Early Postnatal Treatment with Valproate Induces Gad1 Promoter Remodeling in the Brain and Reduces Apnea Episodes in Mecp2-Null Mice


Abstract
:1. Introduction
2. Results
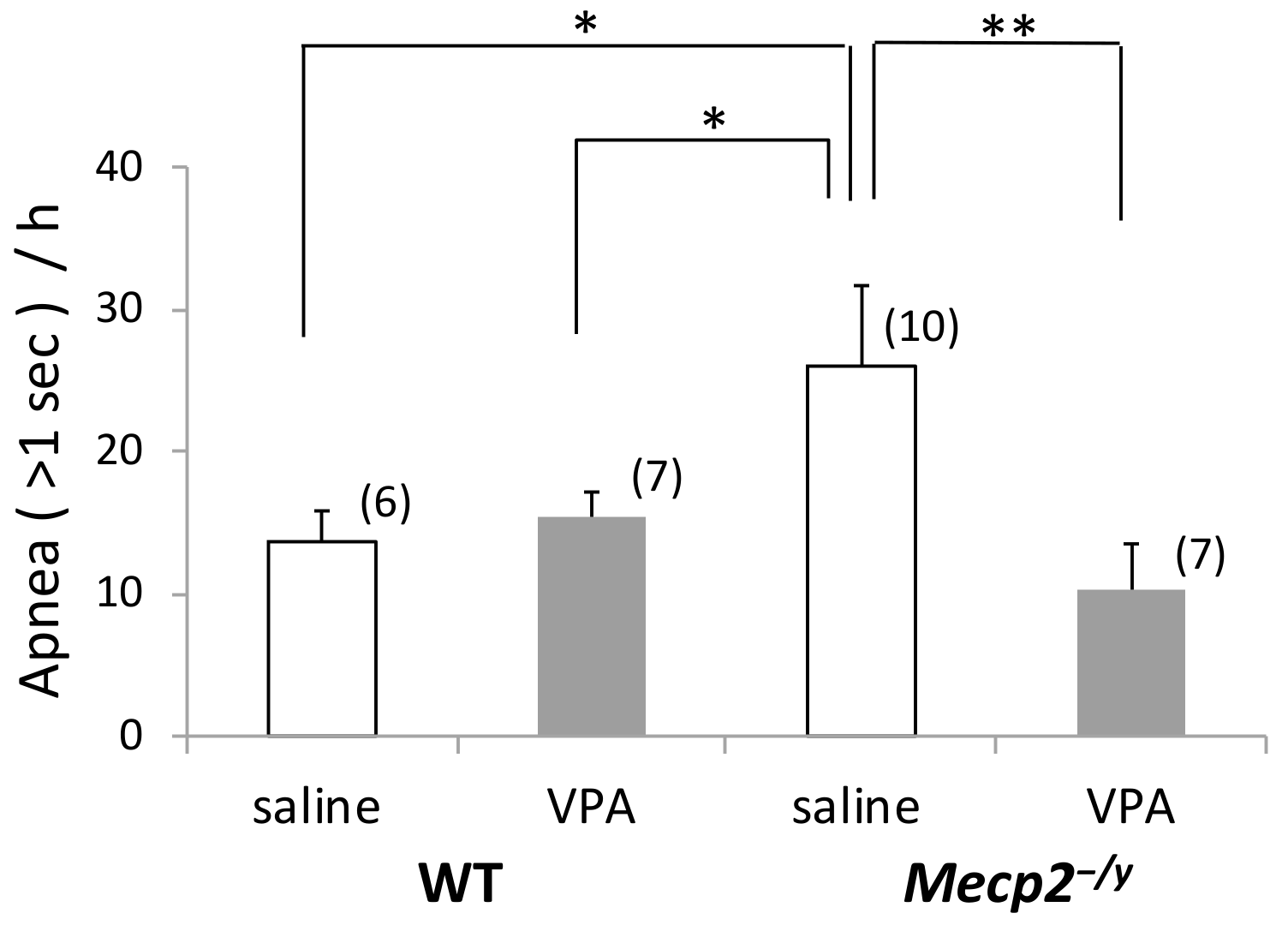
2.1. Spontaneous Apnea Counts are Greater in Mecp2-/y Mice Compared to WT Mice
2.2. VPA Treatment Reduces Apnea Counts in Mecp2-/y Mice
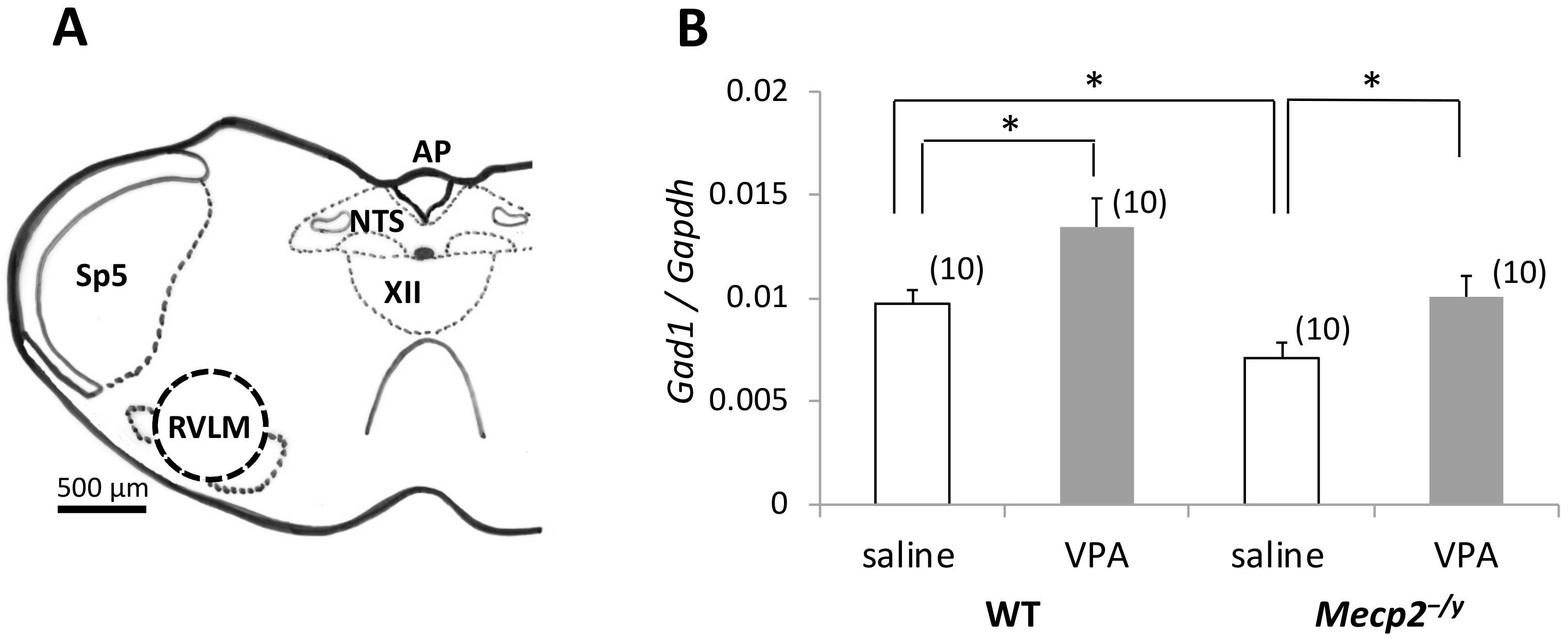
2.3. VPA Treatment Upregulates Gad1 mRNA Expression in the RVLM
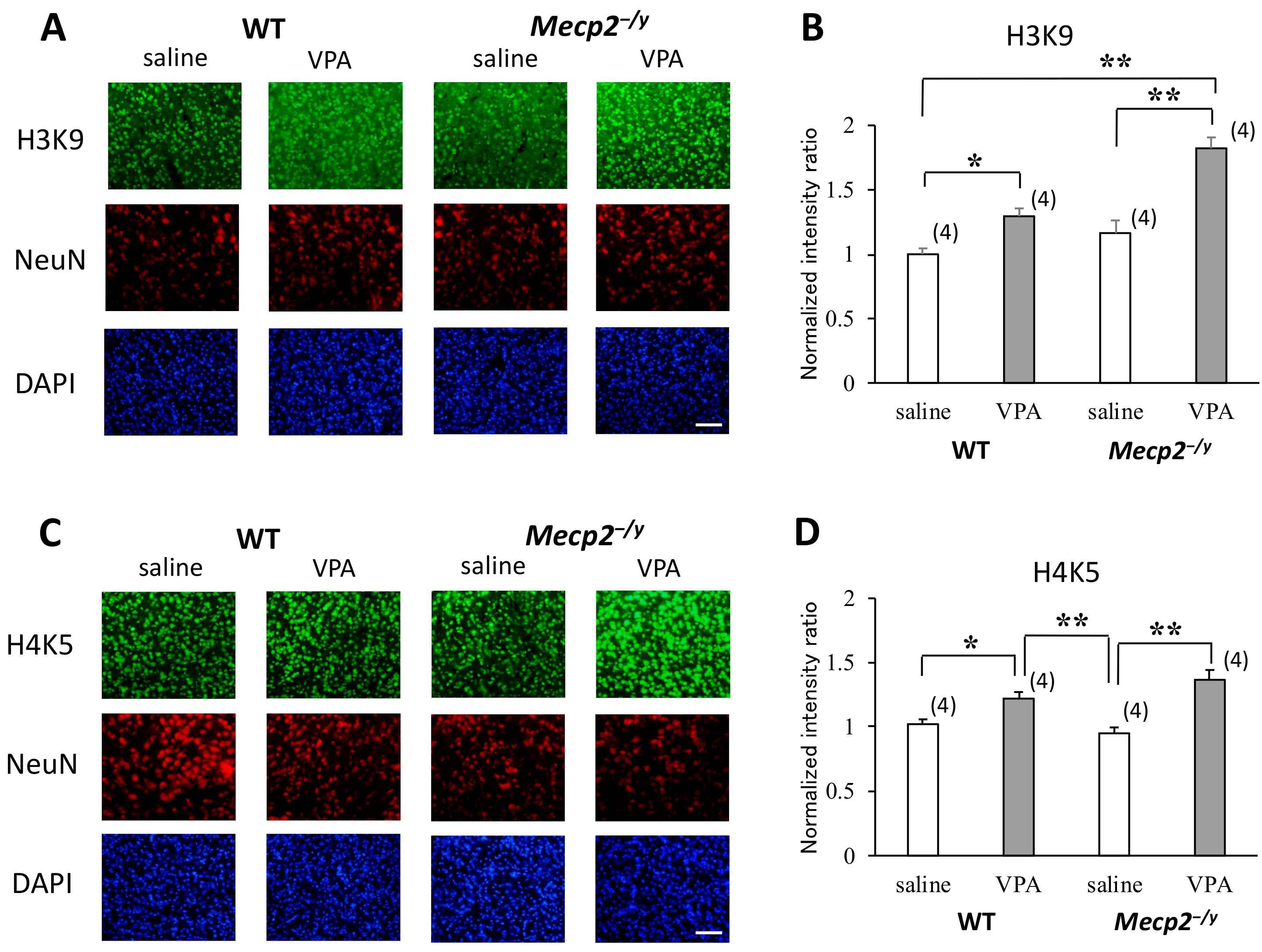
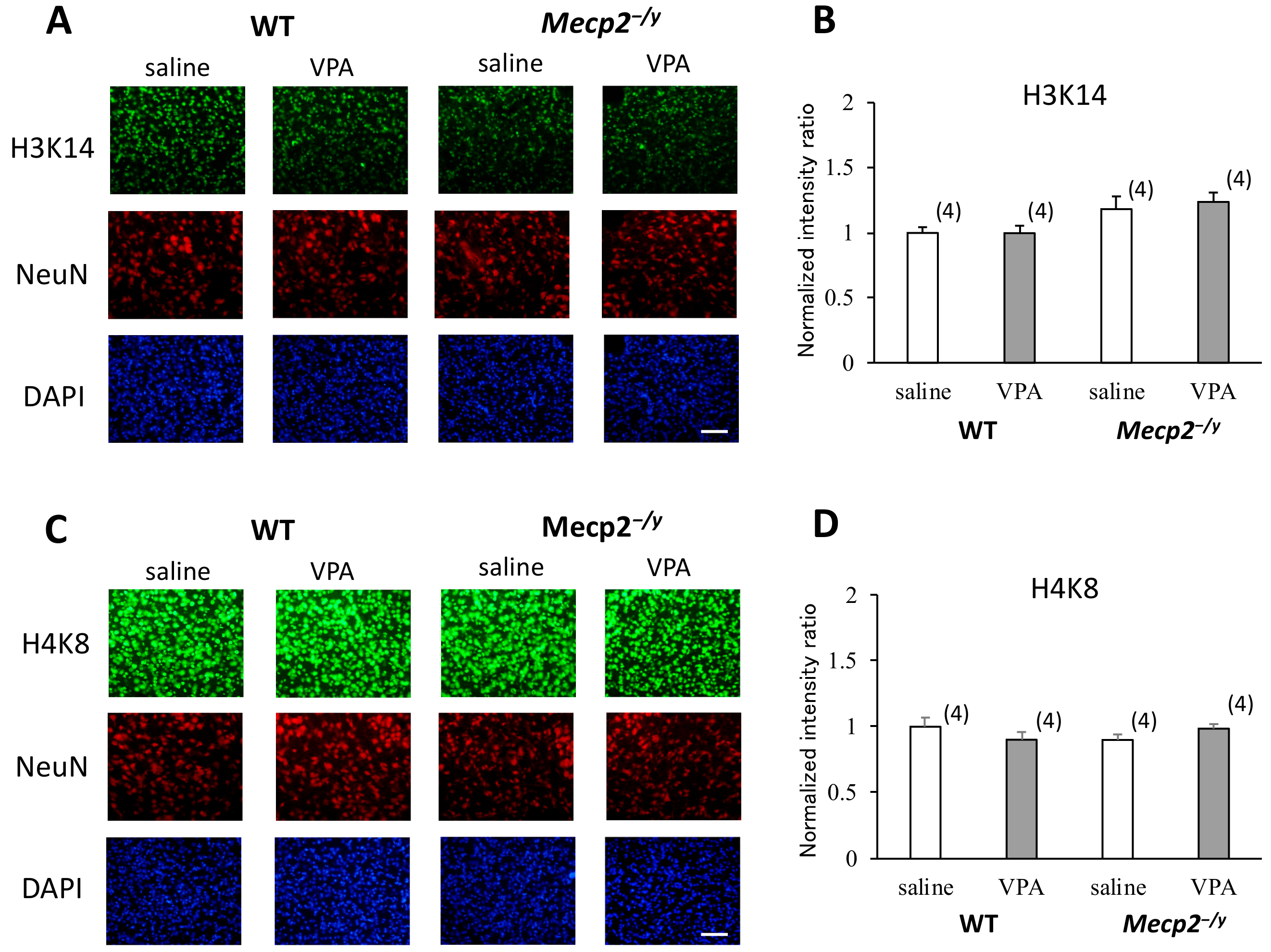
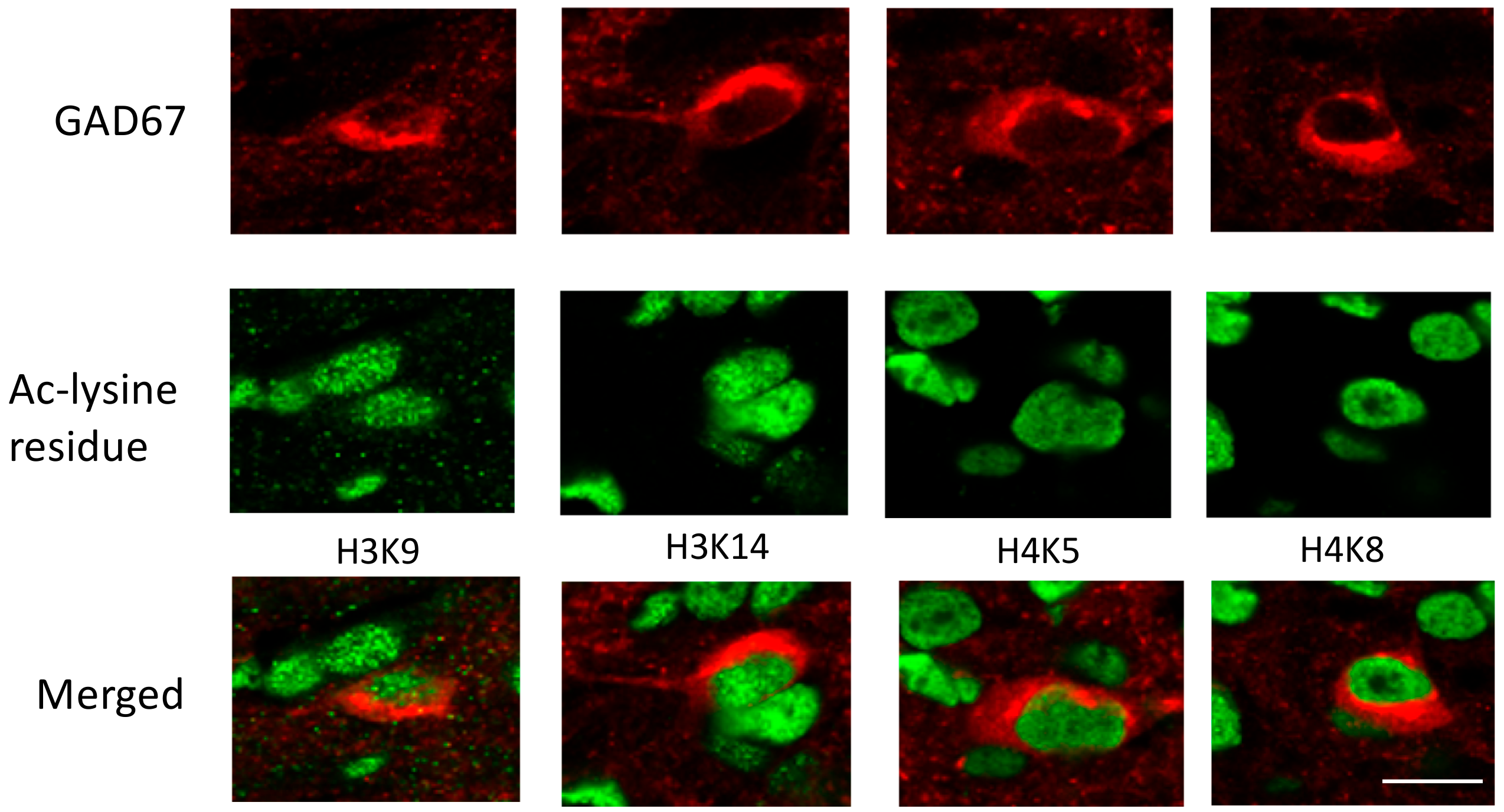
2.4. VPA Treatment Enhances Histone Acetylation in the RVLM
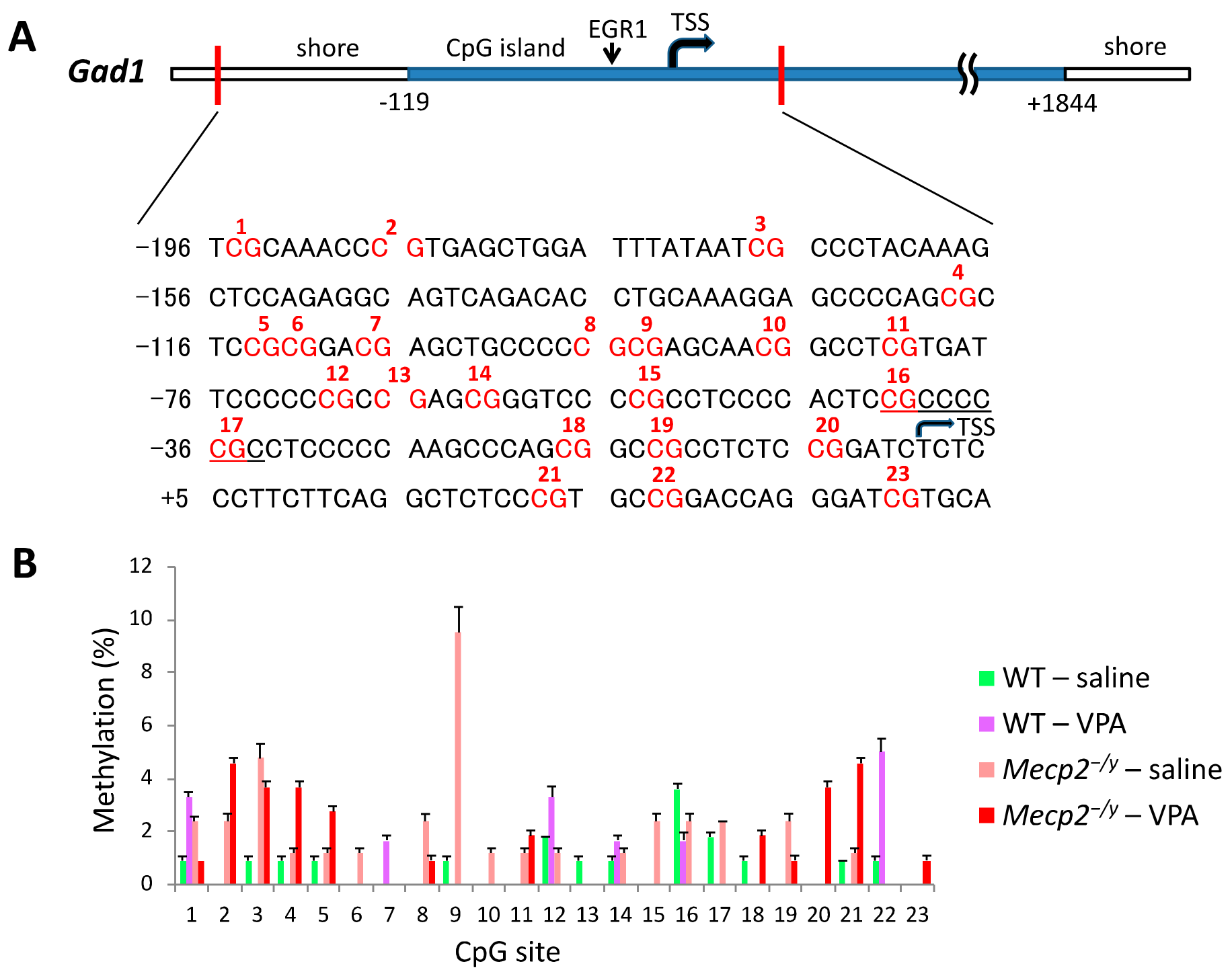
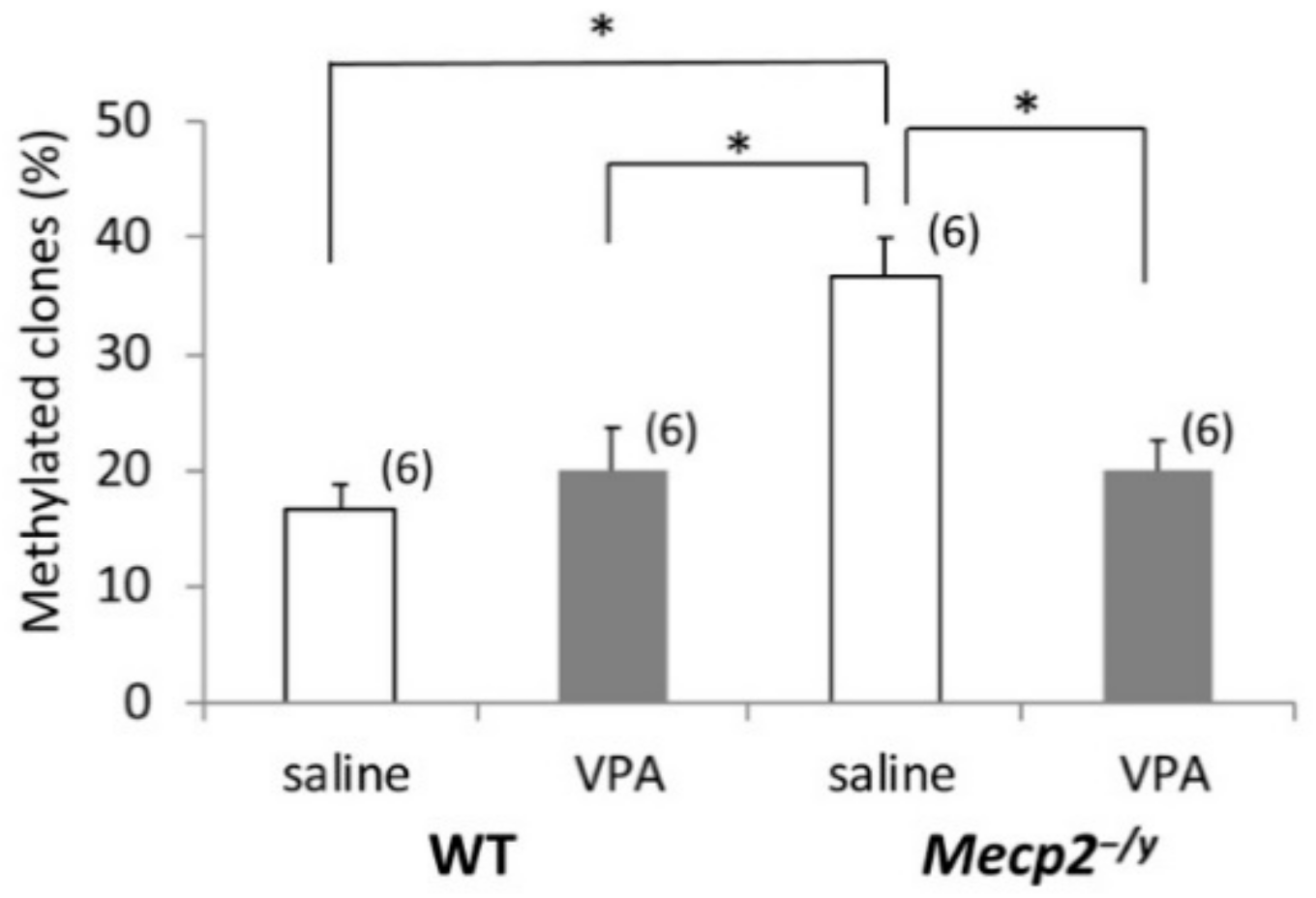
2.5. VPA Treatment Induces Gad1 Promoter Demethylation in Mecp2-/Y Mice
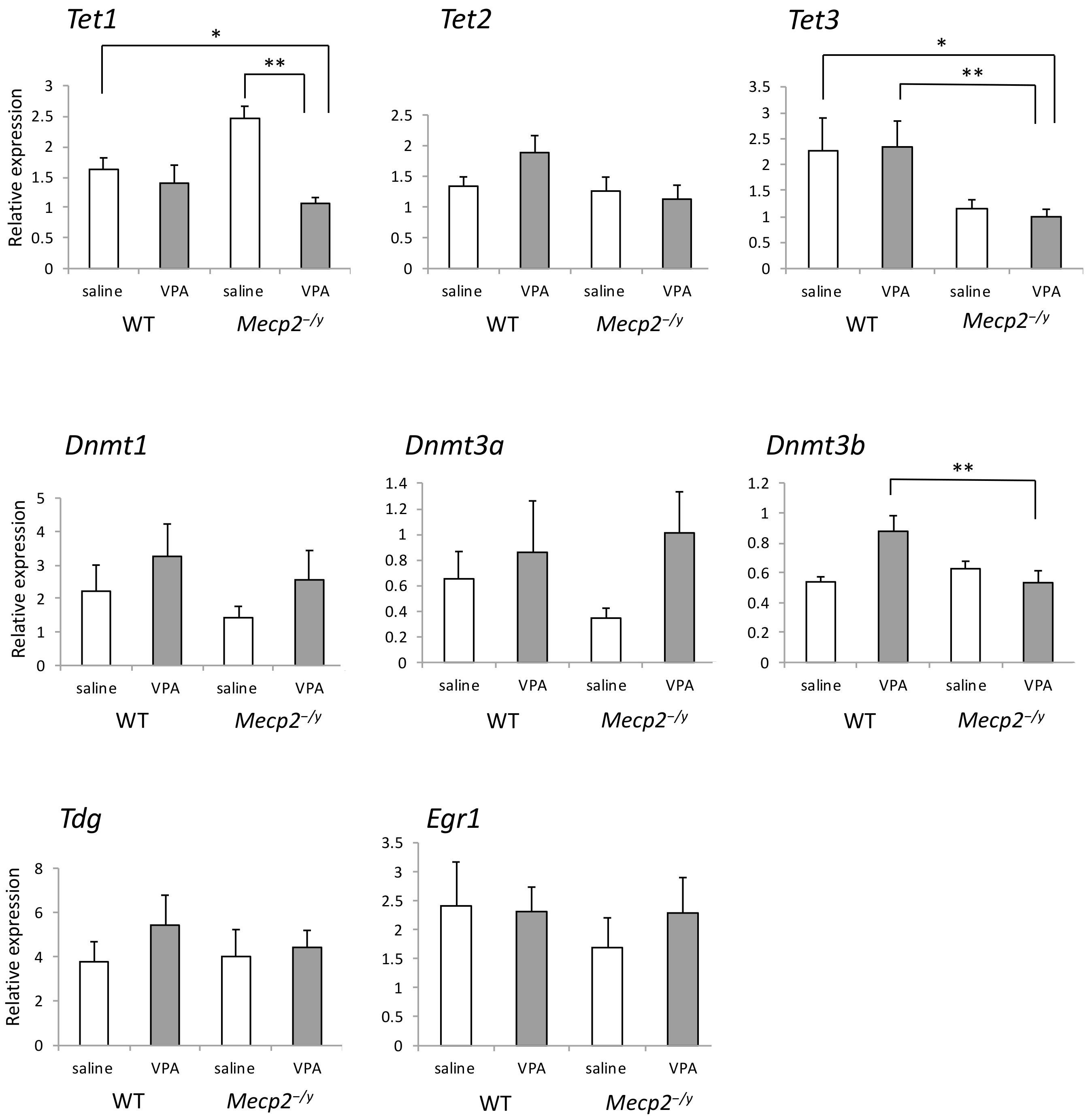
2.6. VPA Treatment Reduces Tet1 mRNA Expression in Mecp2-/Y Mice
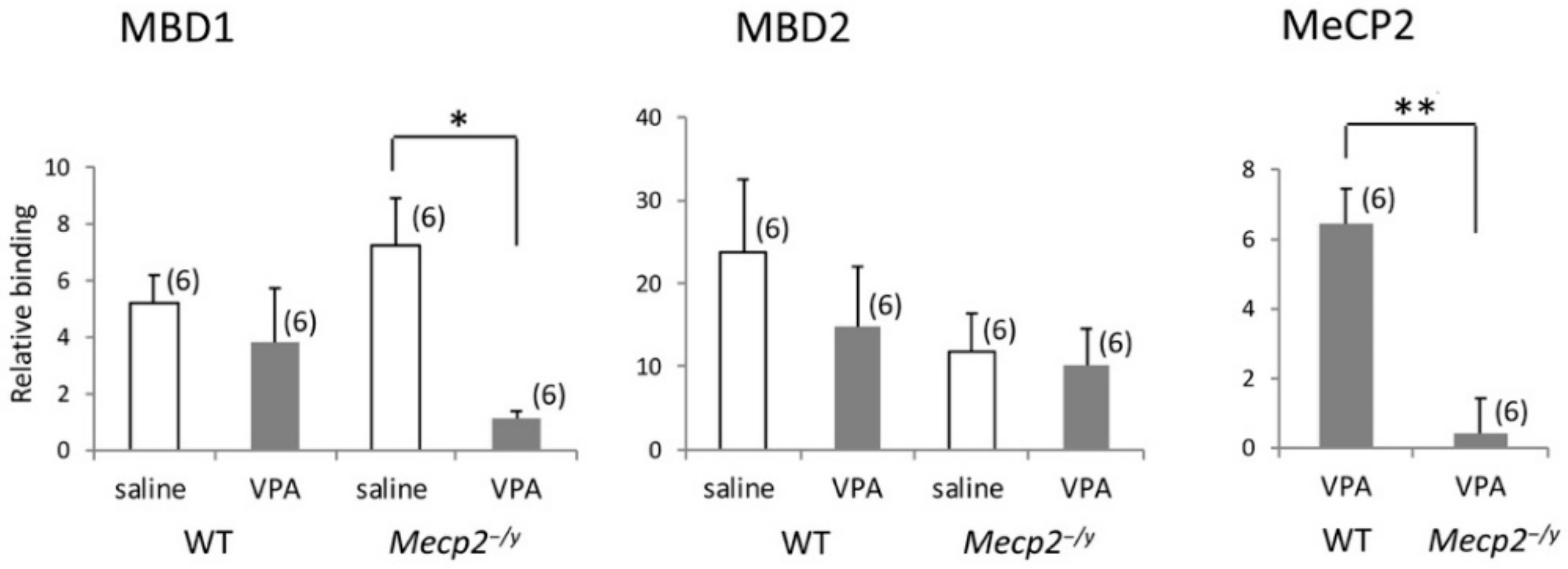
2.7. VPA Treatment Modifies Mbd1 Association with Gad1 Promoter in Mecp2-/Y Mice
3. Discussion
4. Materials and Methods
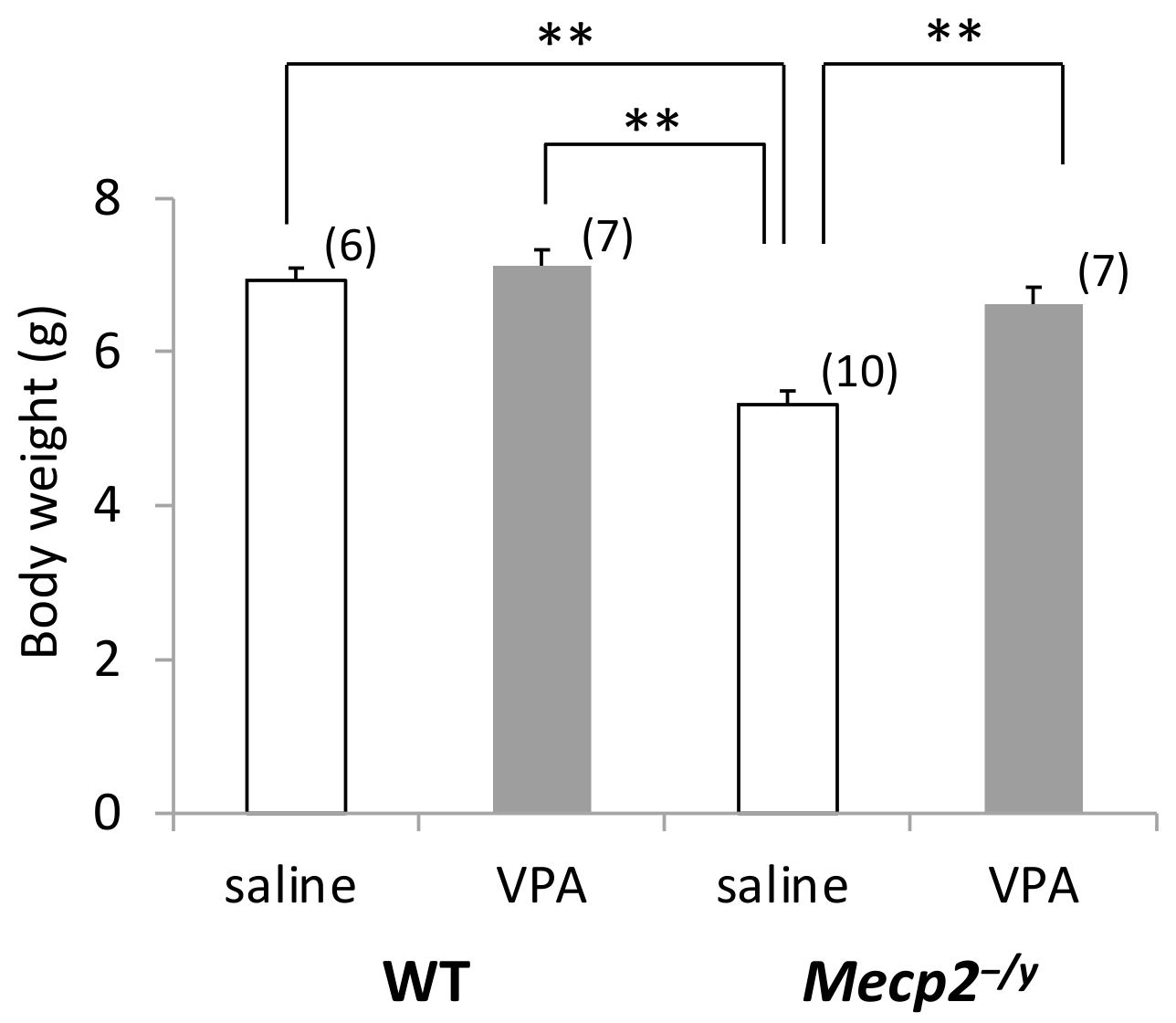
4.1. Animals
4.2. VPA Treatment
4.3. Plethysmographic Assessment of Apnea Episodes
4.4. Isolation of Total RNA and Quantitative Reverse Transcription PCR Analysis
4.5. Quantitative Immunofluorescence Labelling
4.6. Sodium Bisulfite Mapping
4.7. Chromatin Immunoprecipitation Assay
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| AP | area postrema |
| cDNA | complementary DNA |
| DNMT | DNA methyltransferase |
| EGR1 | Early growth response protein 1 |
| GABA | γ-aminobutyric acid |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| HDAC | histone deacetylase |
| MBD | Methyl-CpG binding domain protein |
| MeCP2 | methyl-CpG-binding protein 2 |
| NTS | nucleus tractus solitarius |
| PCR | polymerase chain reaction |
| RT-qPCR | quantitative reverse transcription PCR |
| RTT | Rett syndrome |
| RVLM | rostral ventrolateral medulla |
| Sp5 | spinal trigeminal nucleus |
| TDG | thymine DNA glycosylase |
| TET | ten–eleven translocation |
| TSS | transcriptional start site |
| VPA | valproate |
| XII | hypoglossal nucleus |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | Breathing Frequency (Cycles/min) | Tidal Volume (mL) | Inspiratory Time (s) | Expiratory Time (s) | Peak Inspiratory Flow (mL/s) | Peak Expiratory Flow (mL/s) | |
| WT—saline | 6 | 244.6 ± 2.4 | 0.061 ± 0.001 | 0.073 ± 0.002 | 0.156 ± 0.002 | 1.165 ± 0.005 | 1.031 ± 0.020 |
| WT—VPA | 6 | 253.0 ± 4.7 | 0.051 ± 0.002 | 0.077 ± 0.002 | 0.167 ± 0.010 | 1.106 ± 0.024 | 1.036 ± 0.070 |
