Short-Time Ocular Ischemia Induces Vascular Endothelial Dysfunction and Ganglion Cell Loss in the Pig Retina

 , and
, and
Abstract
:1. Introduction
2. Results
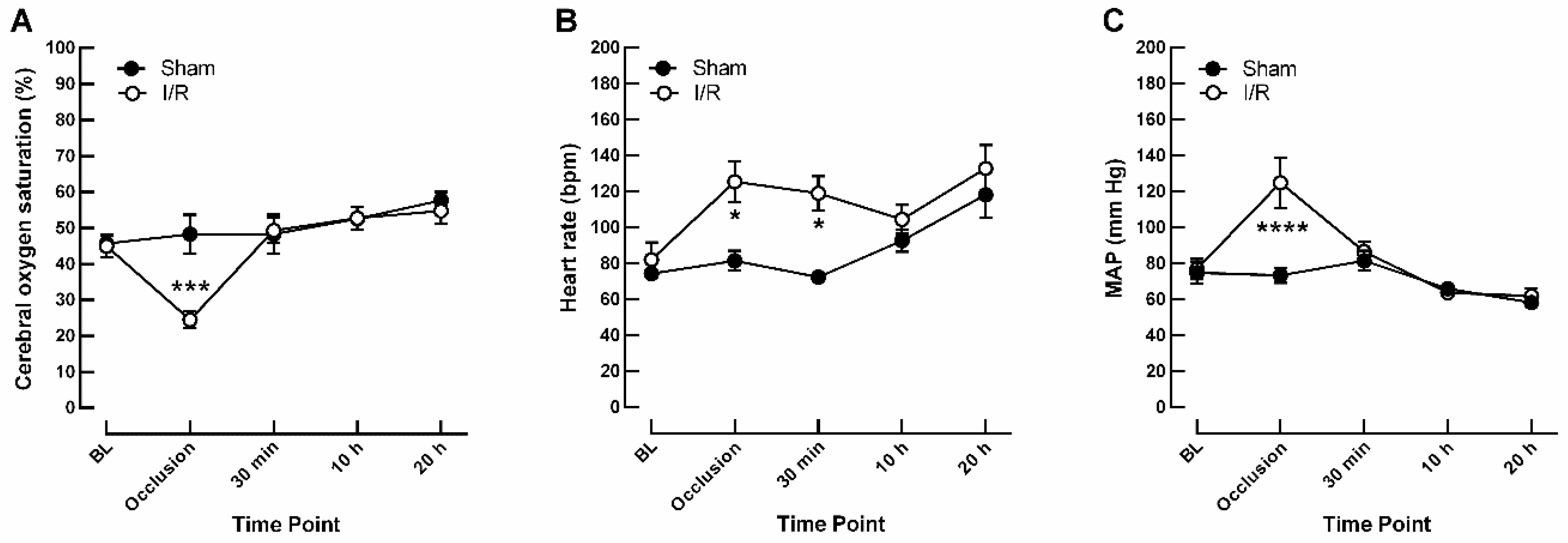
2.1. Effects of I/R on Monitoring Parameters
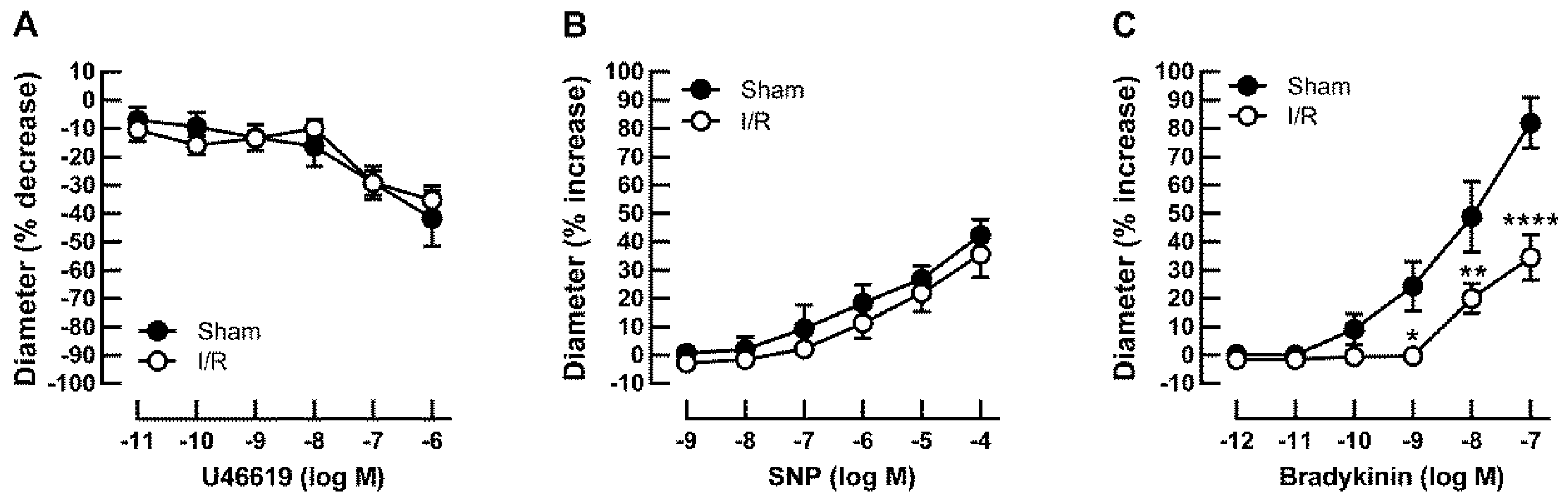
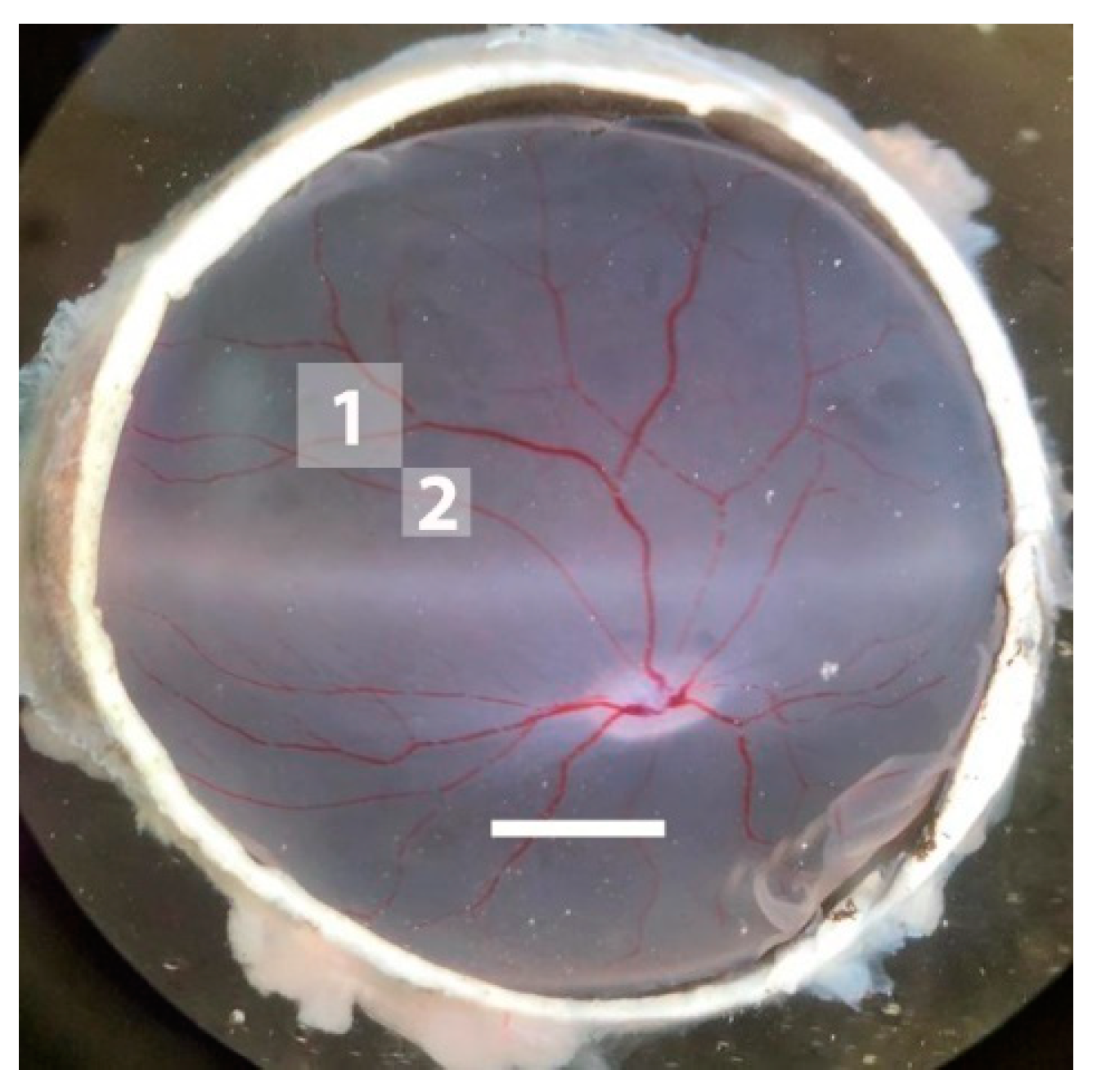
2.2. Effects of I/R on Vascular Responses in Retinal Arterioles
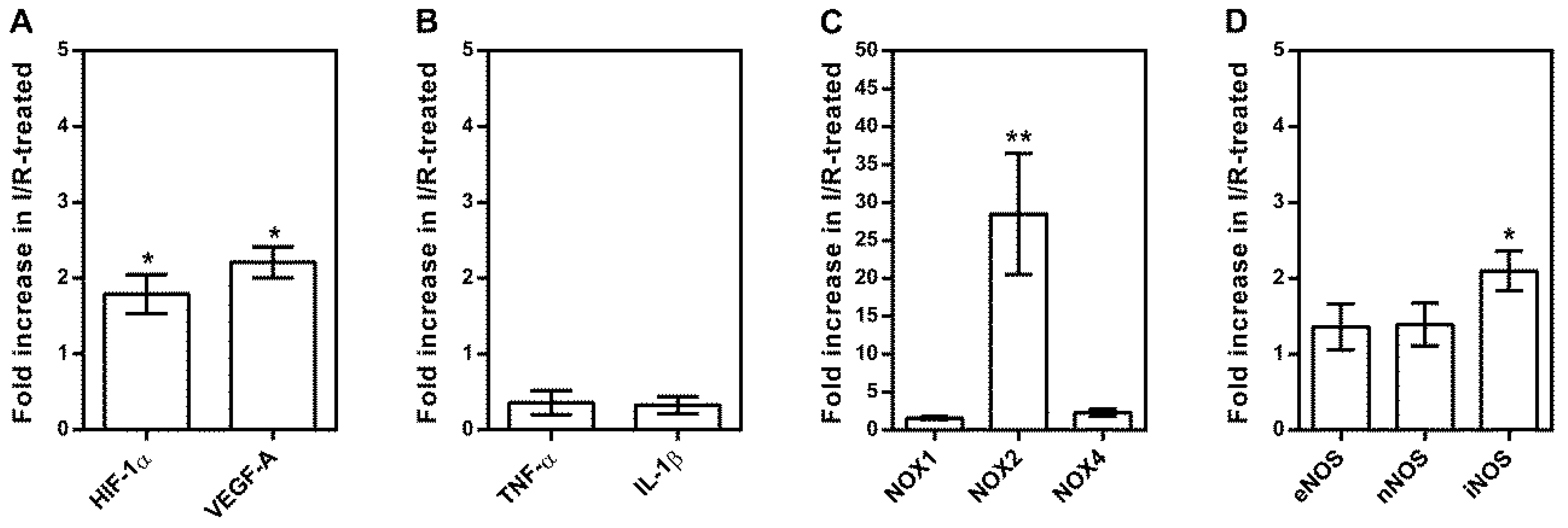
2.3. Messenger RNA Expression Levels in Isolated Retinal Arterioles
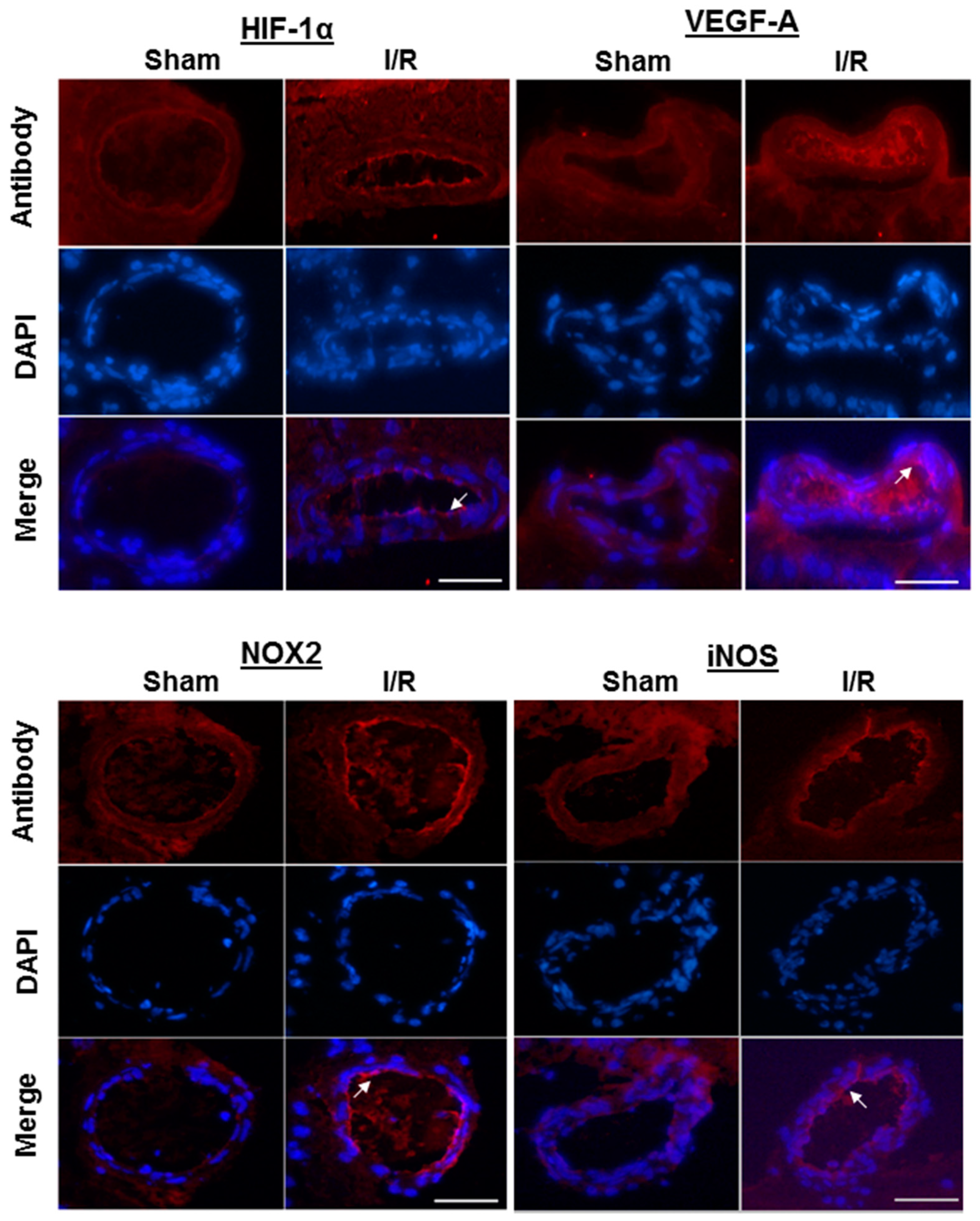
2.4. Immunofluorescence
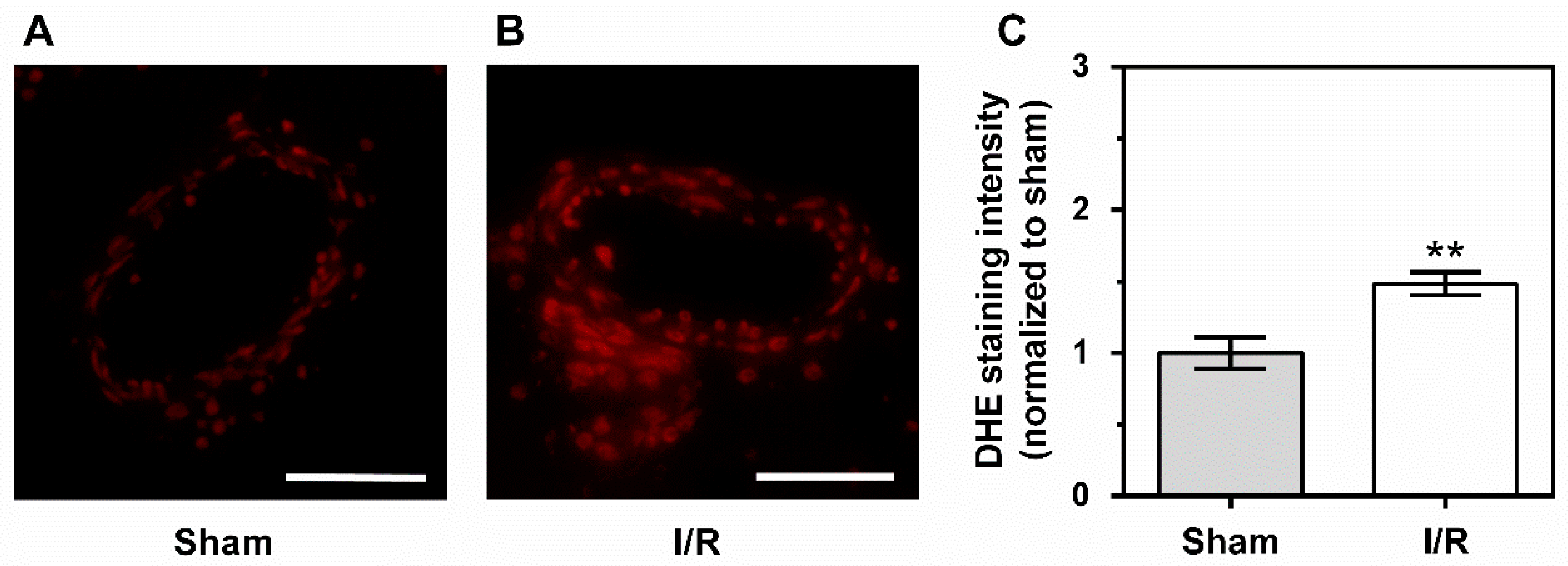
2.5. Levels of Reactive Oxygen Species
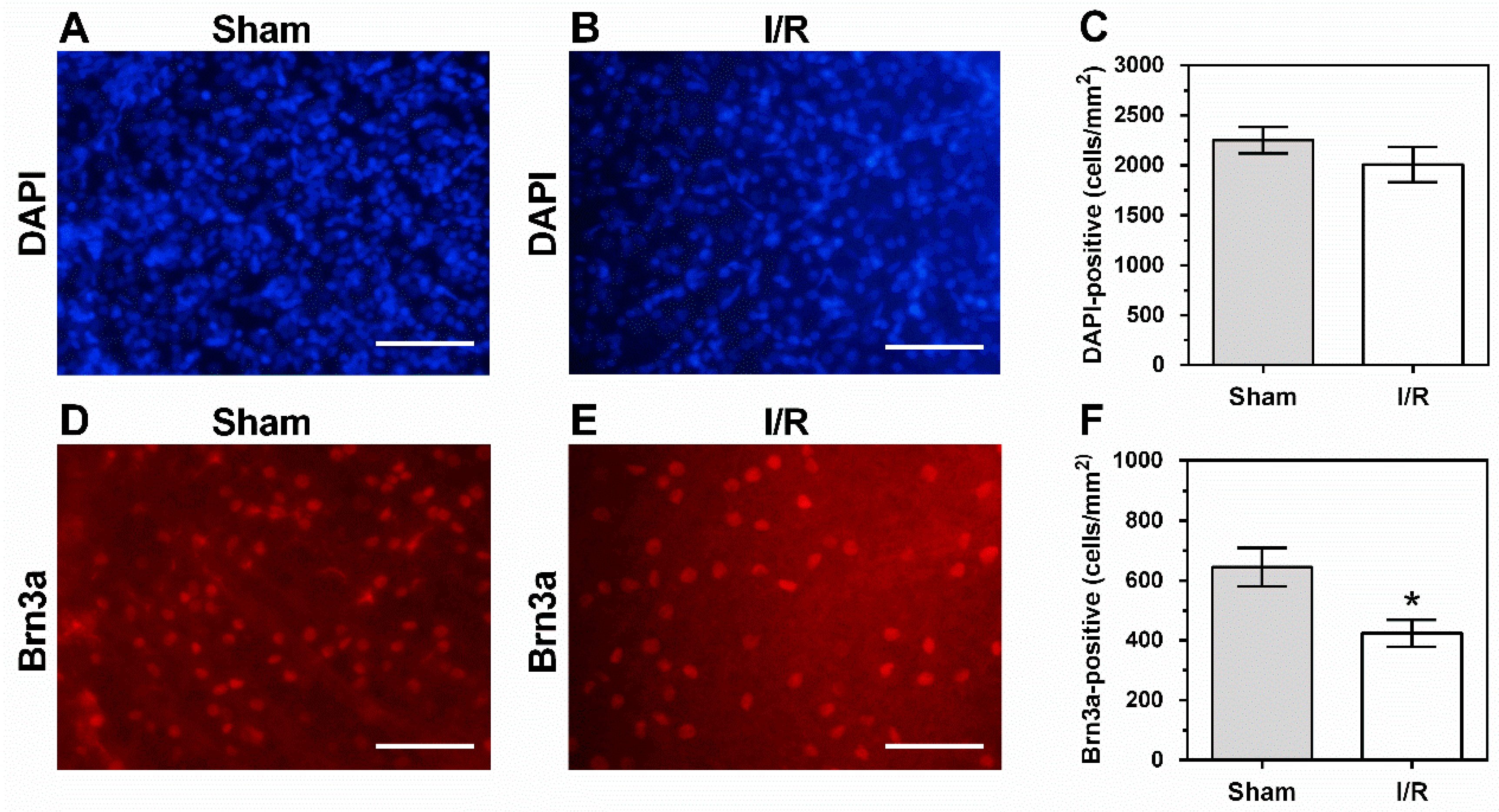
2.6. Cells in the Retinal Ganglion Cell Layer
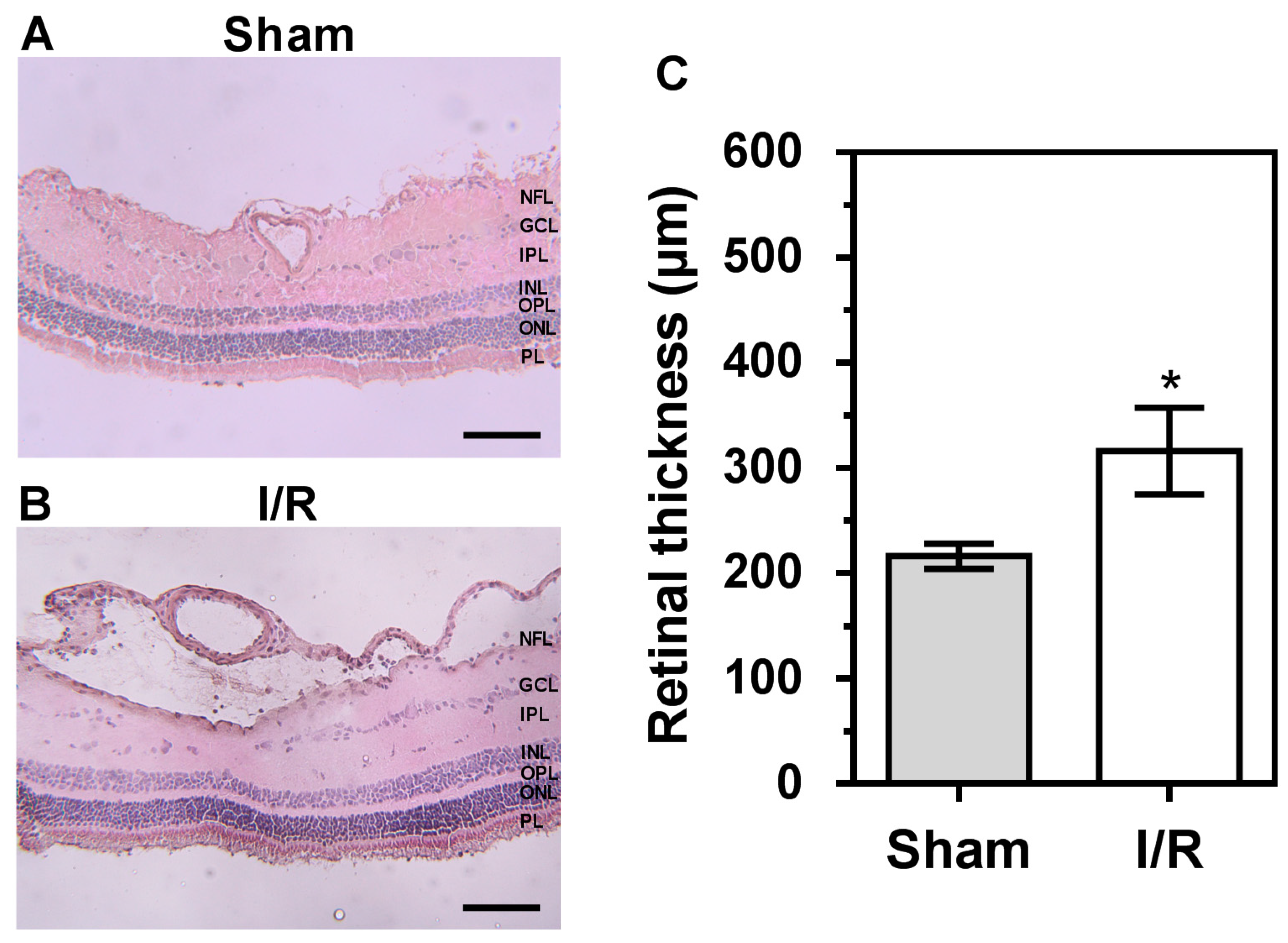
2.7. Retinal Histology
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Materials
4.3. Surgical Procedure
4.4. Measurement of Vascular Reactivity in Retinal Arterioles
4.5. Measurement of Reactive Oxygen Species
4.6. Quantitative PCR
4.7. Immunohistochemistry
4.8. Retinal Histology
4.9. Statistical Methods
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BL | Baseline |
| Brn3a | Brain specific homebox/POU domain protein 3A |
| CRAO | Central retinal artery occlusion |
| DAPI | 4’,6-diamidino-2-phenylindole |
| DHE | Dihydroethidium |
| DMSO | Dimethyl sulfoxide |
| eNOS | Endothelial nitric oxide synthase |
| FCS | Fetal calf serum |
| GCL | Ganglion cell layer |
| HIF-1α | Hypoxia-inducible factor-1α |
| HR | Heartrate |
| iNOS | Inducible nitric oxide synthase |
| INL | Inner nuclear layer |
| IL | Interleukin |
| IPL | Inner plexiform layer |
| I/R | Ischemia-reperfusion |
| MAP | Mean arterial blood pressure |
| NFL | Nerve fiber layer |
| nNOS | Neuronal nitric oxide synthase |
| NOX | Nicotinamide adenine dinucleotide phosphate oxidase |
| ONL | Outer nuclear la |
| OPL | Outer plexiform layer |
| PBS | Phosphate-buffered saline |
| RGC | Retinal ganglion cell |
| ROS | Reactive oxygen species |
| SNP | Sodium nitroprusside |
| TNF- α | Tumor necrosis factor alpha |
| U46619 | 9,11-dideoxy-9α,11α-methanoepoxy prostaglandin F2α |
| VEGF-A | Vascular endothelial growth factor |
References
