Mycelium Polysaccharides from Termitomyces albuminosus Attenuate CCl4-Induced Chronic Liver Injury Via Inhibiting TGFβ1/Smad3 and NF-κB Signal Pathways

 ,
,
Abstract
:1. Introduction
2. Results
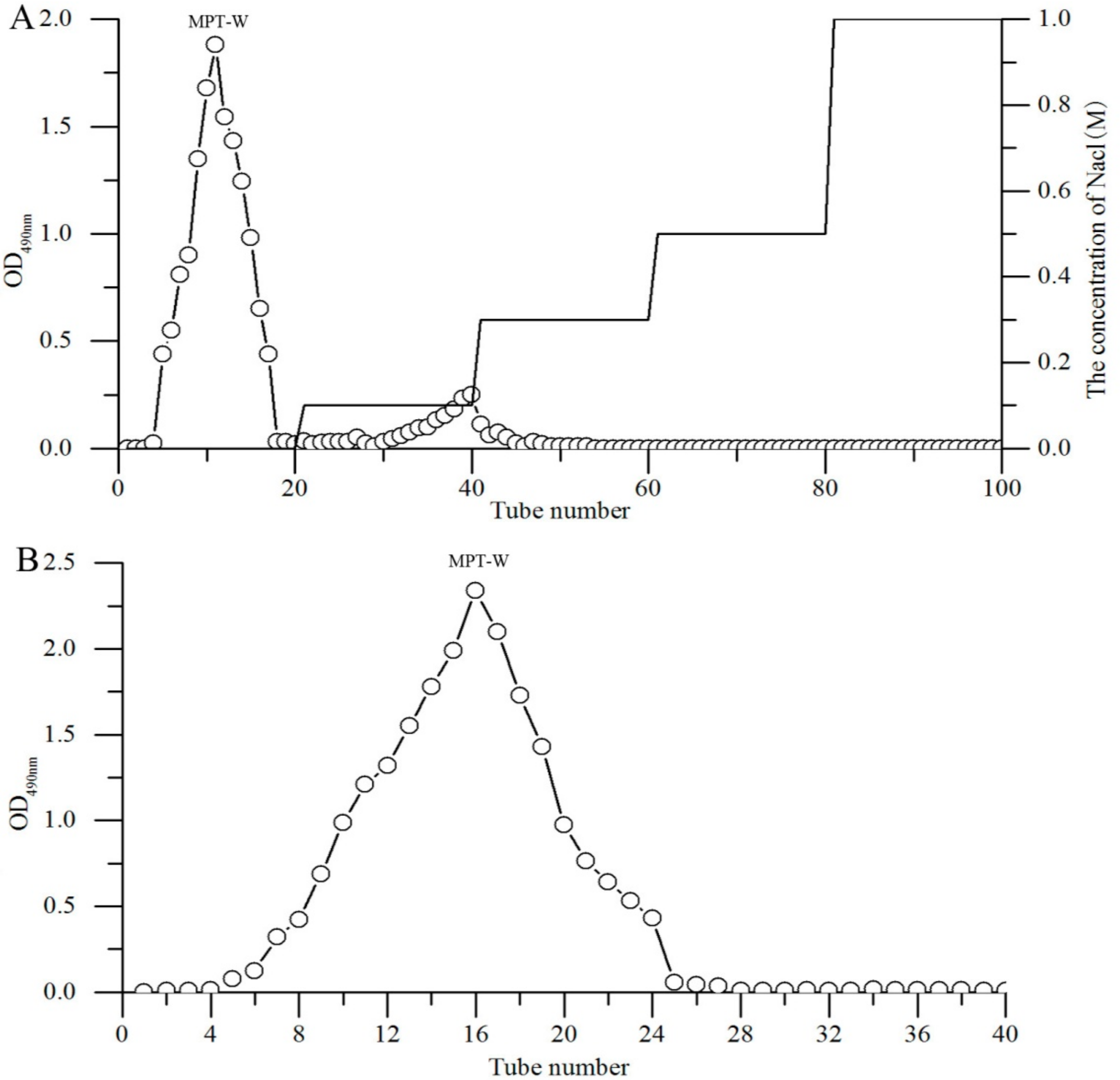
2.1. Purification
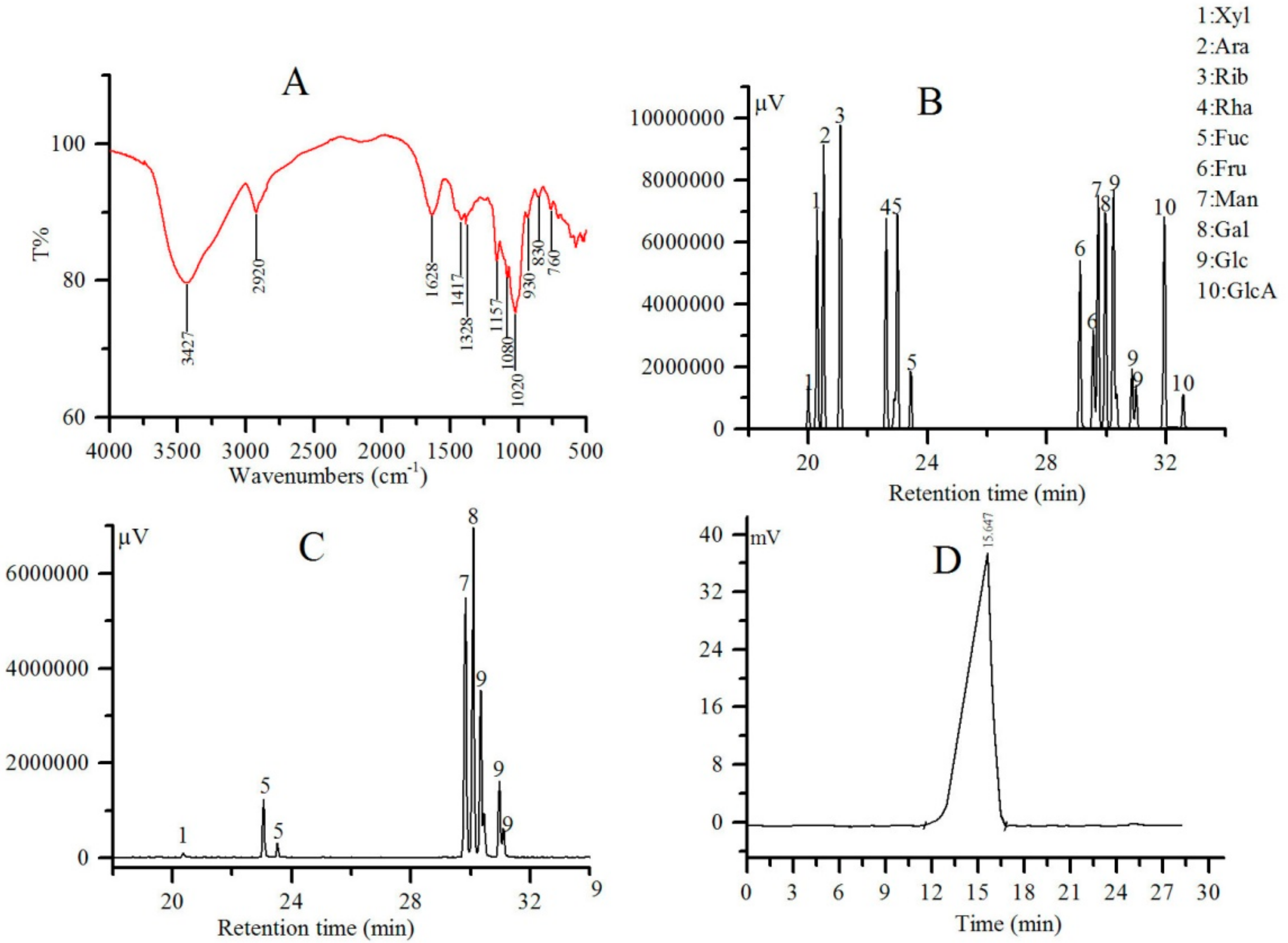
2.2. FT-IR, Monosaccharide Composition and Mw Analysis
2.3. In Vitro Antioxidant Activity of MPT-W
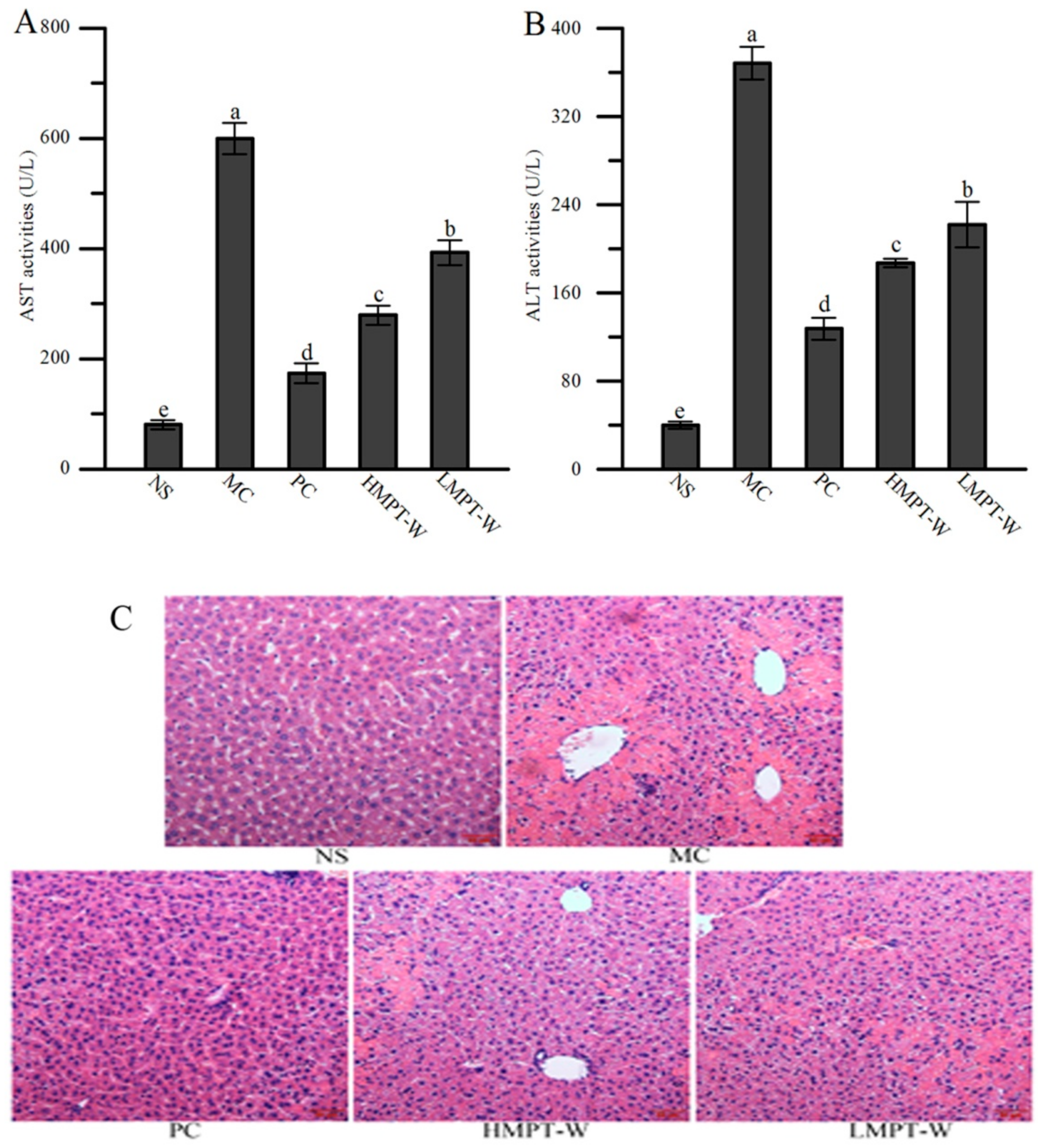
2.4. Effect of MPT-W on Liver Injury in CCl4-Induced Chronic Liver Injury Mice
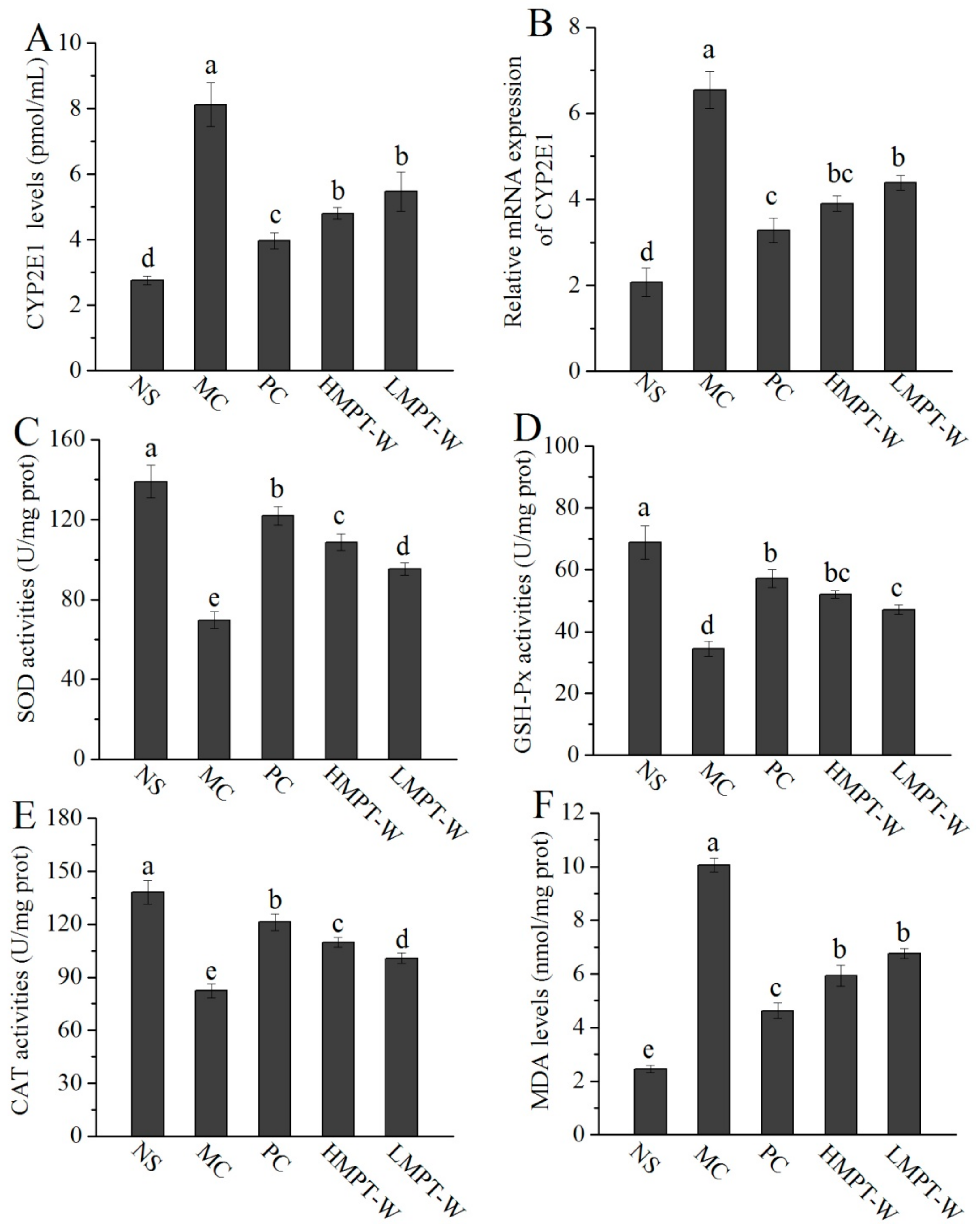
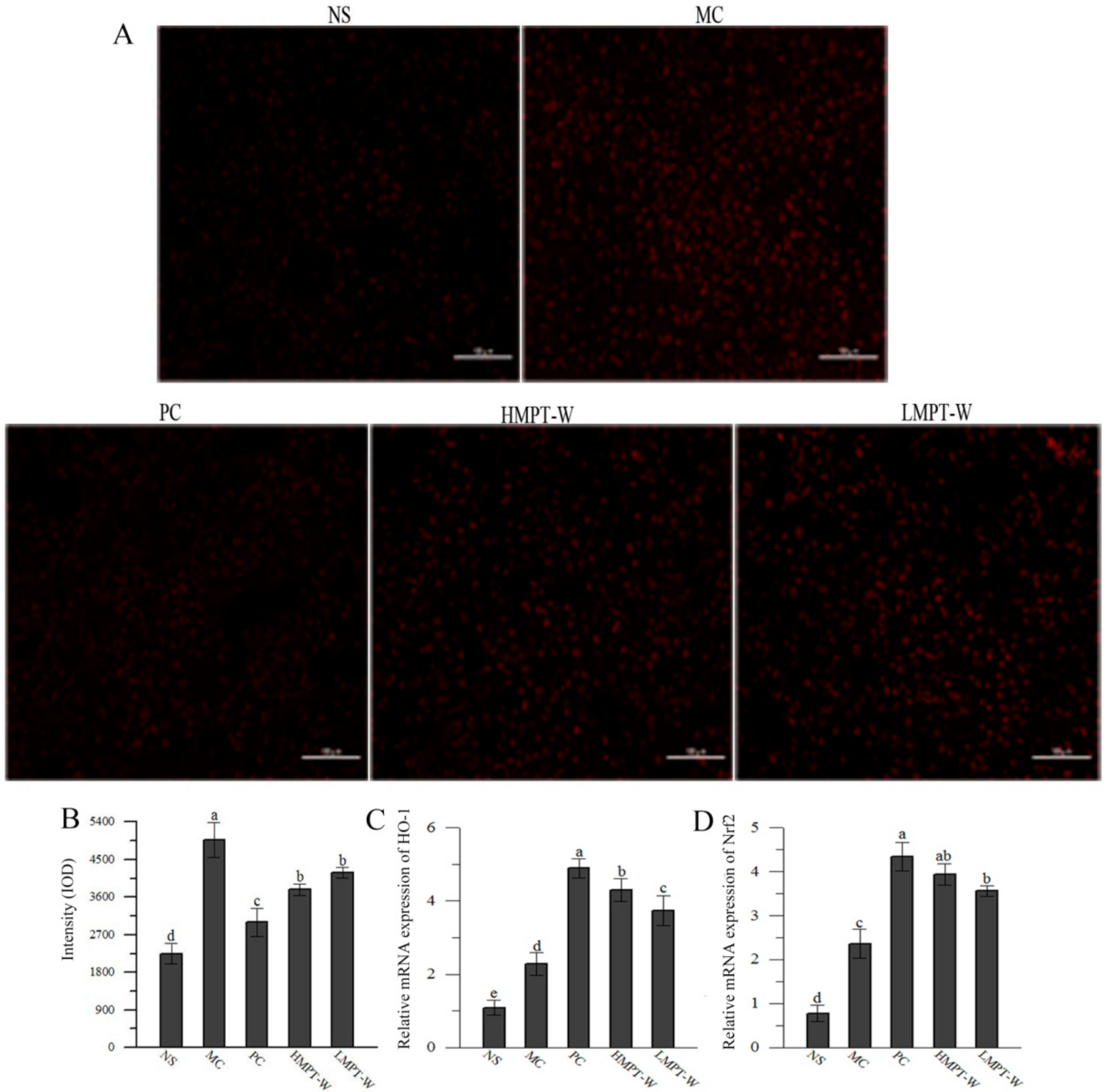
2.5. Effect of MPT-W on Antioxidant Status in CCl4-Induced Chronic Liver Injury Mice
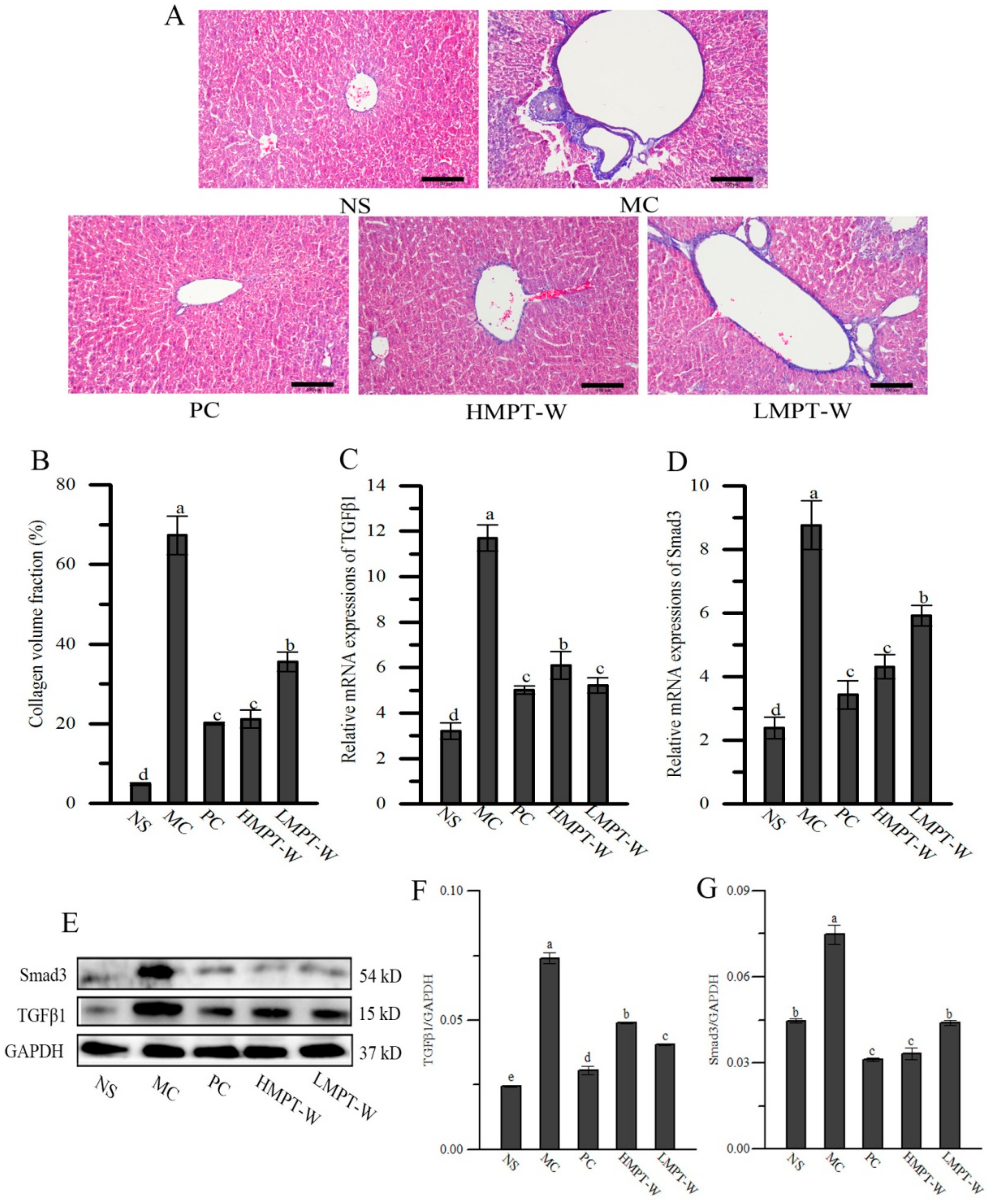
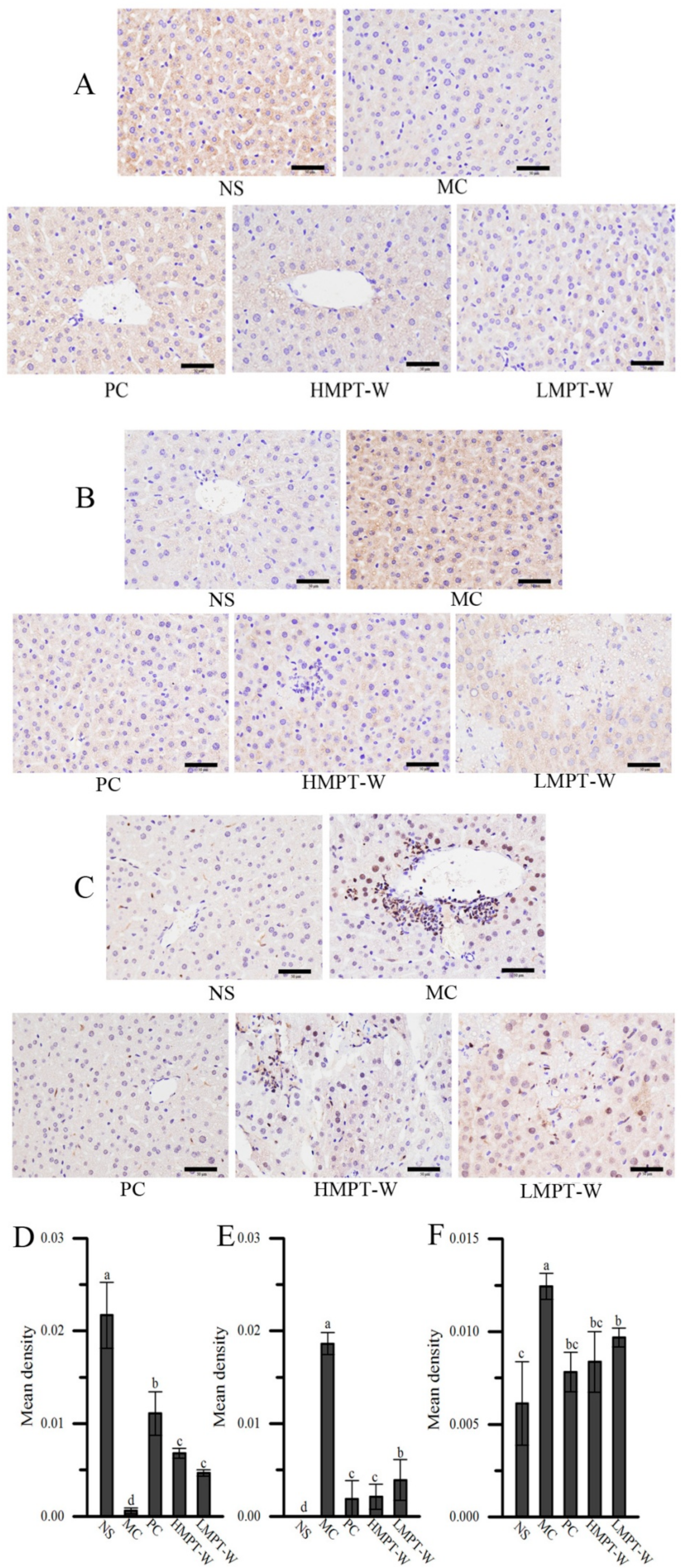
2.6. Effect of MPT-W on Fibrosis in CCl4-Induced Chronic Liver Injury Mice
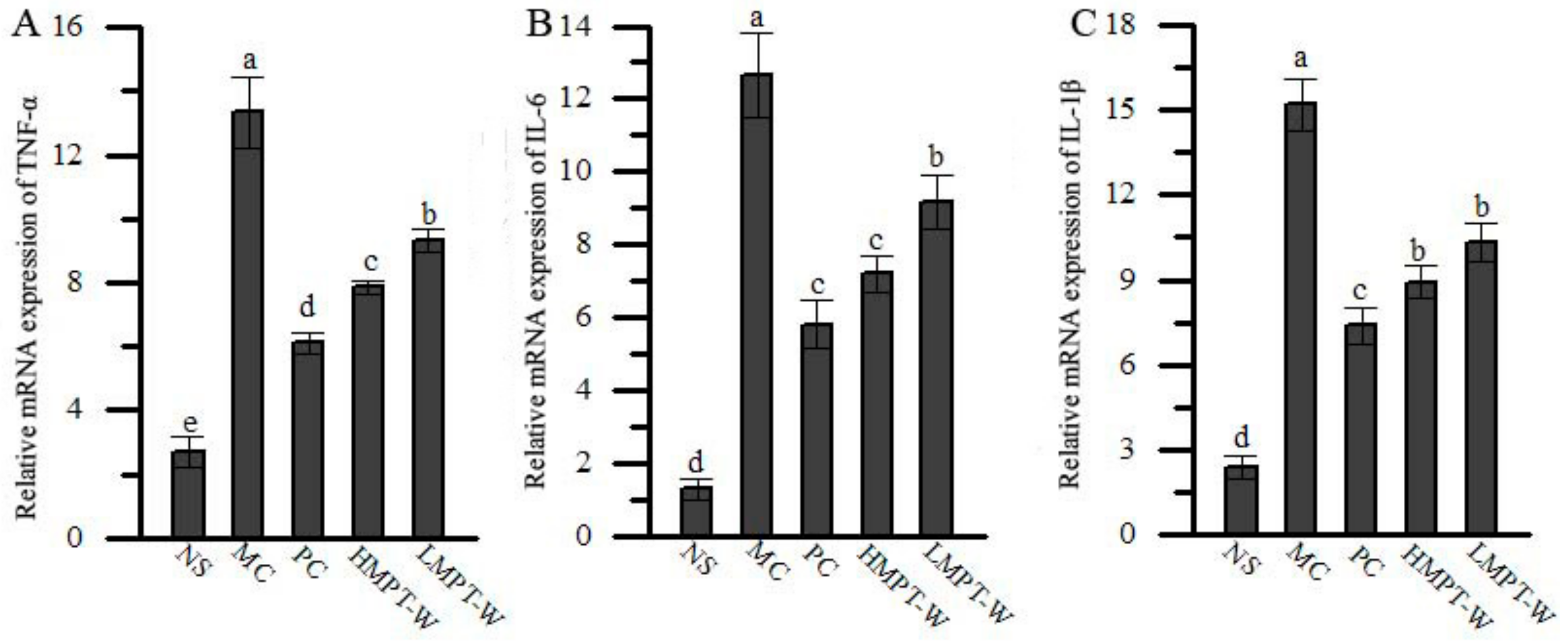
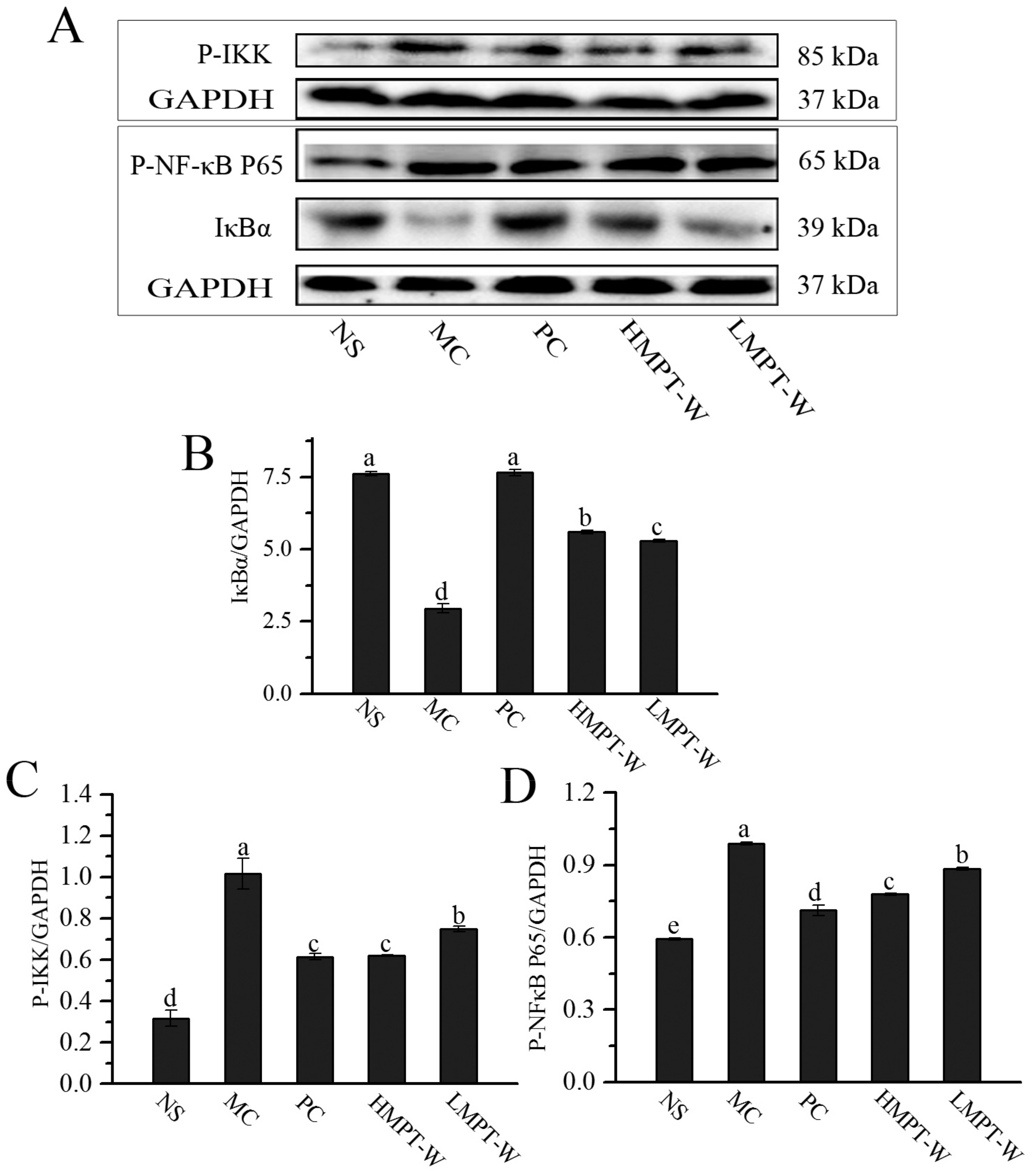
2.7. Effect of MPT-W on Hepatic Inflammatory Response in CCl4-Induced Chronic Liver Injury Mice
3. Discussion
4. Materials and methods
4.1. Materials
4.2. Extraction and Purification of MPT
4.3. Monosaccharide Compositions Analysis
4.4. FT-IR Spectroscopy Analysis
4.5. Mw Determination
4.6. In Vitro Antioxidant Activity of MPT-W
4.7. Animal Experiment
4.8. Enzyme Activities in Serum Assessment
4.9. Biochemical Indices Evaluation in Liver Homogenates
4.10. Western Blotting
4.11. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR) Analysis
4.12. Histopathological Analysis
4.13. Immunohistochemistry Staining
4.14. ROS Level Assay
4.15. Statistical analysis
5. Conclusions
Abbreviation
| Ara | Arabinose |
| CCl4 | Carbon tetrachloride |
| CAT | Catalase |
| CYP2E1 | Cytochrome P4502E1 |
| FT-IR | Fourier-transform infrared spectroscopy |
| Fru | Fructose |
| Fuc | Fucose |
| Gal | Galactose |
| GC-MS | Gas chromatography-mass spectrometry |
| Glc | Glucose |
| GlcA | Glucuronic acid |
| GSH-Px | GSH peroxide |
