Dietary Cholesterol Is Highly Associated with Severity of Hyperlipidemia and Atherosclerotic Lesions in Heterozygous LDLR-Deficient Hamsters

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
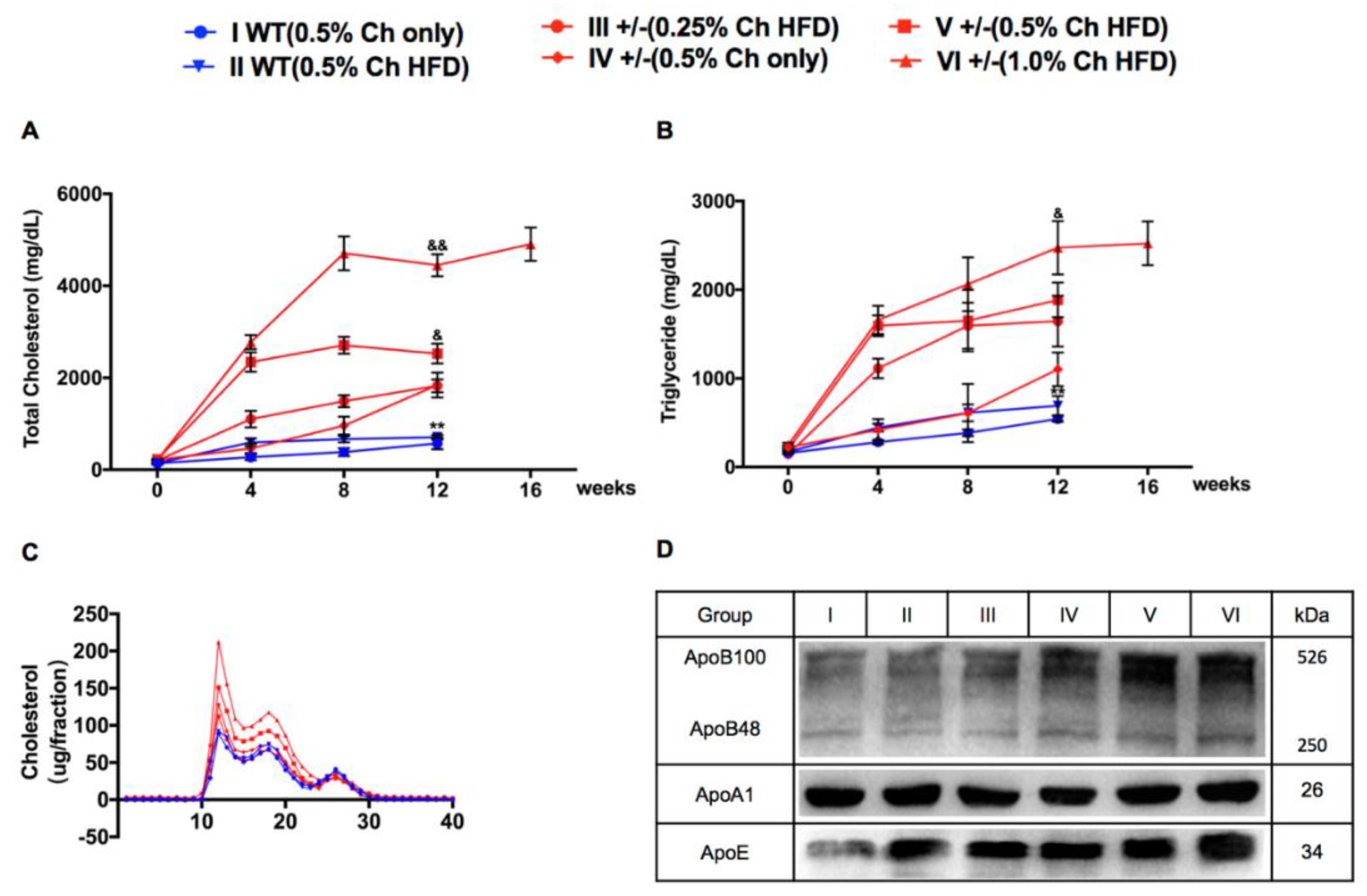
2.1. Characteristics of Plasma Lipids, Apolipoproteins, and Lipoproteins in Wild Type (WT) and LDLR+/− Hamsters
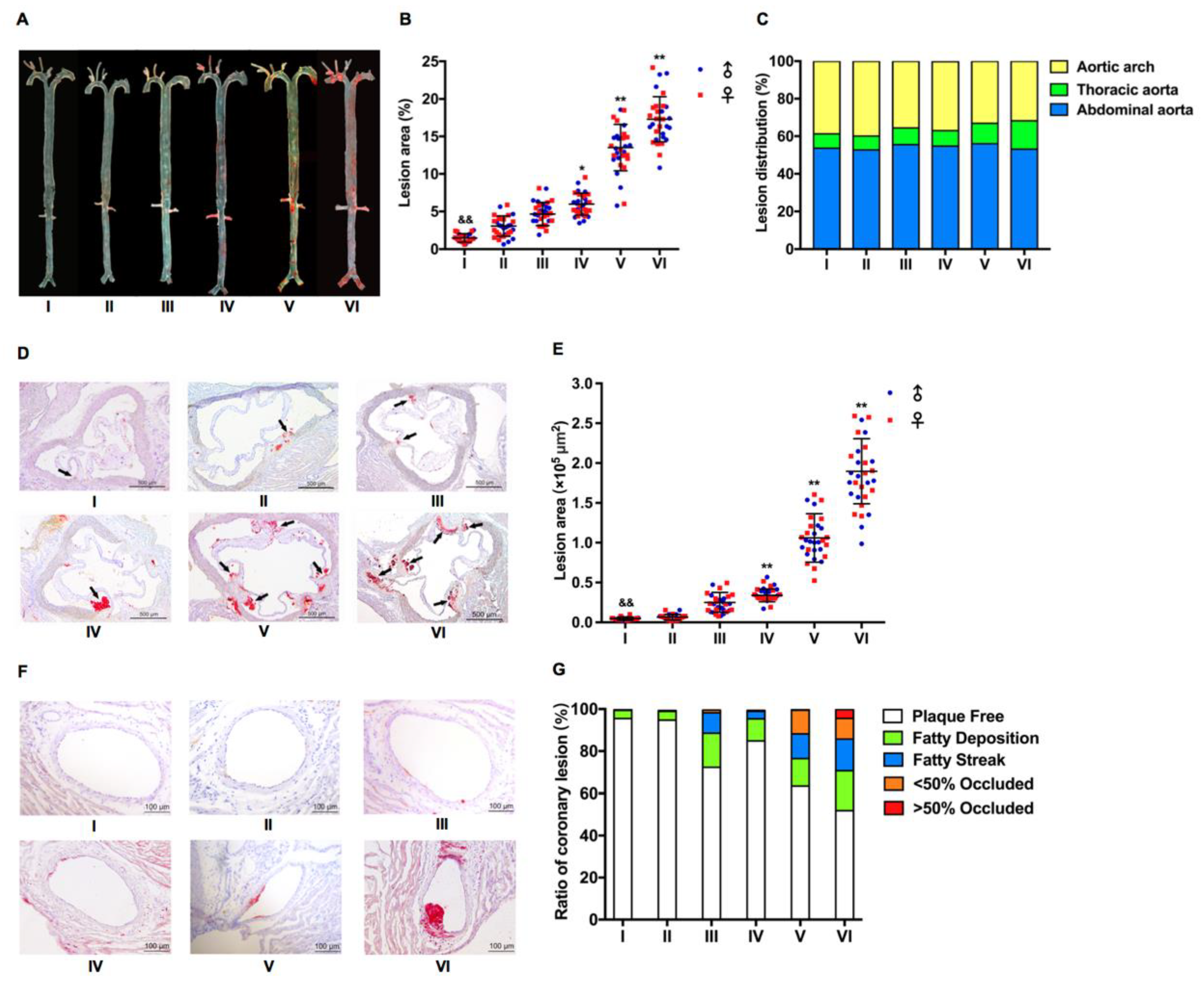
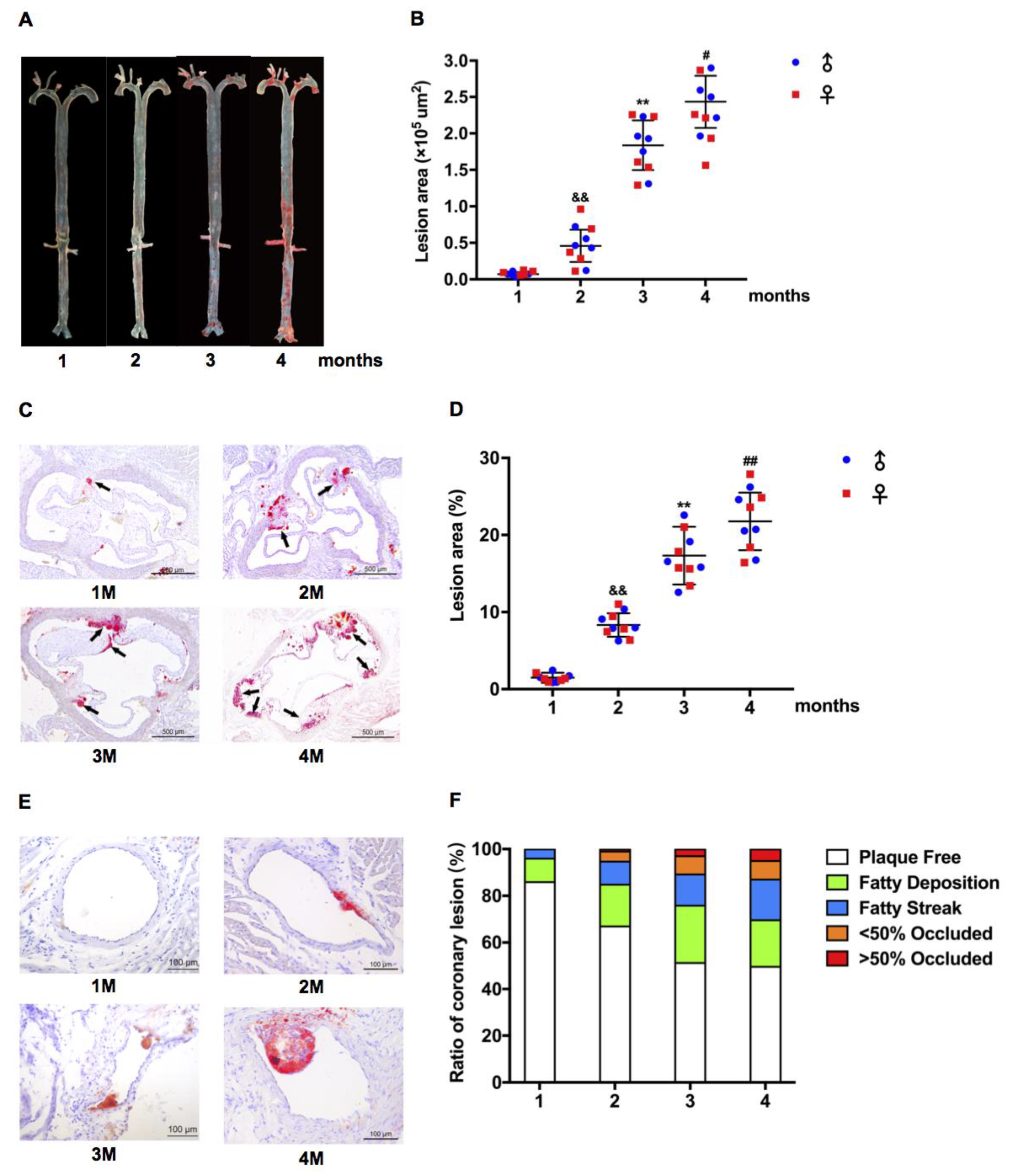
2.2. Atherosclerotic Development in LDLR+/− Hamsters Was Cholesterol Concentration and Time Course-Dependent
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Analysis of Plasma Lipids and (apo)Lipoproteins
4.3. Pathological Analysis
4.4. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hobbs, H.H.; Brown, M.S.; Goldstein, J.L. Molecular genetics of the ldl receptor gene in familial hypercholesterolemia. Hum. Mutat. 1992, 1, 445–466. [Google Scholar] [CrossRef]
- Cuchel, M.; Bruckert, E.; Ginsberg, H.N.; Raal, F.J.; Santos, R.D.; Hegele, R.A.; Kuivenhoven, J.A.; Nordestgaard, B.G.; Descamps, O.S.; Steinhagen-Thiessen, E.; et al. Homozygous familial hypercholesterolaemia: New insights and guidance for clinicians to improve detection and clinical management. A position paper from the consensus panel on familial hypercholesterolaemia of the european atherosclerosis society. Eur. Heart J. 2014, 35, 2146–2157. [Google Scholar] [CrossRef]
