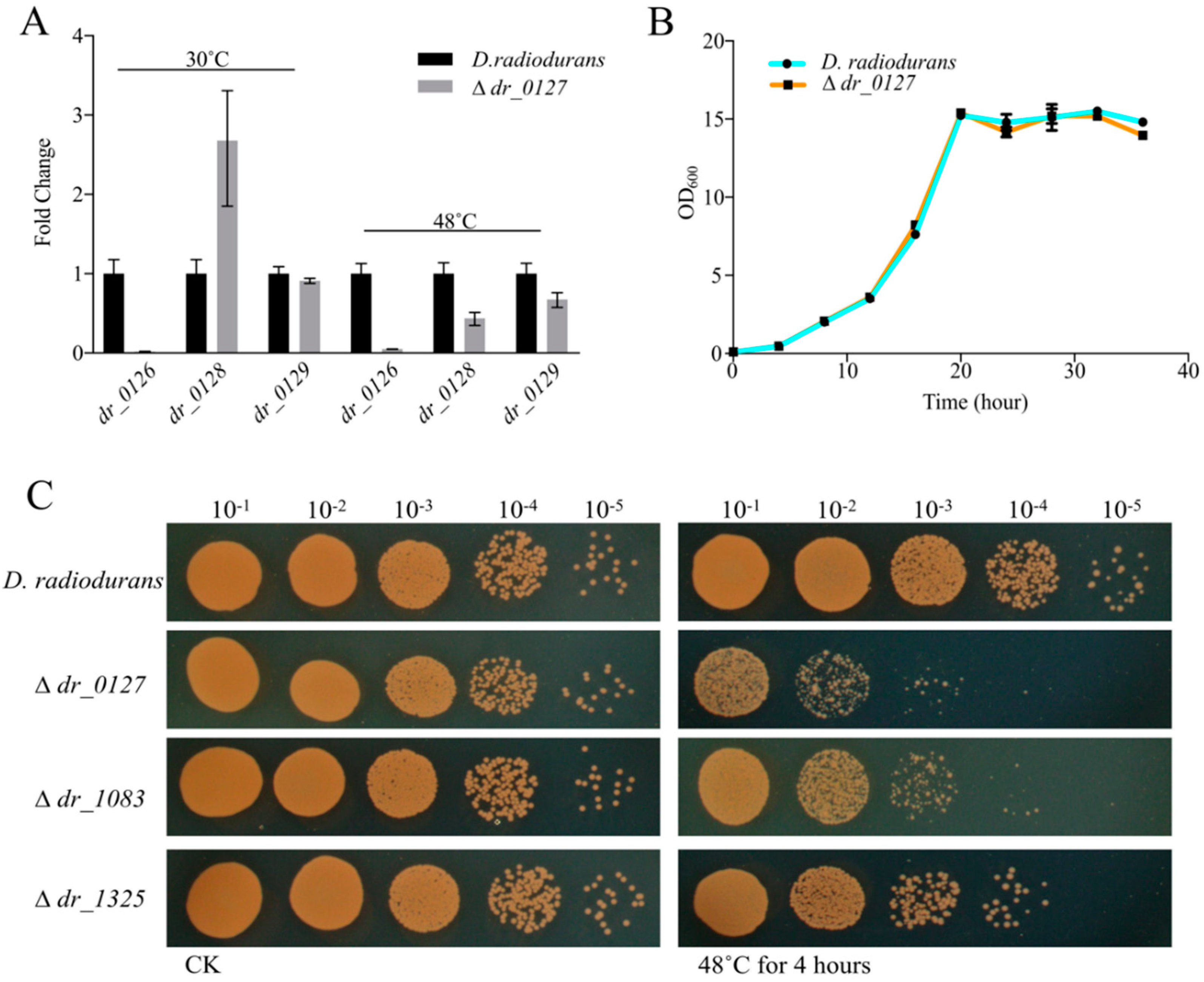
2.5. Function Analysis of the Novel, Potentially Heat-Related Genes
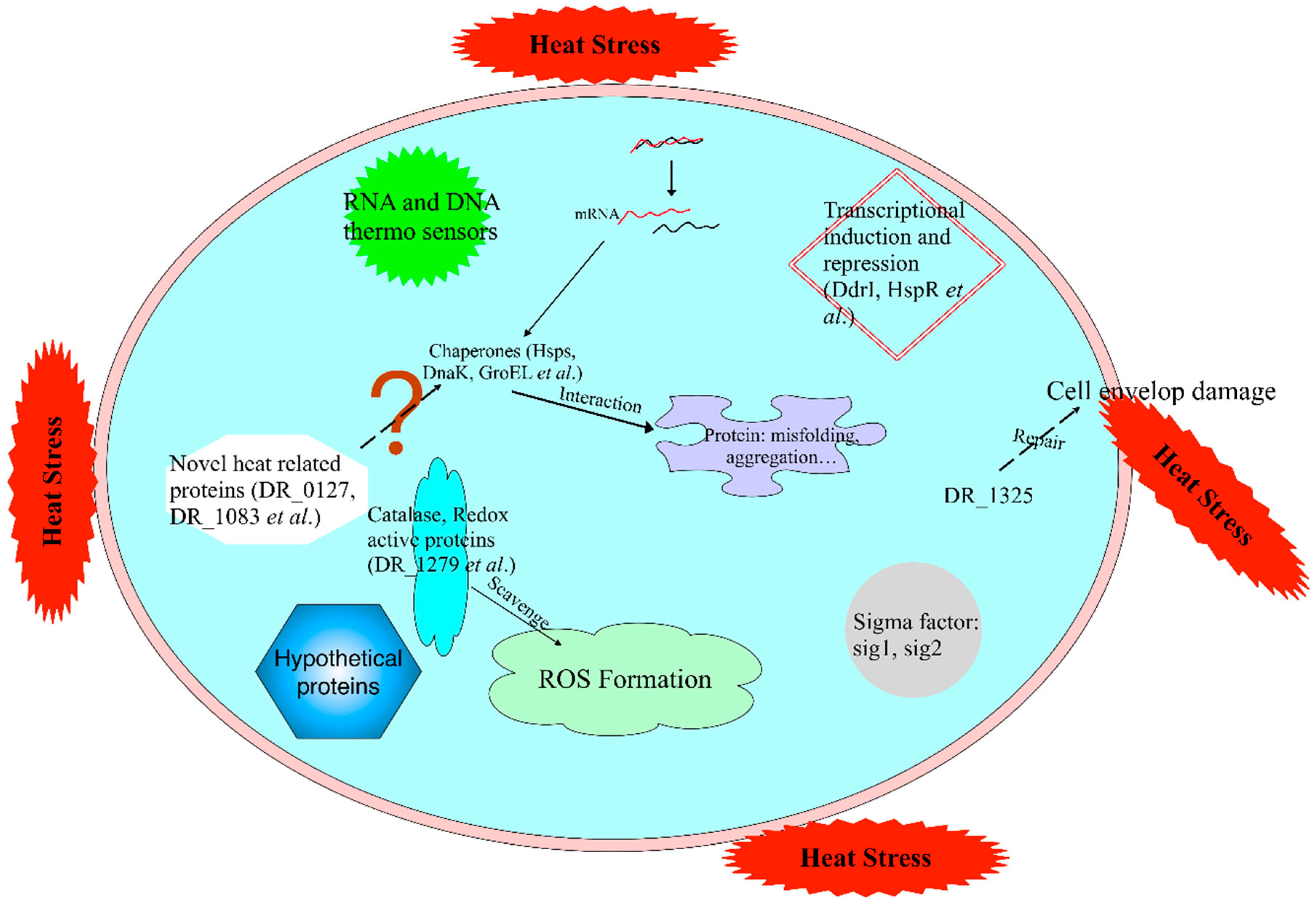
3. Discussion
4. Materials and Methods
4.1. Strain and Growth Conditions
4.2. Heat Stress Treatment of D. radiodurans and Recovery Conditions
4.3. TEM and Fluorescence Assay
4.4. Total RNA Extraction, complementary DNA (cDNA) Library Preparation, and Sequencing
4.5. Assembly and Functional Enrichment Analyses of DEGs
4.6. GO and KEGG Enrichment Analysis
4.7. qRT-PCR Validation
4.8. Construction of Gene Deletion Mutant Strains and Heat Stress Phenotype Assays
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Battista, J.R. Against all odds: the survival strategies of Deinococcus radiodurans. Annu. Rev. Microbiol. 1997, 51, 203–224. [Google Scholar] [CrossRef] [PubMed]
- White, O.; Eisen, J.A.; Heidelberg, J.F.; Hickey, E.K.; Peterson, J.D.; Dodson, R.J.; Haft, D.H.; Gwinn, M.L.; Nelson, W.C.; Richardson, D.L.; et al. Genome sequence of the radioresistant bacterium Deinococcus radiodurans R1. Science 1999, 286, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Mattimore, V.; Battista, J.R. Radioresistance of Deinococcus radiodurans: Functions necessary to survive ionizing radiation are also necessary to survive prolonged desiccation. J. Bacteriol. 1996, 178, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Schellhorn, H.E. Induction of resistance to hydrogen peroxide and radiation in Deinococcus radiodurans. Can. J. Microbiol. 1995, 41, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Bauermeister, A.; Moeller, R.; Reitz, G.; Sommer, S.; Rettberg, P. Effect of relative humidity on Deinococcus radiodurans’ resistance to prolonged desiccation, heat, ionizing, germicidal, and environmentally relevant UV radiation. Microb. Ecol. 2011, 61, 715–722. [Google Scholar] [CrossRef]
- Slade, D.; Radman, M. Oxidative stress resistance in Deinococcus radiodurans. Microbiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef]
- Blasius, M.; Sommer, S.; Hübscher, U. Deinococcus radiodurans: What belongs to the survival kit? Crit. Rev. Biochem. Mol. Biol. 2008, 43, 221–238. [Google Scholar] [CrossRef]
- Harada, K.; Oda, S. Induction of thermotolerance by split-dose hyperthermia at 52 °C in Deinococcus radiodurans. Agric. Biol. Chem. 1988, 52, 2391–2396. [Google Scholar]
