Nuclear Disposition of Alien Chromosome Introgressions into Wheat and Rye Using 3D-FISH

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Morphometric Characteristics of G1 Nuclei of Wheat-Rye and Wheat-Barley Chromosome Introgression Lines
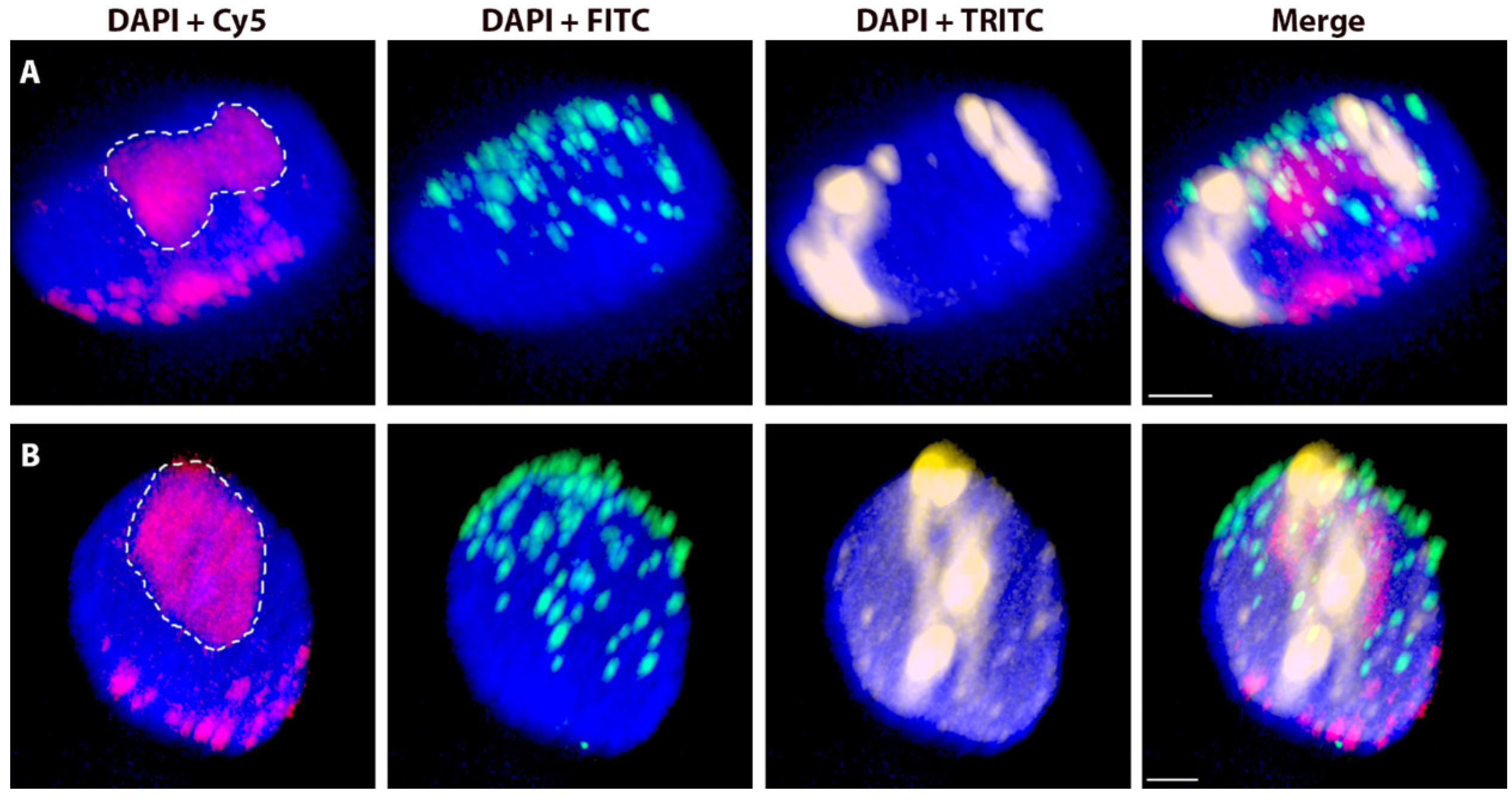
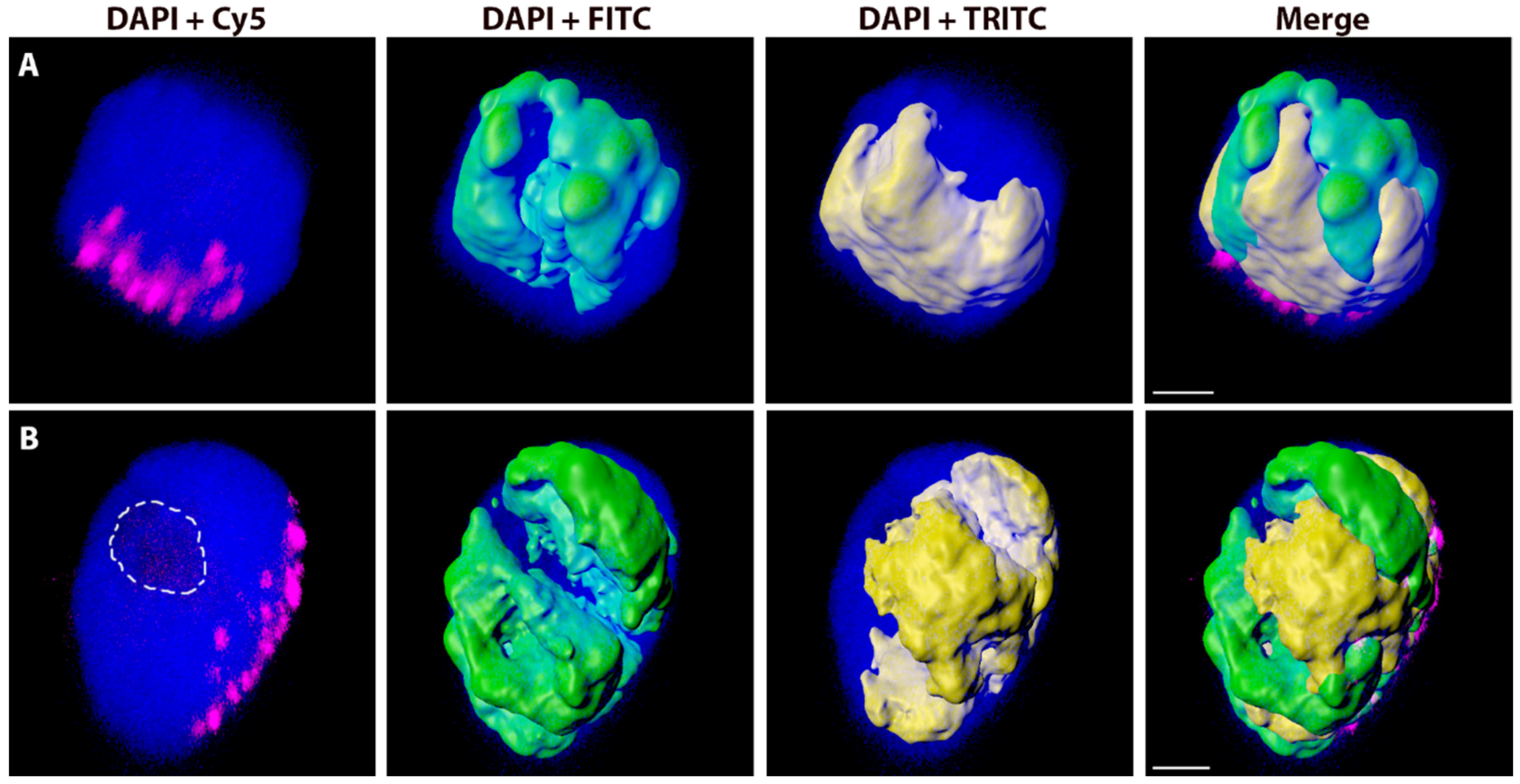
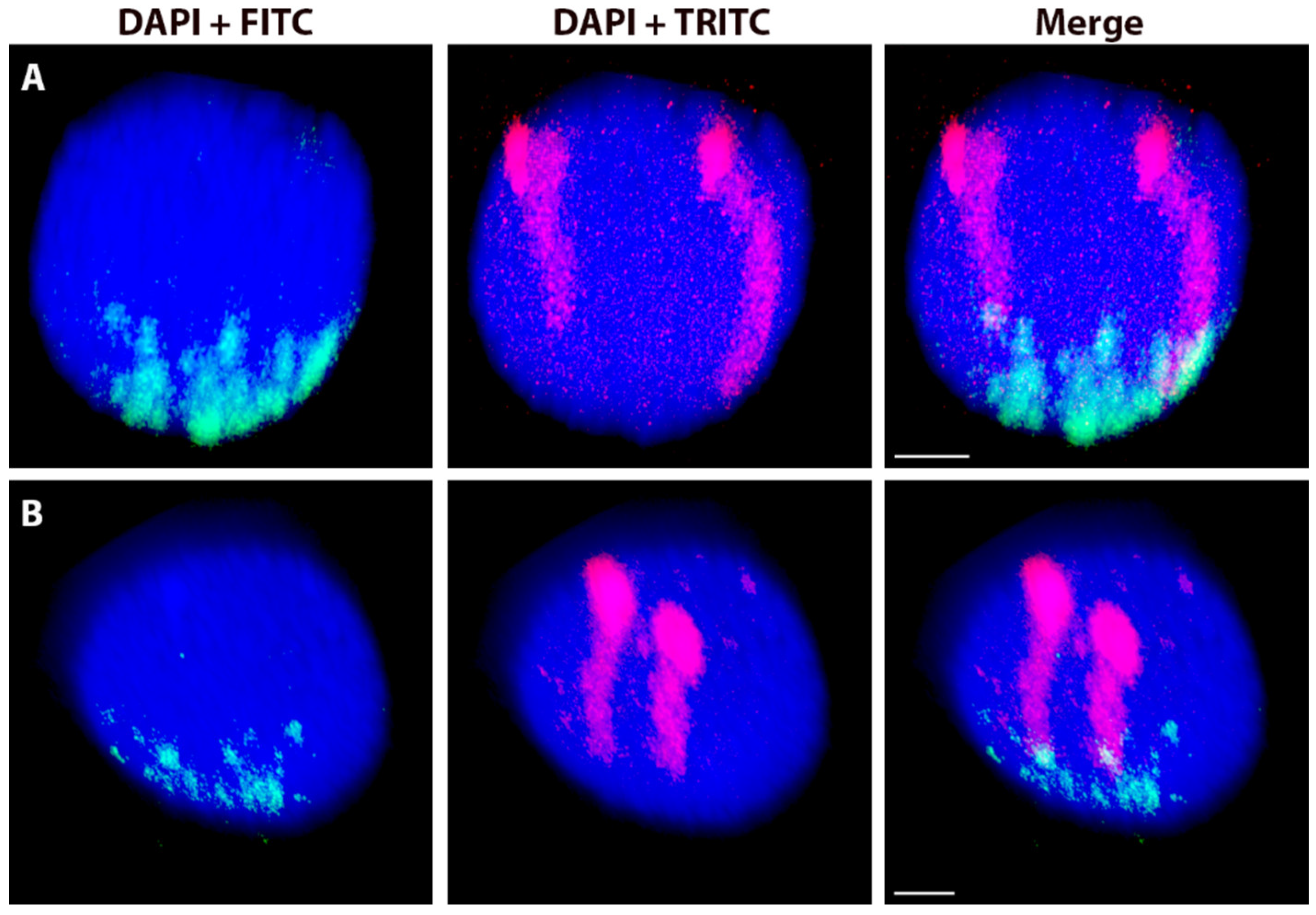
2.2. Variation in the Spatial Arrangements in G1 Nuclei of Alien Chromosome Arms in Various Wheat-Rye and Wheat-Barley Introgression Lines
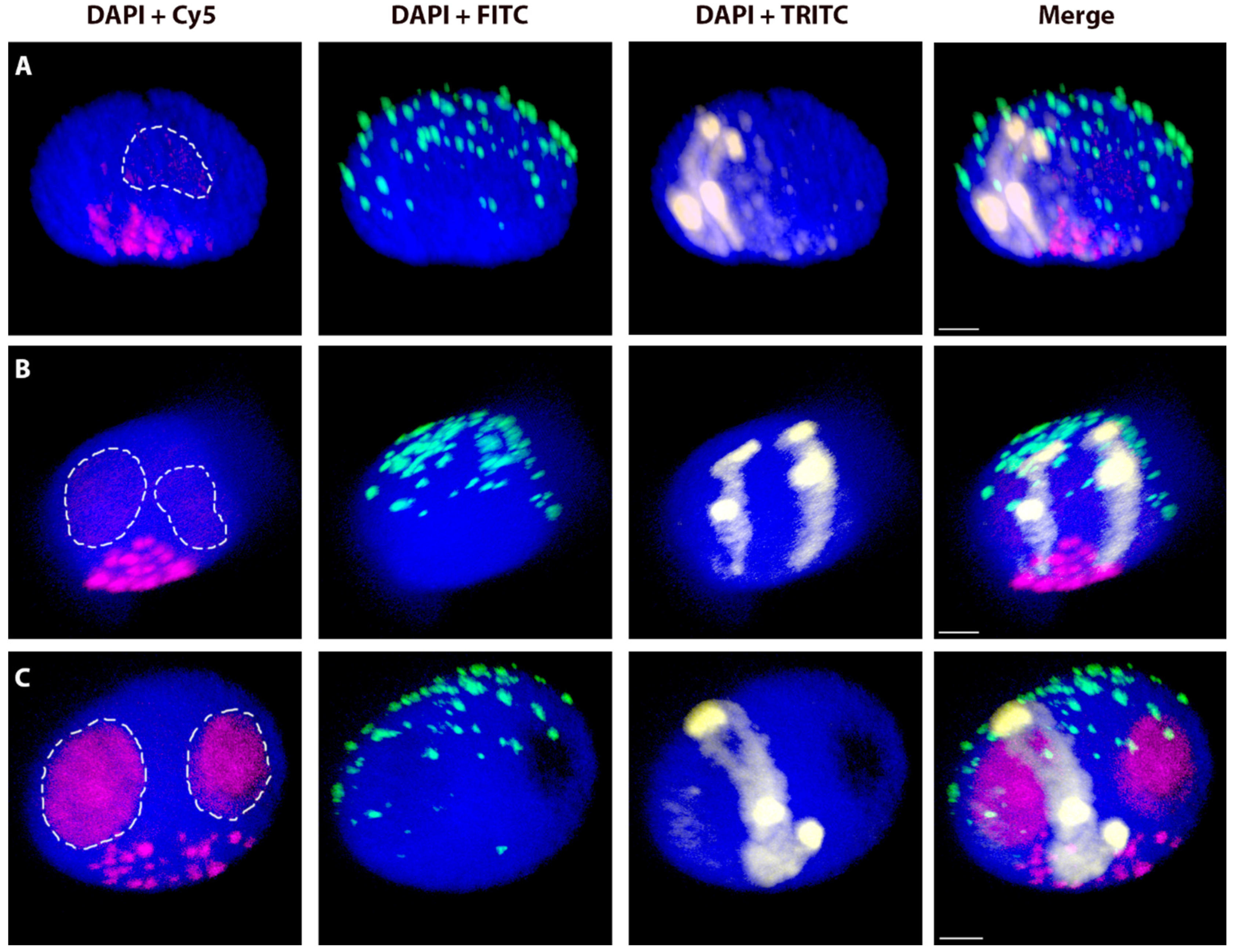
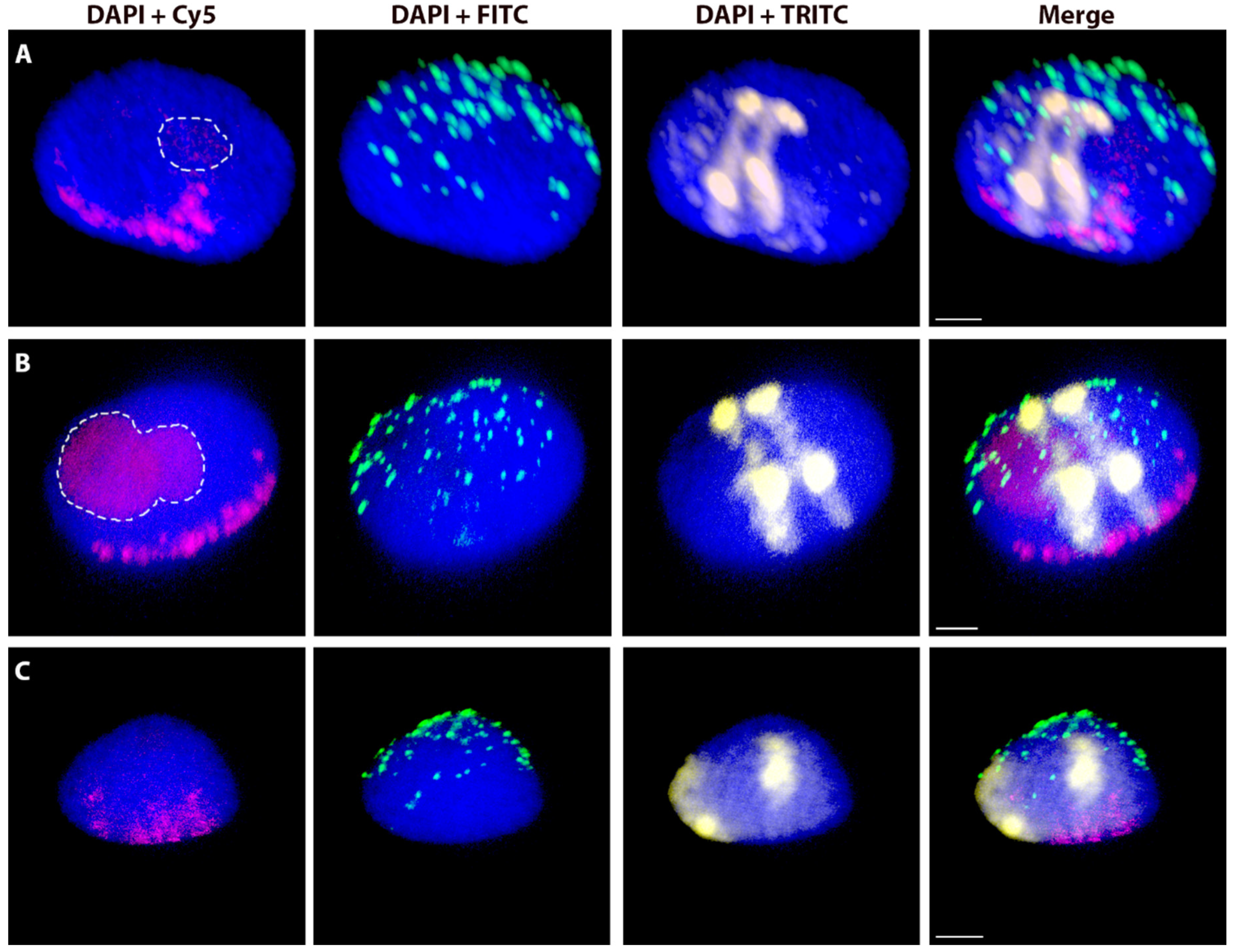
2.3. The Effect of the Cell Cycle Stages and Tissue-Specificity
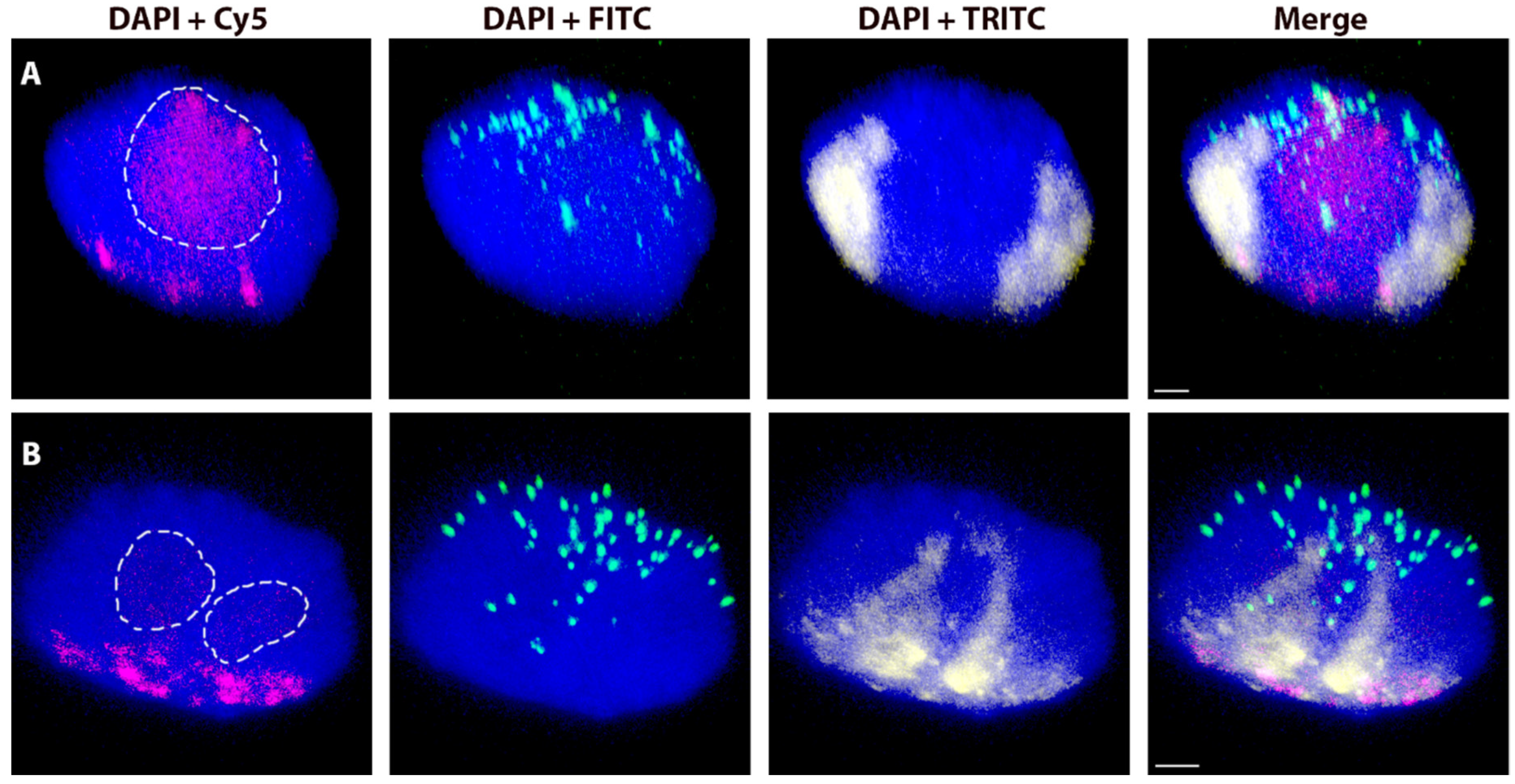
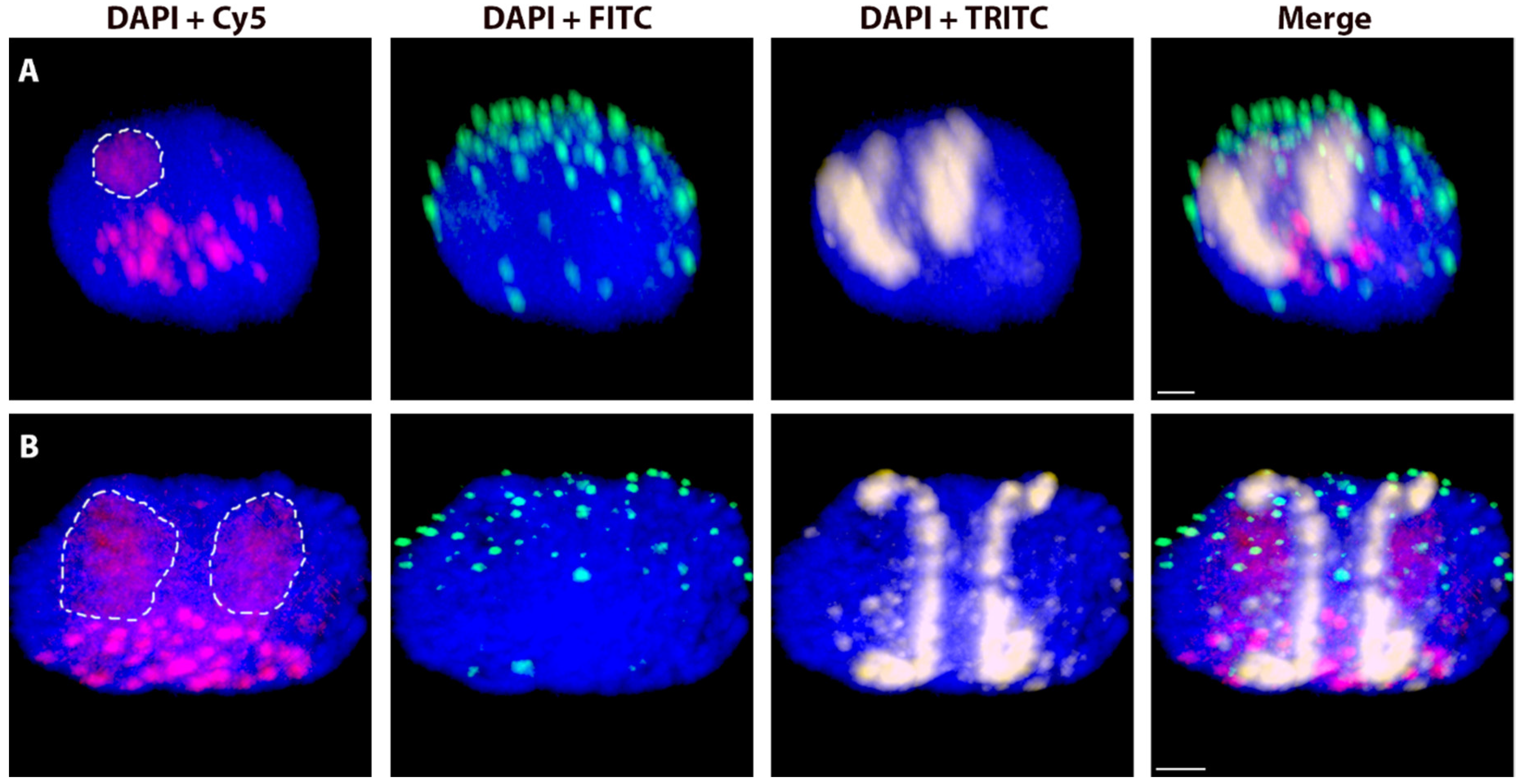
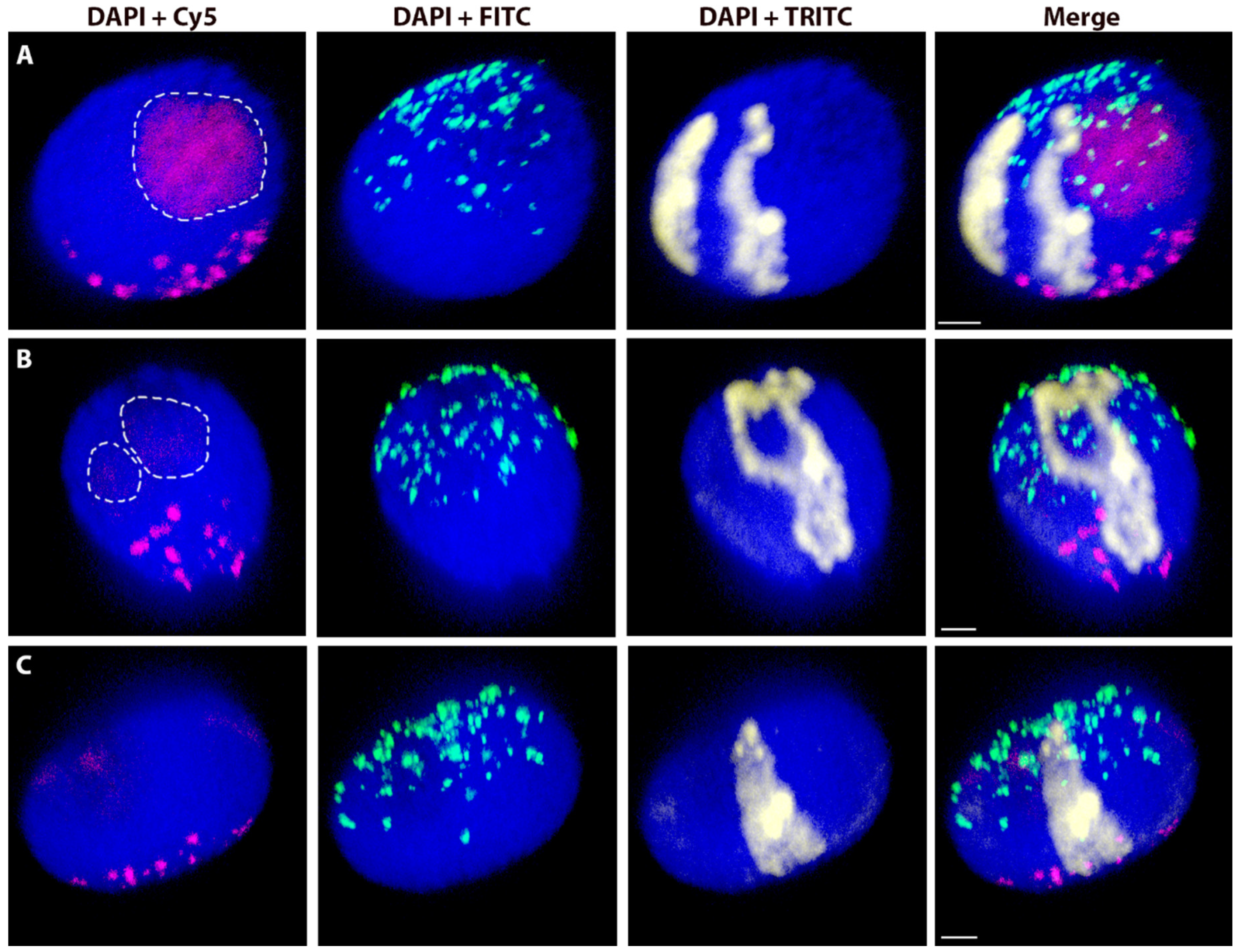
2.4. Spatial Separation vs. Association of Alien Chromosome (Chromosome Arm) Homologues
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Isolation of Nuclei by Flow Cytometry Sorting
4.3. Probe Preparation and 3D-FISH
4.4. Image Acquisition and Analysis
4.5. Association vs. Separation of Rye Homologous Chromosome Arms in the Wheat Nucleus
4.6. Statistical Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D-FISH | Three-dimensional fluorescent in situ hybridization |
| CN | Centre of the nucleus |
| CT | Chromosome territory |
| GISH | Genomic in situ hybridization |
| MA | Arm mid-point |
| NOR | Nucleolar organizing region |
Appendix A




References
- Cremer, T.; Cremer, C. Chromosome territories, nuclear architecture and gene regulation in mammalian cells. Nat. Rev. Genet. 2001, 2, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Fritz, A.J.; Barutcu, A.R.; Martin-Buley, L.; van Wijnen, A.J.; Zaidi, S.K.; Imbalzano, A.N.; Lian, J.B.; Stein, J.L.; Stein, G.S. Chromosomes at work: Organization of chromosome territories in the interphase nucleus. J. Cell. Biochem. 2016, 117, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.L.; Cuaycong, M.H.; Craig, C.A.; Wallrath, L.L.; Locke, J.; Eigin, S.C.R. The fourth chromosome of Drosophila melanogaster: Interspersed euchromatic and heterochromatic domains. Proc. Natl. Acad. Sci. USA 2000, 97, 5340–5345. [Google Scholar] [CrossRef] [PubMed]
- Kozubek, S.; Lukasova, E.; Jirsova, P.; Koutna, I.; Kozubek, M.; Ganova, A.; Bartova, E.; Falk, M.; Pasekova, R. 3D Structure of the human genome: order in randomness. Chromosoma 2002, 111, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Mayer, R.; Brero, A.; von Hase, J.; Schroeder, T.; Cremer, T.; Dietzel, S. Common themes and cell type specific variations of higher order chromatin arrangements in the mouse. Bmc Cell Biol. 2005, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Gorkin, D.U.; Leung, D.; Ren, B. The 3D genome in transcriptional regulation and pluripotency. Cell Stem Cell 2014, 14, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Croft, J.A.; Bridger, J.M.; Boyle, S.; Perry, P.; Teague, P.; Bickmore, W.A. Differences in the localization and morphology of chromosomes in the human nucleus. J. Cell Biol. 1999, 145, 1119–1131. [Google Scholar] [CrossRef]
