Influence of the Type of Diet on the Incidence of Pathogenic Factors and Antibiotic Resistance in Enterococci Isolated from Faeces in Mice

 , ,
, ,
Abstract
:1. Introduction
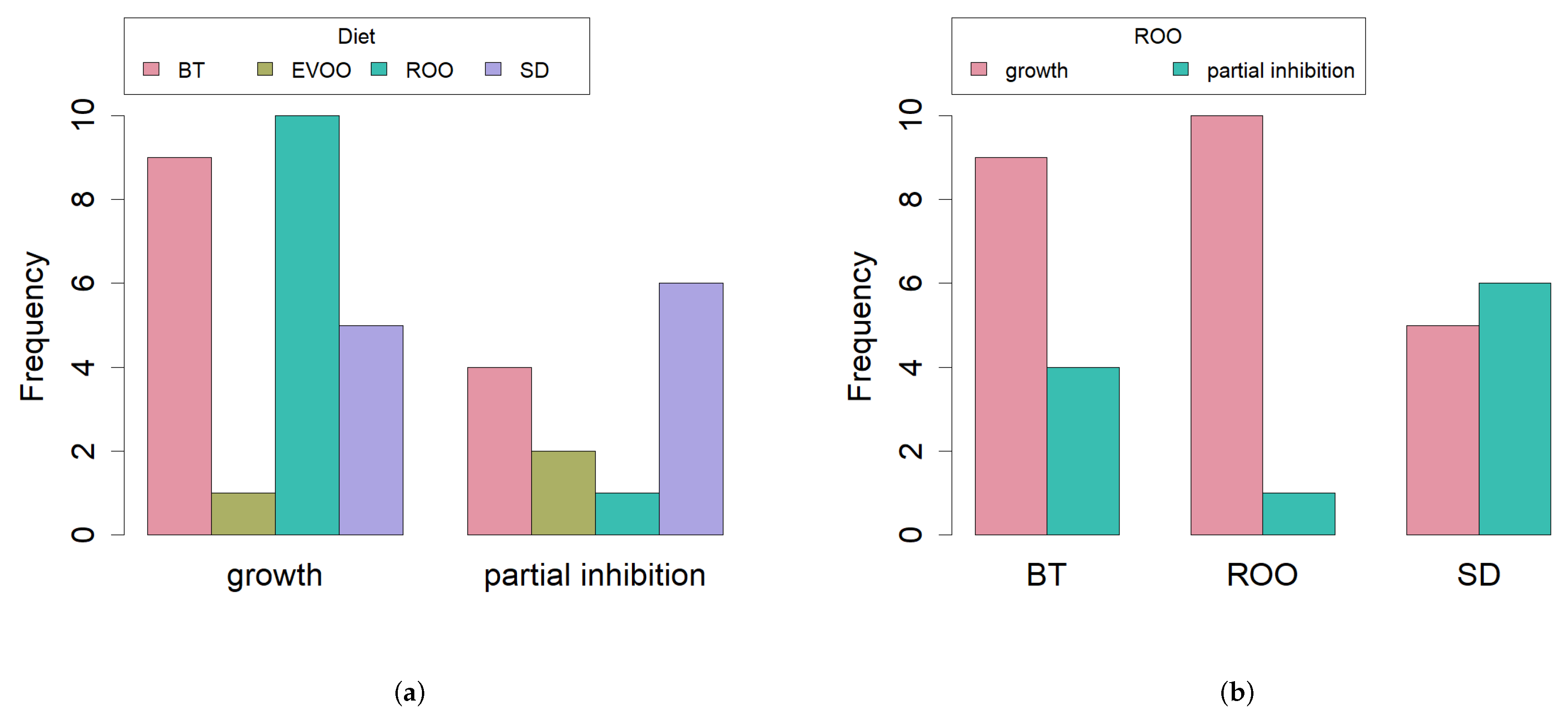
2. Results
2.1. Isolation and Identification of Bacterial Strains
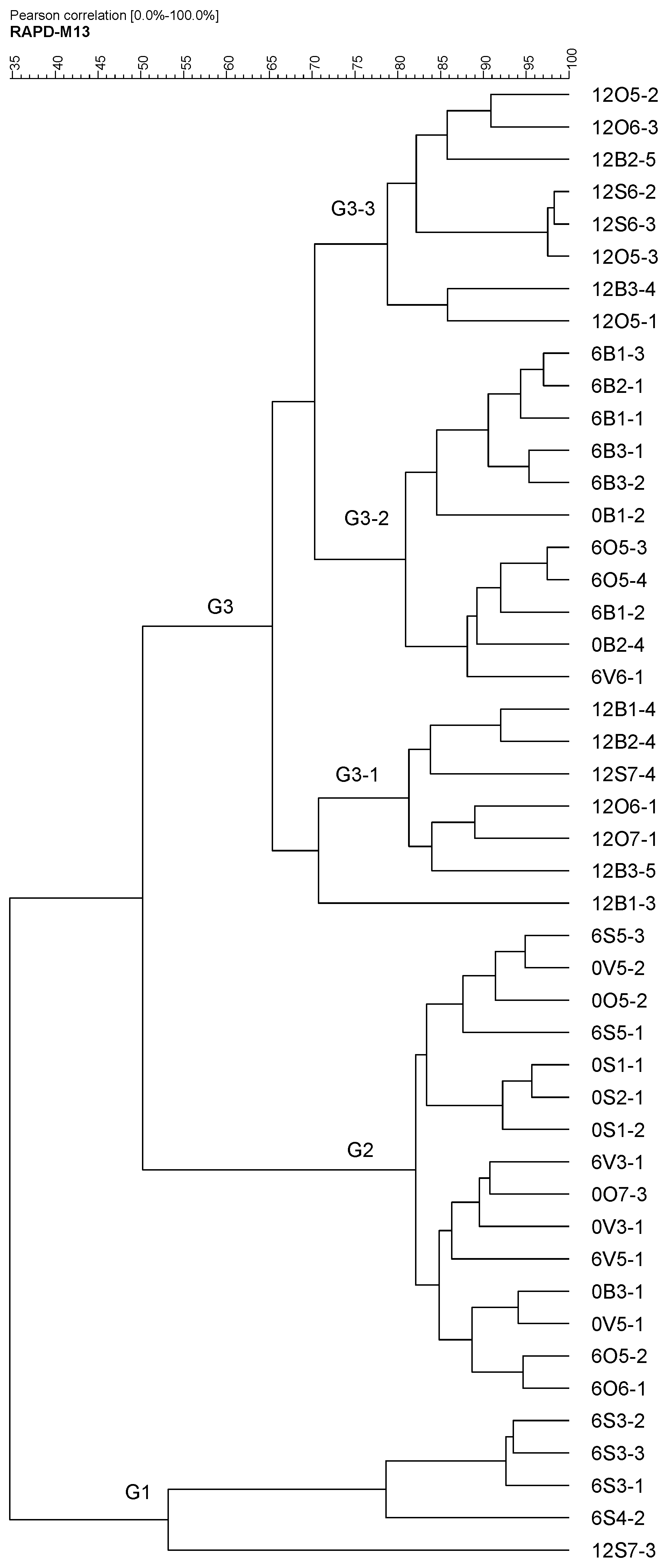
2.2. RAPD Classification
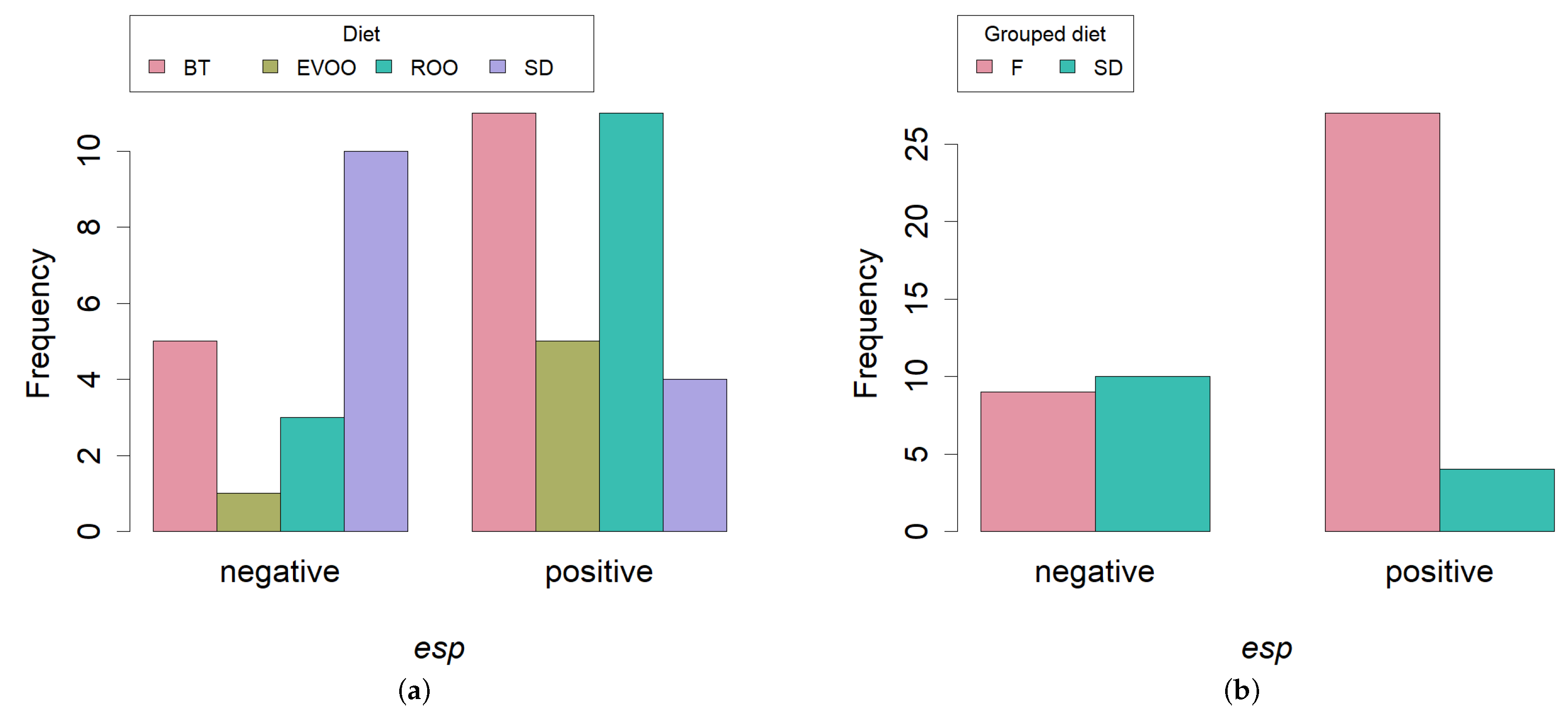
2.3. PCR Amplification of Virulence Factors
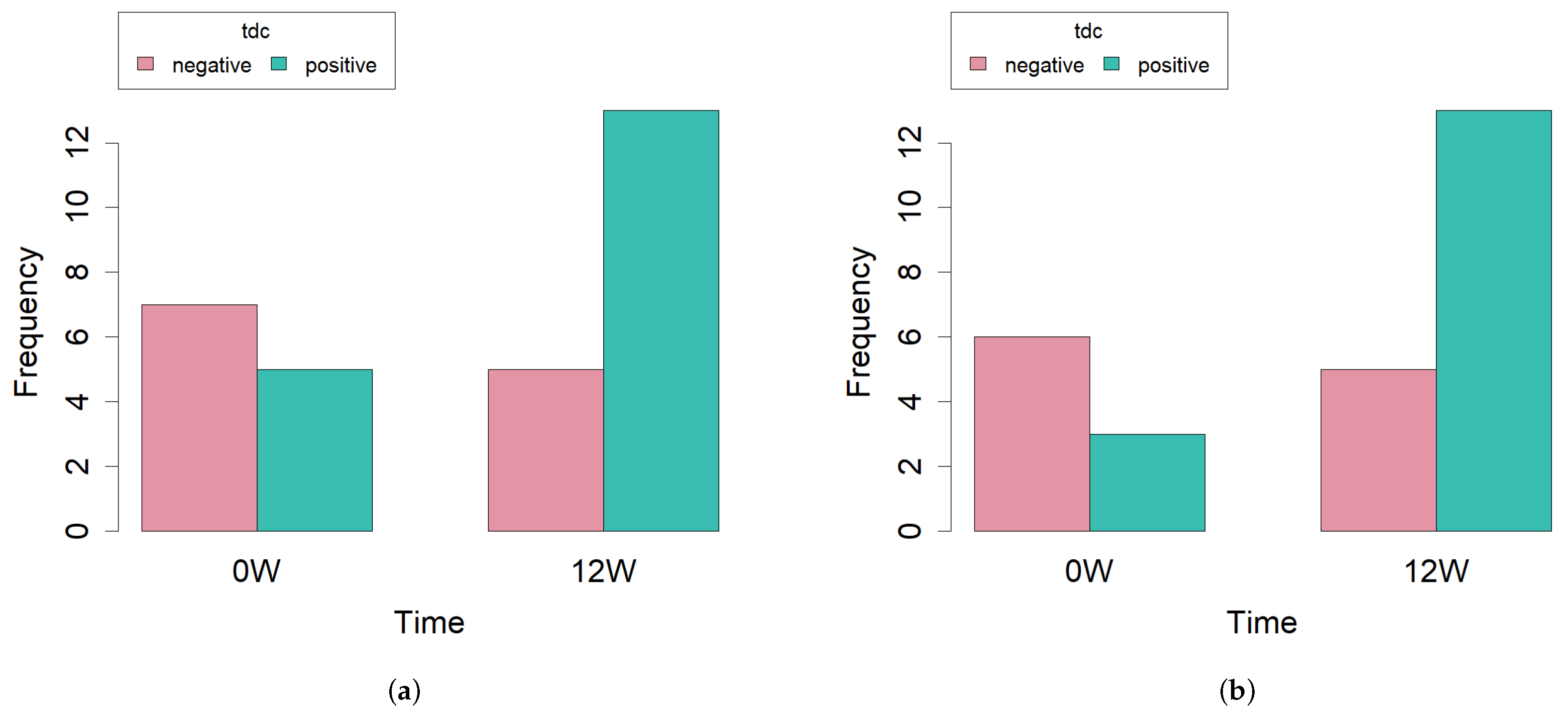
2.4. Biogenic Amine Production
2.5. Antibiotic Resistance
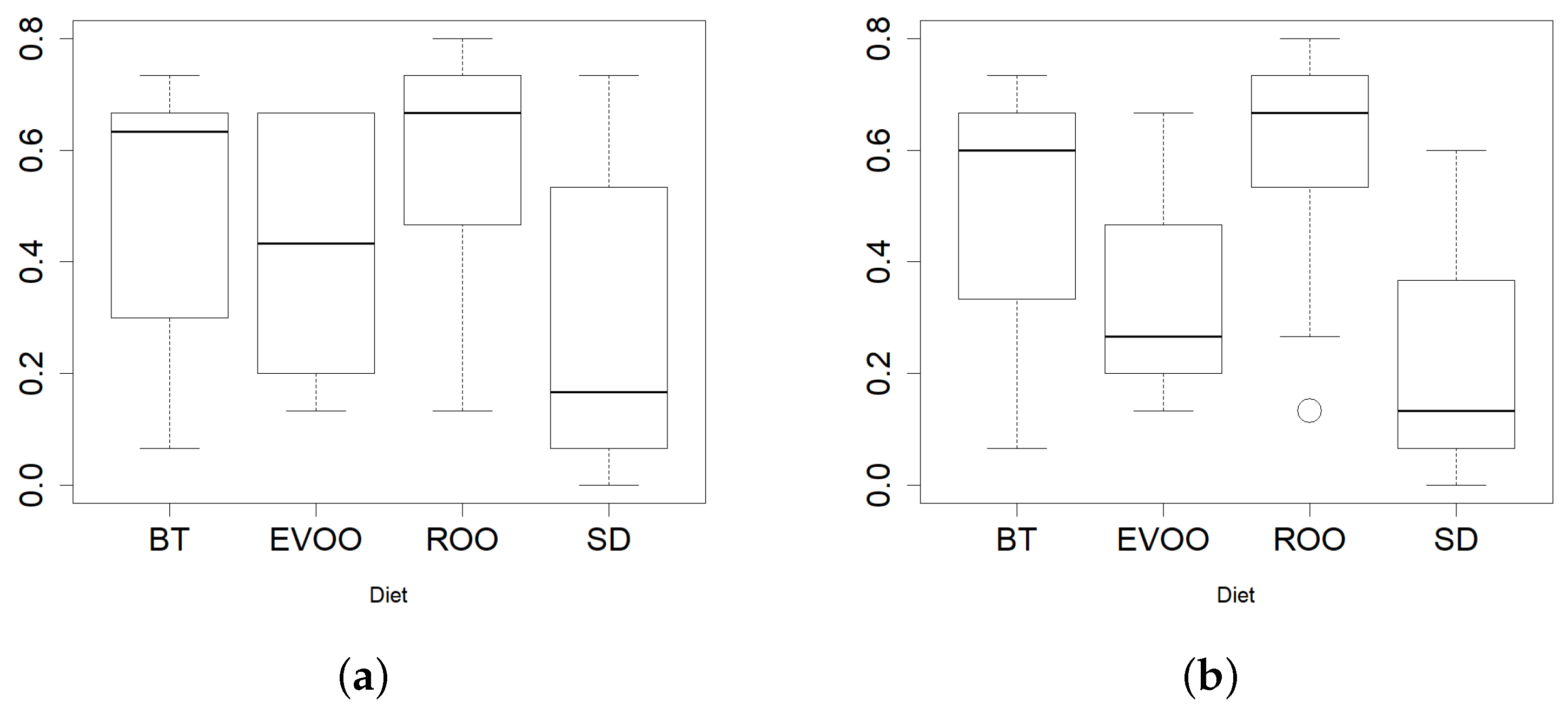
2.6. Growth with Virgin and Refined Olive Oil
3. Discussion
4. Materials and Methods
4.1. Isolation and Identification of Bacterial Strains
4.2. RAPD-PCR Amplification
4.3. PCR Amplification of Virulence Factors
4.4. Biogenic Amine Production
4.5. Antibiotic Resistance
4.6. Growth with Virgin and Refined Olive Oil
4.7. Statistical Studies
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, C.; Zhang, M.; Wang, S.; Han, R.; Cao, Y.; Hua, W. Interactions between gut microbiota, host genetics and diet relevant to development of metabolic syndromes in mice. ISME J. 2010, 4, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Albenberg, L.G.; Wu, G.D. Diet and the intestinal microbiome: associations, functions, and implications for health and disease. Gastroenterology 2014, 146, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Prieto, I.; Abriouel, H.; Cobo, A.; Benomar, N.; Gálvez, A.; Martínez-Cañamero, M. Effect of virgin and refined olive oil consumption on gut microbiota. Comparison to butter. Food Res. Int. 2014, 64, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Prieto, I.; Hidalgo, M.; Segarra, A.B.; Martínez-Rodríguez, A.M.; Cobo, A.; Ramírez, M.; Abriouel, H.; Gálvez, A.; Martínez-Cañamero, M. Influence of a diet enriched in virgin olive oil or butter on mouse gut microbiota and its correlation to physiological and biochemical parameters related to metabolic syndrome. PLoS ONE 2018, 13, e0190368. [Google Scholar] [CrossRef] [PubMed]
- Martínez, N.; Prieto, P.; Hidalgo, M.; Segarra, A.B.; Martínez-Rodríguez, A.M.; Cobo-Molinos, A.; Ramírez, M.; Gálvez, A.; Martínez-Cañamero, M. Refined versus extra virgin olive oil high fat diet impact on intestinal microbiota of mice and its relation to different physiological variables. Microorganisms 2019, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Franz, C.M.; Huch, M.; Abriouel, H.; Holzapfel, W.; Gálvez, A. Enterococci as probiotics and their implications in food safety. Int. J. Food Microbiol. 