Comparative Proteomic Analysis of Wheat Carrying Pm40 Response to Blumeria graminis f. sp. tritici Using Two-Dimensional Electrophoresis

 ,
,
Abstract
:1. Introduction
2. Results
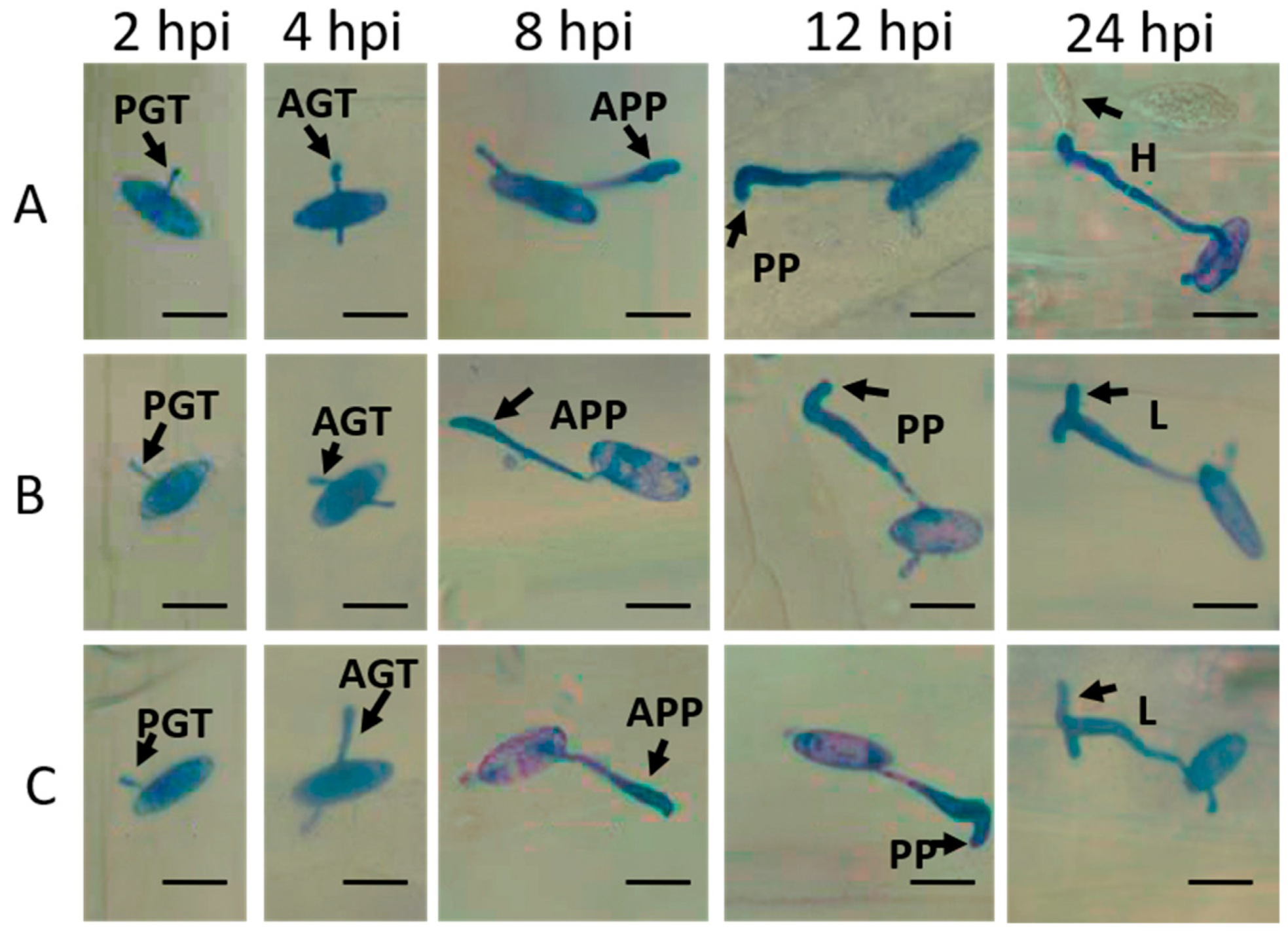
2.1. Phenotypic Differences of Leaves Affected by Bgt
2.2. Estimation of Wheat-Bgt Interactions
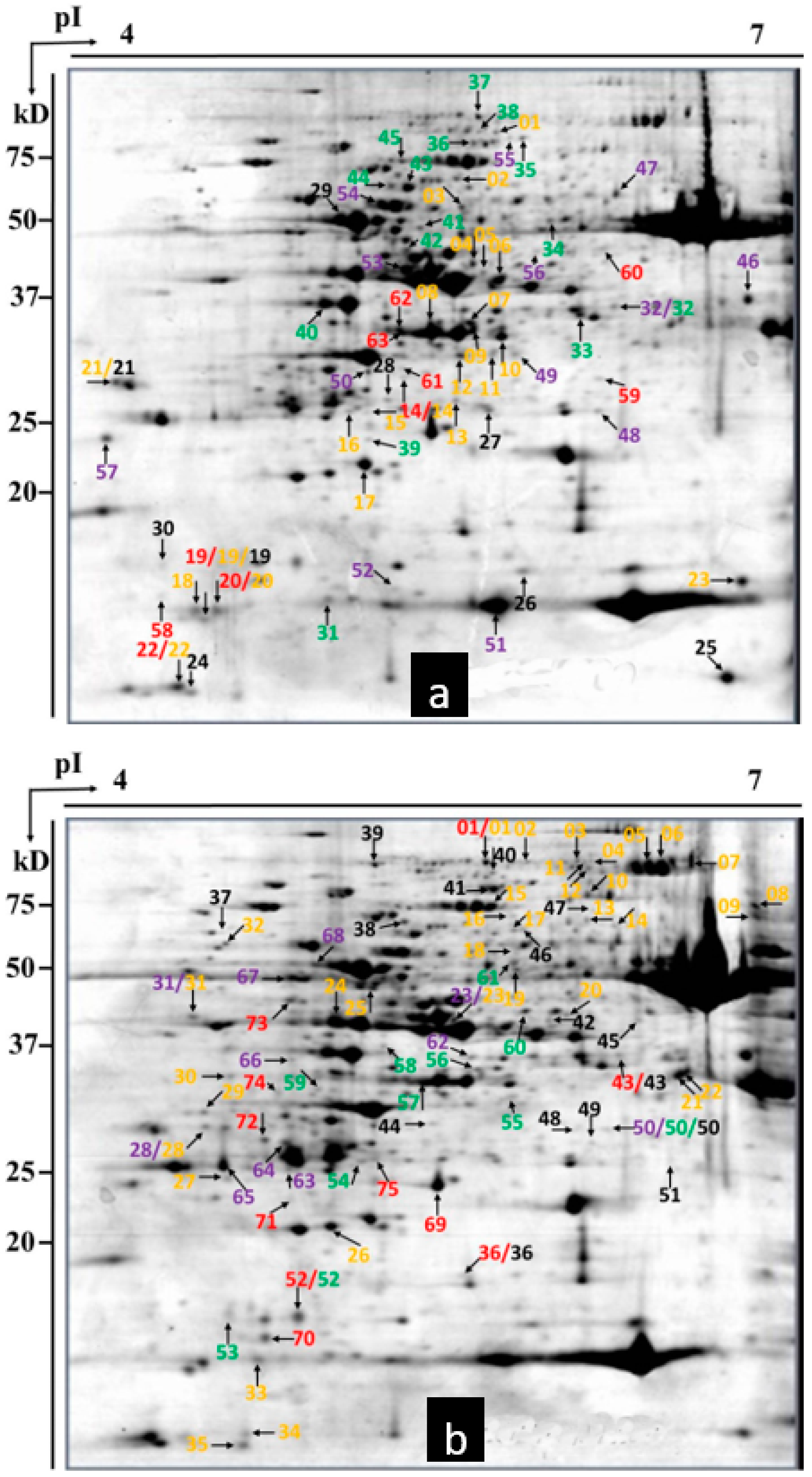
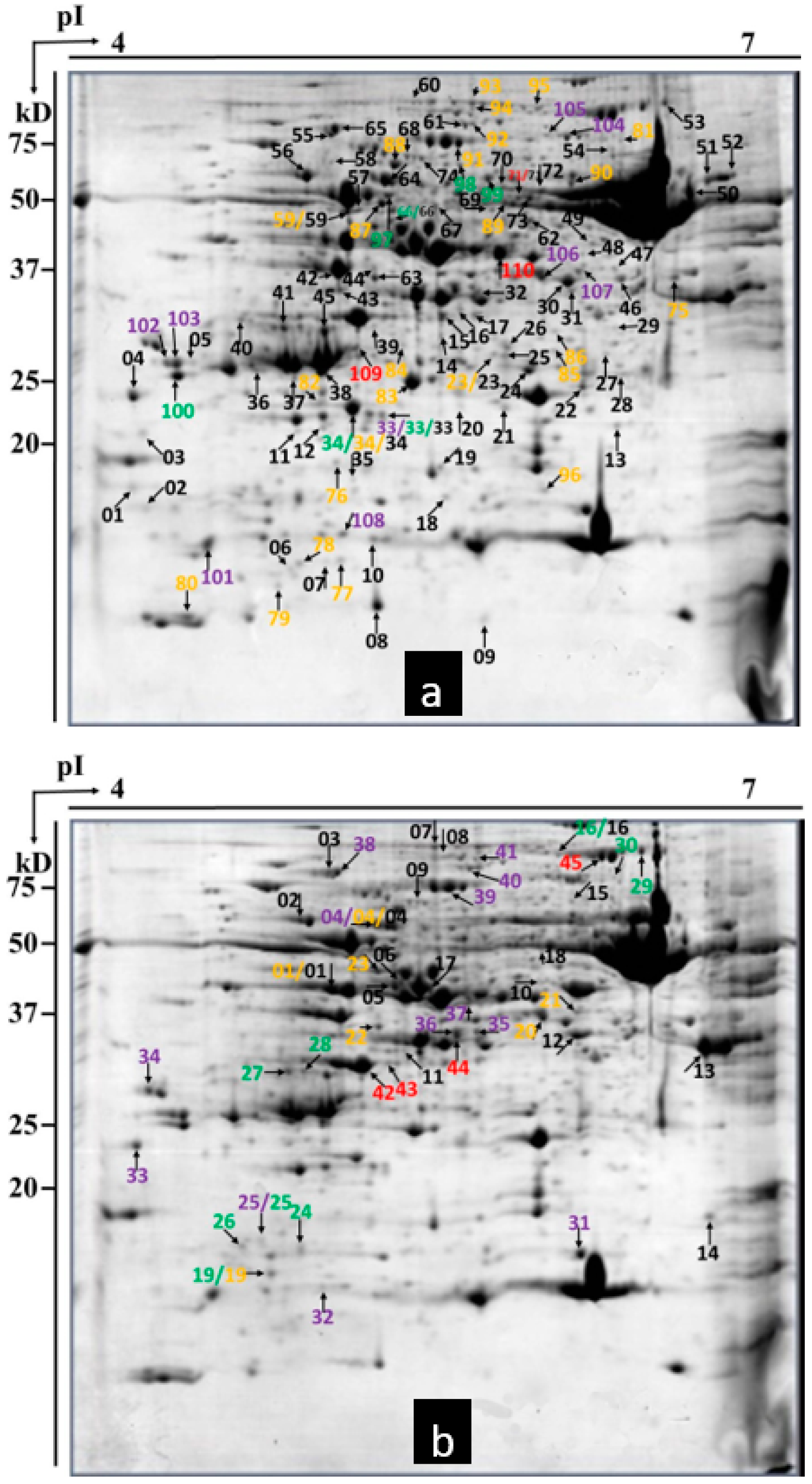
2.3. Detection of Differential Proteins by 2-DE
2.4. Protein Identification
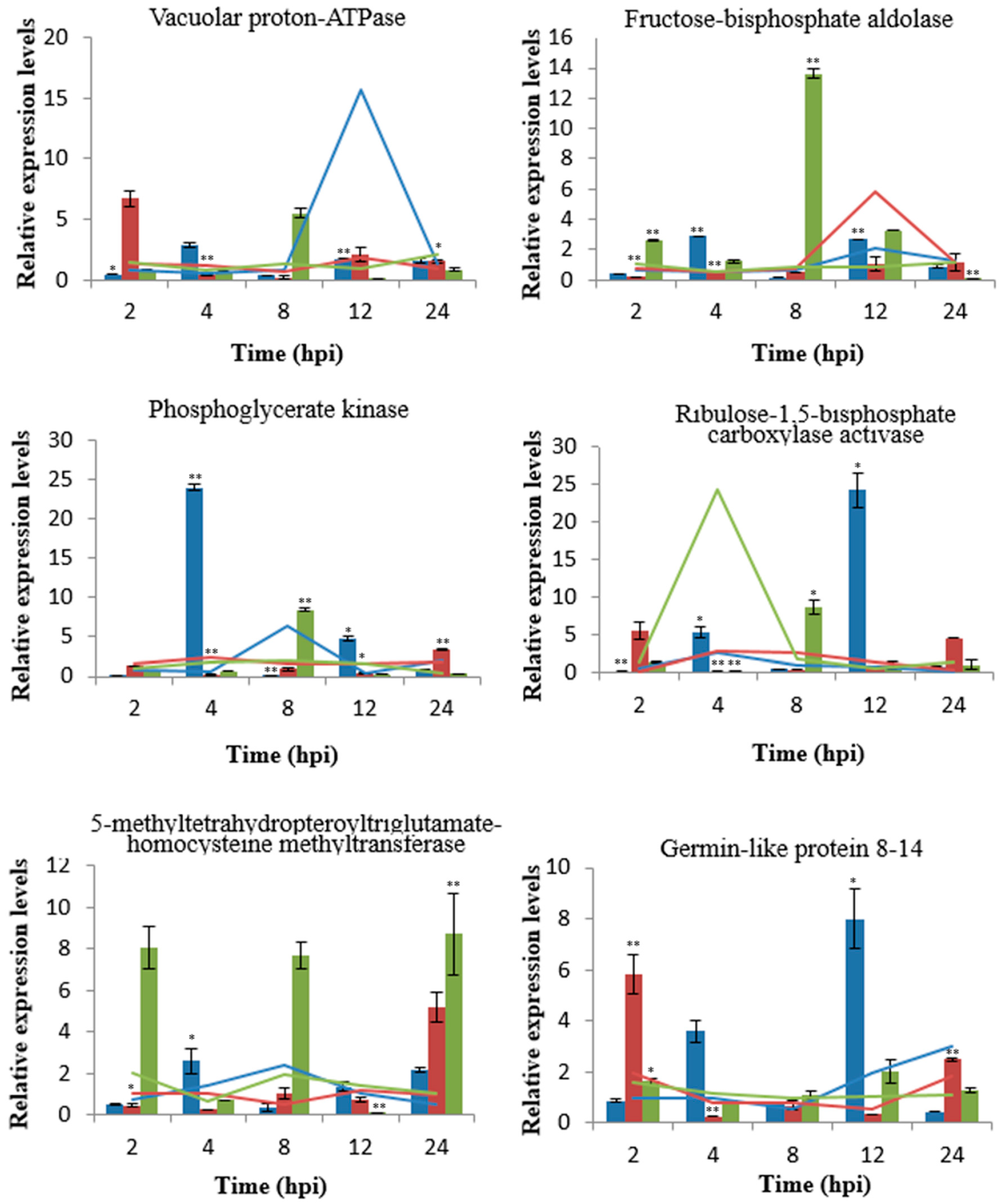
2.5. Validation of Upregulated Proteins by qRT-PCR
3. Discussion
3.1. Stress- and Defense-Related Proteins
3.2. Proteins Related to Photosynthesis
3.3. Proteins Involved in Carbohydrate Metabolism and Energy Pathways
3.4. Proteins Involved in Gene Expression and Protein Turnover
3.5. Proteins Associated with Cell Organization
3.6. The Correlation of mRNA and Protein Expression
3.7. The Novel Proteins Potentially Involed in the Response of Wheat Against Bgt
4. Materials and Methods
4.1. Plant Materials and Inoculation
4.2. Cytological Observation of the Interaction between Wheat and Bgt
4.3. Protein Extraction
4.4. Two-Dimensional Electrophoresis, Protein Visualization, and Image Analysis
4.5. MS and Database Searches
4.6. RNA Isolation and qRT-PCR Assays
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, Z.; Li, L. Seedling and adult plant resistance to powdery mildew in Chinese bread wheat cultivars and lines. Plant Dis. 2005, 89, 457–463. [Google Scholar] [CrossRef]
- Duveiller, E.; Singh, R. The challenges of maintaining wheat productivity: Pests, diseases, and potential epidemics. Euphytica 2007, 157, 417–430. [Google Scholar] [CrossRef]
