The Reason for Growth Inhibition of Ulmus pumila ‘Jinye’: Lower Resistance and Abnormal Development of Chloroplasts Slow Down the Accumulation of Energy

Abstract
:1. Introduction
2. Results and Analysis
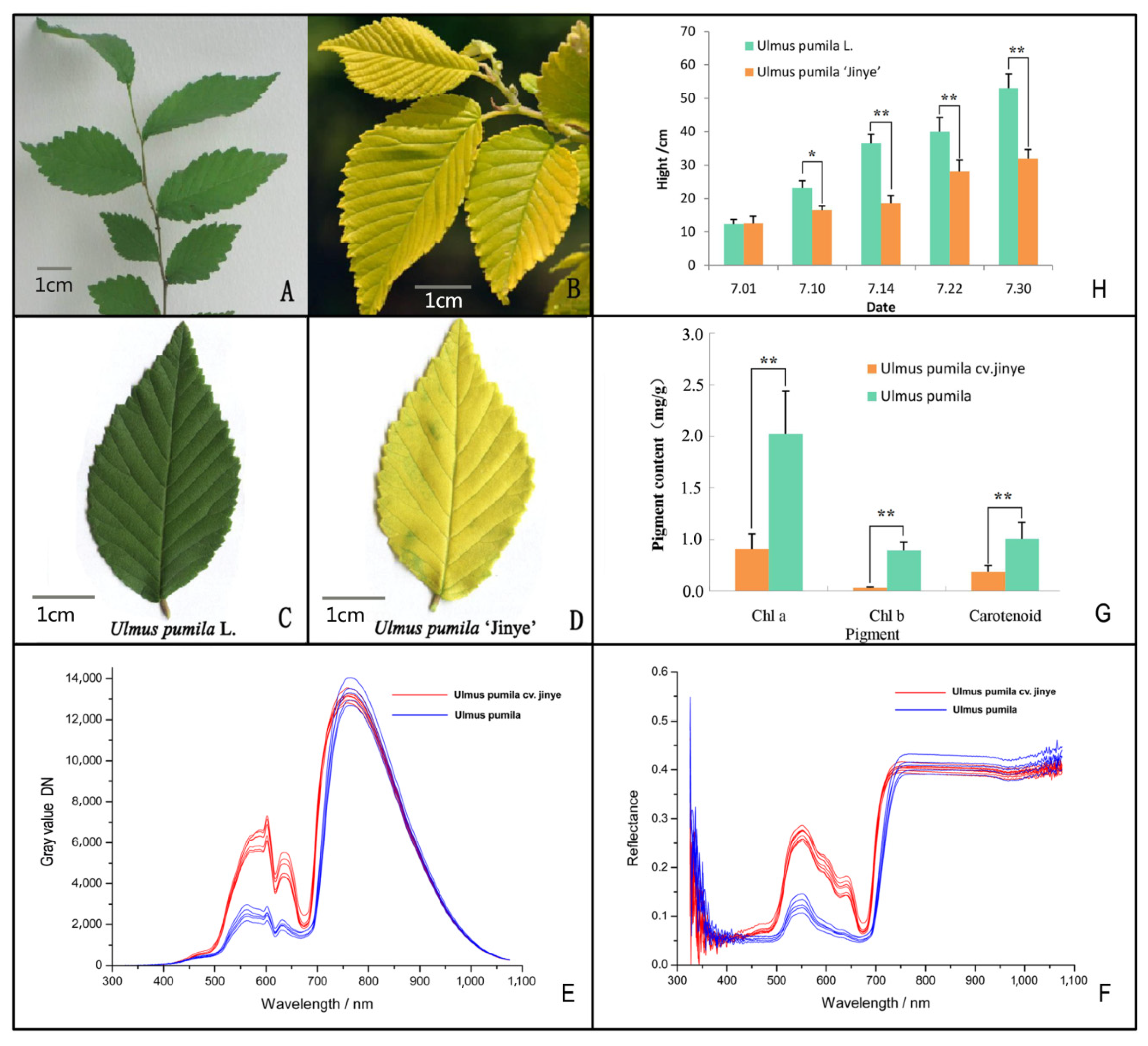
2.1. Growth and Physiological Factors Analysis
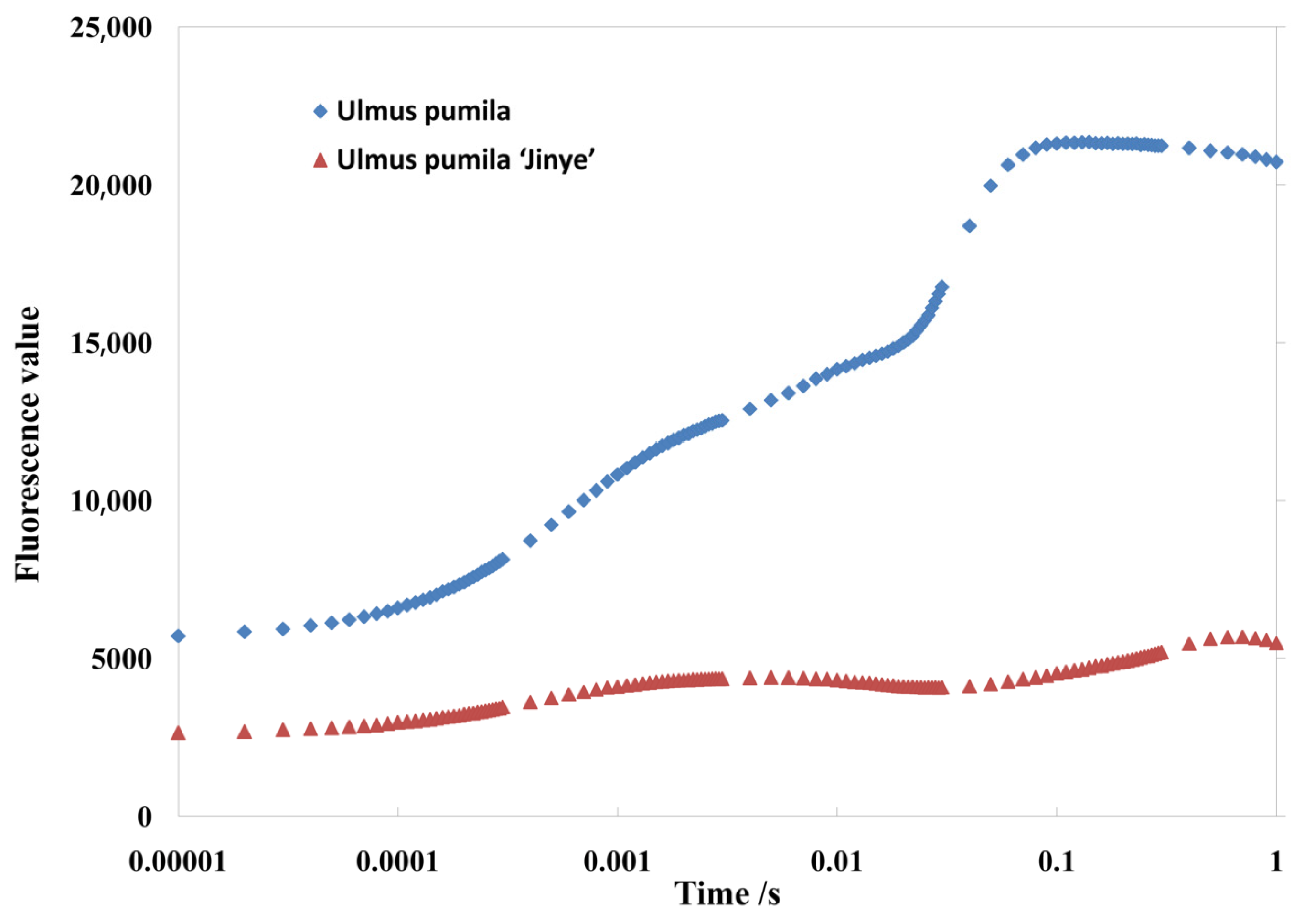
2.2. Photosynthetic Rate and Chlorophyll Fluorescence Analysis
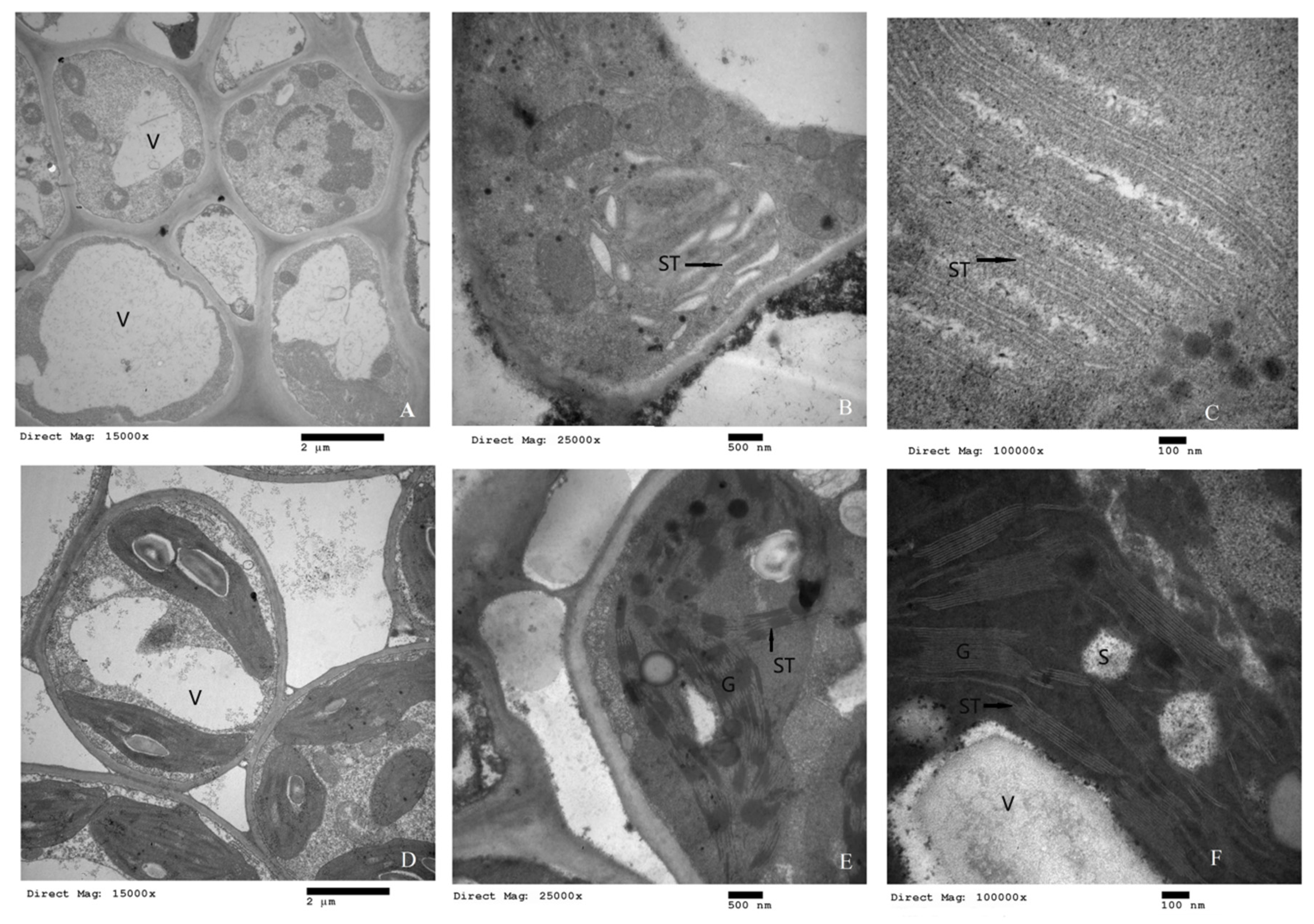
2.3. Cellular Microstructure Analysis
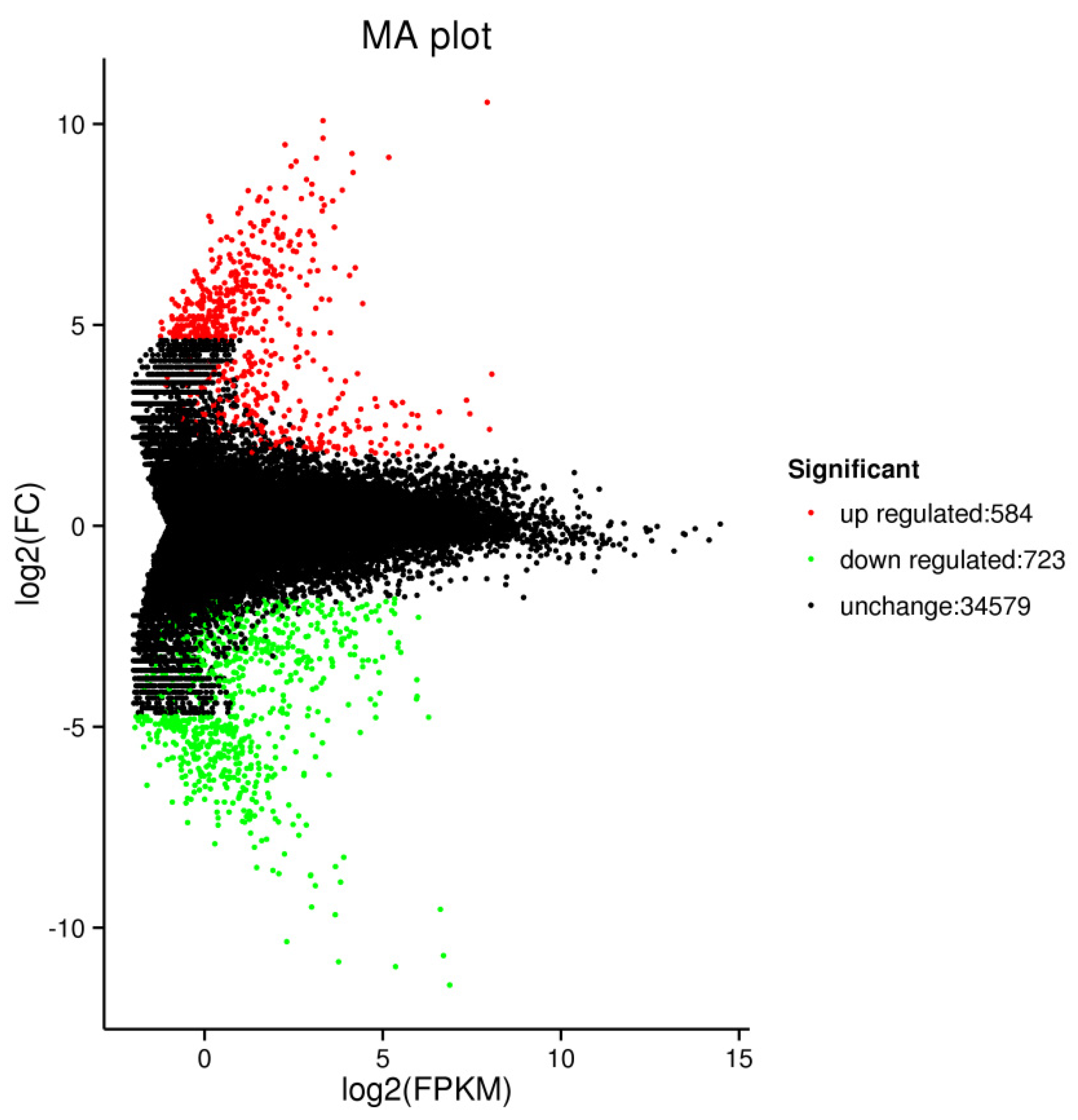
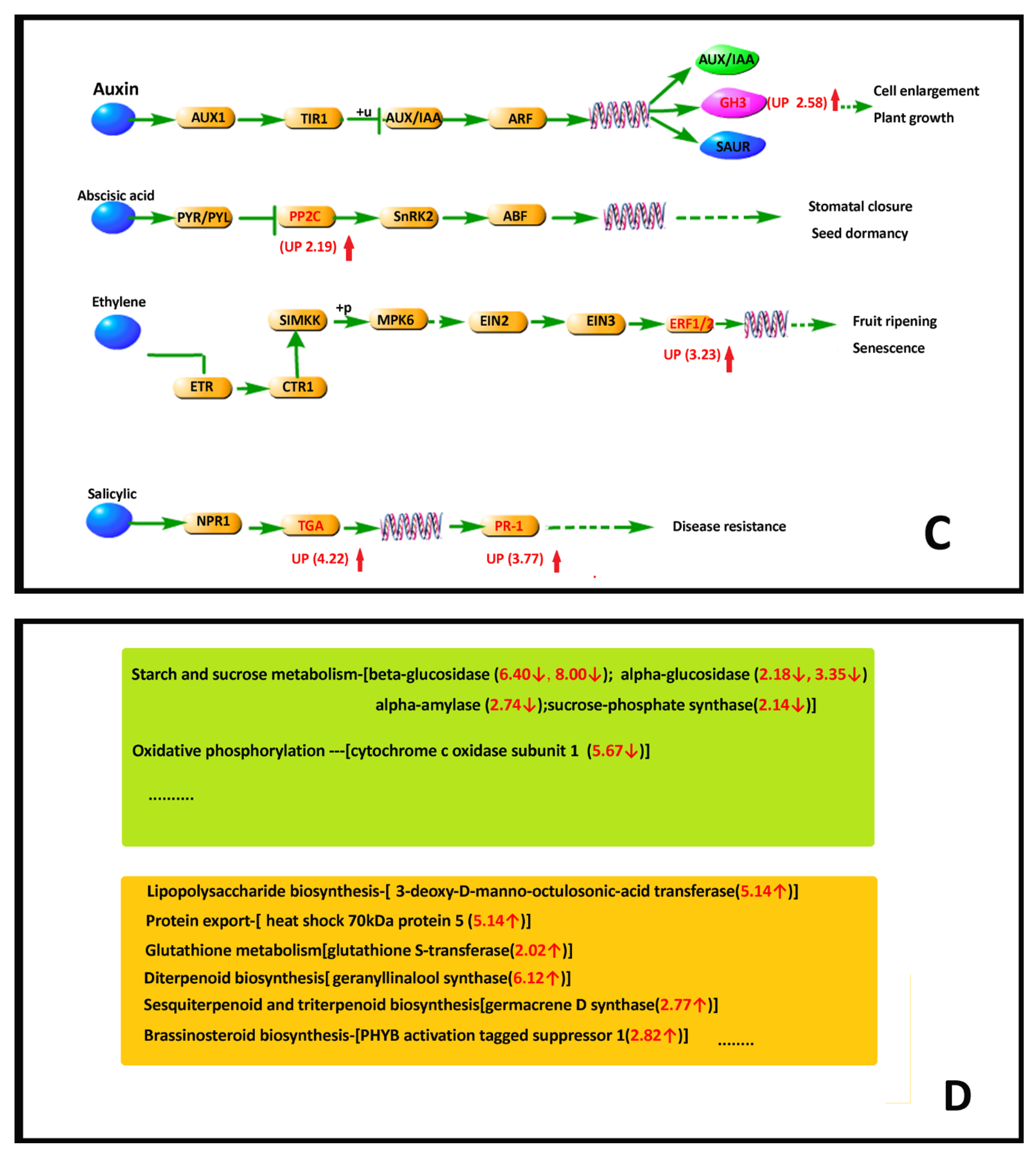
2.4. RNA Sequencing Analysis
2.4.1. DEG Analysis
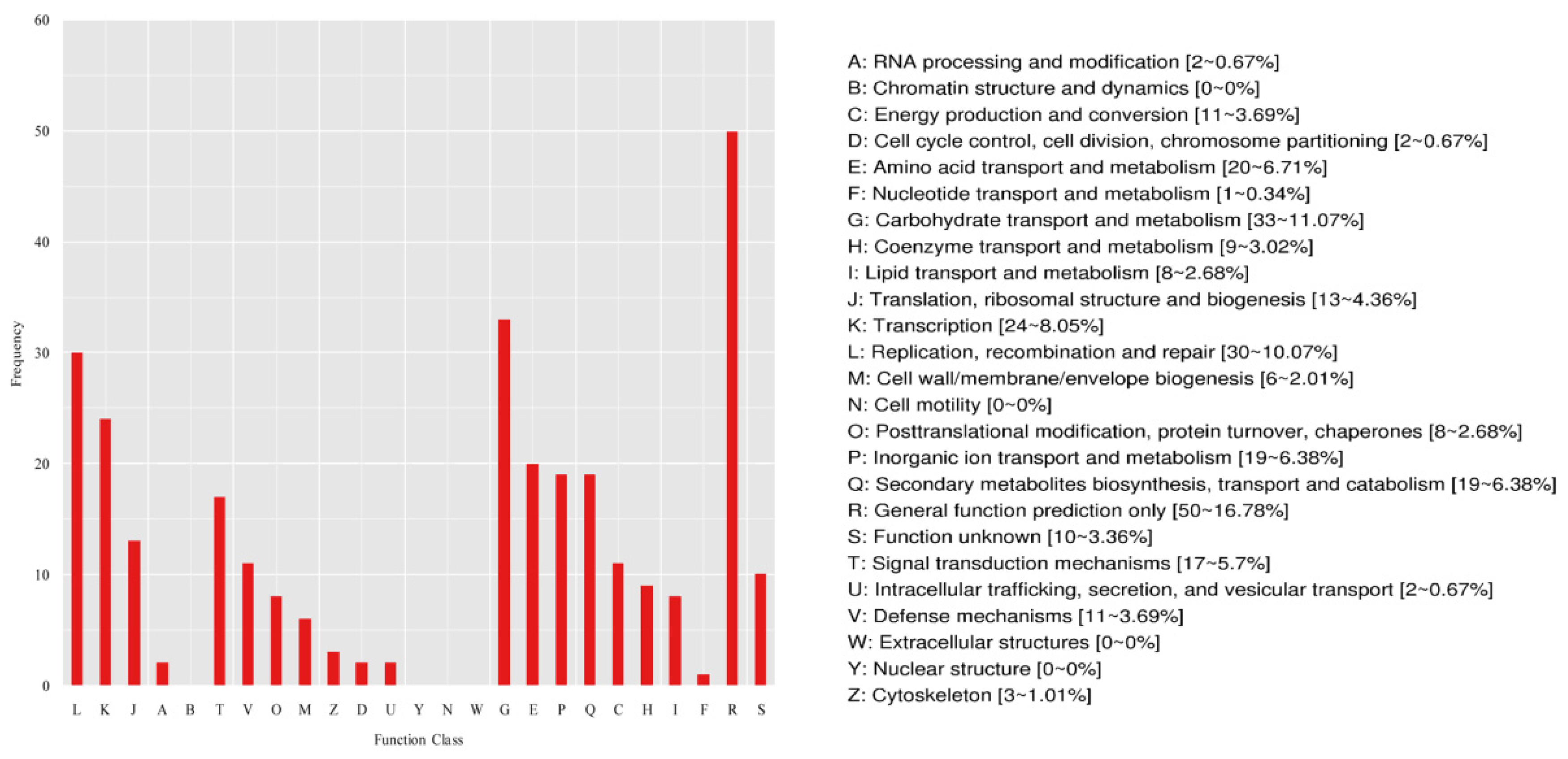
2.4.2. COG Classification of DEGs
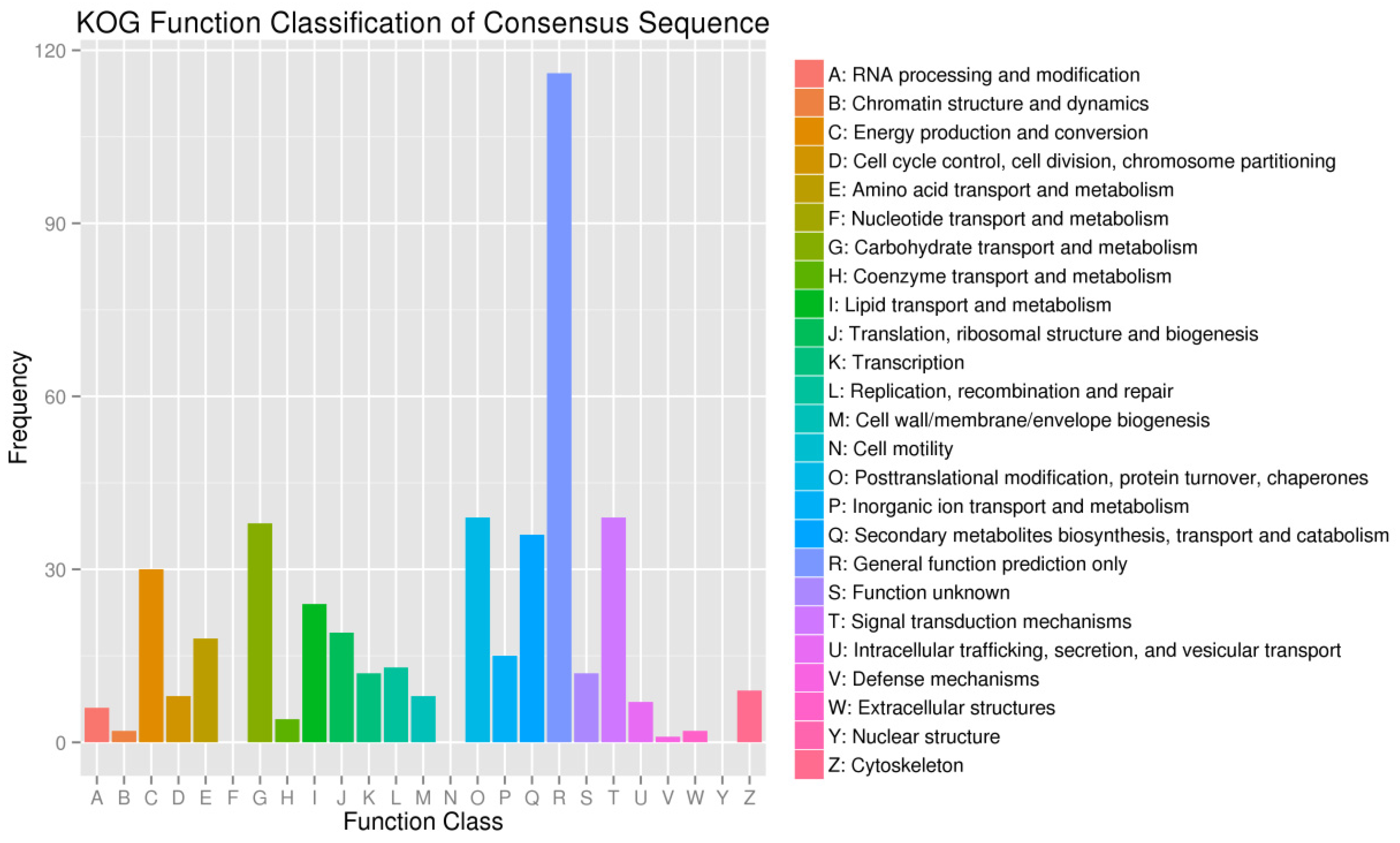
2.4.3. KOG Classification of DEGs
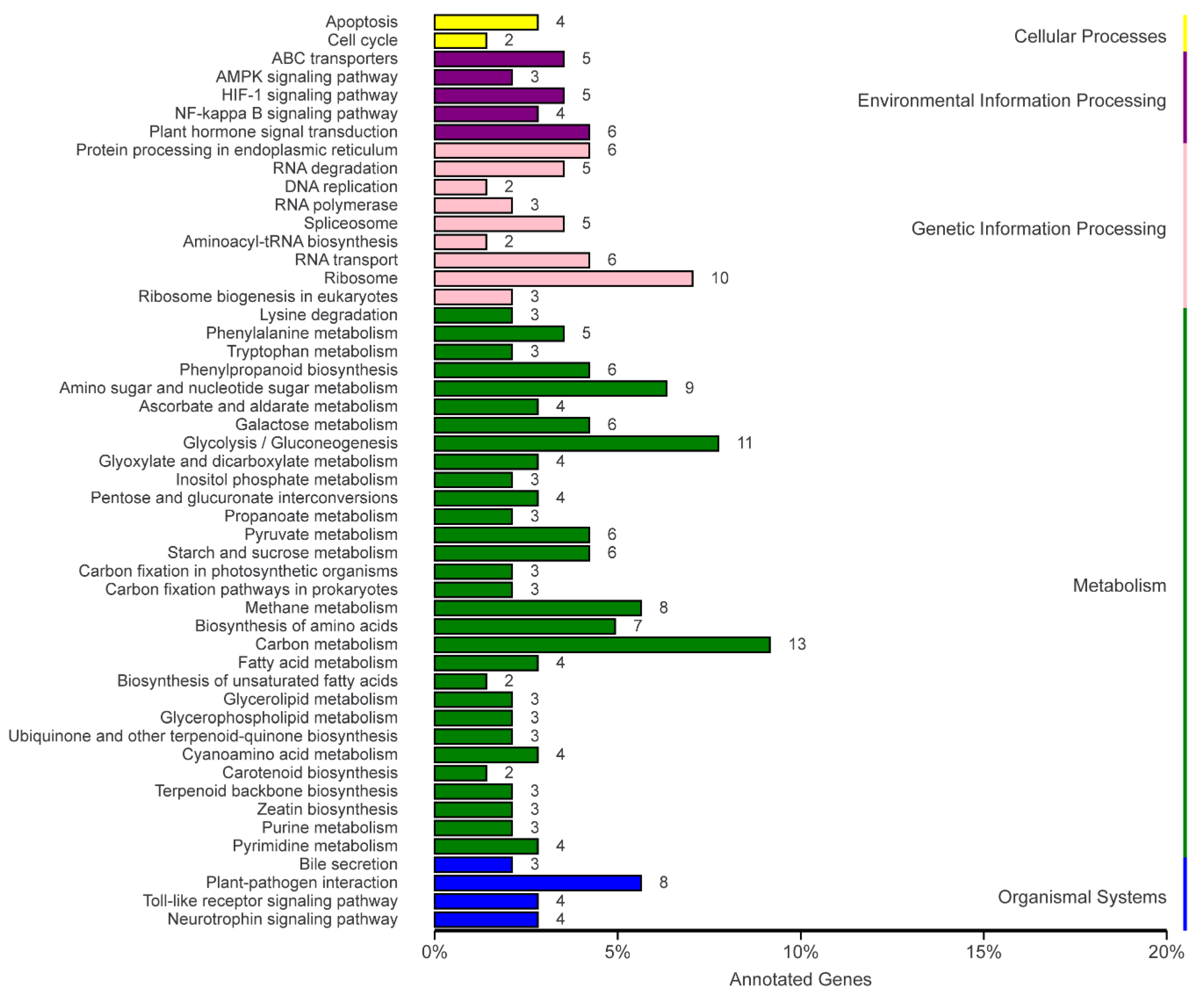
2.4.4. KEGG Metabolism Pathway Analysis
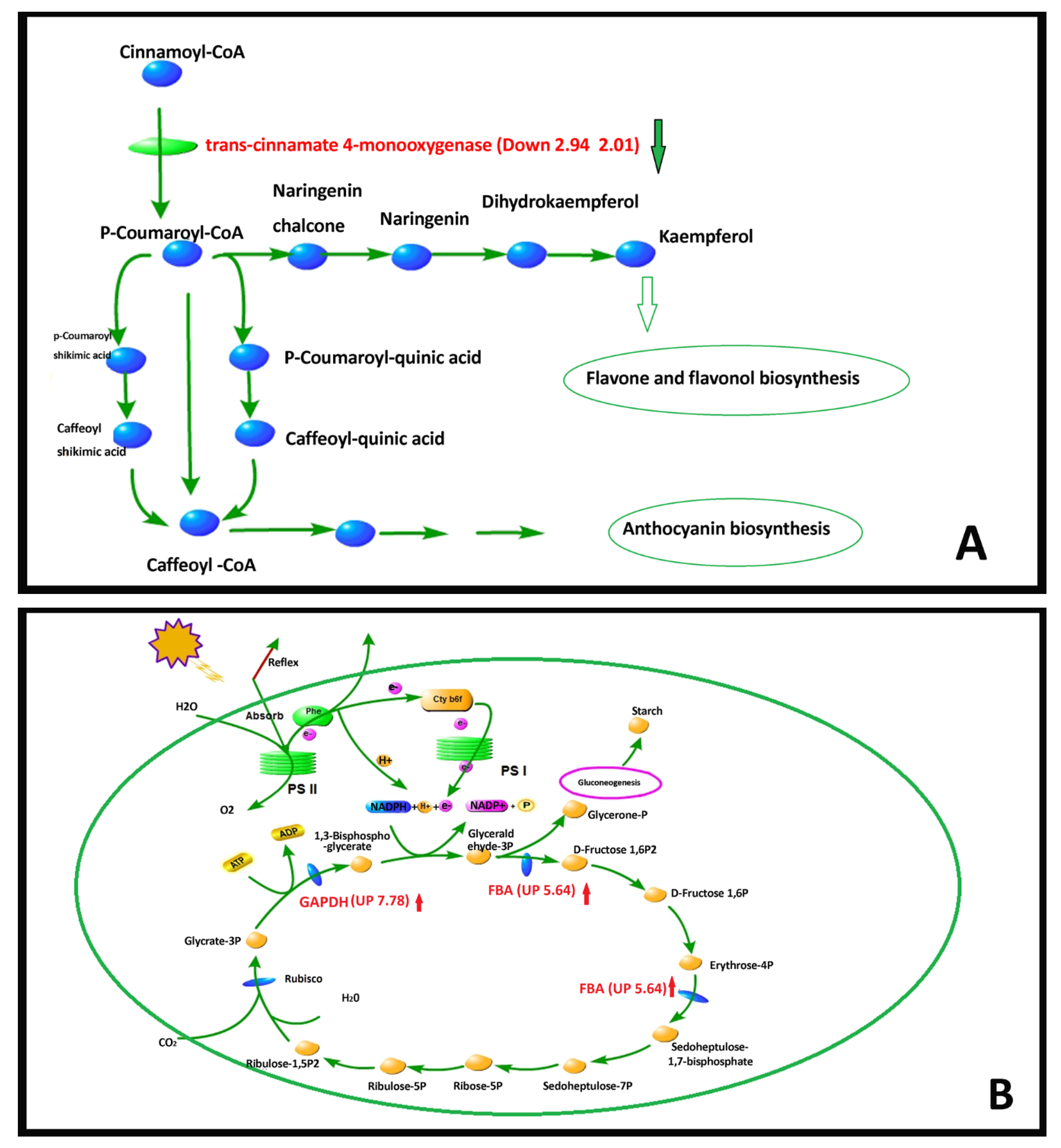
2.4.5. DEG Pathway Analysis
2.4.6. qPCR Verification Analysis
3. Materials and Methods
3.1. Materials
3.2. Determination of Physiological Factors
3.3. Photosynthetic Rate and Chlorophyll Fluorescence Analysis
3.4. Cellular Microstructure Analysis
3.5. RNA Extraction, Library Construction, and Sequencing
3.6. Illumina Sequencing, Assembly, and Functional Annotation
3.7. Real Time Q-PCR Verification
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Airy Show, H.K.; Willis’s, J.C. A Dictionary of the Flowering Plant and Ferns, 8th ed.; Cambridge University Press: Cambridge, UK, 1973; Volume 8. [Google Scholar]
- Dias, M.C.; Pinto, G.; Santos, C. Acclimatization of micropropagated plantlets induces an antioxidative burst: A case study with Ulmus minor Mill. Photosynthetica 2011, 49, 259–266. [Google Scholar] [CrossRef]
- Dias, M.C.; Oliveira, H.; Costa, A.; Santos, C. Improving elms performance under drought stress: The pretreatment with abscisic acid. Environ. Exp. Bot. 2014, 100, 64–73. [Google Scholar] [CrossRef]