2011, 151, 125–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Omar, N.; Castro, A.; Lucas, R.; Abriouel, H.; Yousif, N.M.K.; Franz, C.M.A.P.; Holzapfel, W.H.; Pérez-Pulido, R.; Martínez-Cañamero, M.; Gálvez, A. Functional and safety aspects of enterococci isolated from different Spanish foods. Syst. Appl. Microbiol. 2004, 27, 118–130. [Google Scholar] [CrossRef]
- Ogier, J.C.; Serror, P. Safety assessment of dairy microorganisms: the Enterococcus genus. Int. J. Food Microbiol. 2008, 126, 291–301. [Google Scholar] [CrossRef]
- Tannock, G.W.; Cook, G. Enterococci as members of the intestinal microflora of humans. In The Enterococci: Pathogenesis, Molecular Biology and Antibiotic Resistance; Gilmore, M.S., Ed.; ASM Press: Washington, DC, USA, 2002; pp. 101–132. [Google Scholar]
- Abriouel, H.; Ben Omar, N.; Cobo-Molinos, A.; Lucas López, R.; Grande, M.J.; Martínez-Viedma, P.; Ortega, E.; Martínez-Cañamero, M.; Gálvez, A. Comparative analysis of genetic diversity and incidence of virulence factors and antibiotic resistance among enterococcal populations from raw fruit and vegetable foods, water and soil, and clinical samples. Int. J. Food Microbiol. 2008, 123, 38–49. [Google Scholar] [CrossRef]
- Pérez-Pulido, R.; Omar, N.B.; Lucas, R.; Abriouel, H.; Martínez-Cañamero, M.; Gálvez, A. Resistance to antimicrobial agents in lactobacilli isolated from caper fermentations. Antonie Van Leeuwenhoek 2005, 88, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Cobo-Molinos, A.; Abriouel, H.; Omar, N.B.; López, R.L.; Galvez, A. Detection of ebp (endocarditis- and biofilm-associated pilus) genes in enterococcal isolates from clinical and non-clinical origin. Int. J. Food Microbiol. 2008, 126, 123–126. [Google Scholar] [CrossRef]
- Medina, E.; de Castro, A.; Romero, C.; Brenes, M. Comparison of the concentrations of phenolic compounds in olive oils and other plant oils: correlation with antimicrobial activity. J. Agric. Food Chem. 2006, 12, 4954–4961. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell. Probes. 2001, 15, 209–215. [Google Scholar] [CrossRef]
- Sáenz, Y.; Briñas, L.; Domínguez, E.; Ruiz, J.; Zarazaga, M.; Vila, J.; Torres, C. Mechanisms of Resistance in Multiple-Antibiotic-Resistant Escherichia coli Strains of Human, Animal, and Food Origins. Antimicrob. Agents Chemother. 2004, 48, 3996–4001. [Google Scholar] [CrossRef] [PubMed]