| HPGPC | high performance gel permeation chromatography |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin-6 |
| MDA | Malonaldehyde |
| Man | Mannose |
| MC | model control |
| Mw | Molecular weight |
| MPT | Mycelium polysaccharides from T. albuminosus |
| NS | Normal saline |
| Mn | Number-average molecular weights |
| PC | positive control |
| ROS | Reactive oxygen species |
| Rha | Rhamnose |
| Ri | Ribose |
| SOD | Superoxide dismutase |
| TNF-α | Tumor necrosis factor |
| Xyl | Xylose |
| Mz | Z-average molecular weights |
| HMPT-W | 400 mg/kg MPT-W |
| LMPT-W | 100 mg/kg MPT-W |
Author Contributions
Funding
Conflicts of Interest
References
- Rocha, S.W.S.; França, M.E.R.D.; Rodrigues, G.B.; Barbosa, K.P.S.; Nunes, A.K.S.; Pastor, A.F.; Oliveira, A.G.V.; Oliveira, W.H.; Luna, R.L.A.; Peixoto, C.A. Diethylcarbamazine reduces chronic inflammation and fibrosis in carbon tetrachloride-(CCl4-) induced liver injury in mice. Mediat. Inflamm. 2014. [Google Scholar] [CrossRef] [PubMed]
- Quintal-Novelo, C.; Rangel-Méndez, J.; Ortiz-Tello, Á.; Graniel-Sabido, M.; Pérez-Cabeza, R.; Moo-Puc, R. A Sargassum fluitans borgesen ethanol extract exhibits a hepatoprotective effect in vivo in acute and chronic liver damage Models. BioMed Res. Int. 2018. [Google Scholar] [CrossRef]
- Altaş, S.; Kızıl, G.; Kızıl, M.; Ketani, A.; Haris, P.I. Protective effect of Diyarbakır watermelon juice on carbon tetrachloride-induced toxicity in rats. Food Chem. Toxicol. 2011, 49, 2433–2438. [Google Scholar] [CrossRef]
- Shi, H.; Han, W.; Shi, H.; Ren, F.; Chen, D.; Chen, Y.; Duan, Z. Augmenter of liver regeneration protects against carbon tetrachloride-induced liver injury by promoting autophagy in mice. Oncotarget 2017, 8, 12637–12648. [Google Scholar] [PubMed] [Green Version]