- Zhang, S.; Zuo, L.H.; Zhang, J.; Chen, P.F.; Wang, J.M.; Yang, M.S. Transcriptome analysis of Ulmus pumila ‘Jinye’ responses to different shading involved in chlorophyll metabolism. Tree Genet. Genomes 2017, 13, 64. [Google Scholar] [CrossRef]
- Zuo, L.H.; Shang, A.Q.; Zhang, S.; Yu, X.Y.; Ren, Y.C.; Yang, M.S.; Wang, J.M. The first complete chloroplast genome sequences of Ulmus species by de novo sequencing: Genome comparative and taxonomic position analysis. PLoS ONE 2017, 12, e0171264. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.C.; Pinto, G.; Guerra, C.; Jesus, C.; Amaral, J.; Santos, C. Effect of irradiance during acclimatization on content of proline and phytohormones in micropropagated Ulmus minor. Biol. Plant 2013, 57, 769–772. [Google Scholar] [CrossRef]
- Pontier, D.; Albrieux, C.; Joyard, J.; Lagrange, T.; Block, M.A. Knock-out of the magnesium protoporphyrin IX methyltransferase gene in Arabidopsis. Effects on chloroplast development and on chloroplast-to-nucleus signaling. J. Biol. Chem. 2007, 282, 2297–2304. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Lee, J.Y.; Cho, H.S.; Lee, S.S.; Ha, H.J.; Kim, S.; Choi, D.; Pai, H.S. Inactivation of organellar glutamyl-and seryl-tRNA synthetases leads to developmental arrest of chloroplasts and mitochondria in higher plants. J. Biol. Chem. 2005, 280, 37098–37106. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.R.; Gao, J.X.; Wan, C.M.; Zhang, F.T.; Xu, Z.J.; Huang, X.Q.; Sun, X.Q.; Deng, X.J. Divinyl chlorophyll(ide) a can be converted to monovinyl chlorophyll(ide) a by a divinyl reductase in rice. Plant Physiol. 2010, 153, 994–1003. [Google Scholar] [CrossRef]
- Patrick, G.S.; Matthew, J.T. Light signalling pathways regulating the Mg-chelatase branchpoint of chlorophyll synthesis during de-etiolation in Arabidopsis thaliana. Photochem. Photobiol. Sci. 2008, 7, 1243–1252. [Google Scholar] [CrossRef]
