Innate Immune Cells and Toll-like Receptor–Dependent Responses at the Maternal–Fetal Interface

 ,
,  ,
,  and
and
Abstract
:1. Innate Cell Populations at the Maternal–Fetal Interface
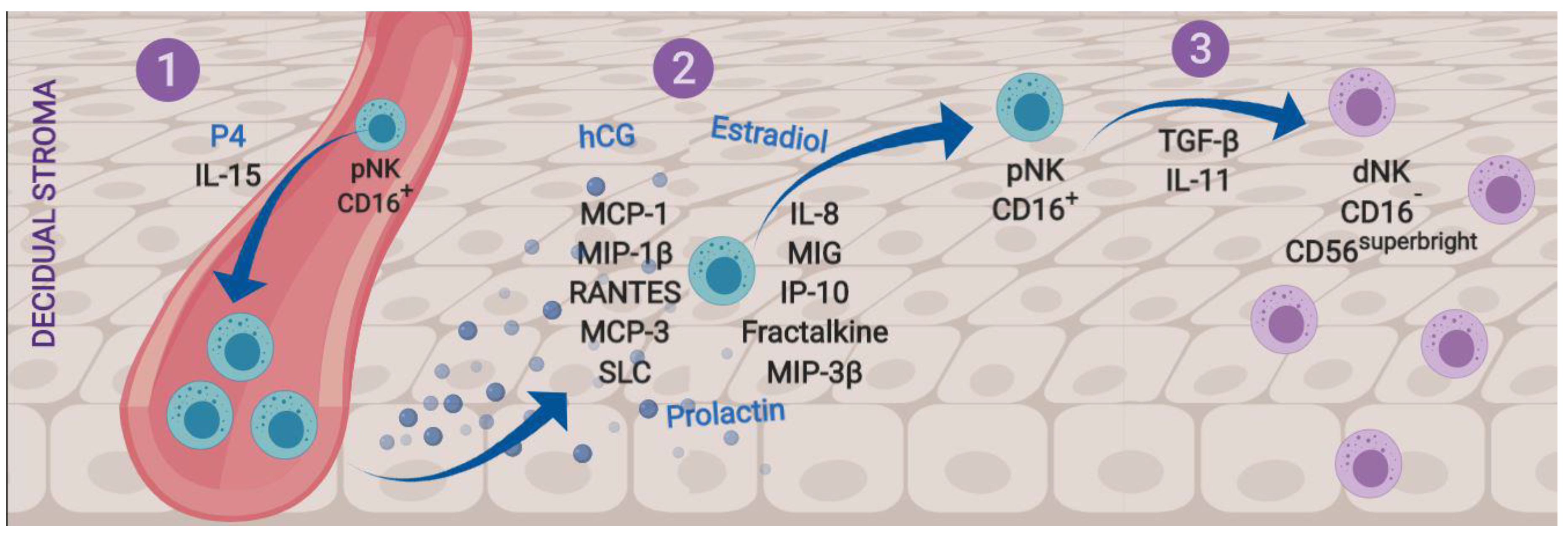
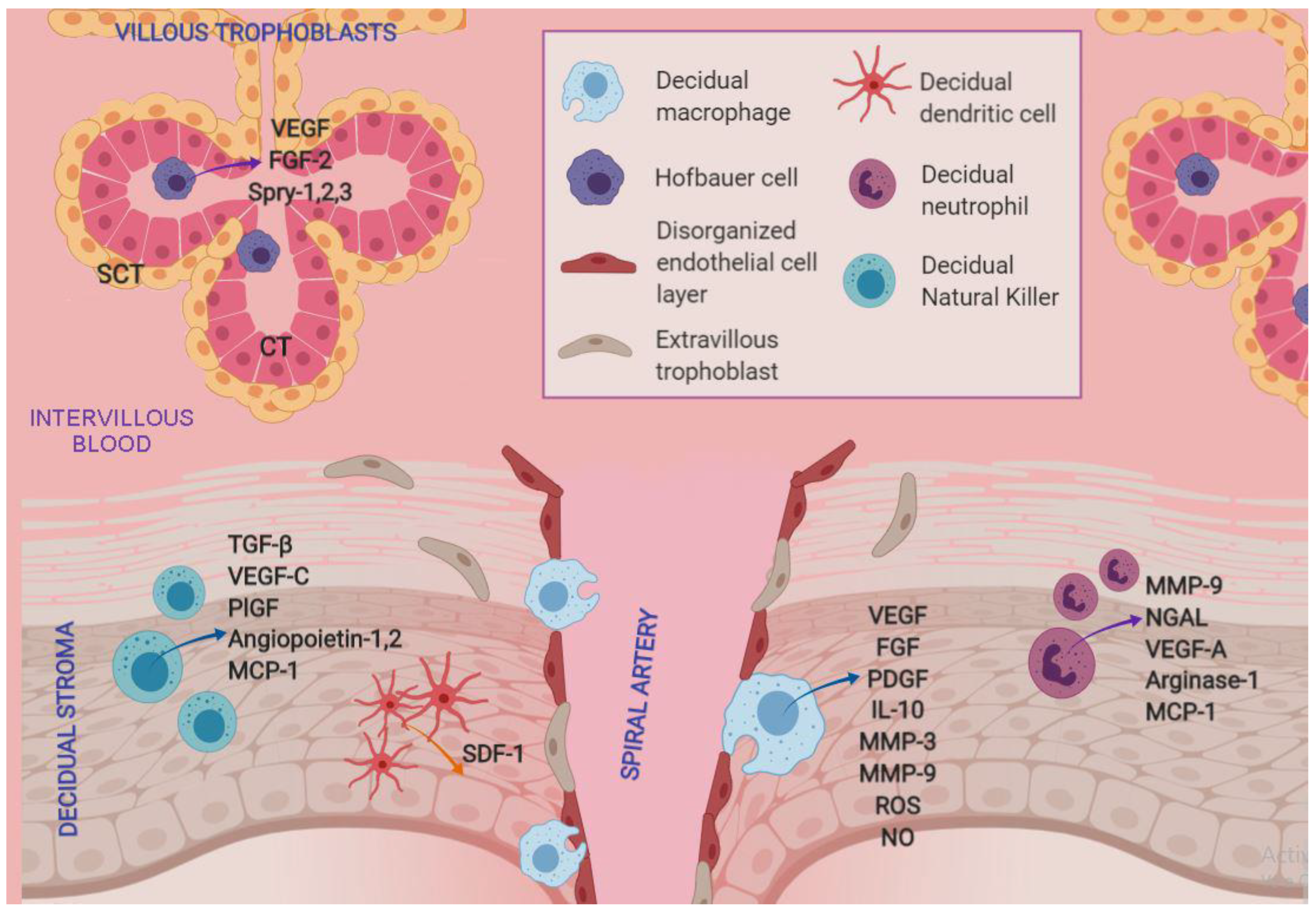
1.1. Natural Killer Cells
1.2. Macrophages
Hofbauer Cells
1.3. Dendritic Cells
1.4. Neutrophils
2. TLR-Dependent Immune Responses During Pregnancy
2.1. Toll-Like Receptors: Structures and Signaling Pathways
2.2. TLRs During Pregnancy
3. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Svensson-Arvelund, J.; Mehta, R.B.; Lindau, R.; Mirrasekhian, E.; Rodriguez-Martinez, H.; Berg, G.; Lash, G.E.; Jenmalm, M.C.; Ernerudh, J. The human fetal placenta promotes tolerance against the semiallogeneic fetus by inducing regulatory T cells and homeostatic M2 macrophages. J. Immunol. 2015, 194, 1534–1544. [Google Scholar] [CrossRef] [PubMed]
- Sacks, G.P.; Studena, K.; Sargent, K.; Redman, C.W. Normal pregnancy and preeclampsia both produce inflammatory changes in peripheral blood leukocytes akin to those of sepsis. Am. J. Obstet. Gynecol. 1998, 179, 80–86. [Google Scholar] [CrossRef]
- Harmon, A.C.; Cornelius, D.C.; Amaral, L.M.; Faulkner, J.L.; Cunningham, M.W., Jr.; Wallace, K.; LaMarca, B. The role of inflammation in the pathology of preeclampsia. Clin. Sci. (Lond.) 2016, 130, 409–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantham, P.; Aye, I.L.; Powell, T.L. Inflammation in maternal obesity and gestational diabetes mellitus. Placenta 2015, 36, 709–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauguel-de Mouzon, S.; Guerre-Millo, M. The placenta cytokine network and inflammatory signals. Placenta 2006, 27, 794–798. [Google Scholar] [CrossRef] [PubMed]
- Olmos-Ortiz, A.; Garcia-Quiroz, J.; Avila, E.; Caldino-Soto, F.; Halhali, A.; Larrea, F.; Diaz, L. Lipopolysaccharide and cAMP modify placental calcitriol biosynthesis reducing antimicrobial peptides gene expression. Am. J. Reprod. Immunol. 2018, 79, e12841. [Google Scholar] [CrossRef] [PubMed]
- Cristiani, C.M.; Palella, E.; Sottile, R.; Tallerico, R.; Garofalo, C.; Carbone, E. Human NK Cell Subsets in Pregnancy and Disease: Toward a New Biological Complexity. Front. Immunol. 2016, 7, 656. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kim, C.J.; Kim, D.J.; Kang, J.H. Immune cells in the female reproductive tract. Immune Netw. 2015, 15, 16–26. [Google Scholar] [CrossRef]
- Lurie, S.; Rahamim, E.; Piper, I.; Golan, A.; Sadan, O. Total and differential leukocyte counts percentiles in normal pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 136, 16–19. [Google Scholar] [CrossRef]
- Mahnke, Y.D.; Beddall, M.H.; Roederer, M. OMIP-029: Human NK-cell phenotypization. Cytom. Part A J. Int. Soc. Anal. Cytol. 2015, 87, 986–988. [Google Scholar] [CrossRef]
- Lanier, L.L.; Le, A.M.; Civin, C.I.; Loken, M.R.; Phillips, J.H. The relationship of CD16 (Leu-11) and Leu-19 (NKH-1) antigen expression on human peripheral blood NK cells and cytotoxic T lymphocytes. J. Immunol. 1986, 136, 4480–4486. [Google Scholar] [PubMed]
- Laskarin, G.; Tokmadzic, V.S.; Strbo, N.; Bogovic, T.; Szekeres-Bartho, J.; Randic, L.; Podack, E.R.; Rukavina, D. Progesterone induced blocking factor (PIBF) mediates progesterone induced suppression of decidual lymphocyte cytotoxicity. Am. J. Reprod. Immunol. 2002, 48, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, A.; Berta, G.; Szekeres-Bartho, J. PIBF positive uterine NK cells in the mouse decidua. J. Reprod. Immunol. 2017, 119, 38–43. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Allan, D.S.; Bowen, M.; Powis, S.J.; Joseph, S.; Verma, S.; Hiby, S.E.; McMichael, A.J.; Loke, Y.W.; Braud, V.M. HLA-E is expressed on trophoblast and interacts with CD94/NKG2 receptors on decidual NK cells. Eur. J. Immunol. 2000, 30, 1623–1631. [Google Scholar] [CrossRef]
- Navarro, F.; Llano, M.; Bellon, T.; Colonna, M.; Geraghty, D.E.; Lopez-Botet, M. The ILT2(LIR1) and CD94/NKG2A NK cell receptors respectively recognize HLA-G1 and HLA-E molecules co-expressed on target cells. Eur. J. Immunol. 1999, 29, 277–283. [Google Scholar] [CrossRef]
- Chazara, O.; Xiong, S.; Moffett, A. Maternal KIR and fetal HLA-C: A fine balance. J. Leukoc. Biol. 2011, 90, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Moffett-King, A. Natural killer cells and pregnancy. Nat. Rev. Immunol. 2002, 2, 656–663. [Google Scholar] [CrossRef]
- Gaynor, L.M.; Colucci, F. Uterine Natural Killer Cells: Functional Distinctions and Influence on Pregnancy in Humans and Mice. Front. Immunol. 2017, 8, 467. [Google Scholar] [CrossRef] [Green Version]
- Fernekorn, U.; Kruse, A. Regulation of leukocyte recruitment to the murine maternal/fetal interface. Chem. Immunol. Allergy 2005, 89, 105–117. [Google Scholar] [CrossRef]
- Williams, P.J.; Searle, R.F.; Robson, S.C.; Innes, B.A.; Bulmer, J.N. Decidual leucocyte populations in early to late gestation normal human pregnancy. J. Reprod. Immunol. 2009, 82, 24–31. [Google Scholar] [CrossRef]
- Wang, F.; Zhu, H.; Li, B.; Liu, M.; Liu, D.; Deng, M.; Wang, Y.; Xia, X.; Jiang, Q.; Chen, D. Effects of human chorionic gonadotropin, estradiol, and progesterone on interleukin-18 expression in human decidual tissues. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2017, 33, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Wilkens, J.; Male, V.; Ghazal, P.; Forster, T.; Gibson, D.A.; Williams, A.R.; Brito-Mutunayagam, S.L.; Craigon, M.; Lourenco, P.; Cameron, I.T.; et al. Uterine NK cells regulate endometrial bleeding in women and are suppressed by the progesterone receptor modulator asoprisnil. J. Immunol. 2013, 191, 2226–2235. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Hiby, S.E.; Loke, Y.W.; King, A. Human decidual natural killer cells express the receptor for and respond to the cytokine interleukin 15. Biol. Reprod. 2000, 62, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Kitaya, K.; Yamaguchi, T.; Honjo, H. Central role of interleukin-15 in postovulatory recruitment of peripheral blood CD16(-) natural killer cells into human endometrium. J. Clin. Endocrinol. Metab. 2005, 90, 2932–2940. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Yun, S.; Ryu, B.J.; Han, A.R.; Lee, S.K. Trophoblasts regulate natural killer cells via control of interleukin-15 receptor signaling. Am. J. Reprod. Immunol. 2017, 78, e12628. [Google Scholar] [CrossRef] [PubMed]
- Santoni, A.; Carlino, C.; Gismondi, A. Uterine NK cell development, migration and function. Reprod. Biomed. Online 2008, 16, 202–210. [Google Scholar] [CrossRef]
- Carlino, C.; Stabile, H.; Morrone, S.; Bulla, R.; Soriani, A.; Agostinis, C.; Bossi, F.; Mocci, C.; Sarazani, F.; Tedesco, F.; et al. Recruitment of circulating NK cells through decidual tissues: A possible mechanism controlling NK cell accumulation in the uterus during early pregnancy. Blood 2008, 111, 3108–3115. [Google Scholar] [CrossRef]