| MeCP2-/y—saline | 6 | 235.6 ± 3.4 | 0.063 ± 0.004 | 0.089 ± 0.002 | 0.184 ± 0.010 | 1.126 ± 0.041 | 1.076 ± 0.044 |
| MeCP2-/y—VPA | 6 | 239.0 ± 5.6 | 0.057 ± 0.001 | 0.085 ± 0.003 | 0.167 ± 0.004 | 1.062 ± 0.045 | 1.006 ± 0.028 |
| Gene | Sequences (F; Forward, R; Reverse) | Genbank Number | |
|---|---|---|---|
| Gad1 | F | 5′-TTCTGGTACATTCCACAAAGCCTTC-3′ | NM_008077.5 |
| R | 5′-CCATGGTTGTTCCTGACTCCATC-3′ | ||
| Tet1 | F | 5′-TCAGCATGAAGTCTCAGTTGACTCC-3′ | NM_001253857.2 |
| R | 5′-GAATTGATGCCTTATTCCCACCA-3′ | ||
| Tet2 | F | 5′-GGTGCTACCCAGATTGCTCCA-3′ | NM_001040400.2 |
| R | 5′-TGGTCTAAGCCTCCACTGTTAGCTC-3′ | ||
| Tet3 | F | 5′-CTGTCCATCTCATGGAGCTTTC-3′ | NM_001347313.1 |
| R | 5′-GCGTATGCACCTCCAATGTGTTA-3′ | ||
| Dnmt1 | F | 5′-CTTCGGCAACATCCTGGACA-3′ | NM_001199431.1 |
| R | 5′-ACTGGACAGCAGGCAGAGCTTA-3′ | ||
| Dnmt3a | F | 5′-GCATACAGCTTGCTGCACTCTC-3′ | NM_153743.4 |
| R | 5′-ACCTGCTGTACGCATTGACC-3′ | ||
| Dnmt3b | F | 5′-TTGCTTTGGTACAAGGGCTGAA-3′ | NM_001003960.4 |
| R | 5′-TCATCCCTGCTGACATCATCATC-3′ | ||
| Tdg | F | 5′-TAGGAAACGTGCGTGTTCAG-3′ | NM_001358517.1 |
| R | 5′-CTCATACTGCCAAACCAGCA-3′ | ||
| Egr1 | F | 5′-TCAGTGGCCACCACCTTTG-3′ | NM_007913.5 |
| R | 5′-AAAGGTCGCTGTCATGTCTGAA-3′ | ||
| Gapdh | F | 5′-TGTGTCCGTCGTGGATCTGA-3′ | NM_008084.3 |
| R | 5′-TTGCTGTTGAAGTCGCAGGAC-3′ |
| No. of Mice Belonging to Each Group | No. of Mothers that Raised Mice Belonging to Each Group | Results of A Two-Factor ANOVA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Genes | WT-saline | WT-VPA | Mecp2-/y-saline | Mecp2-/y-VPA | WT-saline | WT-VPA | Mecp2-/y-saline | Mecp2-/y-VPA | geno-type | treat-ment | inter-action |
| Tet1 | 6 | 6 | 6 | 6 | 5 | 5 | 6 | 6 | n.s. | p < 0.01 | p < 0.05 |
| Tet2 | 6 | 6 | 6 | 6 | 5 | 5 | 6 | 6 | n.s. | n.s. | n.s. |
| Tet3 | 6 | 6 | 6 | 6 | 5 | 5 | 6 | 6 | p < 0.01 | n.s. | n.s. |
| Dnmt1 | 8 | 8 | 8 | 8 | 7 | 7 | 7 | 7 | n.s. | n.s. | n.s. |
| Dnmt3a | 11 | 12 | 10 | 12 | 10 | 10 | 10 | 11 | n.s. | n.s. | n.s. |
| Dnmt3b | 6 | 6 | 6 | 6 | 5 | 5 | 6 | 6 | n.s. | n.s. | p < 0.01 |
| Tdg | 12 | 11 | 10 | 10 | 10 | 10 | 10 | 10 | n.s. | n.s. | n.s. |
| Egr1 | 12 | 11 | 12 | 12 | 10 | 10 | 10 | 11 | n.s. | n.s. | n.s. |



References
- Amir, R.E.; van den Veyver, I.B.; Wan, M.; Tran, C.Q.; Francke, U.; Zoghbi, H.Y. Rett syndrome is caused by mutations in X-linked MECP2, encoding methyl-CpG-binding protein 2. Nat. Genet. 1999, 23, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Percy, A.K. Rett syndrome. Current status and new vistas. Neurol. Clin. 2002, 20, 1125–1141. [Google Scholar] [CrossRef]