- Gao, S.; Andreeva, K.; Cooper, N.G. Ischemia-reperfusion injury of the retina is linked to necroptosis via the ERK1/2-RIP3 pathway. Mol. Vis. 2014, 20, 1374–1387. [Google Scholar] [PubMed]
- Renner, M.; Stute, G.; Alzureiqi, M.; Reinhard, J.; Wiemann, S.; Schmid, H.; Faissner, A.; Dick, H.B.; Joachim, S.C. Optic Nerve Degeneration after Retinal Ischemia/Reperfusion in a Rodent Model. Front Cell Neurosci. 2017, 11, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumelt, S.; Dorenboim, Y.; Rehany, U. Aggressive systematic treatment for central retinal artery occlusion. Am. J. Ophthalmol. 1999, 128, 733–738. [Google Scholar] [CrossRef]
- Terelak-Borys, B.; Skonieczna, K.; Grabska-Liberek, I. Ocular ischemic syndrome—A systematic review. Med. Sci. Monit. 2012, 18, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Feltgen, N.; Neubauer, A.; Jurklies, B.; Schmoor, C.; Schmidt, D.; Wanke, J.; Maier-Lenz, H.; Schumacher, M.; Group, E.A.-S. Multicenter study of the European assessment group for lysis in the eye (EAGLE) for the treatment of central retinal artery occlusion: Design issues and implications. EAGLE Study report no. 1: EAGLE Study report no. 1. Graefes. Arch. Clin. Exp. Ophthalmol. 2006, 244, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.; Schmidt, D.; Jurklies, B.; Gall, C.; Wanke, I.; Schmoor, C.; Maier-Lenz, H.; Solymosi, L.; Brueckmann, H.; Neubauer, A.S.; et al. Central retinal artery occlusion: Local intra-arterial fibrinolysis versus conservative treatment, a multicenter randomized trial. Ophthalmology 2010, 117, 1367–1375. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Alloush, J.; Beck, E.; Weisleder, N. A murine model of myocardial ischemia-reperfusion injury through ligation of the left anterior descending artery. J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [PubMed]
- Masuzawa, K.; Jesmin, S.; Maeda, S.; Kaji, Y.; Oshika, T.; Zaedi, S.; Shimojo, N.; Yaji, N.; Miyauchi, T.; Goto, K. A model of retinal ischemia-reperfusion injury in rats by subconjunctival injection of endothelin-1. Exp. Biol. Med. (Maywood) 2006, 231, 1085–1089. [Google Scholar] [PubMed]
- Larpthaveesarp, A.; Gonzalez, F.F. Transient middle cerebral artery occlusion model of neonatal stroke in P10 Rats. J. Vis. Exp. 2017. [Google Scholar] [CrossRef]