- Hao, Y.; Parks, R. Molecular characterization of a new powdery mildew resistance gene Pm54 in soft red winter wheat. Theor. Appl. Genet. 2015, 128, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Sun, B. Pm55, a developmental-stage and tissue-specific powdery mildew resistance gene introgressed from Dasypyrum villosum into common wheat. Theor. Appl. Genet. 2016, 129, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- Hsam, S.; Huang, X. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell.). 5. Alleles at the Pm1 locus. Theor. Appl. Genet. 1998, 96, 1129–1134. [Google Scholar] [CrossRef]
- Singrün, C.; Hsam, S. Powdery mildew resistance gene Pm22 in cultivar Virest is a member of the complex Pm1 locus in common wheat (Triticum aestivum L. em Thell.). Theor. Appl. Genet. 2003, 106, 1420–1424. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Liu, A. Pm23: A new allele of Pm4 located on chromosome 2AL in wheat. Theor. Appl. Genet. 2008, 117, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Ben-David, R. Suppressed recombination rate in 6VS/6AL translocation region carrying the Pm21 locus introgressed from Haynaldia villosa into hexaploid wheat. Mol. Breed. 2012, 29, 399–412. [Google Scholar] [CrossRef]
- Zou, S.; Wang, H. The NB-LRR gene Pm60 confers powdery mildew resistance in wheat. New Phytol. 2018, 218, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Wiersma, A.T.; Pulman, J.A. Identification of Pm58 from Aegilops tauschii. Theor. Appl. Genet. 2017, 130, 1123–1133. [Google Scholar] [CrossRef]
- Tan, C.; Li, G. Characterization of Pm59, a novel powdery mildew resistance gene in Afghanistan wheat landrace PI 181356. Theor. Appl. Genet. 2018, 131, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Hu, J. Pm61: A recessive gene for resistance to powdery mildew in wheat landrace Xuxusanyuehuang identified by comparative genomics analysis. Theor. Appl. Genet. 