- Terry, M.J.; Kendriek, R.E. Feedbaek inhibition of chlorophyll synthesis in the phytochrome chromophore deficient aurea and yellow-green-2 mutants of tomato. Plant Physiol. 1999, 119, 143–152. [Google Scholar] [CrossRef]
- Gray, J.C. Chloroplast-to-nucleus signalling: A role for Mg-protoporphyrin. Trends Genet. 2003, 19, 526–529. [Google Scholar] [CrossRef] [PubMed]
- Lonosky, P.M.; Zhang, X.S.; Honavar, V.G. A proteomic analysis of maize chloroplast biogenesis. Plant Physiol. 2004, 134, 560–574. [Google Scholar] [CrossRef] [PubMed]
- Highkin, H.; Boardman, N.K.; Goodchild, D.J. Photosynthetic studies on a pea mutant deficient in chlorophyll. Plant Physiol. 1969, 44, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Okabe, K.; Schmed, G.H.; Straub, J. Genetic characterization and high efficiency photosynthesis of an aurea mutant of tobacco. Plant Physiol. 1977, 60, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Stockinger, E.J.; Walling, L.L. A chlorophyll a/b-binding protein gene from soybean (Glycine max [L.] Merr.). Plant Physiol. 1994, 104, 1475–1476. [Google Scholar] [CrossRef] [PubMed]
- Preiss, S.; Thornber, J.P. Stability of the apoproteins of light-harvesting complex and during biogenesis of thylakoids in the chlorophyll b-less barley mutant chlorian f2. Plant Physiol. 1995, 107, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Carol, P.; Bisanz, C.; Breitenbach, J.; Sandmann, G.; Mache, R.; Coupland, G.; Kuntz, M. Mutations in the Arabidopsis gene IMMUTANTS cause a variegated phenotype by inactivating a chloroplast terminal oxidase associated with phytoene desaturation. Plant Cell 1999, 11, 57–68. [Google Scholar] [CrossRef]
- Jung, K.H.; Hur, J.; Ryu, C.H.; Choi, Y.J.; Chung, Y.Y.; Miyao, A.Y.; Hirochika, H.; An, G. Characterization of a chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol. 2003, 44, 463–472. [Google Scholar] [CrossRef]