- Eaton, T.J.; Gasson, M.J. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl. Environ. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef]
- Toledo-Arana, A.; Valle, J.; Solano, C.; Arrizubieta, M.J.; Cucarella, C.; Lamata, M. The enterococcal surface protein, esp, is involved in Enterococcus faecalis biofilm formation. Appl. Environ. Microbiol. 2001, 67, 4538–4545. [Google Scholar] [CrossRef]
- Gardini, F.; Martuscelli, M.; Caruso, M.C.; Galgano, F.; Crudele, M.A.; Favati, F.; Guerzoni, M.E.; Suzzi, G. Effects of pH, temperature and NaCl concentration on growth kinetics, proteolytic activity and biogenic amine production of Enterococcus faecalis. Int. J. Food Microbiol. 2001, 64, 105–117. [Google Scholar] [CrossRef]
- Mariné-Font, A.; Vidal-Carou, M.C.; Izquierdo-Pulido, M.; Veciana-Noguès, M.T.; Hernández-Jover, T. Les amines biogènes dans les aliments: leur signification, leur analyse. Ann. Falsif. Expert Chim. Toxicol. 1995, 88, 119–140. [Google Scholar]
- Pérez-Pulido, R.; Abriouel, H.; Ben Omar, N.; Lucas, R.; Martínez-Cañamero, M.; Gálvez, A. Safety and potential risks of enterococci isolated from traditional fermented capers. Food Chem. Toxicol. 2006, 44, 2070–2077. [Google Scholar] [CrossRef]
- Giraffa, G.; Carminati, D.; Neviani, E. Enterococci isolated from dairy products: A review of risks and potential technological use. J. Food Protect. 1997, 60, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, A.R. Significance of biogenic amines to food safety and human health. Food Res. Int. 1996, 29, 675–690. [Google Scholar] [CrossRef]
- Pacheco, Y.M.; López, S.; Bermúdez, B.; Abia, R.; Muriana, F.J. Extra-virgin vs. refined olive oil on postprandial hemostatic markers in healthy subjects. J. Thromb. Haemost. 2006, 4, 1421–1422. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Lucas, R.; Ben Omar, N.; Valdivia, E.; Maqueda, M.; Martínez-Cañamero, M.; Gálvez, A. Enterocin AS-48RJ: A variant of enterocin AS-48 chromosomally encoded by Enterococcus faecium RJ16 isolated from food. Syst. Appl. Microbiol. 2005, 28, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Martín-Platero, A.M.; Valdivia, E.; Maqueda, M.; Martínez-Bueno, M. Characterization and safety evaluation of enterococci isolated from Spanish goats’ milk cheeses. Int. J. Food Microbiol. 