- Hartsock, M.J.; Cho, H.; Wu, L.; Chen, W.J.; Gong, J.; Duh, E.J. A mouse model of retinal ischemia-reperfusion injury through elevation of intraocular pressure. J. Vis. Exp. 2016. [Google Scholar] [CrossRef]
- Hein, T.W.; Ren, Y.; Potts, L.B.; Yuan, Z.; Kuo, E.; Rosa, R.H., Jr.; Kuo, L. Acute retinal ischemia inhibits endothelium-dependent nitric oxide-mediated dilation of retinal arterioles via enhanced superoxide production. Invest Ophthalmol. Vis. Sci. 2012, 53, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.Y.; Zhang, C.; Li, Z.G. Early activation of caspase-1 after retinal ischemia and reperfusion injury in mice. Chin. Med. J. 2004, 117, 717–721. [Google Scholar]
- Aktas, Z.; Gurelik, G.; Akyurek, N.; Onol, M.; Hasanreisoglu, B. Neuroprotective effect of topically applied brimonidine tartrate 0.2% in endothelin-1-induced optic nerve ischaemia model. Clin. Exp. Ophthalmol. 2007, 35, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg-Cohen, N.; Dadon, S.; Avraham, B.C.; Kramer, M.; Hasanreisoglu, M.; Eldar, I.; Weinberger, D.; Bahar, I. Molecular and histological changes following central retinal artery occlusion in a mouse model. Exp. Eye Res. 2008, 87, 327–333. [Google Scholar] [CrossRef]
- Masuda, T.; Shimazawa, M.; Hara, H. Retinal Diseases Associated with Oxidative Stress and the Effects of a Free Radical Scavenger (Edaravone). Oxid. Med. Cell. Longev. 2017, 2017, 9208489. [Google Scholar] [CrossRef]
- Kruk, J.; Kubasik-Kladna, K.; Aboul-Enein, H.Y. The role oxidative stress in the pathogenesis of eye diseases: Current status and a dual role of physical activity. Mini. Rev. Med. Chem. 2015, 16, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Carden, D.L.; Granger, D.N. Pathophysiology of ischaemia-reperfusion injury. J. Pathol. 2000, 190, 255–266. [Google Scholar] [CrossRef]
- Kowalczyk, A.; Kleniewska, P.; Kolodziejczyk, M.; Skibska, B.; Goraca, A. The role of endothelin-1 and endothelin receptor antagonists in inflammatory response and sepsis. Arch. Immunol. Ther. Exp. (Warsz) 2015, 63, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, I.; Martin, R.; Ussa, F.; Fernandez-Bueno, I. The parameters of the porcine eyeball. Graefes Arch. Clin. Exp. Ophthalmol. 2011, 249, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ederra, J.; Garcia, M.; Hernandez, M.; Urcola, H.; Hernandez-Barbachano, E.; Araiz, J.; Vecino, E. The pig eye as a novel model of glaucoma. Exp. Eye Res. 2005, 81, 561–569. [Google Scholar] [CrossRef]