- Gibson, D.A.; Greaves, E.; Critchley, H.O.; Saunders, P.T. Estrogen-dependent regulation of human uterine natural killer cells promotes vascular remodelling via secretion of CCL2. Hum. Reprod. 2015, 30, 1290–1301. [Google Scholar] [CrossRef]
- Kane, N.; Kelly, R.; Saunders, P.T.; Critchley, H.O. Proliferation of uterine natural killer cells is induced by human chorionic gonadotropin and mediated via the mannose receptor. Endocrinology 2009, 150, 2882–2888. [Google Scholar] [CrossRef]
- Keskin, D.B.; Allan, D.S.; Rybalov, B.; Andzelm, M.M.; Stern, J.N.; Kopcow, H.D.; Koopman, L.A.; Strominger, J.L. TGFbeta promotes conversion of CD16+ peripheral blood NK cells into CD16- NK cells with similarities to decidual NK cells. Proc. Natl. Acad. Sci. USA 2007, 104, 3378–3383. [Google Scholar] [CrossRef]
- Ain, R.; Trinh, M.L.; Soares, M.J. Interleukin-11 signaling is required for the differentiation of natural killer cells at the maternal-fetal interface. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2004, 231, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Manaster, I.; Mizrahi, S.; Goldman-Wohl, D.; Sela, H.Y.; Stern-Ginossar, N.; Lankry, D.; Gruda, R.; Hurwitz, A.; Bdolah, Y.; Haimov-Kochman, R.; et al. Endometrial NK cells are special immature cells that await pregnancy. J. Immunol. 2008, 181, 1869–1876. [Google Scholar] [CrossRef] [PubMed]
- Guimond, M.J.; Luross, J.A.; Wang, B.; Terhorst, C.; Danial, S.; Croy, B.A. Absence of natural killer cells during murine pregnancy is associated with reproductive compromise in TgE26 mice. Biol. Reprod. 1997, 56, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Charalambous, F.; Elia, A.; Georgiades, P. Decidual spiral artery remodeling during early post-implantation period in mice: Investigation of associations with decidual uNK cells and invasive trophoblast. Biochem. Biophys. Res. Commun. 2012, 417, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F.; Charnock-Jones, D.S.; Zhang, E.; Hiby, S.; Malik, S.; Day, K.; Licence, D.; Bowen, J.M.; Gardner, L.; King, A.; et al. Angiogenic growth factor messenger ribonucleic acids in uterine natural killer cells. J. Clin. Endocrinol. Metab. 2001, 86, 1823–1834. [Google Scholar] [CrossRef] [PubMed]
- Lash, G.E.; Schiessl, B.; Kirkley, M.; Innes, B.A.; Cooper, A.; Searle, R.F.; Robson, S.C.; Bulmer, J.N. Expression of angiogenic growth factors by uterine natural killer cells during early pregnancy. J. Leukoc. Biol. 2006, 80, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Faas, M.M.; de Vos, P. Uterine NK cells and macrophages in pregnancy. Placenta 2017, 56, 44–52. [Google Scholar] [CrossRef]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef]
- Brown, M.B.; von Chamier, M.; Allam, A.B.; Reyes, L. M1/M2 macrophage polarity in normal and complicated pregnancy. Front. Immunol. 2014, 5, 606. [Google Scholar] [CrossRef]
- Houser, B.L.; Tilburgs, T.; Hill, J.; Nicotra, M.L.; Strominger, J.L. Two unique human decidual macrophage populations. J. Immunol. 2011, 186, 2633–2642. [Google Scholar] [CrossRef]
- Bulmer, J.N.; Morrison, L.; Longfellow, M.; Ritson, A.; Pace, D. Granulated lymphocytes in human endometrium: Histochemical and immunohistochemical studies. Hum. Reprod. 1991, 6, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Ramhorst, R.; Grasso, E.; Paparini, D.; Hauk, V.; Gallino, L.; Calo, G.; Vota, D.; Perez Leiros, C. Decoding the chemokine network that links leukocytes with decidual cells and the trophoblast during early implantation. Cell Adhes. Migr. 2016, 10, 197–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, F.; Liu, H.; Lash, G.E. The Role of Decidual Macrophages During Normal and Pathological Pregnancy. Am. J. Reprod. Immunol. 2016, 75, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Gnainsky, Y.; Granot, I.; Aldo, P.; Barash, A.; Or, Y.; Mor, G.; Dekel, N. Biopsy-induced inflammatory conditions improve endometrial receptivity: The mechanism of action. Reproduction 2015, 149, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Jasper, M.J.; Hull, M.L.; Aplin, J.D.; Robertson, S.A. Macrophages regulate expression of alpha1,2-fucosyltransferase genes in human endometrial epithelial cells. Mol. Hum. Reprod. 2012, 18, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Rozner, A.E.; Durning, M.; Kropp, J.; Wiepz, G.J.; Golos, T.G. Macrophages modulate the growth and differentiation of rhesus monkey embryonic trophoblasts. Am. J. Reprod. Immunol. 2016, 76, 364–375. [Google Scholar] [CrossRef] [Green Version]
- Kang, X.; Zhang, X.; Zhao, A. Macrophage depletion and TNF-alpha inhibition prevent resorption in CBA/J x DBA/2 model of CpG-induced abortion. Biochem. Biophys. Res. Commun. 2016, 469, 704–710. [Google Scholar] [CrossRef]
- Heikkinen, J.; Mottonen, M.; Komi, J.; Alanen, A.; Lassila, O. Phenotypic characterization of human decidual macrophages. Clin. Exp. Immunol. 2003, 131, 498–505. [Google Scholar] [CrossRef]
- Lash, G.E.; Pitman, H.; Morgan, H.L.; Innes, B.A.; Agwu, C.N.; Bulmer, J.N. Decidual macrophages: Key regulators of vascular remodeling in human pregnancy. J. Leukoc. Biol. 2016, 100, 315–325. [Google Scholar] [CrossRef]
- Smith, S.D.; Dunk, C.E.; Aplin, J.D.; Harris, L.K.; Jones, R.L. Evidence for immune cell involvement in decidual spiral arteriole remodeling in early human pregnancy. Am. J. Pathol. 2009, 174, 1959–1971. [Google Scholar] [CrossRef]
- Moore, S.G.; McCabe, M.S.; Green, J.C.; Newsom, E.M.; Lucy, M.C. The transcriptome of the endometrium and placenta is associated with pregnancy development but not lactation status in dairy cows. Biol. Reprod. 2017, 97, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, V.M.; Kim, Y.M.; Straszewski, S.L.; Romero, R.; Mor, G. Macrophages and apoptotic cell clearance during pregnancy. Am. J. Reprod. Immunol. 2004, 51, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, M.; Hou, Y.; Chen, S.; Zhang, X.; Zhang, M.; Ji, X. Role of decidual CD14(+) macrophages in the homeostasis of maternal-fetal interface and the differentiation capacity of the cells during pregnancy and parturition. Placenta 2016, 38, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Romero, R.; Miller, D.; Kadam, L.; Mial, T.N.; Plazyo, O.; Garcia-Flores, V.; Hassan, S.S.; Xu, Z.; Tarca, A.L.; et al. An M1-like Macrophage Polarization in Decidual Tissue during Spontaneous Preterm Labor That Is Attenuated by Rosiglitazone Treatment. J. Immunol. 2016, 196, 2476–2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubicke, A.; Ekman-Ordeberg, G.; Mazurek, P.; Miller, L.; Yellon, S.M. Density of Stromal Cells and Macrophages Associated with Collagen Remodeling in the Human Cervix in Preterm and Term Birth. Reprod. Sci. 2016, 23, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Egashira, M.; Hirota, Y.; Shimizu-Hirota, R.; Saito-Fujita, T.; Haraguchi, H.; Matsumoto, L.; Matsuo, M.; Hiraoka, T.; Tanaka, T.; Akaeda, S.; et al. F4/80+ macrophages contribute to clearance of senescent cells in the mouse postpartum uterus. Endocrinology 2017, 158, 2344–2353. [Google Scholar] [CrossRef] [PubMed]