2009, 132, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, S.; Weill, F.X.; Cloeckaert, A.; Vrints, M.; Mairiaux, E.; Praud, K.; Dierick, K.; Wildemauve, C.; Godard, C.; Butaye, P.; et al. Clonal emergence of extended-spectrum beta-lactamase (CTX-M-2)-producing Salmonella enterica serovar Virchow isolates with reduced susceptibilities to ciprofloxacin among poultry and humans in Belgium and France (2000 to 2003). J. Clin. Microbiol. 2006, 44, 2897–2903. [Google Scholar] [CrossRef] [PubMed]
- Chiu, CH.; Su, L.H.; Chu, CH.; Wang, M.H.; Yeh, C.M.; Weill, F.X.; Chu, C. Detection of multidrug-resistant Salmonella enterica serovar Typhimurium phage types DT102, DT104, and U302 by multiplex PCR. J. Clin. Microbiol. 2006, 44, 2354–2358. [Google Scholar] [CrossRef]
- Swick, M.C.; Morgan-Linnell, S.K.; Carlson, K.M.; Zechiedrich, L. Expression of multidrug efflux pump genes acrAB-tolC, mdfA, and norE in Escherichia coli clinical isolates as a function of fluoroquinolone and multidrug resistance. Antimicrob. Agents Chemother. 2011, 55, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Soto, S.M.; Argüelles, J.M.; Mendoza, M.C. Multidrug resistance is mediated by large plasmids carrying a class 1 integron in the emergent Salmonella enterica serotype [4,5,12:i:-]. Antimicrob. Agents Chemother. 2001, 45, 1305–1308. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virulence Factors | Biogenic Amines | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | agg | gelE | cylA | cylM | cylB | esp | efaAfs | efaAfm | cob | cpd | ccf | tdc | |
| 0S1-1 | E. faecalis | + | |||||||||||
| 0S1-2 | E. casseliflavus | + | |||||||||||
| 0S2-1 | E. faecalis | + | + | ||||||||||
| 0B1-2 | E. casseliflavus | + | |||||||||||
| 0B2-4 | E. faecalis | + | |||||||||||
| 0B3-1 | E. casseliflavus | + | |||||||||||
| 0O5-2 | E. faecalis | + | + | + | |||||||||
| 0O5-4 | E. faecalis | + | + | ||||||||||
| 0O7-3 | E. faecalis | + | |||||||||||
| 0V3-1 | E. casseliflavus | + | |||||||||||
| 0V5-1 | E. faecalis | + | + | ||||||||||
| 0V5-2 | E. faecalis | + | + | + | |||||||||
| 6S3-3 | E. casseliflavus | + | + | + | |||||||||
| 6S3-4 | E. gallinarum | + | |||||||||||
| 6S4-1 | E. casseliflavus | ||||||||||||
| 6S4-2 | E. faecalis | + | + | ||||||||||
| 6S5-1 | E. faecalis | + | |||||||||||
| 6S5-3 | E. faecalis | + | + | + | |||||||||
| 6B1-1 | E. casseliflavus | + | |||||||||||
| 6B1-2 | E. gallinarum | + | |||||||||||
| 6B1-3 | E. hirae | + | + | ||||||||||
| 6B2-1 | E. faecalis | + | + | ||||||||||