- Cremer, M.; von Hase, J.; Volm, T.; Brero, A.; Kreth, G.; Walter, J.; Fischer, C.; Solovei, I.; Cremer, C.; Cremer, T. Non-random radial higher-order chromatin arrangements in nuclei of diploid human cells. Chromosome Res. 2001, 9, 541–567. [Google Scholar] [CrossRef]
- Tanabe, H.; Muller, S.; Neusser, M.; von Hase, J.; Calcagno, E.; Cremer, M.; Solovei, I.; Cremer, C.; Cremer, T. Evolutionary conservation of chromosome territory arrangements in cell nuclei from higher primates. Proc. Natl. Acad. Sci. USA 2002, 99, 4424–4429. [Google Scholar] [CrossRef] [Green Version]
- Habermann, F.A.; Cremer, M.; Walter, J.; Kreth, G.; von Hase, J.; Bauer, K.; Wienberg, J.; Cremer, C.; Cremer, T.; Solovei, I. Arrangements of macro- and microchromosomes in chicken cells. Chromosome Res. 2001, 9, 569–584. [Google Scholar] [CrossRef]
- Manvelyan, M.; Hunstig, F.; Bhatt, S.; Mrasek, K.; Pellestor, F.; Weise, A.; Simonyan, I.; Aroutiounian, R.; Liehr, T. Chromosome distribution in human sperm—A 3D multicolor banding-study. Mol. Cytogenet. 2008, 1, 25. [Google Scholar] [CrossRef] [PubMed]
- Manvelyan, M.; Hunstig, F.; Mrasek, K.; Bhatt, S.; Pellestor, F.; Weise, A.; Liehr, T. Position of chromosomes 18, 19, 21 and 22 in 3D-preserved interphase nuclei of human and gorilla and white hand gibbon. Mol. Cytogenet. 2008, 1, 9. [Google Scholar] [CrossRef] [PubMed]
- Rabl, C. Über Zellteilung. Morph. Jahrb. 1885, 10, 214–330. [Google Scholar]
- Dong, F.G.; Jiang, J.M. Non-Rabl patterns of centromere and telomere distribution in the interphase nuclei of plant cells. Chromosome Res. 1998, 6, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Schubert, I.; Shaw, P. Organization and dynamics of plant interphase chromosomes. Trends Plant Sci. 2011, 16, 273–281. [Google Scholar] [CrossRef]
- Pecinka, A.; Schubert, V.; Meister, A.; Kreth, G.; Klatte, M.; Lysak, M.A.; Fuchs, J.; Schubert, I. Chromosome territory arrangement and homologous pairing in nuclei of Arabidopsis thaliana are predominantly random except for NOR-bearing chromosomes. Chromosoma 2004, 113, 258–269. [Google Scholar] [CrossRef]
- Fransz, P.; de Jong, J.H.; Lysak, M.; Castiglione, M.R.; Schubert, I. Interphase chromosomes in Arabidopsis are organized as well defined chromocenters from which euchromatin loops emanate. Proc. Natl. Acad. Sci. USA 2002, 99, 14584–14589. [Google Scholar] [CrossRef]
- Schubert, V.; Rudnik, R.; Schubert, I. Chromatin associations in arabidopsis interphase nuclei. Front. Genet. 2014, 5, 389. [Google Scholar] [CrossRef]
- Berr, A.; Schubert, I. Interphase chromosome arrangement in Arabidopsis thaliana is similar in differentiated and meristematic tissues and shows a transient mirror symmetry after nuclear division. Genetics 2007, 176, 853–863. [Google Scholar] [CrossRef]
- Dekker, J.; Rippe, K.; Dekker, M.; Kleckner, N. Capturing chromosome conformation. Science 2002, 295, 1306–1311. [Google Scholar] [CrossRef]