- Schmid, A.K.; Lidstrom, M.E. Involvement of two putative alternative sigma factors in stress response of the radioresistant bacterium Deinococcus radiodurans. J. Bacteriol. 2002, 184, 6182–6189. [Google Scholar] [CrossRef]
- Schmid, A.K.; Howell, H.A.; Battista, J.R.; Peterson, S.N.; Lidstrom, M.E. Global transcriptional and proteomic analysis of the Sig1 heat shock regulon of Deinococcus radiodurans. J. Bacteriol. 2005, 187, 3339–3351. [Google Scholar] [CrossRef]
- Schmid, A.K.; Howell, H.A.; Battista, J.R.; Peterson, S.N.; Lidstrom, M.E. HspR is a global negative regulator of heat shock gene expression in Deinococcus radiodurans. Mol. Microbiol. 2005, 55, 1579–1590. [Google Scholar] [CrossRef] [PubMed]
- Airo, A.; Chan, S.L.; Martinez, Z.; Platt, M.O.; Trent, J.D. Heat shock and cold shock in Deinococcus radiodurans. Cell Biochem. Biophys. 2004, 40, 277–288. [Google Scholar] [CrossRef]
- Schmid, A.K.; Lipton, M.S.; Mottaz, H.; Monroe, M.E.; Smith, R.D.; Lidstrom, M.E. Global whole-cell FTICR mass spectrometric proteomics analysis of the heat shock response in the radioresistant bacterium Deinococcus radiodurans. J. Proteome Res. 2005, 4, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Bauermeister, A.; Hahn, C.; Rettberg, P.; Reitz, G.; Moeller, R. Roles of DNA repair and membrane integrity in heat resistance of Deinococcus radiodurans. Arch. Microbiol. 2012, 194, 959–966. [Google Scholar] [CrossRef]
- Bepperling, A.; Alte, F.; Kriehuber, T.; Braun, N.; Weinkauf, S.; Groll, M.; Haslbeck, M.; Buchner, J. Alternative bacterial two-component small heat shock protein systems. Proc. Natl. Acad. Sci. USA 2012, 109, 20407–20412. [Google Scholar] [CrossRef] [Green Version]
- Meyer, L.; Coste, G.; Sommer, S.; Oberto, J.; Confalonieri, F.; Servant, P.; Pasternak, C. DdrI, a cAMP receptor protein family member, acts as a major regulator for adaptation of Deinococcus radiodurans to various stresses. J. Bacteriol. 2018, 13, e00129-18. [Google Scholar] [CrossRef]
- Lesley, S.A.; Graziano, J.; Cho, C.Y.; Knuth, M.W.; Klock, H.E. Gene expression response to misfolded protein as a screen for soluble recombinant protein. Protein Eng. 2002, 15, 153–160. [Google Scholar] [CrossRef]
- Lim, B.; Gross, C.A. Cellular response to heat shock and cold shock. In Bacterial Stress Responses; ASM Press: Washington, DC, USA, 2011; pp. 93–114. [Google Scholar]