| 6B3-1 | E. avium | + | + | ||||||||||
| 6B3-2 | E. casseliflavus | + | |||||||||||
| 6O5-2 | E. faecalis | + | + | ||||||||||
| 6O5-3 | E. faecalis | + | |||||||||||
| 6O5-4 | E. faecalis | + | + | ||||||||||
| 6O6-1 | E. faecalis | + | + | ||||||||||
| 6O7-1 | E. casseliflavus | + | |||||||||||
| 6V3-1 | E. casseliflavus | + | |||||||||||
| 6V5-1 | E. faecalis | + | + | ||||||||||
| 6V6-1 | E. casseliflavus | + | + | ||||||||||
| 12S6-2 | E. faecalis | + | |||||||||||
| 12S6-3 | E. casseliflavus | + | + | ||||||||||
| 12S7-2 | E. casseliflavus | + | |||||||||||
| 12S7-3 | E. gallinarum | + | + | ||||||||||
| 12S7-4 | E. casseliflavus | + | + | ||||||||||
| 12B1-3 | E. hirae | + | |||||||||||
| 12B1-4 | E. faecalis | + | + | + | |||||||||
| 12B2-4 | E. casseliflavus | + | + | ||||||||||
| 12B2-5 | E. durans | ||||||||||||
| 12B3-2 | E. casseliflavus | + | |||||||||||
| 12B3-4 | E. faecalis | + | |||||||||||
| 12B3-5 | E. faecalis | + | |||||||||||
| 12O5-1 | E. gallinarum | + | + | ||||||||||
| 12O5-2 | E. casseliflavus | + | |||||||||||
| 12O5-3 | E. gallinarum | + | |||||||||||
| 12O6-1 | E. casseliflavus | + | + | ||||||||||
| 12O6-3 | E. casseliflavus | + | |||||||||||
| 12O7-1 | E. casseliflavus | + | |||||||||||
| Antibiotic Resistance | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PEN | PENS | AMPE | CTXP | CTXS/IMIE | KAHES/GEHES | FQPR/MXFPS | LVXS/LVXP | ERYPS/TELPS | QDAE | TETPS/RFAPS | TSU/LNZEP | LNZS | FOSP7FURES | VAN/TEC | |
| 0S1-1 | R | R | I | R | I | S | S | S | R | R | R | R | R | I | R |
| 0S1-2 | R | R | I | R | I | S | S | S | R | R | I | I | R | S | R |
| 0S2-1 | R | R | R | R | I | S | S | S | R | R | R | R | R | R | R |
| 0B1-2 | I | S | S | R | I | S | S | S | S | S | S | S | S | S | S |
| 0B2-4 | R | R | R | R | I | S | S | S | R | R | R | R | R | R | R |
| 0B3-1 | R | R | R | I | I | S | I | S | R | R | R | R | R | R | R |
| 0O5-2 | R | R | R | R | R | S | S | S | R | R | R | I | R | R | R |
| 0O5-4 | R | I | S | R | I | S | S | I | S | I | R | S | I | I | S |
| 0O7-3 | R | R | I | R | I | S | S | S | R | R | R | I | R | I | R |
| 0V3-1 | R | R | R | I | I | S | S | S | R | R | R | I | R | R | R |
| 0V5-1 | R | R | R | R | I | S | S | S | R | R | R | I | R | R | R |
| 0V5-2 | I | I | S | R | I | S | S | S | I | R | R | S | S | S | S |
| 6S3-3 | R | R | I | I | I | S | S | S | I | S | S | S | S | S | S |