- Lieberman-Aiden, E.; van Berkum, N.L.; Williams, L.; Imakaev, M.; Ragoczy, T.; Telling, A.; Amit, I.; Lajoie, B.R.; Sabo, P.J.; Dorschner, M.O.; et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 2009, 326, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Dixon, J.R.; Selvaraj, S.; Yue, F.; Kim, A.; Li, Y.; Shen, Y.; Hu, M.; Liu, J.S.; Ren, B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 2012, 485, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abney, J.R.; Cutler, B.; Fillbach, M.L.; Axelrod, D.; Scalettar, B.A. Chromatin dynamics in interphase nuclei and its implications for nuclear structure. J. Cell Biol. 1997, 137, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Dong, P.; Tu, X.; Li, H.; Zhang, J.; Grierson, D.; Li, P.; Zhong, S. Tissue-specific Hi-C analyses of rice, foxtail millet and maize suggest non-canonical function of plant chromatin domains. J. Intagr. Plant Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Flagel, L.E.; Paterson, A.H.; Rapp, R.A.; Soltis, D.E.; Soltis, P.S.; Wendel, J.F. Evolutionary genetics of genome merger and doubling in plants. Annu. Rev. Genet. 2008, 42, 443–461. [Google Scholar] [CrossRef]
- Schardin, M.; Cremer, T.; Hager, H.D.; Lang, M. Specific staining of human-chromosomes in chinese-hamster x man hybrid cell-lines demonstrates interphase chromosome territories. Hum. Genet. 1985, 71, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, K.; Camps, J.; Mathews, P.; Barenboim-Stapleton, L.; Nguyen, Q.T.; Difilippantonio, M.J.; Ried, T. Position of human chromosomes is conserved in mouse nuclei indicating a species-independent mechanism for maintaining genome organization. Chromosoma 2008, 117, 499–509. [Google Scholar] [CrossRef] [Green Version]
- Heslopharrison, J.S.; Leitch, A.R.; Schwarzacher, T.; Anamthawatjonsson, K. Detection and characterization of 1b/1r translocations in hexaploid wheat. Heredity 1990, 65, 385–392. [Google Scholar] [CrossRef]
- Kopecky, D.; Allen, D.C.; Duchoslav, M.; Dolezel, J.; Lukaszewski, A.J. Condensation of rye chromatin in somatic interphase nuclei of Ph1 and ph1b wheat. Cytogenet. Genome Res. 2007, 119, 263–267. [Google Scholar] [CrossRef]
- Schlegel, R.; Melz, G.; Nestrowicz, R. A Universal reference karyotype in rye, Secale cereale L. Theor. Appl. Genet. 1987, 74, 820–826. [Google Scholar] [CrossRef]
- Naranjo, T. Variable patterning of chromatin remodeling, telomere positioning, synapsis, and chiasma formation of individual rye chromosomes in meiosis of wheat-rye additions. Front. Plant Sci. 2018, 9, 880. [Google Scholar] [CrossRef] [PubMed]
- Gill, B.S.; Friebe, B.; Endo, T.R. Standard karyotype and nomenclature system for description of chromosome bands and structural-aberrations in wheat (Triticum aestivum). Genome 1991, 34, 830–839. [Google Scholar] [CrossRef]
- Paux, E.; Sourdille, P.; Salse, J.; Saintenac, C.; Choulet, F.; Leroy, P.; Korol, A.; Michalak, M.; Kianian, S.; Spielmeyer, W.; et al. A physical map of the 1-gigabase bread wheat chromosome 3B. Science 2008, 322, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Salina, E.A.; Nesterov, M.A.; Frenkel, Z.; Kiseleva, A.A.; Timonova, E.M.; Magni, F.; Vrana, J.; Safar, J.; Simkova, H.; Dolezel, J.; et al. Features of the organization of bread wheat chromosome 5BS based on physical mapping. Bmc Genom. 2018, 19, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. A chromosome conformation capture ordered sequence of the barley genome. Nature 2017, 544, 426. [Google Scholar] [CrossRef] [PubMed]