- Young, O.M.; Tang, Z.; Niven-Fairchild, T.; Tadesse, S.; Krikun, G.; Norwitz, E.R.; Mor, G.; Abrahams, V.M.; Guller, S. Toll-like receptor-mediated responses by placental Hofbauer cells (HBCs): A potential pro-inflammatory role for fetal M2 macrophages. Am. J. Reprod. Immunol. 2015, 73, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Reyes, L.; Wolfe, B.; Golos, T. Hofbauer cells: Placental macrophages of fetal origin. Chapter 3. In Macrophages: Origin, Functions and Biointervention; Kloc, M., Ed.; Springer: Houston, TX, USA, 2017; Volume 62, pp. 45–60. [Google Scholar]
- Kim, J.S.; Romero, R.; Kim, M.R.; Kim, Y.M.; Friel, L.; Espinoza, J.; Kim, C.J. Involvement of Hofbauer cells and maternal T cells in villitis of unknown aetiology. Histopathology 2008, 52, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Seval, Y.; Korgun, E.T.; Demir, R. Hofbauer cells in early human placenta: Possible implications in vasculogenesis and angiogenesis. Placenta 2007, 28, 841–845. [Google Scholar] [CrossRef]
- Grigoriadis, C.; Tympa, A.; Creatsa, M.; Bakas, P.; Liapis, A.; Kondi-Pafiti, A.; Creatsas, G. Hofbauer cells morphology and density in placentas from normal and pathological gestations. Rev. Bras. Ginecol. Obstet. Rev. Fed. Bras. Soc. Ginecol. Obstet. 2013, 35, 407–412. [Google Scholar] [CrossRef]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.L.; Chakraborty, R. Placental Hofbauer cells limit HIV-1 replication and potentially offset mother to child transmission (MTCT) by induction of immunoregulatory cytokines. Retrovirology 2012, 9, 101. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Romero, R.; Tarca, A.L.; Bhatti, G.; Kim, C.J.; Lee, J.; Elsey, A.; Than, N.G.; Chaiworapongsa, T.; Hassan, S.S.; et al. Methylome of fetal and maternal monocytes and macrophages at the feto-maternal interface. Am. J. Reprod. Immunol. 2012, 68, 8–27. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ming, X.F. Functions of arginase isoforms in macrophage inflammatory responses: Impact on cardiovascular diseases and metabolic disorders. Front. Immunol. 2014, 5, 533. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Harada, T.; Koi, H.; Kubota, T.; Azuma, H.; Aso, T. Identification of arginase in human placental villi. Placenta 2007, 28, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Holm, M.B.; Bastani, N.E.; Holme, A.M.; Zucknick, M.; Jansson, T.; Refsum, H.; Morkrid, L.; Blomhoff, R.; Henriksen, T.; Michelsen, T.M. Uptake and release of amino acids in the fetal-placental unit in human pregnancies. PLoS ONE 2017, 12, e0185760. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.L.; Guo, Y.; So, K.H.; Vijayan, M.; Wong, V.H.; Yao, Y.; Lee, K.F.; Chiu, P.C.; Yeung, W.S. Soluble human leukocyte antigen G5 polarizes differentiation of macrophages toward a decidual macrophage-like phenotype. Hum. Reprod. 2015, 30, 2263–2274. [Google Scholar] [CrossRef] [Green Version]
- Hunt, J.S.; Langat, D.L. HLA-G: A human pregnancy-related immunomodulator. Curr. Opin. Pharmacol. 2009, 9, 462–469. [Google Scholar] [CrossRef]
- Abumaree, M.H.; Al Jumah, M.A.; Kalionis, B.; Jawdat, D.; Al Khaldi, A.; Abomaray, F.M.; Fatani, A.S.; Chamley, L.W.; Knawy, B.A. Human placental mesenchymal stem cells (pMSCs) play a role as immune suppressive cells by shifting macrophage differentiation from inflammatory M1 to anti-inflammatory M2 macrophages. Stem Cell Rev. 2013, 9, 620–641. [Google Scholar] [CrossRef]
- Sisino, G.; Bouckenooghe, T.; Aurientis, S.; Fontaine, P.; Storme, L.; Vambergue, A. Diabetes during pregnancy influences Hofbauer cells, a subtype of placental macrophages, to acquire a pro-inflammatory phenotype. Biochim. Biophys. Acta 2013, 1832, 1959–1968. [Google Scholar] [CrossRef] [Green Version]
- Schliefsteiner, C.; Peinhaupt, M.; Kopp, S.; Logl, J.; Lang-Olip, I.; Hiden, U.; Heinemann, A.; Desoye, G.; Wadsack, C. Human Placental Hofbauer Cells Maintain an Anti-inflammatory M2 Phenotype despite the Presence of Gestational Diabetes Mellitus. Front. Immunol. 2017, 8, 888. [Google Scholar] [CrossRef] [PubMed]
- Satosar, A.; Ramirez, N.C.; Bartholomew, D.; Davis, J.; Nuovo, G.J. Histologic correlates of viral and bacterial infection of the placenta associated with severe morbidity and mortality in the newborn. Hum. Pathol. 2004, 35, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, A.Z.; Yu, W.; Hill, D.A.; Reyes, C.A.; Schwartz, D.A. Placental Pathology of Zika Virus: Viral Infection of the Placenta Induces Villous Stromal Macrophage (Hofbauer Cell) Proliferation and Hyperplasia. Arch. Pathol. Lab. Med. 2017, 141, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Ambuhl, L.M.M.; Leonhard, A.K.; Widen Zakhary, C.; Jorgensen, A.; Blaakaer, J.; Dybkaer, K.; Baandrup, U.; Uldbjerg, N.; Sorensen, S. Human papillomavirus infects placental trophoblast and Hofbauer cells, but appears not to play a causal role in miscarriage and preterm labor. Acta Obstet. Gynecol. Scand. 2017, 96, 1188–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simoni, M.K.; Jurado, K.A.; Abrahams, V.M.; Fikrig, E.; Guller, S. Zika virus infection of Hofbauer cells. Am. J. Reprod. Immunol. 2017, 77. [Google Scholar] [CrossRef] [PubMed]
- Loegl, J.; Hiden, U.; Nussbaumer, E.; Schliefsteiner, C.; Cvitic, S.; Lang, I.; Wadsack, C.; Huppertz, B.; Desoye, G. Hofbauer cells of M2a, M2b and M2c polarization may regulate feto-placental angiogenesis. Reproduction 2016, 152, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anteby, E.Y.; Natanson-Yaron, S.; Greenfield, C.; Goldman-Wohl, D.; Haimov-Kochman, R.; Holzer, H.; Yagel, S. Human placental Hofbauer cells express sprouty proteins: A possible modulating mechanism of villous branching. Placenta 2005, 26, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Kammerer, U.; Eggert, A.O.; Kapp, M.; McLellan, A.D.; Geijtenbeek, T.B.; Dietl, J.; van Kooyk, Y.; Kampgen, E. Unique appearance of proliferating antigen-presenting cells expressing DC-SIGN (CD209) in the decidua of early human pregnancy. Am. J. Pathol. 2003, 162, 887–896. [Google Scholar] [CrossRef]
- Barrientos, G.; Tirado-Gonzalez, I.; Freitag, N.; Kobelt, P.; Moschansky, P.; Klapp, B.F.; Thijssen, V.L.; Blois, S.M. CXCR4(+) dendritic cells promote angiogenesis during embryo implantation in mice. Angiogenesis 2013, 16, 417–427. [Google Scholar] [CrossRef]
- Collins, M.K.; Tay, C.S.; Erlebacher, A. Dendritic cell entrapment within the pregnant uterus inhibits immune surveillance of the maternal/fetal interface in mice. J. Clin. Investig. 2009, 119, 2062–2073. [Google Scholar] [CrossRef] [Green Version]