- Galdos, M.; Bayon, A.; Rodriguez, F.D.; Mico, C.; Sharma, S.C.; Vecino, E. Morphology of retinal vessels in the optic disk in a Gottingen minipig experimental glaucoma model. Vet. Ophthalmol. 2012, 15, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.; Johnston, E. The topography of primate retina: A study of the human, bushbaby, and new- and old-world monkeys. J. Comp. Neurol. 1981, 196, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Onken, M.; Berger, S.; Kristian, T. Simple model of forebrain ischemia in mouse. J. Neurosci. Methods 2012, 204, 254–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, T.; Messing, R.O.; Chou, W.H. Mouse model of middle cerebral artery occlusion. J. Vis. Exp. 2011. [Google Scholar] [CrossRef] [PubMed]
- Block, F.; Schwarz, M.; Sontag, K.H. Retinal ischemia induced by occlusion of both common carotid arteries in rats as demonstrated by electroretinography. Neurosci. Lett. 1992, 144, 124–126. [Google Scholar] [CrossRef]
- Bukhari, S.M.; Kiu, K.Y.; Thambiraja, R.; Sulong, S.; Rasool, A.H.; Liza-Sharmini, A.T. Microvascular endothelial function and severity of primary open angle glaucoma. Eye 2016, 30, 1579–1587. [Google Scholar] [CrossRef]
- Resch, H.; Garhofer, G.; Fuchsjager-Mayrl, G.; Hommer, A.; Schmetterer, L. Endothelial dysfunction in glaucoma. Acta Ophthalmol. 2009, 87, 4–12. [Google Scholar] [CrossRef]
- Lim, L.S.; Ling, L.H.; Ong, P.G.; Foulds, W.; Tai, E.S.; Wong, E.; Lee, S.Y.; Wong, D.; Cheung, C.M.; Wong, T.Y. Dynamic responses in retinal vessel caliber with flicker light stimulation in eyes with diabetic retinopathy. Invest. Ophthalmol. Vis. Sci. 2014, 55, 5207–5213. [Google Scholar] [CrossRef]
- Cheng, L.; Yu, H.; Yan, N.; Lai, K.; Xiang, M. Hypoxia-inducible factor-1alpha target genes contribute to retinal neuroprotection. Front. Cell. Neurosci. 2017, 11. [Google Scholar] [CrossRef]