- Dolezel, J.; Greilhuber, J.; Lucretti, S.; Meister, A.; Lysak, M.A.; Nardi, L.; Obermayer, R. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann. Bot. 1998, 82, 17–26. [Google Scholar] [CrossRef]
- Sequeira-Mendes, J.; Gutierrez, C. Genome architecture: from linear organisation of chromatin to the 3D assembly in the nucleus. Chromosoma 2016, 125, 455–469. [Google Scholar] [CrossRef]
- Abranches, R.; Beven, A.F.; Aragon-Alcaide, L.; Shaw, P.J. Transcription sites are not correlated with chromosome territories in wheat nuclei. J. Cell Biol. 1998, 143, 5–12. [Google Scholar] [CrossRef]
- Mikhailova, E.I.; Sosnikhina, S.P.; Kirillova, G.A.; Tikholiz, O.A.; Smirnov, V.G.; Jones, R.N.; Jenkins, G. Nuclear dispositions of subtelomeric and pericentromeric chromosomal domains during meiosis in asynaptic mutants of rye (Secale cereale L.). J. Cell Biol. 2001, 114, 1875–1882. [Google Scholar]
- Corredor, E.; Diez, M.; Shepherd, K.; Naranjo, T. The positioning of rye homologous chromosomes added to wheat through the cell cycle in somatic cells untreated and treated with colchicine. Cytogenet. Genome Res. 2005, 109, 112–119. [Google Scholar] [CrossRef]
- Idziak, D.; Robaszkiewicz, E.; Hasterok, R. Spatial distribution of centromeres and telomeres at interphase varies among brachypodium species. J. Exp. Bot. 2015, 66, 6623–6634. [Google Scholar] [CrossRef] [PubMed]
- Parada, L.A.; Misteli, T. Chromosome positioning in the interphase, nucleus. Trends Cell Biol. 2002, 12, 425–432. [Google Scholar] [CrossRef]
- Schwarzacher, T.; Leitch, A.R.; Bennett, M.D.; Heslop-Harrison, J.S. In situ localization of parental genomes in a wide hybrid. Ann. Bot. 1989, 64, 315–324. [Google Scholar] [CrossRef]
- Schubert, I.; Shi, F.; Fuchs, J.; Endo, T.R. An efficient screening for terminal deletions and translocations of barley chromosomes added to common wheat. Plant J. 1998, 14, 489–495. [Google Scholar] [CrossRef]
- Robaszkiewicz, E.; Idziak-Helmcke, D.; Tkacz, M.A.; Chrominski, K.; Hasterok, R. The arrangement of Brachypodium distachyon chromosomes in interphase nuclei. J. Exp. Bot. 2016, 67, 5571–5583. [Google Scholar] [CrossRef] [PubMed]
- Fung, J.C.; Marshall, W.F.; Dernburg, A.; Agard, D.A.; Sedat, J.W. Homologous chromosome pairing in Drosophila melanogaster proceeds through multiple independent initiations. J. Cell Biol. 1998, 141, 5–20. [Google Scholar] [CrossRef]
- Baroux, C.; Pecinka, A.; Fuchs, J.; Kreth, G.; Schubert, I.; Grossniklaus, U. Non-random chromosome arrangement in triploid endosperm nuclei. Chromosoma 2017, 126, 115–124. [Google Scholar] [CrossRef]
- Hiraoka, Y.; Dernburg, A.F.; Parmelee, S.J.; Rykowski, M.C.; Agard, D.A.; Sedat, J.W. The onset of homologous chromosome-pairing during Drosophila melanogaster embryogenesis. J. Cell Biol. 1993, 120, 591–600. [Google Scholar] [CrossRef]