- Zoghbi, H.Y. Postnatal neurodevelopmental disorders: Meeting at the synapse? Science 2003, 302, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, B.; Aicardi, J.; Dias, K.; Ramos, O. A progressive syndrome of autism, dementia, ataxia, and loss of purposeful hand use in girls: Rett’s syndrome: Report of 35 cases. Ann. Neurol. 1983, 14, 471–479. [Google Scholar] [CrossRef]
- Armstrong, D.D. Review of Rett syndrome. J. Neuropathol. Exp. Neurol. 1997, 56, 843–849. [Google Scholar] [CrossRef]
- Chahrour, M.; Zoghbi, H.Y. The story of Rett syndrome: From clinic to neurobiology. Neuron 2007, 56, 422–437. [Google Scholar] [CrossRef]
- Katz, D.M.; Dutschmann, M.; Ramirez, J.M.; Hilaire, G. Breathing disorders in Rett syndrome: Progressive neurochemical dysfunction in the respiratory network after birth. Respir. Physiol. Neurobiol. 2009, 168, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Guy, J.; Hendrich, B.; Holmes, M.; Martin, J.E.; Bird, A. A mouse Mecp2-null mutation causes neurological symptoms that mimic Rett syndrome. Nat. Genet. 2001, 27, 322–326. [Google Scholar]
- Chen, R.Z.; Akbarian, S.; Tudor, M.; Jaenisch, R. Deficiency of methyl-CpG binding protein-2 in CNS neurons results in a Rett-like phenotype in mice. Nat. Genet. 2001, 27, 327–331. [Google Scholar]
- Viemari, J.C.; Roux, J.C.; Tryba, A.K.; Saywell, V.; Burnet, H.; Peña, F.; Zanella, S.; Bévengut, M.; Barthelemy-Requin, M.; Herzing, L.B.; et al. Mecp2 deficiency disrupts norepinephrine and respiratory systems in mice. J. Neurosci. 2005, 25, 11521–11530. [Google Scholar] [CrossRef]
- Roux, J.C.; Dura, E.; Moncla, A.; Mancini, J.; Villard, L. Treatment with desipramine improves breathing and survival in a mouse model for Rett syndrome. Eur J. Neurosci. 2007, 7, 1915–1922. [Google Scholar]
- Ogier, M.; Katz, D.M. Breathing dysfunction in Rett syndrome: Understanding epigenetic regulation of the respiratory network. Respir. Physiol. Neurobiol. 2008, 164, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, M.; Silva-Fernandes, A.; Oliveira, P.; Sousa, N.; Maciel, P. Evidence for abnormal early development in a mouse model of Rett syndrome. Genes Brain Behav. 2007, 6, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Medrihan, L.; Tantalaki, E.; Aramuni, G.; Sargsyan, V.; Dudanova, I.; Missler, M.; Zhang, W. Early defects of GABAergic synapses in the brain stem of a MeCP2 mouse model of Rett syndrome. J. Neurophysiol. 2008, 99, 112–121. [Google Scholar] [CrossRef]