- Della Bella, S.; Giannelli, S.; Cozzi, V.; Signorelli, V.; Cappelletti, M.; Cetin, I.; Villa, M.L. Incomplete activation of peripheral blood dendritic cells during healthy human pregnancy. Clin. Exp. Immunol. 2011, 164, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Bachy, V.; Williams, D.J.; Ibrahim, M.A. Altered dendritic cell function in normal pregnancy. J. Reprod. Immunol. 2008, 78, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.; Santner-Nanan, B.; Dahlstrom, J.E.; Fadia, M.; Chandra, A.; Peek, M.; Nanan, R. Altered decidual DC-SIGN+ antigen-presenting cells and impaired regulatory T-cell induction in preeclampsia. Am. J. Pathol. 2012, 181, 2149–2160. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhang, H.; Chen, X.; Diao, L.; Lian, R.; Zhang, X.; Hu, L.; Zeng, Y. Association of peripheral blood dendritic cells with recurrent pregnancy loss: A case-controlled study. Am. J. Reprod. Immunol. 2016, 76, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Bozorgmehr, M.; Zarnani, A.H.; Nikoo, S.; Moazzeni, S.M. Suppressive effect of pregnant serum on murine dendritic cell function. J. Obstet. Gynaecol. Res. 2012, 38, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Zarnani, A.H.; Moazzeni, S.M.; Shokri, F.; Salehnia, M.; Dokouhaki, P.; Ghods, R.; Mahmoodi, A.R.; Jeddi-Tehrani, M. Microenvironment of the feto-maternal interface protects the semiallogenic fetus through its immunomodulatory activity on dendritic cells. Fertil. Steril. 2008, 90, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Segerer, S.E.; Muller, N.; van den Brandt, J.; Kapp, M.; Dietl, J.; Reichardt, H.M.; Rieger, L.; Kammerer, U. Impact of female sex hormones on the maturation and function of human dendritic cells. Am. J. Reprod. Immunol. 2009, 62, 165–173. [Google Scholar] [CrossRef]
- Huang, J.; Burke, P.; Yang, Y.; Seiss, K.; Beamon, J.; Cung, T.; Toth, I.; Pereyra, F.; Lichterfeld, M.; Yu, X.G. Soluble HLA-G inhibits myeloid dendritic cell function in HIV-1 infection by interacting with leukocyte immunoglobulin-like receptor B2. J. Virol. 2010, 84, 10784–10791. [Google Scholar] [CrossRef]
- Dauven, D.; Ehrentraut, S.; Langwisch, S.; Zenclussen, A.C.; Schumacher, A. Immune Modulatory Effects of Human Chorionic Gonadotropin on Dendritic Cells Supporting Fetal Survival in Murine Pregnancy. Front. Endocrinol. (Lausanne) 2016, 7, 146. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.F.; Knubel, C.P.; Sanchez, M.C.; Cervi, L.; Motran, C.C. Pregnancy-specific glycoprotein 1a activates dendritic cells to provide signals for Th17-, Th2-, and Treg-cell polarization. Eur. J. Immunol. 2012, 42, 1573–1584. [Google Scholar] [CrossRef]
- Remes Lenicov, F.; Paletta, A.L.; Gonzalez Prinz, M.; Varese, A.; Pavillet, C.E.; Lopez Malizia, A.; Sabatte, J.; Geffner, J.R.; Ceballos, A. Prostaglandin E2 Antagonizes TGF-beta Actions During the Differentiation of Monocytes Into Dendritic Cells. Front. Immunol. 2018, 9, 1441. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.P.; Willekens, B.; Cras, P.; Goossens, H.; Martinez-Caceres, E.; Berneman, Z.N.; Cools, N. Immunomodulatory Effects of 1,25-Dihydroxyvitamin D3 on Dendritic Cells Promote Induction of T Cell Hyporesponsiveness to Myelin-Derived Antigens. J. Immunol. Res. 2016, 2016, 5392623. [Google Scholar] [CrossRef] [PubMed]
- Della Bella, S.; Nicola, S.; Timofeeva, I.; Villa, M.L.; Santoro, A.; Berardi, A.C. Are interleukin-16 and thrombopoietin new tools for the in vitro generation of dendritic cells? Blood 2004, 104, 4020–4028. [Google Scholar] [CrossRef] [PubMed]
- Chernykh, E.R.; Leplina, O.Y.; Tikhonova, M.A.; Seledtsova, N.V.; Tyrinova, T.V.; Khonina, N.A.; Ostanin, A.A.; Pasman, N.M. Elevated levels of dehydroepiandrosterone as a potential mechanism of dendritic cell impairment during pregnancy. BMC Immunol. 2015, 16, 2. [Google Scholar] [CrossRef] [PubMed]
- Abomaray, F.M.; Al Jumah, M.A.; Kalionis, B.; AlAskar, A.S.; Al Harthy, S.; Jawdat, D.; Al Khaldi, A.; Alkushi, A.; Knawy, B.A.; Abumaree, M.H. Human Chorionic Villous Mesenchymal Stem Cells Modify the Functions of Human Dendritic Cells, and Induce an Anti-Inflammatory Phenotype in CD1+ Dendritic Cells. Stem Cell Rev. 2015, 11, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Gorvel, L.; Ben Amara, A.; Ka, M.B.; Textoris, J.; Gorvel, J.P.; Mege, J.L. Myeloid decidual dendritic cells and immunoregulation of pregnancy: Defective responsiveness to Coxiella burnetii and Brucella abortus. Front. Cell. Infect. Microbiol. 2014, 4, 179. [Google Scholar] [CrossRef] [PubMed]
- Talayev, V.Y.; Matveichev, A.V.; Lomunova, M.A.; Talayeva, M.V.; Tsaturov, M.E.; Zaichenko, I.Y.; Babaykina, O.N. The effect of human placenta cytotrophoblast cells on the maturation and T cell stimulating ability of dendritic cells in vitro. Clin. Exp. Immunol. 2010, 162, 91–99. [Google Scholar] [CrossRef]
- Salamone, G.; Fraccaroli, L.; Gori, S.; Grasso, E.; Paparini, D.; Geffner, J.; Perez Leiros, C.; Ramhorst, R. Trophoblast cells induce a tolerogenic profile in dendritic cells. Hum. Reprod. 2012, 27, 2598–2606. [Google Scholar] [CrossRef] [Green Version]
- Du, M.R.; Guo, P.F.; Piao, H.L.; Wang, S.C.; Sun, C.; Jin, L.P.; Tao, Y.; Li, Y.H.; Zhang, D.; Zhu, R.; et al. Embryonic trophoblasts induce decidual regulatory T cell differentiation and maternal-fetal tolerance through thymic stromal lymphopoietin instructing dendritic cells. J. Immunol. 2014, 192, 1502–1511. [Google Scholar] [CrossRef] [PubMed]
- Red-Horse, K.; Rivera, J.; Schanz, A.; Zhou, Y.; Winn, V.; Kapidzic, M.; Maltepe, E.; Okazaki, K.; Kochman, R.; Vo, K.C.; et al. Cytotrophoblast induction of arterial apoptosis and lymphangiogenesis in an in vivo model of human placentation. J. Clin. Investig. 2006, 116, 2643–2652. [Google Scholar] [CrossRef] [Green Version]
- Freitag, N.; Zwier, M.V.; Barrientos, G.; Tirado-Gonzalez, I.; Conrad, M.L.; Rose, M.; Scherjon, S.A.; Plosch, T.; Blois, S.M. Influence of relative NK-DC abundance on placentation and its relation to epigenetic programming in the offspring. Cell Death Dis. 2014, 5, e1392. [Google Scholar] [CrossRef] [PubMed]
- Cordeau, M.; Herblot, S.; Charrier, E.; Audibert, F.; Cordeiro, P.; Harnois, M.; Duval, M. Defects in CD54 and CD86 up-regulation by plasmacytoid dendritic cells during pregnancy. Immunol. Investig. 2012, 41, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Le Gars, M.; Kay, A.W.; Bayless, N.L.; Aziz, N.; Dekker, C.L.; Swan, G.E.; Davis, M.M.; Blish, C.A. Increased Proinflammatory Responses of Monocytes and Plasmacytoid Dendritic Cells to Influenza A Virus Infection During Pregnancy. J. Infect. Dis. 2016, 214, 1666–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escribese, M.M.; Rodriguez-Garcia, M.; Sperling, R.; Engel, S.M.; Gallart, T.; Moran, T.M. Alpha-defensins 1-3 release by dendritic cells is reduced by estrogen. Reprod. Biol. Endocrinol. RB&E 2011, 9, 118. [Google Scholar] [CrossRef]
- Kobayashi, S.D.; Malachowa, N.; DeLeo, F.R. Influence of Microbes on Neutrophil Life and Death. Front. Cell. Infect. Microbiol. 2017, 7, 159. [Google Scholar] [CrossRef] [Green Version]