- Davis, B.T.; Wang, X.J.; Rohret, J.A.; Struzynski, J.T.; Merricks, E.P.; Bellinger, D.A.; Rohret, F.A.; Nichols, T.C.; Rogers, C.S. Targeted disruption of ldlr causes hypercholesterolemia and atherosclerosis in yucatan miniature pigs. PLoS ONE 2014, 9, e93457. [Google Scholar] [CrossRef]
- Johnston, J.M.; Francis, S.E.; Kiss-Toth, E. Experimental models of murine atherosclerosis: Does perception match reality? Cardiovasc. Res. 2018, 114, 1845–1847. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Gao, M.; Wang, Y.; Lin, X.; Yang, L.; Cong, N.; An, X.; Wang, F.; Qu, K.; Yu, L.; et al. Ldl receptor gene-ablated hamsters: A rodent model of familial hypercholesterolemia with dominant inheritance and diet-induced coronary atherosclerosis. Ebiomedicine 2018, 27, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Wold, W.S.; Tollefson, A.E.; Ying, B.; Spencer, J.F.; Toth, K. Drug development against human adenoviruses and its advancement by syrian hamster models. FEMS Microbiol. Rev. 2019, 43, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Morton, R.E.; Izem, L. Cholesteryl ester transfer proteins from different species do not have equivalent activities. J. Lipid Res. 2014, 55, 258–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feingold, K.R.; Grunfeld, C. Introduction to lipids and lipoproteins. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., Kopp, P., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Ishibashi, S.; Brown, M.S.; Goldstein, J.L.; Gerard, R.D.; Hammer, R.E.; Herz, J. Hypercholesterolemia in low density lipoprotein receptor knockout mice and its reversal by adenovirus-mediated gene delivery. J. Clin. Investig. 1993, 92, 883–893. [Google Scholar] [CrossRef] [PubMed]
- von Scheidt, M.; Zhao, Y.; Kurt, Z.; Pan, C.; Zeng, L.; Yang, X.; Schunkert, H.; Lusis, A.J. Applications and limitations of mouse models for understanding human atherosclerosis. Cell Metab. 2017, 25, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Daugherty, A.; Tall, A.R.; Daemen, M.; Falk, E.; Fisher, E.A.; Garcia-Cardena, G.; Lusis, A.J.; Owens, A.P. 3rd; Rosenfeld, M.E.; Virmani, R.; et al. Recommendation on design, execution, and reporting of animal atherosclerosis studies: A scientific statement from the american heart association. Arterioscler. Thromb. Vasc. Biol. 2017, 37, e131–e157. [Google Scholar] [CrossRef] [PubMed]