2018, 131, 2085–2097. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Fan, Y. Pm62, an adult-plant powdery mildew resistance gene introgressed from Dasypyrum villosum chromosome arm 2VL into wheat. Theor. Appl. Genet. 2018, 131, 2613–2620. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Q.; Hsam, S.L.K. Molecular mapping of the wheat powdery mildew resistance gene Pm24 and marker validation for molecular breeding. Theor. Appl. Genet. 2000, 101, 407–414. [Google Scholar] [CrossRef]
- Consortium, I.W.G.S. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [Google Scholar]
- Yahiaoui, N.; Srichumpa, P. Genome analysis at different ploidy levels allows cloning of the powdery mildew resistance gene Pm3b from hexaploid wheat. Plant J. 2004, 37, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Cao, A.; Xing, L. Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc. Natl. Acad. Sci. USA 2011, 108, 7727–7732. [Google Scholar] [CrossRef]
- Hurni, S.; Brunner, S. Rye Pm8 and wheat Pm3 are orthologous genes and show evolutionary conservation of resistance function against powdery mildew. Plant J. 2013, 76, 957–969. [Google Scholar] [CrossRef]
- Sánchez-Martín, J.; Steuernagel, B. Rapid gene isolation in barley and wheat by mutant chromosome sequencing. Genome Biol. 2016, 17, 221. [Google Scholar] [CrossRef]
- Xing, L.; Hu, P. NLR1-V, a CC-NBS-LRR encoding gene, is a potential candidate gene of the wheat powdery mildew resistance gene Pm21. bioRxiv 2017, 114058. [Google Scholar] [CrossRef]
- Luo, P.G.; Luo, H. Characterization and chromosomal location of Pm40 in common wheat: A new gene for resistance to powdery mildew derived from Elytrigia intermedium. Theor. Appl. Genet. 2009, 118, 1059–1064. [Google Scholar] [CrossRef]