- Koch-Weser, J.; Browne, T.R. Drug therapy: Valproate. N. Engl. J. Med. 1980, 302, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, K.; Czapp, M.; Löscher, W. Increase in antiepileptic efficacy during prolonged treatment with valproate: Role of inhibition of histone deacetylases? Epilepsy Res. 2008, 81, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Phiel, C.J.; Zhang, F.; Huang, E.Y.; Guenther, M.G.; Lazar, M.A.; Klein, P.S. Histone deacetylase is a direct target of valproate, a potent anticonvulsant, mood stabilizer, and teratogen. J. Biol. Chem. 2001, 276, 36734–36741. [Google Scholar] [CrossRef] [PubMed]
- Jian, L.; Nagarajan, L.; de Klerk, N.; Ravine, D.; Bower, C.; Anderson, A.; Williamson, S.; Christodoulou, J.; Leonard, H. Predictors of seizure onset in Rett syndrome. J. Pediatr. 2006, 149, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Pintaudi, M.; Calevo, M.G.; Vignoli, A.; Baglietto, M.G.; Hayek, Y.; Traverso, M.; Giacomini, T.; Giordano, L.; Renieri, A.; Russo, S.; et al. Antiepileptic drugs in Rett Syndrome. Eur. J. Paediatr. Neurol. 2015, 19, 446–452. [Google Scholar] [CrossRef]
- Tremolizzo, L.; Carboni, G.; Ruzicka, W.B.; Mitchell, C.P.; Sugaya, I.; Tueting, P.; Sharma, R.; Grayson, D.R.; Costa, E.; Guidotti, A. An epigenetic mouse model for molecular and behavioral neuropathologies related to schizophrenia vulnerability. Proc. Natl. Acad. Sci. USA 2002, 99, 17095–17100. [Google Scholar] [CrossRef] [Green Version]
- Trifonov, S.; Yamashita, Y.; Kase, M.; Maruyama, M.; Sugimoto, T. Glutamate decarboxylase 1 alternative splicing isoforms: Characterization, expression and quantification in the mouse brain. BMC Neurosci. 2014, 15, 114. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, M.; Hayatsu, H. High-speed conversion of cytosine to uracil in bisulfite genomic sequencing analysis of DNA methylation. DNA Res. 2004, 11, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Y.; Hellstrom, I.C.; Bagot, R.C.; Wen, X.; Diorio, J.; Meaney, M.J. Maternal care and DNA methylation of a glutamate decarboxylase 1 promoter in rat hippocampus. J. Neurosci. 2010, 29, 13130–13137. [Google Scholar] [CrossRef] [PubMed]
- Labouesse, M.A.; Dong, E.; Grayson, D.R.; Guidotti, A.; Meyer, U. Maternal immune activation induces GAD1 and GAD2 promoter remodeling in the offspring prefrontal cortex. Epigenetics 2015, 10, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Uchida, T.; Furukawa, T.; Iwata, S.; Yanagawa, Y.; Fukuda, A. Selective loss of parvalbumin-positive GABAergic interneurons in the cerebral cortex of maternally stressed Gad1-heterozygous mouse offspring. Transl. Psychiatry 2014, 4, e371. [Google Scholar] [CrossRef] [PubMed]