- Mandala, W.L.; Gondwe, E.N.; Molyneux, M.E.; MacLennan, J.M.; MacLennan, C.A. Leukocyte counts and lymphocyte subsets in relation to pregnancy and HIV infection in Malawian women. Am. J. Reprod. Immunol. 2017, 78, e12678. [Google Scholar] [CrossRef]
- Fleming, A.F.; Akintunde, E.A.; Harrison, K.A.; Dunn, D. Leucocyte counts during pregnancy and the puerperium and at birth in Nigerians. East Afr. Med. J. 1985, 62, 175–184. [Google Scholar]
- Yang, F.; Feng, C.; Zhang, X.; Lu, J.; Zhao, Y. The diverse biological functions of neutrophils, beyond the defense against infections. Inflammation 2016, 40, 311. [Google Scholar] [CrossRef]
- Kanda, N.; Watanabe, S. 17beta-estradiol enhances the production of granulocyte-macrophage colony-stimulating factor in human keratinocytes. J. Investig. Dermatol. 2004, 123, 329–337. [Google Scholar] [CrossRef]
- Tamura, K.; Kumasaka, K.; Kogo, H. The expression of granulocyte-macrophage colony-stimulating factor (GM-CSF) and its regulation by ovarian steroids in rat uterine stromal cells. Jpn. J. Pharmacol. 1999, 79, 257–262. [Google Scholar] [CrossRef]
- Qiu, X.; Jin, X.; Shao, Z.; Zhao, X. 17beta-estradiol induces the proliferation of hematopoietic stem cells by promoting the osteogenic differentiation of mesenchymal stem cells. Tohoku J. Exp. Med. 2014, 233, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Nagata, H.; Katsuki, T.; Nakashima, C.; Onodera, R.; Hiraoka, A.; Takata, N.; Kobayashi, M.; Kambe, M. Significance of human neutrophil antigen-2a (NB1) expression and neutrophil number in pregnancy. Transfusion 2004, 44, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Felix, J.K.; Calhoun, D.A. Neonatal alloimmune neutropenia in premature monozygous twins. Pediatrics 2000, 106, 340–342. [Google Scholar] [CrossRef] [PubMed]
- Stroncek, D.F.; Skubitz, K.M.; McCullough, J.J. Biochemical characterization of the neutrophil-specific antigen NB1. Blood 1990, 75, 744–755. [Google Scholar] [PubMed]
- Giaglis, S.; Stoikou, M.; Sur Chowdhury, C.; Schaefer, G.; Grimolizzi, F.; Rossi, S.W.; Hoesli, I.M.; Lapaire, O.; Hasler, P.; Hahn, S. Multimodal Regulation of NET Formation in Pregnancy: Progesterone Antagonizes the Pro-NETotic Effect of Estrogen and G-CSF. Front. Immunol. 2016, 7, 565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, V.; Zychlinsky, A. Beneficial suicide: Why neutrophils die to make NETs. Nat. Rev. Microbiol. 2007, 5, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lopez, N.; Romero, R.; Xu, Y.; Miller, D.; Unkel, R.; Shaman, M.; Jacques, S.M.; Panaitescu, B.; Garcia-Flores, V.; Hassan, S.S. Neutrophil Extracellular Traps in the Amniotic Cavity of Women with Intra-Amniotic Infection: A New Mechanism of Host Defense. Reprod. Sci. 2016, 24, 1139–1153. [Google Scholar] [CrossRef]
- Zenclussen, A.C.; Hammerling, G.J. Cellular Regulation of the Uterine Microenvironment That Enables Embryo Implantation. Front. Immunol. 2015, 6, 321. [Google Scholar] [CrossRef]
- Jones, R.L.; Hannan, N.J.; Kaitu’u, T.J.; Zhang, J.; Salamonsen, L.A. Identification of chemokines important for leukocyte recruitment to the human endometrium at the times of embryo implantation and menstruation. J. Clin. Endocrinol. Metab. 2004, 89, 6155–6167. [Google Scholar] [CrossRef]
- Daimon, E.; Wada, Y. Role of neutrophils in matrix metalloproteinase activity in the preimplantation mouse uterus. Biol. Reprod. 2005, 73, 163–171. [Google Scholar] [CrossRef]
- Caballero-Campo, P.; Dominguez, F.; Coloma, J.; Meseguer, M.; Remohi, J.; Pellicer, A.; Simon, C. Hormonal and embryonic regulation of chemokines IL-8, MCP-1 and RANTES in the human endometrium during the window of implantation. Mol. Hum. Reprod. 2002, 8, 375–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovici, R.M.; Betzler, N.K.; Krause, M.S.; Luo, M.; Jauckus, J.; Germeyer, A.; Bloethner, S.; Schlotterer, A.; Kumar, R.; Strowitzki, T.; et al. Gene expression profiling of human endometrial-trophoblast interaction in a coculture model. Endocrinology 2006, 147, 5662–5675. [Google Scholar] [CrossRef] [PubMed]
- Givan, A.L.; White, H.D.; Stern, J.E.; Colby, E.; Gosselin, E.J.; Guyre, P.M.; Wira, C.R. Flow cytometric analysis of leukocytes in the human female reproductive tract: Comparison of fallopian tube, uterus, cervix, and vagina. Am. J. Reprod. Immunol. 1997, 38, 350–359. [Google Scholar] [CrossRef] [PubMed]
- King, A.E.; Critchley, H.O.; Kelly, R.W. Innate immune defences in the human endometrium. Reprod. Biol. Endocrinol. RB&E 2003, 1, 116. [Google Scholar] [CrossRef]
- Giaglis, S.; Stoikou, M.; Grimolizzi, F.; Subramanian, B.Y.; van Breda, S.V.; Hoesli, I.; Lapaire, O.; Hasler, P.; Than, N.G.; Hahn, S. Neutrophil migration into the placenta: Good, bad or deadly? Cell Adhes. Migr. 2016, 10, 208–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, I.; Young, A.; Ledingham, M.A.; Thomson, A.J.; Jordan, F.; Greer, I.A.; Norman, J.E. Leukocyte density and pro-inflammatory cytokine expression in human fetal membranes, decidua, cervix and myometrium before and during labour at term. Mol. Hum. Reprod. 2003, 9, 41–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kliman, H.J.; Sammar, M.; Grimpel, Y.I.; Lynch, S.K.; Milano, K.M.; Pick, E.; Bejar, J.; Arad, A.; Lee, J.J.; Meiri, H.; et al. Placental protein 13 and decidual zones of necrosis: An immunologic diversion that may be linked to preeclampsia. Reprod. Sci. 2012, 19, 16–30. [Google Scholar] [CrossRef]
- Kobara, H.; Miyamoto, T.; Suzuki, A.; Asaka, R.; Yamada, Y.; Ishikawa, K.; Kikuchi, N.; Ohira, S.; Shiozawa, T. Lipocalin2 enhances the matrix metalloproteinase-9 activity and invasion of extravillous trophoblasts under hypoxia. Placenta 2013, 34, 1036–1043. [Google Scholar] [CrossRef] [Green Version]
- Amsalem, H.; Kwan, M.; Hazan, A.; Zhang, J.; Jones, R.L.; Whittle, W.; Kingdom, J.C.; Croy, B.A.; Lye, S.J.; Dunk, C.E. Identification of a novel neutrophil population: Proangiogenic granulocytes in second-trimester human decidua. J. Immunol. 2014, 193, 3070–3079. [Google Scholar] [CrossRef]
- Odum, L.; Andersen, A.S.; Hviid, T.V. Urinary neutrophil gelatinase-associated lipocalin (NGAL) excretion increases in normal pregnancy but not in preeclampsia. Clin. Chem. Lab. Med. 2014, 52, 221–225. [Google Scholar] [CrossRef]
- Yan, L.; Borregaard, N.; Kjeldsen, L.; Moses, M.A. The high molecular weight urinary matrix metalloproteinase (MMP) activity is a complex of gelatinase B/MMP-9 and neutrophil gelatinase-associated lipocalin (NGAL). Modulation of MMP-9 activity by NGAL. J. Biol. Chem. 2001, 276, 37258–37265. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.W. Plasma from preeclamptic women stimulates transendothelial migration of neutrophils. Reprod. Sci. 2009, 16, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.; Giaglis, S.; Hoesli, I.; Hasler, P. Neutrophil NETs in reproduction: From infertility to preeclampsia and the possibility of fetal loss. Front. Immunol. 2012, 3, 362. [Google Scholar] [CrossRef] [PubMed]
