Thymoquinone-Induced Tristetraprolin Inhibits Tumor Growth and Metastasis through Destabilization of MUC4 mRNA

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
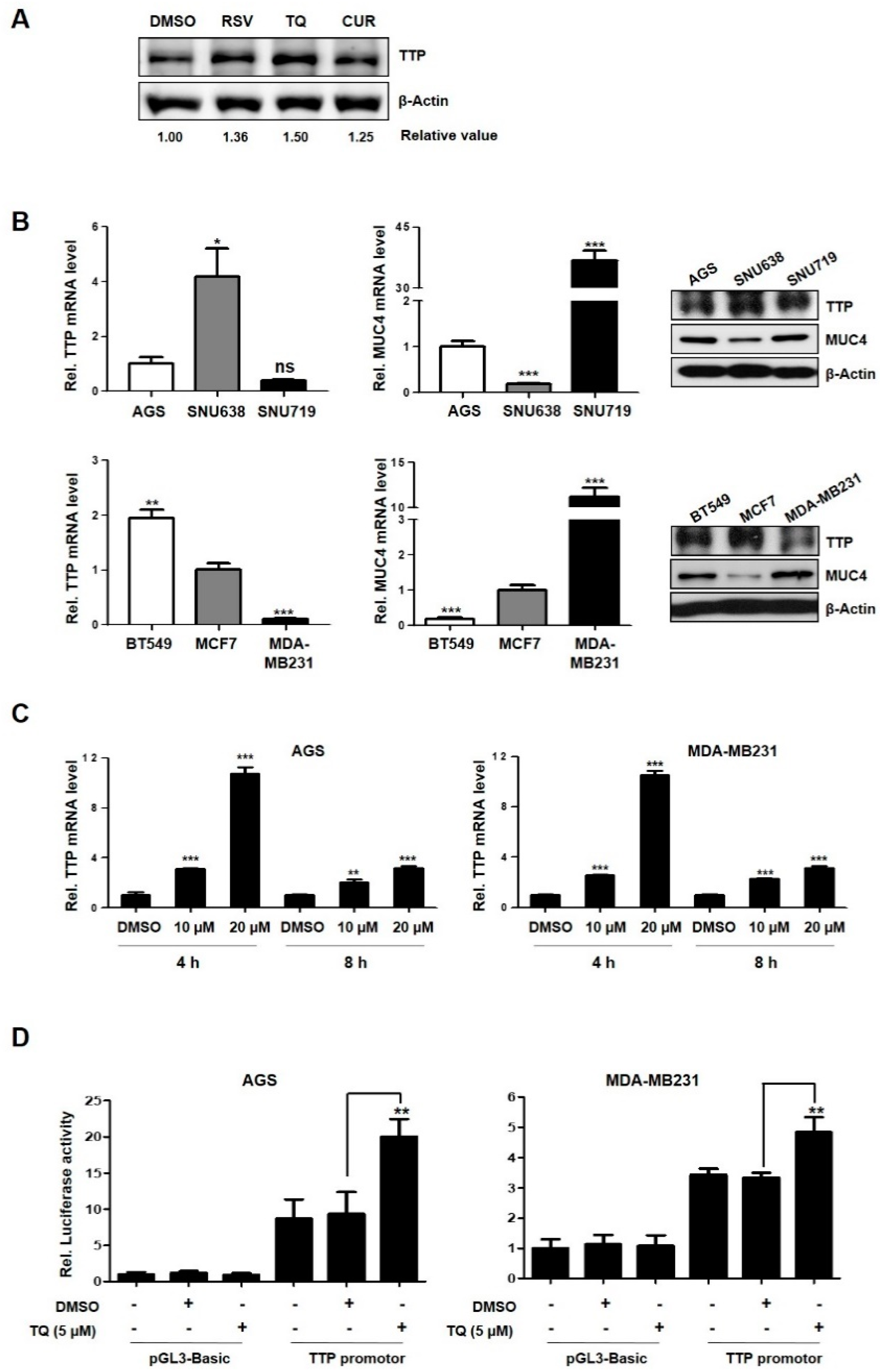
2.1. TQ-Induced TTP Regulates MUC4 Expression in Cancer Cells
2.2. MUC4 mRNA was Identified as a Novel Target for TTP
2.3. TQ-Induced TTP Expression Inhibits Tumor Progression
2.4. TQ-Induced TTP Expression Suppresses Metastasis In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Reagents
4.2. Small Interference RNA (siRNA)
4.3. MTT and Clonogenic Assay
4.4. Plasmid Construction and Luciferase Assay
4.5. Western Blot Analysis
4.6. Quantitative Real-Time (qRT) PCR
4.7. Wound Healing and Invasion Assay
4.8. RNA Electrophoretic Mobility Shift Assay (RNA-EMSA)
4.9. Mouse In Vivo Experiment
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kedar, V.P.; Zucconi, B.E.; Wilson, G.M.; Blackshear, P.J. Direct Binding of Specific AUF1 Isoforms to Tandem Zinc Finger Domains of Tristetraprolin (TTP) Family Proteins. J. Biol. Chem. 2012, 287, 5459–5471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.W.; Vo, M.T.; Kim, H.K.; Lee, H.H.; Yoon, N.A.; Lee, B.J.; Min, Y.J.; Joo, W.D.; Cha, H.J.; Park, J.W.; et al. Ectopic over-expression of tristetraprolin in human cancer cells promotes biogenesis of let-7 by down-regulation of Lin28. Nucleic Acids Res. 2012, 40, 3856–3869. [Google Scholar] [CrossRef]
- Qiu, L.Q.; Lai, W.S.; Bradbury, A.; Zeldin, D.C.; Blackshear, P.J. Tristetraprolin (TTP) coordinately regulates primary and secondary cellular responses to proinflammatory stimuli. J. Leukocyte Biol. 2015, 97, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, E.A.; Smallie, T.; Ding, Q.; O’Neil, J.D.; Cunliffe, H.E.; Tang, T.; Rosner, D.R.; Klevernic, I.; Morrice, N.A.; Monaco, C.; et al. Dominant Suppression of Inflammation via Targeted Mutation of the mRNA Destabilizing Protein Tristetraprolin. J. Immunol. 2015, 195, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Kim, H.J.; Yoon, N.A.; Lee, W.H.; Min, Y.J.; Ko, B.K.; Lee, B.J.; Lee, A.; Cha, H.J.; Cho, W.J.; et al. Tumor suppressor p53 plays a key role in induction of both tristetraprolin and let-7 in human cancer cells. Nucleic Acids Res. 2013, 41, 5614–5625. [Google Scholar] [CrossRef] [Green Version]
- Mahat, D.B.; Brennan-Laun, S.E.; Fialcowitz-White, E.J.; Kishor, A.; Ross, C.R.; Pozharskaya, T.; Rawn, J.D.; Blackshear, P.J.; Hassel, B.A.; Wilson, G.M. Coordinated expression of tristetraprolin post-transcriptionally attenuates mitogenic induction of the oncogenic Ser/Thr kinase Pim-1. PloS ONE 2012, 7, e33194. [Google Scholar] [CrossRef] [PubMed]
- Milke, L.; Schulz, K.; Weigert, A.; Sha, W.; Schmid, T.; Brune, B. Depletion of tristetraprolin in breast cancer cells increases interleukin-16 expression and promotes tumor infiltration with monocytes/macrophages. Carcinogenesis 2013, 34, 850–857. [Google Scholar] [CrossRef]