- Mishra, A.; Newman, E.A. Inhibition of inducible nitric oxide synthase reverses the loss of functional hyperemia in diabetic retinopathy. Glia 2010, 58, 1996–2004. [Google Scholar] [CrossRef] [Green Version]
- Carr, B.C.; Emigh, C.E.; Bennett, L.D.; Pansick, A.D.; Birch, D.G.; Nguyen, C. Towards a treatment for diabetic retinopathy: Intravitreal toxicity and preclinical safety evaluation of inducible nitric oxide synthase inhibitors. Retina 2017, 37, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Yao, Q.; Wang, W.; Yao, H.; Chao, J. iNOS Induces vascular endothelial cell migration and apoptosis via autophagy in ischemia/reperfusion injury. Cell. Physiol. Biochem. 2016, 38, 1575–1588. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.C.; van Wijngaarden, P.; Liu, G.S.; Jiang, F.; Peshavariya, H.; Dusting, G.J. Involvement of Nox2 NADPH oxidase in retinal neovascularization. Invest. Ophthalmol. Vis. Sci. 2013, 54, 7061–7067. [Google Scholar] [CrossRef] [PubMed]
- Zadeh, J.K.; Ruemmler, R.; Hartmann, E.K.; Ziebart, A.; Ludwig, M.; Patzak, A.; Xia, N.; Li, H.; Pfeiffer, N.; Gericke, A. Responses of retinal arterioles and ciliary arteries in pigs with acute respiratory distress syndrome (ARDS). Exp. Eye Res. 2019, 184, 152–162. [Google Scholar] [CrossRef]
- Yuan, G.; Khan, S.A.; Luo, W.; Nanduri, J.; Semenza, G.L.; Prabhakar, N.R. Hypoxia-inducible factor 1 mediates increased expression of NADPH oxidase-2 in response to intermittent hypoxia. J. Cell. Physiol. 2011, 226, 2925–2933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanduri, J.; Vaddi, D.R.; Khan, S.A.; Wang, N.; Makarenko, V.; Semenza, G.L.; Prabhakar, N.R. HIF-1alpha activation by intermittent hypoxia requires NADPH oxidase stimulation by xanthine oxidase. PLoS ONE 2015, 10, e0119762. [Google Scholar] [CrossRef]
- Kurihara, T.; Westenskow, P.D.; Friedlander, M. Hypoxia-inducible factor (HIF)/vascular endothelial growth factor (VEGF) signaling in the retina. Adv. Exp. Med. Biol. 2014, 801, 275–281. [Google Scholar] [PubMed]
- Ritzel, R.M.; Pan, S.J.; Verma, R.; Wizeman, J.; Crapser, J.; Patel, A.R.; Lieberman, R.; Mohan, R.; McCullough, L.D. Early retinal inflammatory biomarkers in the middle cerebral artery occlusion model of ischemic stroke. Mol. Vis. 2016, 22, 575–588. [Google Scholar] [PubMed]