- Kamath, M.S.; Chittawar, P.B.; Kirubakaran, R.; Mascarenhas, M. Use of granulocyte-colony stimulating factor in assisted reproductive technology: A systematic review and meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 214, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhang, T.; Tian, Z.; Zhang, J.; Wang, W.; Zhang, H.; Zeng, Y.; Ou, J.; Yang, Y. Efficacy of intrauterine perfusion of granulocyte colony-stimulating factor (G-CSF) for Infertile women with thin endometrium: A systematic review and meta-analysis. Am. J. Reprod. Immunol. 2017, 78, e12701. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xu, B.; Xie, S.; Zhang, Q.; Li, Y.P. Whether G-CSF administration has beneficial effect on the outcome after assisted reproductive technology? A systematic review and meta-analysis. Reprod. Biol. Endocrinol. RB&E 2016, 14, 62. [Google Scholar] [CrossRef]
- Moffett, A.; Shreeve, N. First do no harm: Uterine natural killer (NK) cells in assisted reproduction. Hum. Reprod. 2015, 30, 1519–1525. [Google Scholar] [CrossRef] [PubMed]
- Roach, J.C.; Glusman, G.; Rowen, L.; Kaur, A.; Purcell, M.K.; Smith, K.D.; Hood, L.E.; Aderem, A. The evolution of vertebrate Toll-like receptors. Proc. Natl. Acad. Sci. USA 2005, 102, 9577–9582. [Google Scholar] [CrossRef] [Green Version]
- Vidya, M.K.; Kumar, V.G.; Sejian, V.; Bagath, M.; Krishnan, G.; Bhatta, R. Toll-like receptors: Significance, ligands, signaling pathways, and functions in mammals. Int. Rev. Immunol. 2018, 37, 20–36. [Google Scholar] [CrossRef]
- Farhat, K.; Riekenberg, S.; Heine, H.; Debarry, J.; Lang, R.; Mages, J.; Buwitt-Beckmann, U.; Roschmann, K.; Jung, G.; Wiesmuller, K.H.; et al. Heterodimerization of TLR2 with TLR1 or TLR6 expands the ligand spectrum but does not lead to differential signaling. J. Leukoc. Biol. 2008, 83, 692–701. [Google Scholar] [CrossRef]
- Kim, H.M.; Park, B.S.; Kim, J.I.; Kim, S.E.; Lee, J.; Oh, S.C.; Enkhbayar, P.; Matsushima, N.; Lee, H.; Yoo, O.J.; et al. Crystal structure of the TLR4-MD-2 complex with bound endotoxin antagonist Eritoran. Cell 2007, 130, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Ohto, U.; Fukase, K.; Miyake, K.; Satow, Y. Crystal structures of human MD-2 and its complex with antiendotoxic lipid IVa. Science 2007, 316, 1632–1634. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFkappaB system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.O.; Sweet, M.J.; Mansell, A.; Kellie, S.; Kobe, B. TRIF-dependent TLR signaling, its functions in host defense and inflammation, and its potential as a therapeutic target. J. Leukoc. Biol. 2016, 100, 27–45. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Yip, T.F.; Yan, S.; Jin, D.Y.; Wei, H.L.; Guo, R.T.; Peiris, J.S.M. Recognition of Double-Stranded RNA and Regulation of Interferon Pathway by Toll-Like Receptor 10. Front. Immunol. 2018, 9, 516. [Google Scholar] [CrossRef] [PubMed]
- Oosting, M.; Cheng, S.C.; Bolscher, J.M.; Vestering-Stenger, R.; Plantinga, T.S.; Verschueren, I.C.; Arts, P.; Garritsen, A.; van Eenennaam, H.; Sturm, P.; et al. Human TLR10 is an anti-inflammatory pattern-recognition receptor. Proc. Natl. Acad. Sci. USA 2014, 111, E4478–E4484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, K.; Mor, G. Toll-like receptors at the maternal-fetal interface in normal pregnancy and pregnancy disorders. Am. J. Reprod. Immunol. 2010, 63, 587–600. [Google Scholar] [CrossRef]
- Mitsunari, M.; Yoshida, S.; Shoji, T.; Tsukihara, S.; Iwabe, T.; Harada, T.; Terakawa, N. Macrophage-activating lipopeptide-2 induces cyclooxygenase-2 and prostaglandin E(2) via toll-like receptor 2 in human placental trophoblast cells. J. Reprod. Immunol. 2006, 72, 46–59. [Google Scholar] [CrossRef]
- Abrahams, V.M.; Bole-Aldo, P.; Kim, Y.M.; Straszewski-Chavez, S.L.; Chaiworapongsa, T.; Romero, R.; Mor, G. Divergent trophoblast responses to bacterial products mediated by TLRs. J. Immunol. 2004, 173, 4286–4296. [Google Scholar] [CrossRef]
- Beijar, E.C.; Mallard, C.; Powell, T.L. Expression and subcellular localization of TLR-4 in term and first trimester human placenta. Placenta 2006, 27, 322–326. [Google Scholar] [CrossRef]
- Holmlund, U.; Cebers, G.; Dahlfors, A.R.; Sandstedt, B.; Bremme, K.; Ekstrom, E.S.; Scheynius, A. Expression and regulation of the pattern recognition receptors Toll-like receptor-2 and Toll-like receptor-4 in the human placenta. Immunology 2002, 107, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Bryant, A.H.; Menzies, G.E.; Scott, L.M.; Spencer-Harty, S.; Davies, L.B.; Smith, R.A.; Jones, R.H.; Thornton, C.A. Human gestation-associated tissues express functional cytosolic nucleic acid sensing pattern recognition receptors. Clin. Exp. Immunol. 2017, 189, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pudney, J.; He, X.; Masheeb, Z.; Kindelberger, D.W.; Kuohung, W.; Ingalls, R.R. Differential expression of toll-like receptors in the human placenta across early gestation. Placenta 2016, 46, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, N.; Nandula, P.; Menden, H.; Jarzembowski, J.; Sampath, V. Placental TLR/NLR expression signatures are altered with gestational age and inflammation. J. Matern. Fetal Neonatal Med. Off. J. Eur. Assoc. Perinat. Med. Fed. Asia Ocean. Perinat. Soc. Int. Soc. Perinat. Obs. 2017, 30, 1588–1595. [Google Scholar] [CrossRef] [PubMed]
- Gierman, L.M.; Stodle, G.S.; Tangeras, L.H.; Austdal, M.; Olsen, G.D.; Follestad, T.; Skei, B.; Rian, K.; Gundersen, A.S.; Austgulen, R.; et al. Toll-like receptor profiling of seven trophoblast cell lines warrants caution for translation to primary trophoblasts. Placenta 2015, 36, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Dulay, A.T.; Buhimschi, C.S.; Zhao, G.; Oliver, E.A.; Mbele, A.; Jing, S.; Buhimschi, I.A. Soluble TLR2 is present in human amniotic fluid and modulates the intraamniotic inflammatory response to infection. J. Immunol. 2009, 182, 7244–7253. [Google Scholar] [CrossRef] [PubMed]
- Kumazaki, K.; Nakayama, M.; Yanagihara, I.; Suehara, N.; Wada, Y. Immunohistochemical distribution of Toll-like receptor 4 in term and preterm human placentas from normal and complicated pregnancy including chorioamnionitis. Hum. Pathol. 2004, 35, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Canavan, T.P.; Simhan, H.N. Innate immune function of the human decidual cell at the maternal-fetal interface. J. Reprod. Immunol. 2007, 74, 46–52. [Google Scholar] [CrossRef]
- Krikun, G.; Lockwood, C.J.; Abrahams, V.M.; Mor, G.; Paidas, M.; Guller, S. Expression of Toll-like receptors in the human decidua. Histol. Histopathol. 2007, 22, 847–854. [Google Scholar] [CrossRef]
- Hayati, A.R.; Mohamed, A.E.; Tan, G.C. An immunohistochemical study of Toll-like receptors 2 and 4 in placenta with and without infection. Malays. J. Pathol. 2010, 32, 13–19. [Google Scholar]