- Jaeschke, H.; McGill, M.R.; Ramachandran, A. Oxidant stress, mitochondria, and cell death mechanisms in drug-induced liver injury: Lessons learned from acetaminophen hepatotoxicity. Drug Metab. Rev. 2012, 44, 88–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wang, S.; Liu, J.; Cai, E.; Zhu, H.; He, Z.; Gao, Y.; Li, P.; Zhao, Y. Sesquiterpenoids from the root of Panax Ginseng protect CCl4-induced acute liver injury by anti-inflammatory and anti-oxidative capabilities in mice. Biomed. Pharmacother. 2018, 102, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Upur, H.; Amat, N.; Blazeković, B.; Talip, A. Protective effect of Cichorium glandulosum root extract on carbon tetrachloride-induced and galactosamine-induced hepatotoxicity in mice. Food Chem. Toxicol. 2009, 47, 2022–2030. [Google Scholar]
- Abe, T. Studies on the distribution and ecological role of termites in a lowland rain forest of West Malaysia. 3. Distribution and abundance of termites in Pasoh Forest Reserve. Jpn. J. Ecol. 1979, 29, 121–135. [Google Scholar]
- Wood, T.G.; Sands, W.A. Role of termites in ecosystems. Int. Biol. Program. 1978, 245–251. [Google Scholar]
- Johjima, T.; Ohkuma, M.; Kudo, T. Isolation and cDNA cloning of novel hydrogen peroxide-dependent phenol oxidase from the basidiomycete Termitomyces albuminosus. Appl. Microbiol. Biotechnol. 2003, 61, 220–225. [Google Scholar] [CrossRef]
- Lu, Y.Y.; Ao, Z.H.; Lu, Z.M.; Xu, H.Y.; Zhang, X.M.; Dou, W.F.; Xu, Z.H. Analgesic and anti-inflammatory effects of the dry matter of culture broth of Termitomyces albuminosus and its extracts. J. Ethnopharmacol. 2008, 120, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Wang, H.; Zhang, G. A novel alkaline protease from wild edible mushroom Termitomyces albuminosus. Acta biochim. Pol. 2011, 58, 269–273. [Google Scholar] [CrossRef] [PubMed]
- De Souza, R.A.; Kamat, N.M.; Nadkarni, V.S. Purification and characterisation of a sulphur rich melanin from edible mushroom Termitomyces albuminosus Heim. Mycology 2018, 9, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, S.; Zhang, J.; Che, G.; Zhou, M.; Liu, M.; Zhang, C.; Xu, N.; Lin, L.; Liu, Y.; et al. The antihyperlipidemic activities of enzymatic and acidic intracellular polysaccharides by Termitomyces albuminosus. Carbohydr. Polym. 2016, 151, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S. Compendium of Materia Medica; People′s Medical Publishing House: Beijing, China, 2002; Volume 2, pp. 17–19. [Google Scholar]