- Zhong, S.; Ma, L. Collinearity analysis and high-density genetic mapping of the wheat powdery mildew resistance gene Pm40 in PI 672538. PLoS ONE 2016, 11, e0164815. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Xu, M. Registration of the novel wheat lines L658, L693, L696, and L699, with resistance to Fusarium Head blight, stripe rust, and powdery mildew. J. Plant Registrat. 2015, 9, 121–124. [Google Scholar] [CrossRef]
- Li, J.; Yang, X. Proteomic analysis of the compatible interaction of wheat and powdery mildew (Blumeria graminis f. sp. tritici). Plant Physiol. Biochem. 2017, 111, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.S.; Elenitoba-Johnson, K.S.J. Proteomics in pathology research. Lab. Investig. 2004, 84, 1227–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marra, R.; Ambrosino, P. Study of the three-way interaction between Trichoderma atroviride, plant and fungal pathogens by using a proteomic approach. Curr. Genet. 2006, 50, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, X. Differences in protein expression and ultrastructure between two wheat near-isogenic lines affected by powdery mildew. Russ. J. Plant Physiol. 2011, 58, 686. [Google Scholar] [CrossRef]
- Wang, B.; Xie, C. Comparative proteomic analysis of wheat response to powdery mildew infection in wheat Pm30 near-lsogenic lines. J. Phytopathol. 2012, 160, 229–236. [Google Scholar] [CrossRef]
- Mandal, M.S.N.; Fu, Y. Proteomic analysis of the defense response of wheat to the powdery mildew fungus, Blumeria graminis f. sp. tritici. Protein J. 2014, 33, 513–524. [Google Scholar] [CrossRef]
- Fu, Y.; Zhang, H. Quantitative proteomics reveals the central changes of wheat in response to powdery mildew. J. Proteom. 2016, 130, 108–119. [Google Scholar] [CrossRef]