- Zou, Q. Experimental Instruction of Plant Physiology; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Strasser, R.J. The Grouping Model of Plant Photosynthesis: Heterogeneity of Photosynthetic Units in Thylakoids. In Photosynthesis III. Structure and Molecular Organization of the Photosynthetic Apparatus; Akoyunoglou, G., Ed.; BISS Press: Philadelphia, PA, USA, 1981; pp. 727–737. [Google Scholar]
- Strasser, R.J.; Tsimill-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Advances in Photosynthesis and Respiration; Govindjee, P.G., Ed.; KAP Press: Dordrecht, The Netherlands, 2004; pp. 1–47. [Google Scholar]
- Eddy, S.R. Profile hidden Markov models. Bioinformatics 1998, 14, 755–763. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A. The COG database: A tool for genome scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Noeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.Z.; Lopez, R.; Magrane, M.; et al. UniProt: The Universal Protein knowledgebase. Nucleic Acids Res. 2004, 32, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Krylov, D.M.; Makarova, K.S.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; Rao, B.S.; et al. A comprehensive evolutionary classification of proteins encoded in complete eukaryotic genomes. Genome Biol. 2004, 5, R7. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.Y.; Li, J.Q.; Wu, S.F.; Zhu, Y.P.; Chen, Y.W.; He, F.C. Integrated nr database in protein annotation system and its localization. Comput. Eng. 2006, 32, 71–74. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L.P. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, 316–322. [Google Scholar] [CrossRef]
- Leng, N.; Dawson, J.A.; Thomson, J.A.; Ruotti, V.; Rissman, A.I.; Smits, B.M.G.; Haag, J.D.; Gould, M.N.; Ron, M.; Kendziorski, C. EBSeq: An empirical bayes hierarchical model for inference in RNA-seq experiments. Bioinformatics 2013, 29, 1035–1043. [Google Scholar] [CrossRef]
- Close, D.C.; Beadle, C.L. The ecophysiology of foliar anthocyanin. Bot. Rev. 2002, 69, 149–161. [Google Scholar] [CrossRef]