- Ventura, M.; Canchaya, C.; Zhang, Z.; Bernini, V.; Fitzgerald, G.F.; Van Sinderen, D. How high G+C Gram-positive bacteria and in particular bifidobacteria cope with heat stress: Protein players and regulators. FEMS Microbiol. Rev. 2006, 30, 734–759. [Google Scholar] [CrossRef]
- Arsène, F.; Tomoyasu, T. The heat shock response of Escherichia coli. Int. J. Food 2000, 55, 3–9. [Google Scholar] [CrossRef]
- Chastanet, A.; Fert, J.; Msadek, T. Comparative genomics reveal novel heat shock regulatory mechanisms in Staphylococcus aureus and other Gram-positive bacteria. Mol. Microbiol. 2003, 47, 1061–1073. [Google Scholar] [CrossRef]
- Wick, L.M.; Egli, T. Molecular components of physiological stress responses in Escherichia coli. Adv. Biochem. Eng. Biotechnol. 2004, 89, 1–45. [Google Scholar] [PubMed]
- Gunasekera, T.S.; Csonka, L.N.; Paliy, O. Genome-wide transcriptional responses of Escherichia coli K-12 to continuous osmotic and heat stresses. J. Bacteriol. 2008, 190, 3712–3720. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.L.; Roberts, C.; Disz, T.; Vonstein, V.; Hwang, K.; Overbeek, R.; Olson, P.D.; Projan, S.J.; Dunman, P.M. Characterization of the Staphylococcus aureus heat shock, cold shock, stringent, and SOS responses and their effects on log-phase mRNA turnover. J. Bacteriol. 2006, 188, 6739–6756. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.K.; Li, Z.; Lu, A.Y.; Sun, F.; Chen, S.; Rothe, M.; Menzel, R.; Sun, F.; Horvitz, H.R. Acyl-CoA dehydrogenase drives heat adaptation by sequestering fatty acids. Cell 2015, 161, 1152–1163. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Zhang, L.; Hao, F.; Zhang, J.; Wang, Y.; Tang, H. Global metabolomic responses of Escherichia coli to heat stress. J. Proteome Res. 2012, 11, 2559–2566. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury Paul, S.; Jain, P.; Mitra, J.; Dutta, S.; Bhattacharya, P.; Bal, B.; Bhattacharyya, D.; Das Gupta, S.; Pal, S. Induction of Cr(VI) reduction activity in an Anoxybacillus strain under heat stress: A biochemical and proteomic study. FEMS Microbiol. Lett. 2012, 331, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.-G.; Priya, K.; Chang, C.-Y.; Abdul Rahman, A.Y.; Tee, K.K.; Yin, W.-F. Transcriptome analysis of Pseudomonas aeruginosa PAO1 grown at both body and elevated temperatures. PeerJ 2016, 4, e2223. [Google Scholar] [CrossRef]