- Szabó, G.; Katarova, Z.; Körtvély, E.; Greenspan, R.J.; Urbán, Z. Structure and the promoter region of the mouse gene encoding the 67-kD form of glutamate decarboxylase. DNA Cell Biol. 1996, 15, 1081–1091. [Google Scholar] [CrossRef]
- Yanagawa, Y.; Kobayashi, T.; Kamei, T.; Ishii, K.; Nishijima, M.; Takaku, A.; Tamura, S. Structure and alternative promoters of the mouse glutamate decarboxylase 67 gene. Biochem. J. 1997, 326, 573–578. [Google Scholar] [CrossRef]
- Chattopadhyaya, B.; Di Cristo, G.; Wu, C.Z.; Knott, G.; Kuhlman, S.; Fu, Y.; Palmiter, R.D.; Huang, Z.J. GAD67-mediated GABA synthesis and signaling regulate inhibitory synaptic innervation in the visual cortex. Neuron 2007, 54, 889–903. [Google Scholar] [CrossRef]
- Tsunekawa, N.; Arata, A.; Obata, K. Development of spontaneous mouth/tongue movement and related neural activity, and their repression in fetal mice lacking glutamate decarboxylase 67. Eur. J. Neurosci. 2005, 21, 173–178. [Google Scholar] [CrossRef]
- Fujii, M.; Arata, A.; Kanbara-Kume, N.; Saito, K.; Yanagawa, Y.; Obata, K. Respiratory activity in brainstem of fetal mice lacking glutamate decarboxylase 65/67 and vesicular GABA transporter. Neuroscience 2007, 146, 1044–1052. [Google Scholar] [CrossRef]
- Chao, H.T.; Chen, H.; Samaco, R.C.; Xue, M.; Chahrour, M.; Yoo, J.; Neul, J.L.; Gong, S.; Lu, H.C.; Heintz, N.; et al. Dysfunction in GABA signalling mediates autism-like stereotypies and Rett syndrome phenotypes. Nature 2010, 468, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.W.; Kochukov, M.Y.; Ward, C.S.; Merritt, J.; Thomas, K.; Nguyen, T.; Arenkiel, B.R.; Neul, J.L. Progressive changes in a distributed neural circuit underlie breathing abnormalities in mice lacking MeCP2. J. Neurosci. 2016, 36, 5572–5586. [Google Scholar] [CrossRef] [PubMed]
- Stettner, G.M.; Huppke, P.; Brendel, C.; Richter, D.W.; Gartner, J.; Dutschmann, M. Breathing dysfunctions associated with impaired control of postinspiratory activity in Mecp2−/y knockout mice. J. Physiol. 2007, 579, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Voituron, N.; Zanella, S.; Menuet, C.; Dutschmann, M.; Hilaire, G. Early breathing defects after moderate hypoxia or hypercapnia in a mouse model of Rett syndrome. Respir. Physiol. Neurobiol. 2009, 168, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Ure, K.; Lu, H.; Wang, W.; Ito-Ishida, A.; Wu, Z.; He, L.; Sztainberg, Y.; Chen, W.; Tang, J.; Zoghbi, H.Y. Restoration of Mecp2 expression in GABAergic neurons is sufficient to rescue multiple disease features in a mouse model of Rett syndrome. eLife 2016, 5, e14198. [Google Scholar] [CrossRef]