- Gebreyohannes, G.; Nyerere, A.; Bii, C.; Sbhatu, D.B. Investigation of antioxidant and antimicrobial activities of different extracts of auricularia and Termitomyces species of mushrooms. Sci. World J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Feng, C.P.; Li, X.; Chang, M.C.; Meng, J.L.; Xu, L.J. Immunomodulatory and antioxidative activity of Cordyceps militaris polysaccharides in mice. Int. J. Biol. Macromol. 2016, 86, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Ozalp, F.O.; Canbek, M.; Yamac, M.; Kanbak, G.; Van Griensven, L.J.; Uyanoglu, M.; Senturk, H.; Kartkaya, K.; Oglakci, A. Consumption of Coprinus comatus polysaccharide extract causes recovery of alcoholic liver damage in rats. Pharm. Biol. 2014, 52, 994–1002. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, F.; Liu, X.; St Ange, K.; Zhang, A.; Li, Q.; Linhardt, R.J. Isolation of a lectin binding rhamnogalacturonan-I containing pectic polysaccharide from pumpkin. Carbohydr. Polym. 2017, 163, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Wang, J.; Zhao, M.; Liu, Y.; Wang, W.; Jiang, Y. Identification of polysaccharides from pericarp tissues of litchi (Litchi chinensis Sonn.) fruit in relation to their antioxidant activities. Carbohydr. Res. 2006, 341, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wu, W.; Ren, Y.; Li, X.; Tang, Y.; Min, T.; Lai, F.; Wu, H. Structural characterization of a novel polysaccharide from Lepidium meyenii (Maca) and analysis of its regulatory function in macrophage polarization in vitro. J. Agric. Food Chem. 2017, 65, 1146–1157. [Google Scholar] [CrossRef]