- Gomide, A.C.P.; de Sá, P.G.; Cavalcante, A.L.Q.; de Jesus Sousa, T.; Gomes, L.G.R.; Ramos, R.T.J.; Azevedo, V.; Silva, A.; Folador, A.R.C. Heat shock stress: Profile of differential expression in Corynebacterium pseudotuberculosis biovar Equi. Gene 2018, 645, 124–130. [Google Scholar] [CrossRef]
- Tsai, C.-H.; Liao, R.; Chou, B.; Contreras, L.M. Transcriptional analysis of Deinococcus radiodurans reveals novel small RNAs that are differentially expressed under ionizing radiation. Appl. Environ. Microbiol. 2015, 81, 1754–1764. [Google Scholar] [CrossRef]
- Tanaka, M.; Earl, A.M.; Howell, H.A.; Park, M.J.; Eisen, J.A.; Peterson, S.N.; Battista, J.R. Analysis of Deinococcus radiodurans’s transcriptional response to ionizing radiation and desiccation reveals novel proteins that contribute to extreme radioresistance. Genetics 2004, 168, 21–33. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, J.; Omelchenko, M.V.; Beliaev, A.S.; Venkateswaran, A.; Stair, J.; Wu, L.; Thompson, D.K.; Xu, D.; Rogozin, I.B.; et al. Transcriptome dynamics of Deinococcus radiodurans recovering from ionizing radiation. Proc. Natl. Acad. Sci. USA 2003, 100, 4191–4196. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- López-Leal, G.; Tabche, M.L.; Castillo-Ramírez, S.; Mendoza-Vargas, A.; Ramírez-Romero, M.A.; Dávila, G. RNA-Seq analysis of the multipartite genome of Rhizobium etli CE3 shows different replicon contributions under heat and saline shock. BMC Genom. 2014, 15, 770. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, L.; Huang, S.; Liu, J.; Ren, X.; Tian, X.; Qiao, J.; Zhang, W. RNA-seq based identification and mutant validation of gene targets related to ethanol resistance in cyanobacterial Synechocystis sp. PCC 6803. Biotechnol. Biofuels 2012, 5, 89. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S. The heat-shock response. Annu. Rev. Biochem. 1986, 55, 1151–1191. [Google Scholar] [CrossRef] [PubMed]
- Guyot, S.; Pottier, L.; Ferret, E.; Gal, L.; Gervais, P. Physiological responses of Escherichia coli exposed to different heat-stress kinetics. Arch. Microbiol. 2010, 8, 651–661. [Google Scholar] [CrossRef]
- Bruhn-Olszewska, B.; Szczepaniak, P.; Matuszewska, E.; Kuczyńska-Wiśnik, D.; Stojowska-Swędrzyńska, K.; Moruno Algara, M.; Laskowska, E. Physiologically distinct subpopulations formed in Escherichia coli cultures in response to heat shock. Microbiol. Res. 2018, 209, 33–42. [Google Scholar] [CrossRef]