- Shirakawa, T.; Honma, S.; Katsuno, Y.; Oguchi, H.; Honma, K.-I. Synchronization of circadian firing rhythms in cultured rat suprachiasmatic neurons. Eur. J. Neurosci. 2000, 12, 2833–2838. [Google Scholar] [CrossRef]
- Jin, X.; Cui, N.; Zhong, W.; Jin, X.T.; Jiang, C. GABAergic synaptic inputs of locus coeruleus neurons in wild-type and Mecp2-null mice. Am. J. Physiol. Cell Physiol. 2013, 304, C844–C857. [Google Scholar] [CrossRef]
- El-Khoury, R.; Panayotis, N.; Matagne, V.; Ghata, A.; Villard, L.; Roux, J.C. GABA and glutamate pathways are spatially and developmentally affected in the brain of Mecp2-deficient mice. PLoS ONE 2014, 9, e92169. [Google Scholar] [CrossRef]
- Zhong, W.; Johnson, C.M.; Wu, Y.; Cui, N.; Xing, H.; Zhang, S.; Jiang, C. Effects of early-life exposure to THIP on phenotype development in a mouse model of Rett syndrome. J. Neurodev. Disord. 2016, 8, 37. [Google Scholar] [CrossRef]
- Takei, H.; Yamamoto, K.; Bae, Y.C.; Shirakawa, T.; Kobayashi, M. Histamine H3 Heteroreceptors Suppress Glutamatergic and GABAergic Synaptic Transmission in the Rat Insular Cortex. Front. Neural Circuits 2017, 11, 85. [Google Scholar] [CrossRef]
- Chen, C.Y.; Di Lucente, J.; Lin, Y.C.; Lien, C.C.; Rogawski, M.A.; Maezawa, I.; Jin, L.W. Defective GABAergic neurotransmission in the nucleus tractus solitarius in Mecp2-null mice, a model of Rett syndrome. Neurobiol. Dis. 2018, 109, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Lozovaya, N.; Nardou, R.; Tyzio, R.; Chiesa, M.; Pons-Bennaceur, A.; Eftekhari, S.; Bui, T.T.; Billon-Grand, M.; Rasero, J.; Bonifazi, P.; et al. Early alterations in a mouse model of Rett syndrome: The GABA developmental shift is abolished at birth. Sci. Rep. 2019, 9, 9276. [Google Scholar] [CrossRef] [PubMed]
- Tsujimura, K.; Irie, K.; Nakashima, H.; Egashira, Y.; Fukao, Y.; Fujiwara, M.; Itoh, M.; Uesaka, M.; Imamura, T.; Nakahata, Y.; et al. miR-199a links MeCP2 with mTOR signaling and its dysregulation leads to Rett syndrome phenotypes. Cell Rep. 2015, 12, 1887–1901. [Google Scholar] [CrossRef] [PubMed]
- Gabel, H.W.; Kinde, B.; Stroud, H.; Gilbert, C.S.; Harmin, D.A.; Kastan, N.R.; Hemberg, M.; Ebert, D.H.; Greenberg, M.E. Disruption of DNA-methylation-dependent long gene repression in Rett syndrome. Nature 2015, 522, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Detich, N.; Bovenzi, V.; Szyf, M. Valproate induces replication-independent active DNA demethylation. J. Biol. Chem. 2003, 278, 27586–27592. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ebihara, S.; Ishii, K.; Kobayashi, T.; Nishijima, M.; Endo, S.; Takaku, A.; Sakagami, H.; Kondo, H.; Tashiro, F.; et al. Structural and functional characterization of mouse glutamate decarboxylase 67 gene promoter. Biochim. Biophys. Acta 2003, 1628, 156–168. [Google Scholar] [CrossRef]