- Luo, L.Y. Defense Responses Induced by Bgt in The Novel Wheat Line L699 Carrying a Pm40 Gene; Sichuan Agricultural University: Ya’an, China, 2016. [Google Scholar]
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.T.; Coaker, G. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Hammond-Kosack, K.E.; Jones, J.D. Resistance gene-dependent plant defense responses. Plant Cell 1996, 8, 1773–1791. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.M.; Senthil-Kumar, M. Regulation of primary plant metabolism during plant-pathogen interactions and its contribution to plant defense. Front. Plant Sci. 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bernier, F.; Berna, A. Germins and germin-like proteins: Plant do-all proteins. But what do they do exactly? Plant Physiol. Biochem. 2001, 39, 545–554. [Google Scholar] [CrossRef]
- Membré, N.; Bernier, F. Arabidopsis thaliana germin-like proteins: Common and specific features point to a variety of functions. Planta 2000, 211, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Park, C.-J.; An, J.-M. Molecular characterization of pepper germin-like protein as the novel PR-16 family of pathogenesis-related proteins isolated during the resistance response to viral and bacterial infection. Planta 2004, 219, 797–806. [Google Scholar] [CrossRef]
- Zimmermann, G.; Bäumlein, H. The multigene family encoding germin-like proteins of barley. Regulation and function in basal host resistance. Plant Physiol. 2006, 142, 181–192. [Google Scholar] [CrossRef]
- Deeba, F.; Sultana, T. Involvement of WRKY, MYB and DOF DNA-binding proteins in interaction with a rice germin-like protein gene promoter. Acta Physiol. Plant. 2017, 39, 189. [Google Scholar] [CrossRef]