- Hoang, M.; Potter, J.A.; Gysler, S.M.; Han, C.S.; Guller, S.; Norwitz, E.R.; Abrahams, V.M. Human fetal membranes generate distinct cytokine profiles in response to bacterial Toll-like receptor and nod-like receptor agonists. Biol. Reprod. 2014, 90, 39. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, V.M.; Potter, J.A.; Bhat, G.; Peltier, M.R.; Saade, G.; Menon, R. Bacterial modulation of human fetal membrane Toll-like receptor expression. Am. J. Reprod. Immunol. 2013, 69, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Romero, R.; Chaiworapongsa, T.; Kim, G.J.; Kim, M.R.; Kuivaniemi, H.; Tromp, G.; Espinoza, J.; Bujold, E.; Abrahams, V.M.; et al. Toll-like receptor-2 and -4 in the chorioamniotic membranes in spontaneous labor at term and in preterm parturition that are associated with chorioamnionitis. Am. J. Obstet. Gynecol. 2004, 191, 1346–1355. [Google Scholar] [CrossRef] [PubMed]
- Flores-Espinosa, P.; Pineda-Torres, M.; Vega-Sanchez, R.; Estrada-Gutierrez, G.; Espejel-Nunez, A.; Flores-Pliego, A.; Maida-Claros, R.; Paredes-Vivas, Y.; Morales-Mendez, I.; Sosa-Gonzalez, I.; et al. Progesterone elicits an inhibitory effect upon LPS-induced innate immune response in pre-labor human amniotic epithelium. Am. J. Reprod. Immunol. 2014, 71, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Tchirikov, M.; Schlabritz-Loutsevitch, N.; Maher, J.; Buchmann, J.; Naberezhnev, Y.; Winarno, A.S.; Seliger, G. Mid-trimester preterm premature rupture of membranes (PPROM): Etiology, diagnosis, classification, international recommendations of treatment options and outcome. J. Perinat. Med. 2018, 46, 465–488. [Google Scholar] [CrossRef]
- Zaga-Clavellina, V.; Flores-Espinosa, P.; Pineda-Torres, M.; Sosa-Gonzalez, I.; Vega-Sanchez, R.; Estrada-Gutierrez, G.; Espejel-Nunez, A.; Flores-Pliego, A.; Maida-Claros, R.; Estrada-Juarez, H.; et al. Tissue-specific IL-10 secretion profile from term human fetal membranes stimulated with pathogenic microorganisms associated with preterm labor in a two-compartment tissue culture system. J. Matern. Fetal Neonatal Med. Off. J. Eur. Assoc. Perinat. Med. Fed. Asia Ocean. Perinat. Soc. Int. Soc. Perinat. Obs. 2014, 27, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Zaga-Clavellina, V.; Martha, R.V.; Flores-Espinosa, P. In vitro secretion profile of pro-inflammatory cytokines IL-1beta, TNF-alpha, IL-6, and of human beta-defensins (HBD)-1, HBD-2, and HBD-3 from human chorioamniotic membranes after selective stimulation with Gardnerella vaginalis. Am. J. Reprod. Immunol. 2012, 67, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Flores-Espinosa, P.; Preciado-Martinez, E.; Mejia-Salvador, A.; Sedano-Gonzalez, G.; Bermejo-Martinez, L.; Parra-Covarruvias, A.; Estrada-Gutierrez, G.; Vega-Sanchez, R.; Mendez, I.; Quesada-Reyna, B.; et al. Selective immuno-modulatory effect of prolactin upon pro-inflammatory response in human fetal membranes. J. Reprod. Immunol. 2017, 123, 58–64. [Google Scholar] [CrossRef]
- Pineda-Torres, M.; Flores-Espinosa, P.; Espejel-Nunez, A.; Estrada-Gutierrez, G.; Flores-Pliego, A.; Maida-Claros, R.; Zaga-Clavellina, V. Evidence of an immunosuppressive effect of progesterone upon in vitro secretion of proinflammatory and prodegradative factors in a model of choriodecidual infection. BJOG 2015, 122, 1798–1807. [Google Scholar] [CrossRef]
- Dudley, D.J.; Edwin, S.S.; Dangerfield, A.; Jackson, K.; Trautman, M.S. Regulation of decidual cell and chorion cell production of interleukin-10 by purified bacterial products. Am. J. Reprod. Immunol. 1997, 38, 246–251. [Google Scholar] [CrossRef]
- Bayraktar, M.; Peltier, M.; Vetrano, A.; Arita, Y.; Gurzenda, E.; Joseph, A.; Kazzaz, J.; Sharma, S.; Hanna, N. IL-10 modulates placental responses to TLR ligands. Am. J. Reprod. Immunol. 2009, 62, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, V.M.; Visintin, I.; Aldo, P.B.; Guller, S.; Romero, R.; Mor, G. A role for TLRs in the regulation of immune cell migration by first trimester trophoblast cells. J. Immunol. 2005, 175, 8096–8104. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, G.; Flores-Espinosa, P.; Preciado-Martinez, E.; Bermejo-Martinez, L.; Espejel-Nunez, A.; Estrada-Gutierrez, G.; Maida-Claros, R.; Flores-Pliego, A.; Zaga-Clavellina, V. In vitro progesterone modulation on bacterial endotoxin-induced production of IL-1beta, TNFalpha, IL-6, IL-8, IL-10, MIP-1alpha, and MMP-9 in pre-labor human term placenta. Reprod. Biol. Endocrinol. RB&E 2015, 13, 115. [Google Scholar] [CrossRef]
- Zaga-Clavellina, V.; Garcia-Lopez, G.; Flores-Espinosa, P. Evidence of in vitro differential secretion of human beta-defensins-1, -2, and -3 after selective exposure to Streptococcus agalactiae in human fetal membranes. J. Matern. Fetal Neonatal Med. Off. J. Eur. Assoc. Perinat. Med. Fed. Asia Ocean. Perinat. Soc. Int. Soc. Perinat. Obs. 2012, 25, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lopez, G.; Flores-Espinosa, P.; Zaga-Clavellina, V. Tissue-specific human beta-defensins (HBD)1, HBD2, and HBD3 secretion from human extra-placental membranes stimulated with Escherichia coli. Reprod. Biol. Endocrinol. RB&E 2010, 8, 146. [Google Scholar] [CrossRef]
- Dudley, D.J.; Edwin, S.S.; Van Wagoner, J.; Augustine, N.H.; Hill, H.R.; Mitchell, M.D. Regulation of decidual cell chemokine production by group B streptococci and purified bacterial cell wall components. Am. J. Obstet. Gynecol. 1997, 177, 666–672. [Google Scholar] [CrossRef]
- Preciado-Martinez, E.; Garcia-Ruiz, G.; Flores-Espinosa, P.; Bermejo-Martinez, L.; Espejel-Nunez, A.; Estrada-Gutierrez, G.; Razo-Aguilera, G.; Granados-Cepeda, M.; Helguera-Repetto, A.C.; Irles, C.; et al. Progesterone suppresses the lipopolysaccharide-induced pro-inflammatory response in primary mononuclear cells isolated from human placental blood. Immunol. Investig. 2018, 47, 181–195. [Google Scholar] [CrossRef]
- Liu, H.Y.; Hong, Y.F.; Huang, C.M.; Chen, C.Y.; Huang, T.N.; Hsueh, Y.P. TLR7 negatively regulates dendrite outgrowth through the Myd88-c-Fos-IL-6 pathway. J. Neurosci. 2013, 33, 11479–11493. [Google Scholar] [CrossRef]
- Kato, E.; Yamamoto, T.; Chishima, F. Effects of Cytokines and TLR Ligands on the Production of PlGF and sVEGFR1 in Primary Trophoblasts. Gynecol. Obstet. Investig. 2017, 82, 39–46. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Main Ligands | TLR Expression | References | |||
|---|---|---|---|---|---|
| 1st Trimester | 2nd Trimester | 3rd Trimester | |||
| TLR1 | Triacylated lipopeptides and lipoproteins from Gram-positive bacteria. When forming a heterodimer with TLR2, it recognizes peptidoglycan and triacylated lipoproteins | ++ | ++ | - | [149,150,154] |
| TLR2 | Lipoteichoic acid and lipoproteins from Gram-positive bacteria, lipoarabinomannans from mycobacteria and zymosan from yeast. Diacylated or triacylated lipopeptides depending on heterodimerization patterns | ++ | ++ | +++ | [150,152] |