- Dillard, A.; Matthan, N.R.; Lichtenstein, A.H. Use of hamster as a model to study diet-induced atherosclerosis. Nutr. Metab. 2010, 7, 89. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; He, K.; Yang, C.; Lin, X.; Zhang, X.; Wang, Y.; Liu, G.; Xian, X. Dietary Cholesterol Is Highly Associated with Severity of Hyperlipidemia and Atherosclerotic Lesions in Heterozygous LDLR-Deficient Hamsters. Int. J. Mol. Sci. 2019, 20, 3515. https://doi.org/10.3390/ijms20143515
Wang J, He K, Yang C, Lin X, Zhang X, Wang Y, Liu G, Xian X. Dietary Cholesterol Is Highly Associated with Severity of Hyperlipidemia and Atherosclerotic Lesions in Heterozygous LDLR-Deficient Hamsters. International Journal of Molecular Sciences. 2019; 20(14):3515. https://doi.org/10.3390/ijms20143515
Chicago/Turabian StyleWang, Jinjie, Kunxiang He, Chun Yang, Xiao Lin, Xin Zhang, Yuhui Wang, George Liu, and Xunde Xian. 2019. "Dietary Cholesterol Is Highly Associated with Severity of Hyperlipidemia and Atherosclerotic Lesions in Heterozygous LDLR-Deficient Hamsters" International Journal of Molecular Sciences 20, no. 14: 3515. https://doi.org/10.3390/ijms20143515