- Fujita, N.; Takebayashi, S.; Okumura, K.; Kudo, S.; Chiba, T.; Saya, H.; Nakao, M. Methylation-mediated transcriptional silencing in euchromatin by methyl-CpG binding protein MBD1 isoforms. Mol. Cell. Biol. 1999, 19, 6415–6426. [Google Scholar] [CrossRef]
- Clouaire, T.; de Las Heras, J.I.; Merusi, C.; Stancheva, I. Recruitment of MBD1 to target genes requires sequence-specific interaction of the MBD domain with methylated DNA. Nucleic Acids Res. 2010, 38, 4620–4634. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Chen, B.F.; Chan, W.Y. An epigenetic regulator: Methyl-CpG-binding domain protein 1 (MBD1). Int. J. Mol. Sci. 2015, 16, 5125. [Google Scholar] [CrossRef]
- Kavalali, E.T.; Nelson, E.D.; Monteggia, L.M. Role of MeCP2, DNA methylation, and HDACs in regulating synapse function. J. Neurodev. Disord. 2011, 3, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Kohli, R.M.; Zhang, Y. TET enzymes, TDG and the dynamics of DNA demethylation. Nature 2013, 502, 472–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, A.; Shinoda, M.; Honda, K.; Shirakawa, T.; Iwata, K. Regulation of transient receptor potential vanilloid 1 expression in trigeminal ganglion neurons via methyl-CpG binding protein 2 signaling contributes tongue heat sensitivity and inflammatory hyperalgesia in mice. Mol. Pain 2016, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Abdala, A.P.; Bissonnette, J.M.; Newman-Tancredi, A. Pinpointing brainstem mechanisms responsible for autonomic dysfunction in Rett syndrome: Therapeutic perspectives for 5-HT1A agonists. Front. Physiol. 2014, 5, 205. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, B.; Nativio, P.; Fabbri, A.; Ricceri, L.; Adriani, W.; Lacivita, E.; Leopoldo, M.; Passarelli, F.; Fuso, A.; Laviola, G. Pharmacological stimulation of the brain serotonin receptor 7 as a novel therapeutic approach for Rett syndrome. Neuropsychopharmacology 2014, 39, 2506–2518. [Google Scholar] [CrossRef]
- Abdala, A.P.; Lioy, D.T.; Garg, S.K.; Knopp, S.J.; Paton, J.F.R.; Bissonnette, J.M. Effect of sarizotan, a 5-HT1a and D2-like receptor agonist, on respiration in three mouse models of Rett syndrome. Am. J. Respir. Cell Mol. Biol. 2014, 50, 1031–1039. [Google Scholar] [CrossRef]