- Baneyx, F.; Mujacic, M. Recombinant protein folding and misfolding in Escherichia coli. Nat. Biotechnol. 2004, 11, 1399. [Google Scholar] [CrossRef]
- Parsell, D.A.; Lindquist, S. The function of heat-Shock proteins in stress tolerance: degradation and reactivation of damaged proteins. Annu. Rev. Genet. 1993, 1, 437–496. [Google Scholar] [CrossRef]
- Schumann, W. Regulation of bacterial heat shock stimulons. Cell Stress Chaperones 2016, 21, 959–968. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, A.; Nomura, K.; Ohtomo, R.; Kato, J.; Ikeda, T.; Takiguchi, N.; Ohtake, H.; Kornberg, A. Role of inorganic polyphosphate in promoting ribosomal protein degradation by the Lon protease in E. coli. Science 2001, 293, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Servant, P.; Jolivet, E.; Bentchikou, E.; Mennecier, S.; Bailone, A.; Sommer, S. The ClpPX protease is required for radioresistance and regulates cell division after γ-irradiation in Deinococcus radiodurans. Mol. Microbiol. 2007, 66, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Matuszewska, M.; Kuczyńska-Wiśnik, D.; Laskowska, E.; Liberek, K. The small heat shock protein IbpA of Escherichia coli cooperates with IbpB in stabilization of thermally aggregated proteins in a disaggregation competent state. J. Biol. Chem. 2005, 280, 12292–12298. [Google Scholar] [CrossRef] [PubMed]
- Kuczyńska-Wiśnik, D.; Kȩdzierska, S.; Matuszewska, E.; Lund, P.; Taylor, A.; Lipińska, B.; Laskowska, E. The Escherichia coli small heat-shock proteins IbpA and IbpB prevent the aggregation of endogenous proteins denatured in vivo during extreme heat shock. Microbiology 2002, 148, 1757–1765. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Appukuttan, D.; Lim, S. Hsp20, a small heat shock protein of Deinococcus radiodurans, confers tolerance to hydrogen peroxide in Escherichia coli. J. Microbiol. Biotechnol. 2014, 24, 1118–1122. [Google Scholar] [CrossRef]
- Azam, T.A.; Ishihama, A. Twelve species of the nucleoid-associated protein from Escherichia coli. Sequence recognition specificity and DNA binding affinity. J. Biol. Chem. 1999, 274, 33105–33113. [Google Scholar] [CrossRef]
- Dillon, S.C.; Dorman, C.J. Bacterial nucleoid-associated proteins, nucleoid structure and gene expression. Nat. Rev. Microbiol. 2010, 8, 185–195. [Google Scholar] [CrossRef]