| TLR3 | Double strand RNA | ++ | ++ | +++ | [153,154] |
| TLR4 | Lipopolysaccharides from Gram-negative bacteria, paclitaxel, heat shock proteins, heparan sulphate, reactive oxygen species (ROS), fibrinogen and fibronectin | ++ | +++ | +++ | [150,151,152] |
| TLR5 | Flagellin from both Gram-positive and Gram-negative bacteria | ++ | ++ | ++ | [154,155] |
| TLR6 | Diacylated lipopeptides from mycoplasmas (also heterodimerizes with TLR2) | -/+ | + | ++ | [149,150,154] |
| TLR7 | Single strand RNA and small synthetic compounds such as guanosine analogs or imidazoquinoline | + | ++ | + | [153,154] |
| TLR8 | Single strand RNA and small synthetic compounds such as guanosine analogs or imidazoquinoline | + | ++ | +++ | [153,154] |
| TLR9 | Viral single-strand unmethylated CpG DNA and also fetal DNA | - | + | + | [153,154,155] |
| TLR10 | Double strand RNA | + | + | + | [154] |
| Cultured Term Placenta | ||||||||||||||
| Cytokines | Chemokines | Antimicrobial Peptides | ||||||||||||
| Agonist of: | IL-6 | IL-1β | TNF-α | IL-10 | IL-2 | IL-8 | MCP-1 | GRO-α | RANTES | MIP-1α | HBD1 | HBD2 | HBD3 | Reference |
| TLR2 | ||||||||||||||
| Lipoteichoic acid | ++ | ++ | + | ++ | + | +++ | [172] | |||||||
| TLR3 | ||||||||||||||
| High MW Poly I:C | + | + | [153] | |||||||||||
| Low MW Poly I:C | ++ | ++ | + | + | + | [153] | ||||||||
| TLR4 | ||||||||||||||
| LPS from E. coli | + | + | + | [173] | ||||||||||
| LPS from E. coli | + | +++ | ++ | + | ++ | ++ | [174] | |||||||
| LPS from E. coli | UC | ++ | UC | [6] | ||||||||||
| LPS from E. coli | ++ | +++ | +++ | ++ | + | +++ | [172] | |||||||
| TLR7 | ||||||||||||||
| Imiquimod | UC | + | [153] | |||||||||||
| TLR8 | ||||||||||||||
| ssRNA40 | ++ | ++ | [153] | |||||||||||
| TLR9 | ||||||||||||||
| ODN21798 | UC | UC | [153] | |||||||||||
| Cultured Term Chorion | ||||||||||||||
| Cytokines | Chemokines | Antimicrobial Peptides | ||||||||||||
| Agonist of: | IL-6 | IL-1β | TNF-α | IL-10 | IL-2 | IL-8 | MCP-1 | GRO-α | RANTES | MIP-1α | HBD1 | HBD2 | HBD3 | Reference |
| TLR2 | ||||||||||||||
| Streptococcus agalactiae | + | [167] | ||||||||||||
| Streptococcus agalactiae | UC | + | UC | [175] | ||||||||||
| Lipoteichoic acid from Streptococcus pyogenes | + | [171] | ||||||||||||
| Candida albicans | +++ | [167] | ||||||||||||
| TLR3 | ||||||||||||||
| High MW Poly I:C | + | + | [153] | |||||||||||
| Low MW Poly I:C | ++ | ++ | [153] | |||||||||||
| TLR4 | ||||||||||||||
| LPS from E. coli | UC | ++ | +++ | + | [169] | |||||||||
| LPS from E. coli | ++ | +++ | +++ | + | + | [170] | ||||||||
| LPS from E. coli | ++ | [171] | ||||||||||||
| Escherichia coli | UC | + | UC | [176] | ||||||||||
| Gardnerella vaginalis | +++ | [167] | ||||||||||||
| Gardnerella vaginalis | ++ | ++ | UC | UC | + | UC | [168] | |||||||
| TLR7 | ||||||||||||||
| Imiquimod | UC | + | [153] | |||||||||||
| TLR8 | ||||||||||||||
| ssRNA40 | + | ++ | [153] | |||||||||||
| TLR9 | ||||||||||||||
| ODN21798 | UC | UC | [153] | |||||||||||
| Cultured Term Amnion | ||||||||||||||
| Cytokines | Chemokines | Antimicrobial Peptides | ||||||||||||
| Agonist of: | IL-6 | IL-1β | TNF-α | IL-10 | IL-2 | IL-8 | MCP-1 | GRO-α | RANTES | MIP-1α | HBD1 | HBD2 | HBD3 | Reference |
| TLR2 | ||||||||||||||
| Peptidoglycans from Staphylococcus aureus | ++ | ++ | ++ | UC | ++ | UC | UC | +++ | + | [162] | ||||
| TLR4 | ||||||||||||||
| LPS from E. coli | UC | +++ | +++ | +++ | UC | + | UC | ++ | +++ | UC | [162] | |||
| TLR5 | ||||||||||||||
| Flagellin from Salmonella typhimurium | +++ | +++ | +++ | UC | UC | +++ | UC | + | +++ | + | [162] | |||
| TLR9 | ||||||||||||||
| ODN21798 | UC | UC | UC | UC | UC | UC | + | UC | - | - | [162] | |||
| Cultured Term Chorioamniotic Membranes | ||||||||||||||
| Cytokines | Chemokines | Antimicrobial Peptides | ||||||||||||
| Agonist of: | IL-6 | IL-1β | TNF-α | IL-10 | IL-2 | IL-8 | MCP-1 | GRO-α | RANTES | MIP-1α | HBD1 | HBD2 | HBD3 | Reference |
| TLR2 | ||||||||||||||
| Peptidoglycans from Staphylococcus aureus | ++ | ++ | ++ | UC | ++ | UC | UC | +++ | +++ | [162] | ||||
| TLR4 | ||||||||||||||
| LPS from E. coli | UC | +++ | +++ | +++ | UC | + | UC | ++ | +++ | UC | [162] | |||
| TLR5 | ||||||||||||||
| Flagellin | +++ | +++ | +++ | UC | UC | +++ | UC | + | +++ | +++ | [162] | |||
| TLR9 | ||||||||||||||
| ODN21798 | UC | UC | UC | UC | UC | UC | + | UC | - | - | [162] | |||
| Cultured Term Decidual Cells | ||||||||||||||
| Cytokines | Chemokines | Antimicrobial Peptides | ||||||||||||
| Agonist of: | IL-6 | IL-1β | TNF-α | IL-10 | IL-2 | IL-8 | MCP-1 | GRO-α | RANTES | MIP-1α | HBD1 | HBD2 | HBD3 | Reference |
| TLR2 | ||||||||||||||
| Lipoteichoic acid from Streptococcus pyogenes | ++ | ++ | [177] | |||||||||||
| Lipoteichoic acid from Streptococcus pyogenes | +++ | [171] | ||||||||||||
| Group B Streptococci | ++ | ++ | [177] | |||||||||||
| TLR4 | ||||||||||||||
| LPS from E. coli | +++ | +++ | [177] | |||||||||||
| LPS from E. coli | +++ | [171] | ||||||||||||
| Cultured Term Placental Blood Mononuclear Cells | ||||||||||||||
| Cytokines | Chemokines | Antimicrobial Peptides | ||||||||||||
| Agonist of: | IL-6 | IL-1β | TNF-α | IL-10 | IL-2 | IL-8 | MCP-1 | GRO-α | RANTES | MIP-1α | HBD1 | HBD2 | HBD3 | Reference |
| TLR4 | ||||||||||||||
| LPS from E. coli | ++ | ++ | UC | ++ | ++ | [178] | ||||||||
| Cultured Term Hofbauer Cells | ||||||||||||||
| Cytokines | Chemokines | Antimicrobial Peptides | ||||||||||||
| Agonist of: | IL-6 | IL-1β | TNF-α | IL-10 | IL-2 | IL-8 | MCP-1 | GRO-α | RANTES | MIP-1α | HBD1 | HBD2 | HBD3 | Reference |
| TLR2 | ||||||||||||||
| Peptidoglycans from Staphylococcus aureus | UC | UC | [57] | |||||||||||
| TLR3 | ||||||||||||||
| Poly I:C | ++ | + | [57] | |||||||||||
| TLR4 | ||||||||||||||
| LPS from E. coli | +++ | + | [57] | |||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olmos-Ortiz, A.; Flores-Espinosa, P.; Mancilla-Herrera, I.; Vega-Sánchez, R.; Díaz, L.; Zaga-Clavellina, V. Innate Immune Cells and Toll-like Receptor–Dependent Responses at the Maternal–Fetal Interface. Int. J. Mol. Sci. 2019, 20, 3654. https://doi.org/10.3390/ijms20153654
Olmos-Ortiz A, Flores-Espinosa P, Mancilla-Herrera I, Vega-Sánchez R, Díaz L, Zaga-Clavellina V. Innate Immune Cells and Toll-like Receptor–Dependent Responses at the Maternal–Fetal Interface. International Journal of Molecular Sciences. 2019; 20(15):3654. https://doi.org/10.3390/ijms20153654
Chicago/Turabian StyleOlmos-Ortiz, Andrea, Pilar Flores-Espinosa, Ismael Mancilla-Herrera, Rodrigo Vega-Sánchez, Lorenza Díaz, and Verónica Zaga-Clavellina. 2019. "Innate Immune Cells and Toll-like Receptor–Dependent Responses at the Maternal–Fetal Interface" International Journal of Molecular Sciences 20, no. 15: 3654. https://doi.org/10.3390/ijms20153654