- Liu, F.; Guo, J.; Zhang, Q.; Liu, D.; Wen, L.; Yang, Y.; Yang, L.; Liu, Z. The Expression of Tristetraprolin and Its Relationship with Urinary Proteins in Patients with Diabetic Nephropathy. PloS ONE 2015, 10, e0141471. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Yang, S.S.; Vo, M.T.; Cho, W.J.; Lee, B.J.; Leem, S.H.; Lee, S.H.; Cha, H.J.; Park, J.W. Tristetraprolin down-regulates IL-23 expression in colon cancer cells. Mol. Cells 2013, 36, 571–576. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.H.; Yoon, N.A.; Vo, M.T.; Kim, C.W.; Woo, J.M.; Cha, H.J.; Cho, Y.W.; Lee, B.J.; Cho, W.J.; Park, J.W. Tristetraprolin down-regulates IL-17 through mRNA destabilization. FEBS Lett. 2012, 586, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Yoon, N.A.; Seong, H.; Jeong, J.Y.; Kang, S.; Park, N.; Choi, J.; Lee, D.H.; Roh, G.S.; Kim, H.J.; et al. Resveratrol Induces Glioma Cell Apoptosis through Activation of Tristetraprolin. Mol. Cells 2015, 38, 991–997. [Google Scholar] [PubMed] [Green Version]
- Jafri, S.H.; Glass, J.; Shi, R.H.; Zhang, S.L.; Prince, M.; Kleiner-Hancock, H. Thymoquinone and cisplatin as a therapeutic combination in lung cancer: In vitro and in vivo. J. Exp. Clin. Canc. Res. 2010, 29, 87. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, A.G.M.; Hossain, M.K.; Basak, D.; Bin Sayeed, M.S. Thymoquinone as a Potential Adjuvant Therapy for Cancer Treatment: Evidence from Preclinical Studies. Front. Pharmacol. 2017, 8, 295. [Google Scholar] [CrossRef]
- Shakibaei, M.; Mobasheri, A.; Lueders, C.; Busch, F.; Shayan, P.; Goel, A. Curcumin enhances the effect of chemotherapy against colorectal cancer cells by inhibition of NF-kappaB and Src protein kinase signaling pathways. PloS ONE 2013, 8, e57218. [Google Scholar] [CrossRef]
- Gupta, S.C.; Kannappan, R.; Reuter, S.; Kim, J.H.; Aggarwal, B.B. Chemosensitization of tumors by resveratrol. Ann. New York Acad. Sci. 2011, 1215, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Ke, X.Q.; Zhao, Y.; Lu, X.L.; Wang, Z.; Liu, Y.Y.; Ren, M.D.; Lu, G.F.; Zhang, D.; Sun, Z.G.; Xu, Z.P.; et al. TQ inhibits hepatocellular carcinoma growth in vitro and in vivo via repression of Notch signaling. Oncotarget 2015, 6, 32610–32621. [Google Scholar] [CrossRef]
- Arafa el, S.A.; Zhu, Q.; Shah, Z.I.; Wani, G.; Barakat, B.M.; Racoma, I.; El-Mahdy, M.A.; Wani, A.A. Thymoquinone up-regulates PTEN expression and induces apoptosis in doxorubicin-resistant human breast cancer cells. Mutat. Res. 2011, 706, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attoub, S.; Sperandio, O.; Raza, H.; Arafat, K.; Al-Salam, S.; Al Sultan, M.A.; Al Safi, M.; Takahashi, T.; Adem, A. Thymoquinone as an anticancer agent: Evidence from inhibition of cancer cells viability and invasion in vitro and tumor growth in vivo. Fundam. Clin. Pharmacol. 2013, 27, 557–569. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Bhardwaj, A.; Aggarwal, R.S.; Seeram, N.P.; Shishodia, S.; Takada, Y. Role of resveratrol in prevention and therapy of cancer: Preclinical and clinical studies. Anticancer. Res. 2004, 24, 2783–2840. [Google Scholar]