- Pan, L.; Yang, Z.; Feng, L.; Gan, L. Functional equivalence of Brn3 POU-domain transcription factors in mouse retinal neurogenesis. Development 2005, 132, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Garca, M.; Ruiz-Ederra, J.; Hernandez-Barbachano, H.; Vecino, E. Topography of pig retinal ganglion cells. J. Comp. Neurol. 2005, 486, 361–372. [Google Scholar] [CrossRef]
- Hebel, R. Distribution of retinal ganglion cells in five mammalian species (pig, sheep, ox, horse, dog). Anat. Embryol. (Berl) 1976, 150, 45–51. [Google Scholar] [PubMed]
- Bardy, M.; Tsacopoulos, M. Metabolic changes in the retina after experimental microembolism in the miniature pig (author’s transl). Klin. Monbl. Augenheilkd. 1978, 172, 451–460. [Google Scholar] [PubMed]
- Osborne, N.N.; Block, F.; Sontag, K.H. Reduction of ocular blood flow results in glial fibrillary acidic protein (GFAP) expression in rat retinal Muller cells. Vis. Neurosci. 1991, 7, 637–639. [Google Scholar] [CrossRef] [PubMed]
- Hayreh, S.S.; Kolder, H.E.; Weingeist, T.A. Central retinal artery occlusion and retinal tolerance time. Ophthalmology 1980, 87, 75–78. [Google Scholar] [CrossRef]
- Hayreh, S.S.; Zimmerman, M.B.; Kimura, A.; Sanon, A. Central retinal artery occlusion. retinal survival time. Exp. Eye Res. 2004, 78, 723–736. [Google Scholar] [CrossRef]
- Hayreh, S.S.; Weingeist, T.A. Experimental occlusion of the central artery of the retina. IV: Retinal tolerance time to acute ischaemia. Br. J. Ophthalmol. 1980, 64, 818–825. [Google Scholar] [CrossRef]
- Leeb-Lundberg, L.M.; Marceau, F.; Muller-Esterl, W.; Pettibone, D.J.; Zuraw, B.L. International union of pharmacology. XLV. Classification of the kinin receptor family: From molecular mechanisms to pathophysiological consequences. Pharmacol. Rev. 2005, 57, 27–77. [Google Scholar] [CrossRef] [PubMed]
- Dalsgaard, T.; Kroigaard, C.; Bek, T.; Simonsen, U. Role of calcium-activated potassium channels with small conductance in bradykinin-induced vasodilation of porcine retinal arterioles. Invest. Ophthalmol. Vis. Sci. 2009, 50, 3819–3825. [Google Scholar] [CrossRef]