- Gogliotti, R.G.; Senter, R.K.; Fisher, N.M.; Adams, J.; Zamorano, R.; Walker, A.G.; Blobaum, A.L.; Engers, D.W.; Hopkins, C.R.; Daniels, J.S.; et al. mGlu7 potentiation rescues cognitive, social, and respiratory phenotypes in a mouse model of Rett syndrome. Sci. Transl. Med. 2017, 9, eaai7459. [Google Scholar] [CrossRef]
- Bittolo, T.; Raminelli, C.A.; Deiana, C.; Baj, G.; Vaghi, V.; Ferrazzo, S.; Bernareggi, A.; Tongiorgi, E. Pharmacological treatment with mirtazapine rescues cortical atrophy and respiratory deficits in MeCP2 null mice. Sci. Rep. 2016, 6, 19796. [Google Scholar] [CrossRef]
- Guo, W.; Tsujimura, K.; Otsuka, I.M.; Irie, K.; Igarashi, K.; Nakashima, K.; Zhao, X. VPA alleviates neurological deficits and restores gene expression in a mouse model of Rett syndrome. PLoS ONE 2014, 9, e100215. [Google Scholar] [CrossRef]
- Thurston, J.H.; Hauhart, R.E. Amelioration of adverse effects of valproate on ketogenesis and liver coenzyme A metabolism by cotreatment with pantothenate and carnitine in developing mice: Possible clinical significance. Pediatr. Res. 1992, 31, 419–423. [Google Scholar] [CrossRef]
- Schaevitz, L.R.; Nicolai, R.; Lopez, C.M.; D’Iddio, S.; Iannoni, E.; Berger-Sweeney, J.E. Acetyl-L-carnitine improves behavior and dendritic morphology in a mouse model of Rett syndrome. PLoS ONE 2012, 7, e51586. [Google Scholar] [CrossRef]
- Franklin, K.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates, 3rd ed.; Academic Press: Cambridge, MA, USA, 2008; pp. 86–96. [Google Scholar]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishiyama, M.; Tamura, S.; Ito, H.; Takei, H.; Hoshi, M.; Asano, M.; Itoh, M.; Shirakawa, T. Early Postnatal Treatment with Valproate Induces Gad1 Promoter Remodeling in the Brain and Reduces Apnea Episodes in Mecp2-Null Mice. Int. J. Mol. Sci. 2019, 20, 5177. https://doi.org/10.3390/ijms20205177
Ishiyama M, Tamura S, Ito H, Takei H, Hoshi M, Asano M, Itoh M, Shirakawa T. Early Postnatal Treatment with Valproate Induces Gad1 Promoter Remodeling in the Brain and Reduces Apnea Episodes in Mecp2-Null Mice. International Journal of Molecular Sciences. 2019; 20(20):5177. https://doi.org/10.3390/ijms20205177
Chicago/Turabian StyleIshiyama, Misa, Satoko Tamura, Hisanori Ito, Hiroki Takei, Manami Hoshi, Masatake Asano, Masayuki Itoh, and Tetsuo Shirakawa. 2019. "Early Postnatal Treatment with Valproate Induces Gad1 Promoter Remodeling in the Brain and Reduces Apnea Episodes in Mecp2-Null Mice" International Journal of Molecular Sciences 20, no. 20: 5177. https://doi.org/10.3390/ijms20205177