- Hughes, N.M.; Carpenter, K.L.; Cannon, J.G. Estimating contribution of anthocyanin pigments to osmotic adjustment during winter leaf reddening. J. Plant Physiol. 2014, 170, 230–233. [Google Scholar] [CrossRef]
- Lin, C.S.; Lai, Y.H.; Sun, C.W.; Liu, N.T.; Tsay, H.S.; Chang, W.C.; Teremy, J.; Chen, W. Identification of ESTs differentially expressed in green and albino mutantbamboo (Bambusaedulis) by suppressive subtractive hybridization (SSH) and microarray analysis. Plant Cell Tissue Organ. Cult. 2006, 86, 169–175. [Google Scholar] [CrossRef]
- Hörtensteiner, S. Stay-green regulates chlorophyll and chlorophyll-binding protein degradation during senescence. Trends Plant Sci. 2009, 14, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Melis, A. Excitation energy transfer: Functional and dynamic aspects of Lhc(cab) proteins. In Oxygenic Photosynthesis: The Light Reactions; Yocum, C.F., Heichel, I.F., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 523–538. [Google Scholar]
- Yang, Y.L.; Xu, J.; Rao, Y.C.; Zeng, Y.J.; Liu, H.J.; Zheng, T.T.; Zhang, G.H.; Hu, J.; Guo, L.B.; Qian, Q.; et al. Cloning and functional analysis of pale-green leaf(PGL10) in rice (Oryza sativa L.). Plant Growth Regul. 2016, 78, 69–77. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulae and Terms | U. pumila L. | U. pumila ’Jinye’ | |
|---|---|---|---|
| Net photosynthetic rate | Pn | 17.41 ± 1.42 a | 12.57 ± 2.80 b |
| Fluorescence intensity | Fo | 5695.32 ± 468.22 a | 2657.62 ± 248.74 b |
| Fm | 21,337.45 ± 715.39 a | 5686.73 ± 482.54 b | |
| Fv | 15,642.73 ± 657.22 a | 3029.86 ± 397.61 b | |
| Yields or flux ratios | φPo | 0.73 ± 0.13 a | 0.53 ± 0.08 b |
| ψO | 0.84 ± 0.11 a | 0.73 ± 0.11 b | |
| φEo | 0.62 ± 0.07 a | 0.39 ± 0.04 b | |
| φDo | 0.27 ± 0.04 b | 0.47 ± 0.05 a | |
| Phenomenological energy fluxes | ABS/CSO | 5695.73 ± 451.35 a | 2657.06 ± 426.33 b |