- Bornfleth, H.; Edelmann, P.; Zink, D.; Cremer, T.; Cremer, C. Quantitative motion analysis of subchromosomal foci in living cells using four-dimensional microscopy. Biophys. J. 1999, 77, 2871–2886. [Google Scholar] [CrossRef]
- Lucas, J.N.; Cervantes, E. Significant large-scale chromosome territory movement occurs as a result of mitosis, but not during interphase. Int. J. Rad. Biol. 2002, 78, 449–455. [Google Scholar] [CrossRef]
- Lam, E.; Kato, N.; Watanabe, K. Visualizing chromosome structure/organization. Annu. Rev. Plant Biol. 2004, 55, 537–554. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Lam, E. Chromatin of endoreduplicated pavement cells has greater range of movement than that of diploid guard cells in Arabidopsis thaliana. J. Cell Sci. 2003, 116, 2195–2201. [Google Scholar] [CrossRef] [PubMed]
- Gartenberg, M.R.; Neumann, F.R.; Laroche, T.; Blaszczyk, M.; Gasser, S.M. Sir-mediated repression can occur independently of chromosomal and subnuclear contexts. Cell 2004, 119, 955–967. [Google Scholar] [CrossRef] [PubMed]
- Tiang, C.L.; He, Y.; Pawlowski, W.P. Chromosome organization and dynamics during Interphase, mitosis, and meiosis in plants. Plant Physiol. 2012, 158, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Soutoglou, E.; Misteli, T. Mobility and immobility of chromatin in transcription and genome stability. Curr. Opin. Genet. Dev. 2007, 17, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasencakova, Z.; Meister, A.; Schubert, I. Chromatin organization and its relation to replication and histone acetylation during the cell cycle in barley. Chromosoma 2001, 110, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Bolzer, A.; Kreth, G.; Solovei, I.; Koehler, D.; Saracoglu, K.; Fauth, C.; Muller, S.; Eils, R.; Cremer, C.; Speicher, M.R.; et al. Three-dimensional maps of all chromosomes in human male fibroblast nuclei and prometaphase rosettes. PLoS Biol. 2005, 3, 826–842. [Google Scholar] [CrossRef] [PubMed]
- Bauer, E.; Schmutzer, T.; Barilar, I.; Mascher, M.; Gundlach, H.; Martis, M.M.; Twardziok, S.O.; Hackauf, B.; Gordillo, A.; Wilde, P.; et al. Towards a whole-genome sequence for rye (Secale cereale L.). Plant J. 2017, 89, 853–869. [Google Scholar] [CrossRef] [PubMed]
- Schwarzacher-Robinson, T.; Finch, R.A.; Smith, J.B.; Bennett, M.D. Genotypic control of centromere positions of parental genomes in Hordeum x Secale hybrid metaphases. J. Cell Sci. 1987, 87, 291–304. [Google Scholar]
- Leitch, A.R.; Schwarzacher, T.; Mosgoller, W.; Bennett, M.D.; Heslop-Harrison, J.S. Parental genomes are separated throughout the cell cycle in a plant hybrid. Chromosoma 1991, 101, 206–213. [Google Scholar] [CrossRef]
- Lukaszewski, A.J. Behavior of Centromeres in Univalents and Centric Misdivision in Wheat. Cytogenet. Genome Res. 2010, 129, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Pernickova, K.; Kolackova, V.; Lukaszewski, A.J.; Fan, C.L.; Vrana, J.; Duchoslav, M.; Jenkins, G.; Phillips, D.; Samajova, O.; Sedlarova, M.; et al. Instability of alien chromosome introgressions in wheat associated with improper positioning in the nucleus. Int. J. Mol. Sci. 2019, 20, 1448. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Nasuda, S.; Endo, T.R. A direct repeat sequence associated with the centromeric retrotransposons in wheat. Genome 2004, 47, 747–756. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Introgression | Introgressed Chromosome Arm Length3 (Mb) | Chromosome Length (Mb) | Arm Ratio | Nuclear Volume (µm3) | Arm Length (µm) | C-C (µm) | MA1-MA2 (µm) | T-T (µm) | CN-MA (µm) |