- Habijanic, J.; Berovic, M.; Boh, B.; Plankl, M.; Wraber, B. Submerged cultivation of Ganoderma lucidum and the effects of its polysaccharides on the production of human cytokines TNF-α, IL-12, IFN-γ, IL-2, IL-4, IL-10 and IL-17. N. Biotechnol. 2015, 32, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fan, Y.; Wang, W.; Liu, N.; Zhang, H.; Zhu, Z.; Liu, A. Polysaccharides from Lycium barbarum leaves: Isolation, characterization and splenocyte proliferation activity. Int. J. Biol. Macromol. 2012, 51, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Li, J.; Xu, N.; Zhang, J.; Song, X.; Wang, X.; Gao, Z.; Jing, H.; Li, S.; Zhang, C.; et al. Anti-hyperlipidemic and antioxidant effects of alkali-extractable mycelia polysaccharides by Pleurotus eryngii var. tuolensis. Carbohydr. Polym. 2017, 175, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Kosecik, M.; Erel, O.; Sevinc, E.; Selek, S. Increased oxidative stress in children exposed to passive smoking. Int. J. Cardiol. 2005, 100, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Ban, X.; He, J.; Zeng, H.; Zhang, P.; Wang, Y. Hepatoprotective and antioxidant effects of the methanolic extract from Halenia elliptica. J. Ethnopharmacol. 2010, 131, 276–281. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. The importance of free radicals and catalytic metal ions in human diseases. Mol. Asp. Med. 1985, 8, 89–193. [Google Scholar] [CrossRef]
- Liang, D.; Zhou, Q.; Gong, W.; Wang, Y.; Nie, Z.; He, H.; Li, J.; Wu, J.; Wu, C.; Zhang, J. Studies on the antioxidant and hepatoprotective activities of polysaccharides from Talinum triangulare. J. Ethnopharmacol. 2011, 136, 316–321. [Google Scholar] [CrossRef]
- Li, R.; Chen, W.C.; Wang, W.P.; Tian, W.; Zhang, X.G. Antioxidant activity of Astragalus polysaccharides and antitumour activity of the polysaccharides and siRNA. Carbohydr. Polym. 2010, 82, 220–244. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, M.; Yang, Y.; Lin, L.; Xu, N.; Zhao, H.; Jia, L. Purification, characterization and hepatoprotective activities of mycelia zinc polysaccharides by Pleurotus djamor. Carbohydr. Polym. 2016, 136, 588–597. [Google Scholar] [CrossRef]