| 6S3-4 | I | S | S | I | I | S | S | S | S | S | S | S | S | S | S |
| 6S4-1 | R | R | I | R | I | S | S | S | R | R | R | I | R | S | R |
| 6S4-2 | R | R | S | R | I | S | S | S | I | S | S | S | S | S | S |
| 6S5-1 | I | I | S | I | I | S | S | S | I | I | I | S | S | I | S |
| 6S5-3 | I | I | S | S | S | S | S | S | I | I | R | S | S | I | S |
| 6B1-1 | R | I | S | R | I | I | S | S | I | R | R | S | S | I | S |
| 6B1-2 | R | R | R | R | I | S | I | S | R | R | R | I | R | R | R |
| 6B1-3 | I | S | S | R | I | S | S | S | I | I | I | S | I | S | S |
| 6B2-1 | R | R | R | R | R | R | R | R | R | S | I | I | S | S | S |
| 6B3-1 | R | R | R | R | I | S | R | S | R | R | R | I | R | R | R |
| 6B3-2 | R | I | S | R | I | S | S | S | R | R | R | S | S | I | S |
| 6O5-2 | R | R | R | I | S | S | S | R | R | R | R | I | R | R | R |
| 6O5-3 | R | R | R | R | I | S | S | S | R | R | R | I | R | R | R |
| 6O5-4 | R | R | R | R | R | S | S | S | R | R | R | I | R | R | R |
| 6O6-1 | R | R | R | R | I | S | S | S | R | R | R | I | R | R | R |
| 6O7-1 | R | I | S | R | I | S | S | S | I | R | R | S | S | S | S |
| 6V3-1 | R | R | R | R | I | S | I | S | R | R | R | I | R | R | R |
| 6V5-1 | I | I | S | R | I | S | S | S | R | R | R | S | S | I | S |
| 6V6-1 | I | I | S | I | I | S | S | S | I | R | R | S | S | S | S |
| 12S6-2 | R | R | I | S | S | S | S | S | R | R | R | R | R | I | R |
| 12S6-3 | I | I | S | R | I | S | S | S | I | I | I | S | S | I | S |
| 12S7-2 | I | I | S | R | I | S | S | S | I | I | I | S | S | I | S |
| 12S7-3 | R | R | S | I | S | S | S | S | I | S | S | I | S | S | S |
| 12S7-4 | R | R | I | R | I | S | S | S | R | R | R | R | R | S | R |
| 12B1-3 | R | R | R | R | R | R | R | R | R | S | R | I | S | S | S |
| 12B1-4 | R | R | R | R | R | R | R | R | R | S | R | I | S | S | S |
| 12B2-4 | R | I | S | R | I | S | S | S | I | R | R | S | S | R | S |
| 12B2-5 | R | I | S | R | I | S | S | S | I | R | R | S | S | R | S |
| 12B3-2 | R | R | R | R | R | R | R | R | R | S | R | I | S | S | S |
| 12B3-4 | R | R | R | R | R | R | R | R | R | I | R | S | S | I | I |
| 12B3-5 | I | I | S | I | I | S | S | S | I | S | R | S | S | I | S |
| 12O5-1 | R | R | S | R | I | S | S | S | R | R | R | I | R | I | S |
| 12O5-2 | R | R | R | R | R | S | R | I | R | R | R | I | R | R | R |
| 12O5-3 | I | S | S | R | I | S | S | S | S | S | R | S | S | I | S |
| 12O6-1 | R | R | R | R | R | S | S | S | R | R | R | I | R | R | R |