- Lu, R.Q.; Serrero, G. Resveratrol, a natural product derived from grape, exhibits antiestrogenic activity and inhibits the growth of human breast cancer cells. J. Cell. Physiol. 1999, 179, 297–304. [Google Scholar] [CrossRef]
- Bai, Y.; Mao, Q.Q.; Qin, J.; Zheng, X.Y.; Wang, Y.B.; Yang, K.; Shen, H.F.; Xie, L.P. Resveratrol induces apoptosis and cell cycle arrest of human T24 bladder cancer cells in vitro and inhibits tumor growth in vivo. Cancer Sci. 2010, 101, 488–493. [Google Scholar] [CrossRef]
- Vanamala, J.; Reddivari, L.; Radhakrishnan, S.; Tarver, C. Resveratrol suppresses IGF-1 induced human colon cancer cell proliferation and elevates apoptosis via suppression of IGF-1R/Wnt and activation of p53 signaling pathways. BMC Cancer 2010, 10. [Google Scholar] [CrossRef]
- Albrecht, H.; Carraway, K.L., 3rd. MUC1 and MUC4: Switching the emphasis from large to small. Cancer Biother. Radiopharm. 2011, 26, 261–271. [Google Scholar] [CrossRef]
- Carraway, K.L.; Price-Schiavi, S.A.; Komatsu, M.; Jepson, S.; Perez, A.; Carraway, C.A. Muc4/sialomucin complex in the mammary gland and breast cancer. J. Mammary Gland Biol. Neoplasia 2001, 6, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.P.; Ponnusamy, M.P.; Chakraborty, S.; Smith, L.M.; Das, S.; Arafat, H.A.; Batra, S.K. Effects of Thymoquinone in the Expression of Mucin 4 in Pancreatic Cancer Cells: Implications for the Development of Novel Cancer Therapies. Mol. Cancer Ther. 2010, 9, 1419–1431. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Son, Y.J.; Lee, W.H.; Park, Y.W.; Chae, S.W.; Cho, W.J.; Kim, Y.M.; Choi, H.J.; Choi, D.H.; Jung, S.W.; et al. Tristetraprolin regulates expression of VEGF and tumorigenesis in human colon cancer. Int. J. Cancer 2010, 126, 1817–1827. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Lee, S.R.; Leem, S.H. Tristetraprolin Regulates Prostate Cancer Cell Growth Through Suppression of E2F1. J. Microbiol. Biotechn. 2014, 24, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Gali-Muhtasib, H.; Ocker, M.; Kuester, D.; Krueger, S.; El-Hajj, Z.; Diestel, A.; Evert, M.; El-Najjar, N.; Peters, B.; Jurjus, A.; et al. Thymoquinone reduces mouse colon tumor cell invasion and inhibits tumor growth in murine colon cancer models. J. Cell. Mol. Med. 2008, 12, 330–342. [Google Scholar] [CrossRef]
- Tran, D.D.H.; Koch, A.; Saran, S.; Armbrecht, M.; Ewald, F.; Koch, M.; Wahlicht, T.; Wirth, D.; Braun, A.; Nashan, B.; et al. Extracellular-signal regulated kinase (Erk1/2), mitogen-activated protein kinase-activated protein kinase 2 (MK2) and tristetraprolin (TTP) comprehensively regulate injury-induced immediate early gene (IEG) response in in vitro liver organ culture. Cell Signal 2016, 28, 438–447. [Google Scholar] [CrossRef]