|---|---|---|---|---|---|---|---|---|---|
| 1AS.1RL | 626 | 902 | 2.27 | 1545 ± 228 | 7.73 ± 1.48 | 4.01 ± 1.98 | 5.11 ± 2.28 | 4.88 ± 1.71 | 3.55 ± 0.84a |
| 1RS.1BL | 423 | 959 | 1.27 | 1501 ± 362 | 8.21 ± 1.69 | 3.49 ± 1.50 | 4.42 ± 1.64 | 4.18 ± 1.66 | 3.57 ± 1.01a |
| 1RS.1DL | 423 | 804 | 0.90 | 1998 ± 285 | 6.86 ± 1.31 | 2.83 ± 1.15 | 3.55 ± 1.63 | 3.71 ± 1.74 | 2.82 ± 0.78b |
| 2RS.2BL | 595 | 1102 | 0.85 | 1597 ± 490 | 8.51 ± 2.58 | 3.38 ± 1.70 | 4.38 ± 2.04 | 4.09 ± 1.95 | 3.51 ± 1.22a |
| 2BS.2RL | 693 | 1116 | 1.64 | 1510 ± 357 | 8.96 ± 1.62 | 4.05 ± 1.80 | 4.99 ± 2.13 | 4.99 ± 2.25 | 3.76 ± 0.97a |
| 5RS.5BL | 346 | 928 | 1.68 | 1656 ± 329 | 6.95 ± 1.54 | 3.06 ± 1.29 | 3.68 ± 1.47 | 3.97 ± 1.48 | 2.99 ± 0.91b |
| Test statistics | F (dfg, dfe)p-Value | - | - | - | - | - | - | - | 13.55 (5, 195.6)*** |
| del1RS.1RL | S1: 271 | 897 | 2.31 | 1986 ± 390 | 4.04 ± 1.03 | 2.88 ± 1.37 | 3.52 ± 1.50 | 3.86 ± 1.79 | 2.82 ± 0.98a |
| L: 626 | 7.56 ± 1.67 | 3.87 ± 1.74 | 3.56 ± 1.87 | 3.13 ± 0.87A | |||||
| 1RS.del1RL | S: 423 | 861 | 1.04 | 2006 ± 500 | 6.18 ± 1.58 | 3.57 ± 1.69 | 4.19 ± 1.88 | 4.20 ± 1.92 | 3.05 ± 1.00a |
| L2: 438 | 5.21 ± 1.41 | 4.06 ± 1.83 | 4.12 ± 1.53 | 3.09 ± 1.00A | |||||
| 1R(1A) | S: 423 | 1049 | 1.48 | 1196 ± 143 | 8.58 ± 1.79 | 4.09 ± 2.41 | 5.45 ± 2.27 | 4.99 ± 2.32 | 4.12 ± 1.27b |
| L: 626 | 9.88 ± 1.87 | 5.22 ± 2.76 | 4.23 ± 2.47 | 4.09 ± 1.24B | |||||
| t1RS | 423 | 423 | - | 1190 ± 109 | 8.84 ± 1.77 | 4.15 ± 1.86 | 4.94 ± 2.26 | 4.67 ± 2.09 | 4.21 ± 1.19b |
| Test statistics | F (dfg, dfe)p-Value | - | - | - | - | - | - | - | S: 18.76 (3, 93.9.)*** L: 10.57 (2, 82.5)*** |
| 3B | S: 433 | 995 | 1.30 | 1535 ± 315 | 5.63 ± 1.47 | 4.30 ± 1.52 | 5.39 ± 1.7 | 5.35 ± 2.02 | 3.83 ± 0.98 |
| L: 562 | 6.91 ± 1.81 | 4.96 ± 1.79 | 4.22 ± 1.93 | 3.76 ± 1.03 | |||||
| 5B | S: 290 | 870 | 2.00 | 1492 ± 383 | 4.30 ± 1.67 | 4.48 ± 1.93 | 4.96 ± 1.86 | 4.82 ± 1.62 | 3.65 ± 1.24 |
| L: 580 | 6.96 ± 1.94 | 5.13 ± 1.86 | 3.61 ± 1.77 | 3.60 ± 1.07 | |||||
| Test statistics | F (dfg, dfe)p-Value | - | - | - | - | - | - | - | S: 0.54 (1, 74.2)n.s. L: 0.46 (1, 77.9)n.s. |
| 3H | S: 294 | 700 | 1.38 | 1316 ± 328 | 5.74 ± 1.48 | 3.7 ± 2.2 | 4.54 ± 2.26 | 5.25 ± 2.46 | 3.98 ± 1.43 |
| L: 406 | 6.96 ± 1.94 | 3.95 ± 1.98 | 4.36 ± 1.88 | 3.70 ± 1.25 | |||||
| 6H | S: 261 | 583 | 1.23 | 1848 ± 352 | 4.13 ± 1.78 (3.46 ± 1.08) | 3.77 ± 1.59 | 4.22 ± 1.71 | 4.99 ± 1.76 | 3.18 ± 1.00 |
| L: 322 | 5.68 ± 1.69 | 4.29 ± 2.08 | 4.71 ± 1.96 | 3.05 ± 1.08 | |||||
| Test statistics | F (dfg, dfe)p-Value | - | - | - | - | - | - | - | S: 8.30 (1, 69.9)** L: 6.13 (1, 76.4)* |
| 3HL | 406 | 406 | - | 1419 ± 358 | 8.19 ± 1.91 | 2.88 ± 1.49 | 3.81 ± 2 | 3.93 ± 2.64 | 3.65 ± 1.12 |