- Zhao, H.; Lai, Q.; Zhang, J.; Huang, C.; Jia, L. Antioxidant and hypoglycemic effects of acidic-extractable polysaccharides from Cordyceps militaris on type 2 diabetes mice. Oxid. Med. Cell. Longev. 2018. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, D.D.; Su, L.; Wang, Q.; Li, Y. Protective effect of polysaccharide from Agaricus bisporus in Tibet area of China against tetrachloride-induced acute liver injury in mice. Int. J. Biol. Macromol. 2018, 118, 1488–1493. [Google Scholar] [CrossRef] [PubMed]
- Rahmat, A.A.; Dar, F.A.; Choudhary, I.M. Protection of CCl4-induced liver and kidney damage by phenolic compounds in leaf extracts of cnestis ferruginea (de Candolle). Pharmacognosy Res. 2014, 6, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, A.; Spasic, S.; Jovanovic-Galovic, A.; Djordjevic, R.; Grubor-Lajsic, G. Oxidative stress in diabetic pregnancy: Sod, cat and GSH-Px activity and lipid peroxidation products. J. Matern. Fetal Neonatal Med. 2004, 16, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Sabir, S.M.; Ahmad, S.D.; Hamid, A. Antioxidant and hepatoprotective activity of ethanolic extract of leaves of Solidago microglossa containing polyphenolic compounds. Food Chem. 2012, 131, 741–747. [Google Scholar] [CrossRef]
- Tebay, L.E.; Robertson, H.; Durant, S.T.; Vitale, S.R.; Penning, T.M.; Dinkovakostova, A.T. Mechanisms of activation of the transcription factor Nrf2 by redox stressors, nutrient cues, and energy status and the pathways through which it attenuates degenerative disease. Free Radic. Biol. Med. 2015, 88, 108–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Dai, C.; Liu, Q.; Li, J.; Qiu, J. Curcumin attenuates on carbon tetrachloride-induced acute liver injury in mice via modulation of the Nrf2/HO-1 and TGF-β1/Smad3 Pathway. Molecules 2018, 23, 215. [Google Scholar] [CrossRef]