| TRO/CSO | 4174.96 ± 391.57 a | 1415.42 ± 206.54 b | |
| ETO/CSO | 3523.71 ± 299.36 a | 1039.25 ± 255.32 b | |
| DIO/CSO | 1520.04 ± 268.13 a | 1241.58 ± 143.68 b | |
| Density of reaction centers | RC/CSO | 2823.42 ± 162.47 a | 890.16 ± 118.67 b |
| Samples | Read Number | Base Number | GC Content | % ≥ Q30 | Mapped Reads | Mapped Ratio |
|---|---|---|---|---|---|---|
| U. pumila L. | 25,920,122 | 7,815,753,169 | 45.72% | 92.08% | 21,715,012 | 83.78% |
| U. pumila ‘Jinye’ | 26,788,118 | 8,080,109,712 | 45.70% | 92.40% | 22,450,089 | 83.81% |
| KEGG Pathway | DEG | Total | Enrichment Factor | p-Value | Corrected p-Value |
|---|---|---|---|---|---|
| Carbon metabolism | 13 | 643 | 1.22 | 0.27 | 1.00 |
| Glycolysis/Gluconeogenesis | 11 | 315 | 2.11 | 0.02 | 1.00 |
| Ribosome | 10 | 823 | 0.73 | 0.89 | 1.00 |
| Amino sugar and nucleotide sugar metabolism | 9 | 192 | 2.83 | 0.00 | 0.30 |
| Methane metabolism | 8 | 178 | 2.71 | 0.01 | 0.63 |
| Plant–pathogen interaction | 8 | 239 | 2.02 | 0.04 | 1.00 |
| Biosynthesis of amino acids | 7 | 480 | 0.88 | 0.69 | 1.00 |
| Galactose metabolism | 6 | 114 | 3.18 | 0.01 | 0.77 |
| Starch and sucrose metabolism | 6 | 265 | 1.37 | 0.28 | 1.00 |
| Pyruvate metabolism | 6 | 254 | 1.43 | 0.24 | 1.00 |
| Phenylpropanoid biosynthesis | 6 | 171 | 2.12 | 0.06 | 1.00 |
| RNA transport | 6 | 317 | 1.14 | 0.43 | 1.00 |
| Plant hormone signal transduction | 6 | 189 | 1.92 | 0.09 | 1.00 |
| Protein processing in endoplasmic reticulum | 6 | 402 | 0.90 | 0.66 | 1.00 |
| Phenylalanine metabolism | 5 | 116 | 2.60 | 0.04 | 1.00 |
| ABC transporters | 5 | 56 | 5.39 | 0.00 | 0.15 |
| RNA degradation | 5 | 208 | 1.45 | 0.26 | 1.00 |
| Spliceosome | 5 | 350 | 0.86 | 0.69 | 1.00 |
| HIF-1 signaling pathway | 5 | 150 | 2.01 | 0.10 | 1.00 |
| Pentose and glucuronate interconversions | 4 | 140 | 1.73 | 0.20 | 1.00 |
| Ascorbate and aldarate metabolism | 4 | 90 | 2.68 | 0.06 | 1.00 |