- Camejo, D.; Guzmán-Cedeño, Á. Reactive oxygen species, essential molecules, during plant-pathogen interactions. Plant Physiol. Biochem. 2016, 103, 10–23. [Google Scholar]
- Dunwell, J.M.; Gibbings, J.G. Germin and germin-like proteins: Evolution, structure, and function. Crit. Rev. Plant Sci. 2008, 27, 342–375. [Google Scholar] [CrossRef]
- Schweizer, P.; Christoffel, A. Transient expression of members of the germin-like gene family in epidermal cells of wheat confers disease resistance. Plant J. 1999, 20, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed]
- Ritossa, F. A new puffing pattern induced by temperature shock and DNP in Drosophila. Experientia 1962, 18, 571–573. [Google Scholar] [CrossRef]
- Tissiéres, A.; Mitchell, H.K. Protein synthesis in salivary glands of Drosophila melanogaster: Relation to chromosome puffs. J. Mol. Biol. 1974, 84, 389–398. [Google Scholar] [CrossRef]
- Neckers, L.; Tatu, U. Molecular chaperones in pathogen virulence: Emerging new targets for therapy. Cell Host Microbe 2008, 4, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Bakthisaran, R.; Tangirala, R. Small heat shock proteins: Role in cellular functions and pathology. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2015, 1854, 291–319. [Google Scholar] [CrossRef] [Green Version]
- Siedow, J.N. Plant lipoxygenase: Structure and function. Annu. Rev. Plant Biol. 1991, 42, 145–188. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Vick, B.A.; Zimmerman, D. Oxidative systems for modification of fatty acids: The lipoxygenase pathway. Biochem. Plants 1987, 9, 53–90. [Google Scholar]
- Feussner, L.; Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 2002, 53, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Bilgin, D.D.; Zavala, J.A. Biotic stress globally downregulates photosynthesis genes. Plant Cell Environ. 2010, 33, 1597–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholes, J.; Lee, P. Photosynthetic metabolism in leaves infected with powdery mildew. Curr. Res. Photosynth. 1990, 4, 219–222. [Google Scholar]