| 3HS | 294 | 294 | - | 2182 ± 646 | 4.14 ± 0.97 | 2.73 ± 1.66 | 3.08 ± 1.93 | 3.33 ± 1.99 | 3.19 ± 1.10 |
| 6HL | 322 | 322 | - | 1695 ± 307 | 5.46 ± 1.33 | 2.9 ± 2.06 | 3.98 ± 2.83 | 3.97 ± 2.97 | 3.34 ± 1.29 |
| 6HS | 261 | 261 | - | 1868 ± 609 | 6.65 ± 1.79 (4.23 ± 1.06) | 3.15 ± 1.76 | 4.26 ± 2.24 | 4.59 ± 2.45 | 3.35 ± 1.16 |
| Test statistics | F (dfg, dfe)p-Value | - | - | - | - | - | - | - | 1.37 (3, 92.7)n.s. |
| Triticale | Number of Nuclei | Nuclear Volume (µm3) | Rye Chromatin (%) | Wheat Chromatin (%) |
|---|---|---|---|---|
| 4× | 22 | 993 ± 260 | 53.6 ± 4.7 | 46.4 ± 4.7 |
| 6× | 22 | 1412 ± 274 | 37.7 ± 10.0 | 62.3 ± 10.1 |
| Introgression | Cell Cycle Stage | Nuclear Volume (µm3) | Arm Length (µm) | C-C (µm) | MA1-MA2 (µm) | T-T (µm) | MA-CN (µm) |
|---|---|---|---|---|---|---|---|
| 1AS.1RL | G1 | 1545 ± 228a | 7.73 ± 1.48a | 4.01 ± 1.98 | 5.11 ± 2.28 | 4.88 ± 1.71 | 3.55 ± 0.84a |
| S | 1791 ± 400b | 7.50 ± 1.60a | 3.17 ± 1.24 | 4.28 ± 1.76 | 3.73 ± 1.47 | 3.23 ± 0.95a | |
| G2 | 2562 ± 428c | 6.13 ± 1.15b | 2.12 ± 0.99 | 2.28 ± 0.74 | 2.63 ± 1.27 | 2.43 ± 0.57b | |
| Test statistics | F (dfg, dfe)p-Value | 104.6 (2, 77.2)*** | 20.8 (2, 85.9)*** | - | - | - | 33.3 (2, 83.2)*** |
| del1RS.1RL1 | G1 | 1986 ± 390a | S: 4.04 ± 1.03a | 2.88 ± 1.37 | 3.52 ± 1.50 | 3.86 ± 1.79 | 2.82 ± 0.98 |
| L: 7.56 ± 1.67A | 3.87 ± 1.74 | 3.56 ± 1.87 | 3.13 ± 0.87A | ||||
| S | 2118 ± 367a | S: 3.68 ± 0.85ab | 2.74 ± 1.45 | 3.45 ± 1.58 | 3.91 ± 1.61 | 2.70 ± 0.92 | |
| L: 6.26 ± 1.07B | 3.71 ± 1.71 | 3.09 ± 1.13 | 2.87 ± 0.91AB | ||||
| G2 | 2308 ± 246b | S: 3.38 ± 0.79b | 2.17 ± 0.91 | 2.59 ± 1.27 | 2.94 ± 1.54 | 2.54 ± 0.72 | |
| L: 6.25 ± 1.12B | 2.75 ± 1.46 | 2.15 ± 1.23 | 2.59 ± 0.67B | ||||
| Test statistics | F (dfg, dfe)p-Value | 12.7 (2, 88.1)*** | S: 6 (2, 90.5)** L: 12.1 (2, 89.5)*** | - | - | - | S: 1.3 (2, 90.0)n.s. L: 5.72 (2, 89.6)** |
| 1R(1A) | G1 | 1196 ± 143a | S: 8.58 ± 1.79a | 4.09 ± 2.41 | 5.45 ± 2.27 | 4.99 ± 2.32 | 4.12 ± 1.27 |
| L: 9.88 ± 1.87A | 5.22 ± 2.76 | 4.23 ± 2.47 | 4.09 ± 1.24A | ||||
| S | 2147 ± 507a | S: 5.88 ± 1.41a | 2.47 ± 1.08 | 3.49 ± 1.74 | 3.31 ± 1.83 | 2.74 ± 0.80 | |
| L: 6.66 ± 1.59B | 3.38 ± 1.77 | 2.79 ± 1.27 | 2.77 ± 0.85AB | ||||
| G2 | 2584 ± 450b | S: 5.32 ± 1.15b | 2.48 ± 1.02 | 3.29 ± 1.48 | 3.13 ± 1.11 | 2.33 ± 0.74 | |
| L: 6.03 ± 1.32B | 3.16 ± 1.37 | 2.46 ± 1.08 | 2.36 ± 0.65B | ||||
| Test statistics | F (dfg, dfe)p-Value | 12.7 (2, 88.1)*** | S: 6.0 (2, 90.5)** L: 12.1 (2, 89.5)*** | - | - | - | S: 1.3 (2, 90.0)n.s. L: 5.7 (2, 89.6)** |
| t1RS | G1 | 1190 ± 109a | 8.84 ± 1.77a | 4.15 ± 1.86 | 4.94 ± 2.26 | 4.67 ± 2.09 | 4.21 ± 1.19a |
| S | 2250 ± 367b | 5.72 ± 1.23b | 3.38 ± 1.69 | 3.77 ± 1.67 | 3.31 ± 1.68 | 2.59 ± 0.76b | |
| G2 | 2371 ± 402b | 5.64 ± 1.19b | 2.52 ± 1.3 | 3.21 ± 1.71 | 2.96 ± 1.51 | 2.39 ± 0.86b | |
| Test statistics | F (dfg, dfe)p-Value | 323.7 (2, 65.0)*** | 57.7 (2, 82.8)*** | - | - | - | 37.4 (2, 83.2)*** |
| Introgression | Tissue Type | Nuclear Volume (µm3) | Arm Length (µm) | C-C (µm) | MA1-MA2 (µm) | T-T (µm) | MA-CN (µm) |
|---|---|---|---|---|---|---|---|