| 12O6-3 | R | R | R | R | I | S | S | S | R | R | R | I | R | S | R |
| 12O7-1 | R | R | R | R | R | S | S | S | R | R | R | I | R | R | R |
| -Lactamases | Tetracyclines | Efflux Pumps | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Group I | Group II | ||||||||||||
| bla | bla | bla | bla | tetB | tetC | tetD | tetA | tetE | tetG | acrB | aadA1 | mdfA | |
| 0S1-1 | + | + | + | + | + | ||||||||
| 0S1-2 | + | + | + | + | + | + | + | ||||||
| 0S2-1 | + | + | + | + | + | ||||||||
| 0B1-2 | + | ||||||||||||
| 0B2-4 | + | + | |||||||||||
| 0B3-1 | + | + | |||||||||||
| 0O5-2 | + | + | + | ||||||||||
| 0O5-4 | + | ||||||||||||
| 0O7-3 | + | + | + | + | + | ||||||||
| 0V3-1 | + | + | + | + | + | + | |||||||
| 0V5-1 | + | + | + | ||||||||||
| 0V5-2 | + | + | |||||||||||
| 6S3-3 | + | + | + | + | + | ||||||||
| 6S3-4 | + | + | + | + | + | + | + | ||||||
| 6S4-1 | + | + | + | ||||||||||
| 6S4-2 | + | + | + | + | |||||||||
| 6S5-1 | + | + | + | + | |||||||||
| 6S5-3 | + | + | + | + | + | ||||||||
| 6B1-1 | + | + | + | ||||||||||
| 6B1-2 | + | + | + | ||||||||||
| 6B1-3 | + | + | + | ||||||||||
| 6B2-1 | + | + | + | + | |||||||||
| 6B3-1 | + | + | + | + | |||||||||
| 6B3-2 | + | + | + | + | |||||||||
| 6O5-2 | + | + | + | ||||||||||
| 6O5-3 | + | + | + | + | |||||||||
| 6O5-4 | + | + | + | + | |||||||||
| 6O6-1 | + | + | + | + | |||||||||
| 6O7-1 | + | + | + | + | |||||||||
| 6V3-1 | + | + | |||||||||||
| 6V5-1 | + | + | + | + | + | + | |||||||
| 6V6-1 | + | + | |||||||||||
| 12S6-2 | + | + | |||||||||||
| 12S6-3 | + | + | + | + | + | + | + | ||||||
| 12S7-2 | + | + | |||||||||||
| 12S7-3 | + | + | + | + | |||||||||
| 12S7-4 | + | + | + | ||||||||||
| 12B1-3 | + | + | + | + | + | ||||||||
| 12B1-4 | + | + | |||||||||||
| 12B2-4 | + | + | + | ||||||||||
| 12B2-5 | + | + | + | ||||||||||
| 12B3-2 | + | + | + | + | + | + | |||||||
| 12B3-4 | + | + | + | ||||||||||
| 12B3-5 | |||||||||||||
| 12O5-1 | + | + | + | ||||||||||
| 12O5-2 | + | + | + | ||||||||||
| 12O5-3 | + | + | |||||||||||
| 12O6-1 | + | + | + | + | + | + | |||||||
| 12O6-3 | + | + | + | + | |||||||||
| 12O7-1 | + | + | + | + | + | + | |||||||
| Cholic Acid 1% | Lecitine 3% | Lecitine 3% + Cholic Acid 1% | Filter Paper | |||||
|---|---|---|---|---|---|---|---|---|
| ROO | EVOO | ROO | EVOO | ROO | EVOO | ROO | EVOO | |
| 0S1-1 | PI | PI | PI | PI | ||||
| 0S1-2 | PI | PI | ||||||
| 0S2-1 | PI | |||||||
| 0B1-2 | PI | |||||||