- Niu, L.; Cui, X.; Qi, Y.; Xie, D.; Wu, Q.; Chen, X.; Ge, J.; Liu, Z. Involvement of TGF-β1/Smad3 signaling in carbon tetrachloride-induced acute liver injury in mice. PLoS ONE 2016. [Google Scholar] [CrossRef]
- Bai, F.; Huang, Q.; Nie, J.; Lu, S.; Lu, C.; Zhu, X.; Wang, Y.; Zhuo, L.; Lu, Z.; Lin, X. Trolline Ameliorates Liver Fibrosis by Inhibiting the NF-κB Pathway, Promoting HSC Apoptosis and Suppressing Autophagy. Cell. Physiol. Biochem. 2017, 44, 436–446. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Yuan, Y.; Yue, T. Immunomodulatory of selenium nano-particles decorated by sulfated Ganoderma lucidum polysaccharides. Food Chem. Toxicol. 2014, 68, 183–189. [Google Scholar] [CrossRef]
- Li, B.; Cong, M.; Zhu, Y.; Xiong, Y.; Jin, W.; Wan, Y.; Zhou, Y.; Ao, Y.; Wang, H. Indole-3-carbinol induces apoptosis of hepatic stellate cells through K63 de-ubiquitination of RIP1 in Rats. Cell. Physiol. Biochem. 2017, 41, 1481–1490. [Google Scholar] [CrossRef]
- Staub, A.M. Removal of proteins from polysaccharides methods. Carbohydr. Chem. 1965, 5, 5–6. [Google Scholar]
- Cheng, N.; Ren, N.; Gao, H.; Lei, X.; Zheng, J.; Cao, W. Antioxidant and hepatoprotective effects of Schisandra chinensis pollen extract on CCl4-induced acute liver damage in mice. Food Chem. Toxicol. 2013, 55, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Irudayaraj, S.S.; Sunil, C.; Duraipandiyan, V.; Ignacimuthu, S. In vitro antioxidant and antihyperlipidemic activities of Toddalia asiatica (L) Lam. leaves in Triton WR-1339 and high fat diet induced hyperlipidemic rats. Food Chem. Toxicol. 2013, 60, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Nishimiki, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in thereaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Bioph. Res. Co. 1972, 46, 849–853. [Google Scholar] [CrossRef]
- Shan, Q.; Li, X.; Zheng, M.; Lin, X.; Lu, G.; Su, D.; Lu, X. Protective effects of dimethyl itaconate in mice acute cardiotoxicity induced by doxorubicin. Biochem. Biophys. Res. Commun. 2019, 517, 538–544. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Mn (Da) | Mw (Da) | Mz (Da) | Mw/Mn |