| 1AS.1RL | Root tip | 1545 ± 228a | 7.73 ± 1.48a | 4.01 ± 1.98 | 5.11 ± 2.28 | 4.88 ± 1.71 | 3.55 ± 0.84a |
| Embryo | 1390 ± 277b | 8.43 ± 1.90a | 3.79 ± 2.00 | 4.65 ± 2.12 | 5.25 ± 2.59 | 3.56 ± 1.07a | |
| Leaf | 678 ± 73c | 11.69 ± 2.6b | 7.67 ± 3.56 | 6.79 ± 2.95 | 6.01 ± 2.8 | 5.58 ± 1.68b | |
| Test statistics | F (dfg, dfe)p-Value | 428.3 (2, 70.3)*** | 37.8 (2, 79.9)*** | - | - | - | 26.6 (2, 78.3)*** |
| del1RS.1RL1 | Root tip | 1986 ± 390a | S: 4.04 ± 1.03a | 2.88 ± 1.37 | 3.52 ± 1.50 | 3.86 ± 1.79 | 2.82 ± 0.98a |
| L: 7.56 ± 1.67A | 3.87 ± 1.74 | 3.56 ± 1.87 | 3.13 ± 0.87A | ||||
| Embryo | 1210 ± 188b | S: 4.45 ± 0.97a | 4.76 ± 2.31 | 5.13 ± 2.50 | 5.52 ± 2.35 | 4.33 ± 1.06b | |
| L: 8.78 ± 2.03B | 4.85 ± 2.58 | 5.40 ± 3.23 | 4.23 ± 1.14B | ||||
| Leaf | 626 ± 131c | S: 5.75 ± 1.74b | 7.10± 3.25 | 8.73 ± 3.07 | 9.46 ± 3.01 | 6.58 ± 2.30c | |
| L: 12.36 ± 2.98C | 8.48 ± 3.88 | 7.8± 3.86 | 6.21 ± 2.44C | ||||
| Test statistics | F (dfg, dfe)p-Value | 337.7 (2, 82.3)*** | S: 14.9 (2, 79.0)*** L: 41.1 (2, 77.6)*** | - | - | - | S: 58.9 (2, 75.6)*** L: 36.6 (2, 72.8)*** |
| Introgression | Complete Separation | Partial Association | Complete Association |
|---|---|---|---|
| 1AS.1RL (embryo, G1) | 19 | 1 | 2 |
| 1AS.1RL (leaf, G1) | 16 | 4 | 1 |
| 1AS.1RL (root tip, G1) | 22 | 3 | |
| 1AS.1RL (root tip, S) | 18 | 3 | |
| 1AS.1RL (root tip, G2) | 20 | 2 | 1 |
| 1RS.1BL | 32 | 7 | 1 |
| 1RS.1DL | 37 | 3 | |
| 2RS.2BL | 32 | 8 | |
| 2BS.2RL | 33 | 6 | 1 |
| 5RS.5BL | 31 | 7 | 2 |
| del1RS.1RL 1 (embryo, G1) | 16 | 4 | 2 |
| del1RS.1RL 1 (leaf, G1) | 17 | 3 | |
| del1RS.1RL 1 (root tip, G1) | 22 | 3 | |
| del1RS.1RL 1 (root tip, S) | 21 | 1 | 1 |
| del1RS.1RL 1 (root tip, G2) | 15 | 6 | 1 |
| 1RS.del1RL 2 | 17 | 3 | 1 |
| 1R(1A) (G1) | 15 | 5 | 2 |
| 1R(1A) (S) | 15 | 4 | 1 |
| 1R(1A) (G2) | 18 | 2 | 2 |
| t1RS (G1) | 20 | 1 | 1 |
| t1RS (S) | 20 | 1 | |
| t1RS (G2) | 19 | 2 | 2 |
| 3B | 20 | ||
| 5B | 18 | 2 | |
| 3H | 14 | 3 | 3 |
| 3HL | 18 | 4 | 2 |
| 3HS | 18 | 2 | 2 |
| 6H | 17 | 2 | 1 |
| 6HL | 14 | 4 | 2 |
| 6HS | 19 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koláčková, V.; Perničková, K.; Vrána, J.; Duchoslav, M.; Jenkins, G.; Phillips, D.; Turkosi, E.; Šamajová, O.; Sedlářová, M.; Šamaj, J.; et al. Nuclear Disposition of Alien Chromosome Introgressions into Wheat and Rye Using 3D-FISH. Int. J. Mol. Sci. 2019, 20, 4143. https://doi.org/10.3390/ijms20174143
Koláčková V, Perničková K, Vrána J, Duchoslav M, Jenkins G, Phillips D, Turkosi E, Šamajová O, Sedlářová M, Šamaj J, et al. Nuclear Disposition of Alien Chromosome Introgressions into Wheat and Rye Using 3D-FISH. International Journal of Molecular Sciences. 2019; 20(17):4143. https://doi.org/10.3390/ijms20174143
Chicago/Turabian StyleKoláčková, Veronika, Kateřina Perničková, Jan Vrána, Martin Duchoslav, Glyn Jenkins, Dylan Phillips, Edina Turkosi, Olga Šamajová, Michaela Sedlářová, Jozef Šamaj, and et al. 2019. "Nuclear Disposition of Alien Chromosome Introgressions into Wheat and Rye Using 3D-FISH" International Journal of Molecular Sciences 20, no. 17: 4143. https://doi.org/10.3390/ijms20174143