| Pyrimidine metabolism | 4 | 221 | 1.09 | 0.50 | 1.00 |
| Cyanoamino acid metabolism | 4 | 67 | 3.61 | 0.02 | 1.00 |
| Glyoxylate and dicarboxylate metabolism | 4 | 187 | 1.29 | 0.37 | 1.00 |
| Fatty acid metabolism | 4 | 178 | 1.36 | 0.34 | 1.00 |
| NF-kappa B signaling pathway | 4 | 79 | 3.06 | 0.04 | 1.00 |
| Apoptosis | 4 | 110 | 2.20 | 0.11 | 1.00 |
| Toll-like receptor signaling pathway | 4 | 111 | 2.18 | 0.11 | 1.00 |
| Neurotrophin signaling pathway | 4 | 165 | 1.46 | 0.29 | 1.00 |
| Ubiquinone and other terpenoid-quinone biosynthesis | 3 | 48 | 3.78 | 0.04 | 1.00 |
| Purine metabolism | 3 | 300 | 0.60 | 0.88 | 1.00 |
| Lysine degradation | 3 | 98 | 1.85 | 0.22 | 1.00 |
| Tryptophan metabolism | 3 | 99 | 1.83 | 0.23 | 1.00 |
| Glycerolipid metabolism | 3 | 116 | 1.56 | 0.30 | 1.00 |
| Inositol phosphate metabolism | 3 | 103 | 1.76 | 0.24 | 1.00 |
| Glycerophospholipid metabolism | 3 | 143 | 1.27 | 0.42 | 1.00 |
| Propanoate metabolism | 3 | 98 | 1.85 | 0.22 | 1.00 |
| Carbon fixation in photosynthetic organisms | 3 | 163 | 1.11 | 0.51 | 1.00 |
| Carbon fixation pathways in prokaryotes | 3 | 110 | 1.65 | 0.27 | 1.00 |
| Terpenoid backbone biosynthesis | 3 | 75 | 2.42 | 0.13 | 1.00 |
| Zeatin biosynthesis | 3 | 23 | 7.88 | 0.01 | 0.41 |
| Ribosome biogenesis in eukaryotes | 3 | 183 | 0.99 | 0.59 | 1.00 |
| RNA polymerase | 3 | 74 | 2.45 | 0.12 | 1.00 |
| AMPK signaling pathway | 3 | 213 | 0.85 | 0.69 | 1.00 |
| Bile secretion | 3 | 64 | 2.83 | 0.09 | 1.00 |
| Fructose and mannose metabolism | 2 | 109 | 1.11 | 0.54 | 1.00 |
| Fatty acid degradation | 2 | 129 | 0.94 | 0.63 | 1.00 |
| Photosynthesis | 2 | 56 | 2.16 | 0.24 | 1.00 |
| Valine, leucine, and isoleucine degradation | 2 | 128 | 0.94 | 0.63 | 1.00 |
| Glutathione metabolism | 2 | 133 | 0.91 | 0.65 | 1.00 |
| Riboflavin metabolism | 2 | 24 | 5.03 | 0.06 | 1.00 |
| Carotenoid biosynthesis | 2 | 29 | 4.17 | 0.08 | 1.00 |