- Browning, D.F.; Grainger, D.C.; Busby, S.J.W. Effects of nucleoid-associated proteins on bacterial chromosome structure and gene expression. Curr. Opin. Microbiol. 2010, 13, 773–780. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 4, 357–359. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, E.I.; Weng, S.; Gollub, J.; Jin, H.; Botstein, D.; Cherry, J.M.; Sherlock, G. GO::TermFinder—Open source software for accessing Gene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics 2004, 20, 3710–3715. [Google Scholar] [CrossRef] [PubMed]
- Sheng, D.; Gao, G.; Tian, B.; Xu, Z.; Zheng, Z.; Hua, Y. RecX is involved in antioxidant mechanisms of the radioresistant bacterium Deinococcus radiodurans. FEMS Microbiol. Lett. 2005, 244, 251–257. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Log2 (Fold Change) | p Value | Function Description | References |
|---|---|---|---|---|
| DR_A0081 | 13.0548 | 0.02115134 | Hypothetical protein | |
| DR_A0078 | 9.3231 | 0.01952321 | Hypothetical protein | |
| DR_2307 | 8.5243 | 1.16 × 10−5 | Multidrug-efflux transporter, putative | [10] |
| DR_0518 | 8.3853 | 0.00804687 | Hypothetical protein | |
| DR_0524 | 8.1827 | 0.00604411 | Hypothetical protein | |
| DR_A0082 | 7.8581 | 0.02528407 | Hypothetical protein | |
| DR_A0079 | 7.745 | 0.0212175 | Hypothetical protein | |
| DR_A0080 | 7.6759 | 0.0183724 | Hypothetical protein | |
| DR_A0083 | 7.4529 | 0.02027402 | Hypothetical protein | |
| DR_A0086 | 7.3763 | 0.01788636 | Hypothetical protein | |
| DR_0519 | 7.3405 | 0.00011836 | Hypothetical protein | |
| DR_A0101 | 7.2857 | 0.0161855 | Hypothetical protein | |
| DR_A0077 | 7.1326 | 0.01621666 | Hypothetical protein | |
| DR_A0075 | 6.9708 | 0.04170709 | Transposase, putative | |
| DR_A0087 | 6.5568 | 0.01613077 | Hypothetical protein | |
| DR_B0072 | 6.0839 | 5.39 × 10−6 | Salicylate monooxygenase-related protein | |
| DR_B0074 | 6.0164 | 7.87 × 10−7 | 1-Phosphofructokinase | |
| DR_B0073 | 5.8393 | 1.1 × 10−5 | PTS system, fructose-specific IIBC component | |
| DR_A0085 | 5.833 | 0.02305785 | Hypothetical protein | |
| DR_A0211 | 5.5236 | 4.38 × 10−6 | Transcriptional regulator, GntR family | |
| DR_C0023 | 5.1883 | 2.24 × 10−5 | Hypothetical protein | |
| DR_A0076 | 5.1069 | 0.02545438 | ATP-dependent target DNA activator | |
| DR_B0141 | 5.0516 | 9 × 10−5 | HicB-related protein | |
| DR_B0142 | 4.9077 | 5.11 × 10−5 | Hypothetical protein | |
| DR_0422 | 4.8592 | 0.00024562 | Trans-aconitate 2-methyltransferase | |
| DR_A0182 | 4.799 | 0.00068635 | Hypothetical protein | [13] |