- Chen, Y.L.; Jiang, Y.W.; Su, Y.L.; Lee, S.C.; Chang, M.S.; Chang, C.J. Transcriptional regulation of tristetraprolin by NF-kappa B signaling in LPS-stimulated macrophages. Mol. Biol. Rep. 2013, 40, 2867–2877. [Google Scholar] [CrossRef]
- Chehl, N.; Chipitsyna, G.; Gong, Q.K.; Yeo, C.J.; Arafat, H.A. Anti-inflammatory effects of the Nigella sativa seed extract, thymoquinone, in pancreatic cancer cells. HPB 2009, 11, 373–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, Y.; Higashi, M.; Kitamoto, S.; Yokoyama, S.; Osako, M.; Horinouchi, M.; Shimizu, T.; Tabata, M.; Batra, S.K.; Goto, M.; et al. MUC4 and MUC1 expression in adenocarcinoma of the stomach correlates with vessel invasion and lymph node metastasis: An immunohistochemical study of early gastric cancer. PloS ONE 2012, 7, e49251. [Google Scholar] [CrossRef]
- Rachagani, S.; Macha, M.A.; Ponnusamy, M.P.; Haridas, D.; Kaur, S.; Jain, M.; Batra, S.K. MUC4 potentiates invasion and metastasis of pancreatic cancer cells through stabilization of fibroblast growth factor receptor 1. Carcinogenesis 2012, 33, 1953–1964. [Google Scholar] [CrossRef]
- Wang, W.H.; Chen, J.; Zhao, F.; Zhang, B.R.; Yu, H.S.; Jin, H.Y.; Dai, J.H. MiR-150-5p suppresses colorectal cancer cell migration and invasion through targeting MUC4. Asian Pac. J. Cancer Prev. APJCP 2014, 15, 6269–6273. [Google Scholar] [CrossRef]
- Kayl, A.E.; Meyers, C.A. Side-effects of chemotherapy and quality of life in ovarian and breast cancer patients. Curr. Opin. Obstet. Gynecol. 2006, 18, 24–28. [Google Scholar] [CrossRef]
- Simonelli, L.E.; Siegel, S.D.; Duffy, N.M. Fear of cancer recurrence: A theoretical review and its relevance for clinical presentation and management. Psycho-oncology 2016, 26, 1444–1454. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-R.; Mun, J.-Y.; Jeong, M.-S.; Lee, H.-H.; Roh, Y.-G.; Kim, W.-T.; Kim, M.-H.; Heo, J.; Choi, Y.H.; Kim, S.J.; et al. Thymoquinone-Induced Tristetraprolin Inhibits Tumor Growth and Metastasis through Destabilization of MUC4 mRNA. Int. J. Mol. Sci. 2019, 20, 2614. https://doi.org/10.3390/ijms20112614
Lee S-R, Mun J-Y, Jeong M-S, Lee H-H, Roh Y-G, Kim W-T, Kim M-H, Heo J, Choi YH, Kim SJ, et al. Thymoquinone-Induced Tristetraprolin Inhibits Tumor Growth and Metastasis through Destabilization of MUC4 mRNA. International Journal of Molecular Sciences. 2019; 20(11):2614. https://doi.org/10.3390/ijms20112614
Chicago/Turabian StyleLee, Se-Ra, Jeong-Yeon Mun, Mi-So Jeong, Hyun-Hee Lee, Yun-Gil Roh, Won-Tae Kim, Min-Hye Kim, Jeonghoon Heo, Yung Hyun Choi, Su Jin Kim, and et al. 2019. "Thymoquinone-Induced Tristetraprolin Inhibits Tumor Growth and Metastasis through Destabilization of MUC4 mRNA" International Journal of Molecular Sciences 20, no. 11: 2614. https://doi.org/10.3390/ijms20112614