| Nitrogen metabolism | 2 | 43 | 2.81 | 0.16 | 1.00 |
| Flavonoid biosynthesis | 2 | 38 | 3.18 | 0.13 | 1.00 |
| Stilbenoid, diarylheptanoid, and gingerol biosynthesis | 2 | 24 | 5.03 | 0.06 | 1.00 |
| Tropane, piperidine, and pyridine alkaloid biosynthesis | 2 | 46 | 2.63 | 0.18 | 1.00 |
| Aminoacyl-tRNA biosynthesis | 2 | 123 | 0.98 | 0.61 | 1.00 |
| Biosynthesis of unsaturated fatty acids | 2 | 84 | 1.44 | 0.41 | 1.00 |
| Degradation of aromatic compounds | 2 | 23 | 5.25 | 0.05 | 1.00 |
| DNA replication | 2 | 92 | 1.31 | 0.45 | 1.00 |
| Protein export | 2 | 78 | 1.55 | 0.37 | 1.00 |
| Cell cycle | 2 | 228 | 0.53 | 0.90 | 1.00 |
| Ubiquitin mediated proteolysis | 2 | 220 | 0.55 | 0.88 | 1.00 |
| Endocytosis | 2 | 211 | 0.57 | 0.87 | 1.00 |
| Insulin signaling pathway | 2 | 220 | 0.55 | 0.88 | 1.00 |
| Estrogen signaling pathway | 2 | 125 | 0.97 | 0.62 | 1.00 |
| Thyroid hormone synthesis | 2 | 56 | 2.16 | 0.24 | 1.00 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, L.; Zhang, S.; Liu, Y.; Huang, Y.; Yang, M.; Wang, J. The Reason for Growth Inhibition of Ulmus pumila ‘Jinye’: Lower Resistance and Abnormal Development of Chloroplasts Slow Down the Accumulation of Energy. Int. J. Mol. Sci. 2019, 20, 4227. https://doi.org/10.3390/ijms20174227
Zuo L, Zhang S, Liu Y, Huang Y, Yang M, Wang J. The Reason for Growth Inhibition of Ulmus pumila ‘Jinye’: Lower Resistance and Abnormal Development of Chloroplasts Slow Down the Accumulation of Energy. International Journal of Molecular Sciences. 2019; 20(17):4227. https://doi.org/10.3390/ijms20174227
Chicago/Turabian StyleZuo, Lihui, Shuang Zhang, Yichao Liu, Yinran Huang, Minsheng Yang, and Jinmao Wang. 2019. "The Reason for Growth Inhibition of Ulmus pumila ‘Jinye’: Lower Resistance and Abnormal Development of Chloroplasts Slow Down the Accumulation of Energy" International Journal of Molecular Sciences 20, no. 17: 4227. https://doi.org/10.3390/ijms20174227