| 0B2-4 | PI | PI | PI | PI | PI | |||
| 0B3-1 | PI | PI | ||||||
| ROO | EVOO | ROO | EVOO | ROO | EVOO | ROO | EVOO | |
| 0O5-2 | PI | |||||||
| 0O5-4 | PI | PI | PI | |||||
| 0O7-3 | PI | PI | ||||||
| 0V3-1 | PI | PI | ||||||
| 0V5-1 | PI | |||||||
| 0V5-2 | PI | PI | PI | PI | ||||
| 6S3-3 | PI | PI | ||||||
| 6S3-4 | PI | |||||||
| 6S4-1 | PI | PI | PI | PI | ||||
| 6S4-2 | PI | PI | ||||||
| 6S5-1 | PI | PI | ||||||
| 6S5-3 | PI | PI | PI | PI | PI | |||
| 6B1-1 | PI | |||||||
| 6B1-2 | PI | PI | ||||||
| 6B1-3 | PI | |||||||
| 6B2-1 | PI | PI | PI | |||||
| 6B3-1 | PI | PI | ||||||
| 6B3-2 | PI | PI | PI | PI | PI | |||
| 6O5-2 | PI | PI | PI | PI | PI | PI | ||
| 6O5-3 | PI | PI | PI | |||||
| 6O5-4 | PI | PI | ||||||
| 6O6-1 | PI | |||||||
| 6O7-1 | PI | |||||||
| 6V3-1 | PI | |||||||
| 6V5-1 | PI | PI | PI | |||||
| 6V6-1 | PI | PI | ||||||
| 12S6-2 | PI | PI | PI | |||||
| 12S6-3 | PI | PI | ||||||
| 12S7-2 | PI | |||||||
| 12S7-3 | PI | PI | PI | |||||
| 12S7-4 | PI | |||||||
| 12B1-3 | PI | |||||||
| 12B1-4 | PI | |||||||
| 12B2-4 | PI | |||||||
| 12B2-5 | PI | |||||||
| 12B3-2 | PI | |||||||
| 12B3-4 | PI | PI | ||||||
| 12B3-5 | PI | PI | ||||||
| 12O5-1 | PI | PI | PI | PI | PI | |||
| 12O5-2 | PI | |||||||
| 12O5-3 | PI | PI | ||||||
| 12O6-1 | PI | PI | ||||||
| 12O6-3 | PI | |||||||
| 12O7-1 | PI | PI | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, B.; Cobo, A.; Hidalgo, M.; Martínez-Rodríguez, A.M.; Prieto, I.; Gálvez, A.; Martínez-Cañamero, M. Influence of the Type of Diet on the Incidence of Pathogenic Factors and Antibiotic Resistance in Enterococci Isolated from Faeces in Mice. Int. J. Mol. Sci. 2019, 20, 4290. https://doi.org/10.3390/ijms20174290
Sánchez B, Cobo A, Hidalgo M, Martínez-Rodríguez AM, Prieto I, Gálvez A, Martínez-Cañamero M. Influence of the Type of Diet on the Incidence of Pathogenic Factors and Antibiotic Resistance in Enterococci Isolated from Faeces in Mice. International Journal of Molecular Sciences. 2019; 20(17):4290. https://doi.org/10.3390/ijms20174290
Chicago/Turabian StyleSánchez, Beatriz, Antonio Cobo, Marina Hidalgo, Ana M. Martínez-Rodríguez, Isabel Prieto, Antonio Gálvez, and Magdalena Martínez-Cañamero. 2019. "Influence of the Type of Diet on the Incidence of Pathogenic Factors and Antibiotic Resistance in Enterococci Isolated from Faeces in Mice" International Journal of Molecular Sciences 20, no. 17: 4290. https://doi.org/10.3390/ijms20174290