| DR_0516 | 4.7289 | 0.00382339 | Hypothetical protein | |
| DR_2374 | 4.6897 | 0.0005564 | Ribonucleoside-diphosphate reductase-related protein | [13] |
| DR_0532 | 4.6735 | 0.00051046 | Hypothetical protein | |
| DR_0423 | 4.5152 | 0.00011999 | Hypothetical protein | |
| DR_A0364 | −4.3096 | 5.93 × 10−5 | Oxidoreductase, short-chain dehydrogenase/reductase family | [13] |
| DR_0201 | −4.321 | 0.00785386 | Hypothetical protein | |
| DR_0392 | −4.3342 | 0.00434331 | Hypothetical protein | [10] |
| DR_0334 | −4.3572 | 0.01241591 | Lipase, putative | |
| DR_B0038 | −4.3573 | 0.01463299 | Hypothetical protein | |
| DR_A0352 | −4.3662 | 0.00155047 | Methyl-accepting chemotaxis protein | |
| DR_2240 | −4.3942 | 0.00195473 | Hypothetical protein | |
| DR_2527 | −4.4273 | 0.00068251 | Hypothetical protein | [10] |
| DR_A0233 | −4.4361 | 1.05 × 10−5 | Oxidoreductase, iron-sulfur subunit | [13] |
| DR_1987 | −4.4998 | 3.18 × 10−6 | Hypothetical protein | |
| DR_A0231 | −4.5169 | 0.00510755 | Oxidoreductase | [13] |
| DR_1778 | −4.5277 | 0.00016505 | 3-Isopropylmalate dehydratase, large subunit | |
| DR_2263 | −4.5338 | 6.47 × 10−8 | DNA-binding stress response protein, Dps family | [10] |
| DR_2563 | −4.6247 | 0.00080366 | Hypothetical protein | |
| DR_1712 | −4.7284 | 0.00739225 | Extracellular solute-binding protein, family 5 | [13] |
| DR_A0232 | −4.7289 | 0.00081206 | Oxidoreductase | |
| DR_1277 | −4.7754 | 0.01673083 | ABC transporter, periplasmic substrate-binding protein, putative | |
| DR_1711 | −4.8264 | 0.00028517 | N-Acyl-L-amino acid amidohydrolase | |
| DR_1067 | −4.8305 | 5.2 × 10−6 | Hypothetical protein | |
| DR_0791 | −4.9522 | 0.00226901 | Chloride peroxidase, putative | |
| DR_0644 | −5.4271 | 0.00078552 | Hypothetical protein | |
| DR_2560 | −5.4566 | 0.0002672 | Hypothetical protein | |
| DR_1315 | −5.4743 | 0.00327873 | Hypothetical protein | [10,13] |
| DR_0105 | −5.4893 | 1.54 × 10−6 | Hypothetical protein | |
| DR_1066 | −5.6713 | 0.02719424 | Hypothetical protein | |
| DR_1483 | −5.8009 | 0.00071264 | Hypothetical protein | |
| DR_1790 | −5.8218 | 1.62 × 10−6 | Yellow-related protein | |
| DR_0465 | −5.8342 | 0.00052829 | Conserved hypothetical protein | |
| DR_1314 | −6.1749 | 0.00281391 | Conserved hypothetical protein | [10] |
| DR_0891 | −6.4635 | 0.00113242 | DNA-binding response regulator |