|---|---|---|---|---|
| MPT-W | 1.13 × 105 | 1.30 × 105 | 1.49 × 105 | 1.15 |
| Indexes | Sample | Concentrations (mg/L) | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 200 | 400 | 600 | 800 | 1000 | 1200 | ||
| DPPH radicals | MPT-W | 0 | 11.91 ± 0.84 | 24.87 ± 2.17 | 42.10 ± 1.70 | 63.84 ± 2.80 | 72.01 ± 2.05 | 76.81 ± 1.31 |
| Vc | 0 | 61.07 ± 3.47 | 75.45 ± 2.07 | 87.24 ± 2.01 | 89.58 ± 1.07 | 91.04 ± 0.69 | 92.65 ± 1.13 | |
| Hydroxyl radicals | MPT-W | 0 | 19.58 ± 1.98 | 38.10 ± 1.83 | 44.19 ± 1.20 | 54.84 ± 1.91 | 67.17 ± 1.97 | 74.11 ± 1.52 |
| Vc | 0 | 41.26 ± 2.68 | 59.63 ± 0.89 | 69.45 ± 3.67 | 85.89 ± 2.17 | 92.75 ± 1.56 | 95.96 ± 0.84 | |
| Superoxide anion radicals | MPT-W | 0 | 18.88 ± 1.29 | 36.12 ± 0.33 | 52.54 ± 1.169 | 57.49 ± 1.84 | 66.16 ± 1.71 | 72.59 ± 0.58 |
| Vc | 0 | 32.69 ± 1.46 | 48.35 ± 0.79 | 65.50 ± 2.65 | 77.88 ± 1.30 | 82.84 ± 1.07 | 88.09 ± 1.49 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Li, H.; Feng, Y.; Zhang, Y.; Yuan, F.; Zhang, J.; Ren, H.; Jia, L. Mycelium Polysaccharides from Termitomyces albuminosus Attenuate CCl4-Induced Chronic Liver Injury Via Inhibiting TGFβ1/Smad3 and NF-κB Signal Pathways. Int. J. Mol. Sci. 2019, 20, 4872. https://doi.org/10.3390/ijms20194872
Zhao H, Li H, Feng Y, Zhang Y, Yuan F, Zhang J, Ren H, Jia L. Mycelium Polysaccharides from Termitomyces albuminosus Attenuate CCl4-Induced Chronic Liver Injury Via Inhibiting TGFβ1/Smad3 and NF-κB Signal Pathways. International Journal of Molecular Sciences. 2019; 20(19):4872. https://doi.org/10.3390/ijms20194872
Chicago/Turabian StyleZhao, Huajie, Huaping Li, Yanbo Feng, Yiwen Zhang, Fangfang Yuan, Jianjun Zhang, Haixia Ren, and Le Jia. 2019. "Mycelium Polysaccharides from Termitomyces albuminosus Attenuate CCl4-Induced Chronic Liver Injury Via Inhibiting TGFβ1/Smad3 and NF-κB Signal Pathways" International Journal of Molecular Sciences 20, no. 19: 4872. https://doi.org/10.3390/ijms20194872