- Friederich, J.A.; Butterworth, J.F. t. Sodium nitroprusside: Twenty years and counting. Anesth. Analg. 1995, 81, 152–162. [Google Scholar]
- Coleman, R.A.; Humphrey, P.P.; Kennedy, I.; Levy, G.P.; Lumley, P. Comparison of the actions of U-46619, a prostaglandin H2-analogue, with those of prostaglandin H2 and thromboxane A2 on some isolated smooth muscle preparations. Br. J. Pharmacol. 1981, 73, 773–778. [Google Scholar] [CrossRef]
- Nadal-Nicolas, F.M.; Jimenez-Lopez, M.; Sobrado-Calvo, P.; Nieto-Lopez, L.; Canovas-Martinez, I.; Salinas-Navarro, M.; Vidal-Sanz, M.; Agudo, M. Brn3a as a marker of retinal ganglion cells: Qualitative and quantitative time course studies in naive and optic nerve-injured retinas. Invest. Ophthalmol. Vis. Sci. 2009, 50, 3860–3868. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Romero, C.; Aviles-Trigueros, M.; Jimenez-Lopez, M.; Valiente-Soriano, F.J.; Salinas-Navarro, M.; Nadal-Nicolas, F.; Villegas-Perez, M.P.; Vidal-Sanz, M.; Agudo-Barriuso, M. Axotomy-induced retinal ganglion cell death in adult mice: Quantitative and topographic time course analyses. Exp. Eye Res. 2011, 92, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Gericke, A.; Mayer, V.G.; Steege, A.; Patzak, A.; Neumann, U.; Grus, F.H.; Joachim, S.C.; Choritz, L.; Wess, J.; Pfeiffer, N. Cholinergic responses of ophthalmic arteries in M3 and M5 muscarinic acetylcholine receptor knockout mice. Invest. Ophthalmol. Vis. Sci. 2009, 50, 4822–4827. [Google Scholar] [CrossRef] [PubMed]
- Manicam, C.; Staubitz, J.; Brochhausen, C.; Grus, F.H.; Pfeiffer, N.; Gericke, A. The gatekeepers in the mouse ophthalmic artery: Endothelium-dependent mechanisms of cholinergic vasodilation. Sci. Rep. 2016, 6, 20322. [Google Scholar] [CrossRef]
- Laspas, P.; Zhutdieva, M.B.; Brochhausen, C.; Musayeva, A.; Zadeh, J.K.; Pfeiffer, N.; Xia, N.; Li, H.; Wess, J.; Gericke, A. The M1 muscarinic acetylcholine receptor subtype is important for retinal neuron survival in aging mice. Sci. Rep. 2019, 9, 5222. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Teister, J.; Anders, F.; Beck, S.; Funke, S.; von Pein, H.; Prokosch, V.; Pfeiffer, N.; Grus, F. Decelerated neurodegeneration after intravitreal injection of alpha-synuclein antibodies in a glaucoma animal model. Sci. Rep. 2017, 7, 6260. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diameter Changes [%] | |||||