| Gene ID | Gene Name | Functions | Log2(Fold Change) | p Value |
|---|---|---|---|---|
| DR_A0101 | Hypothetical protein | 6.7807 | 0.00190534 | |
| DR_0128 | grpE | Protein turnover and chaperone function | −3.1812 | 0.00160996 |
| DR_0349 | lon | Protein turnover and chaperone function | −3.5087 | 0.00188947 |
| DR_0607 | groEL | Protein turnover and chaperone function | −3.5102 | 0.00128474 |
| DR_0129 | dnaK | Protein turnover and chaperone function | −3.9169 | 0.04542787 |
| DR_0985 | Protein turnover and chaperone function | −4.1346 | 0.00249103 | |
| DR_1046 | clpB | Protein turnover and chaperone function | −4.4467 | 0.01832036 |
| DR_0606 | groES | Protein turnover and chaperone function | −3.3489 | 0.00035202 |
| DR_0127 | DNA-binding protein | −3.0322 | 0.00135957 | |
| DR_1083 | Hypothetical protein | −3.1333 | 4.23 × 10−5 | |
| DR_1325 | LysM peptidoglycan-binding protein | −3.3623 | 0.00886153 | |
| DR_2311 | Nucleotide metabolism and transport function | −4.3949 | 9.48 × 10−5 | |
| DR_2312 | Nucleotide metabolism and transport function | −4.2404 | 1.03 × 10−6 | |
| DR_1501 | Energy production and conversion | −3.1352 | 0.00033852 | |
| DR_0693 | Membrane transport | −3.7209 | 0.00754623 | |
| DR_1042 | padR | Transcription | −3.0467 | 1.6 × 10−5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, D.; Liu, W.; Chen, Y.; Liu, Y.; Han, J.; Geng, X.; Li, J.; Jiang, S.; Zhou, Z.; Zhang, W.; et al. RNA-Seq-Based Comparative Transcriptome Analysis Highlights New Features of the Heat-Stress Response in the Extremophilic Bacterium Deinococcus radiodurans. Int. J. Mol. Sci. 2019, 20, 5603. https://doi.org/10.3390/ijms20225603
Xue D, Liu W, Chen Y, Liu Y, Han J, Geng X, Li J, Jiang S, Zhou Z, Zhang W, et al. RNA-Seq-Based Comparative Transcriptome Analysis Highlights New Features of the Heat-Stress Response in the Extremophilic Bacterium Deinococcus radiodurans. International Journal of Molecular Sciences. 2019; 20(22):5603. https://doi.org/10.3390/ijms20225603
Chicago/Turabian StyleXue, Dong, Wenzheng Liu, Yun Chen, Yingying Liu, Jiahui Han, Xiuxiu Geng, Jiang Li, Shijie Jiang, Zhengfu Zhou, Wei Zhang, and et al. 2019. "RNA-Seq-Based Comparative Transcriptome Analysis Highlights New Features of the Heat-Stress Response in the Extremophilic Bacterium Deinococcus radiodurans" International Journal of Molecular Sciences 20, no. 22: 5603. https://doi.org/10.3390/ijms20225603