|---|---|---|---|---|---|
| Baseline [µm] | Myogenic Tone [%] | U46619 at 10−6 M[%] | SNP at 10−4 M [%] | Bradykinin at 10−7 M [%] | |
| Sham | 85.5 ± 20.6 | −46.1 ± 5.79 | −41.6 ± 9.89 | 42.3 ± 5.62 | 82.0 ± 8.85 |
| I/R | 91.8 ± 24.7 | −42.7 ± 4.18 | −35.2 ± 5.11 | 35.6 ± 8.06 | 34.5 ± 8.02 |
| Gene Name | Primer Sequence |
|---|---|
| NOX1 | F: TCAGTTTTATTTCTGGCTGCTTGG R: CTTTCTCAGGGTGCGCCTAC |
| NOX2 | F: CACTTCACGCCACGATTCAC R: TTGACTCGGGCGTTCACAC |
| NOX4 | F: GTCCCAGTGTGTCTGCGTTAG R: TCTCGAAATCGTTCTGTCCAGTC |
| eNOS | F: CTACAGGACCCAAGATGGGC R: TGAAGCAGGGTACAGGGTCT |
| nNOS | F: ATTTTCGGAGTCACCCTGCG R: AGCTGAAAACCTCATCTGTGTCT |
| iNOS | F: ACTATTTCTTCCAGCTTAAGAGCC R: CTCGTAGGGAAATACAGCACCA |
| TNF-α | F: TTCTGCCTACTGCACTTCGAG R: TGAGACGATGATCTGAGTCCTT |
| IL-1β | F: ATAGTACCTGAACCCGCCAAG R: GTGCAGCACTTCATCTCTTTGG |
| HIF-1α | F: CGTGCGACCATGAGGAAATG R: GTGAAGTACTTTCCATGTTGCAG |
| VEGF-A | F: ATAGAGCGAGGCAAGAAAATCCC R: ACACGTCTGCGGATCTTGTA |
| β-actin | F: TGGACTACCTCCTGTCTGCT R:CCTAGGGGTGGGTTTCTGTG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zadeh, J.K.; Garcia-Bardon, A.; Hartmann, E.K.; Pfeiffer, N.; Omran, W.; Ludwig, M.; Patzak, A.; Xia, N.; Li, H.; Gericke, A. Short-Time Ocular Ischemia Induces Vascular Endothelial Dysfunction and Ganglion Cell Loss in the Pig Retina. Int. J. Mol. Sci. 2019, 20, 4685. https://doi.org/10.3390/ijms20194685
Zadeh JK, Garcia-Bardon A, Hartmann EK, Pfeiffer N, Omran W, Ludwig M, Patzak A, Xia N, Li H, Gericke A. Short-Time Ocular Ischemia Induces Vascular Endothelial Dysfunction and Ganglion Cell Loss in the Pig Retina. International Journal of Molecular Sciences. 2019; 20(19):4685. https://doi.org/10.3390/ijms20194685
Chicago/Turabian StyleZadeh, Jenia Kouchek, Andreas Garcia-Bardon, Erik Kristoffer Hartmann, Norbert Pfeiffer, Wael Omran, Marion Ludwig, Andreas Patzak, Ning Xia, Huige Li, and Adrian Gericke. 2019. "Short-Time Ocular Ischemia Induces Vascular Endothelial Dysfunction and Ganglion Cell Loss in the Pig Retina" International Journal of Molecular Sciences 20, no. 19: 4685. https://doi.org/10.3390/ijms20194685