- Swarbrick, P.J.; Schulze-lefert, P. Metabolic consequences of susceptibility and resistance (race-specific and broad-spectrum) in barley leaves challenged with powdery mildew. Plant Cell Environ. 2006, 29, 1061–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, J.; So, B.R. Resampling and editing of mischarged tRNA prior to translation elongation. Mol. Cell 2009, 33, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Zaher, H.S.; Green, R. Quality control by the ribosome following peptide bond formation. Nature 2009, 457, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Park, S.G.; Ewalt, K.L. Functional expansion of aminoacyl-tRNA synthetases and their interacting factors: New perspectives on housekeepers. Trends Biochem. Sci. 2005, 30, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Henty-Ridilla, J.L.; Shimono, M. The plant actin cytoskeleton responds to signals from microbe-associated molecular patterns. PLoS Pathog. 2013, 9, e1003290. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analy. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Candiano, G.; Bruschi, M. Blue silver: A very sensitive colloidal Coomassie G-250 staining for proteome analysis. Electrophoresis 2004, 25, 1327–1333. [Google Scholar] [CrossRef]
- Balaji, B.; Bucholtz, D.B. Barley yellow dwarf virus and Cereal yellow dwarf virus quantification by real-time polymerase chain reaction in resistant and susceptible plants. Phytopathology 2003, 93, 1386–1392. [Google Scholar] [CrossRef] [PubMed]
- Livaka, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
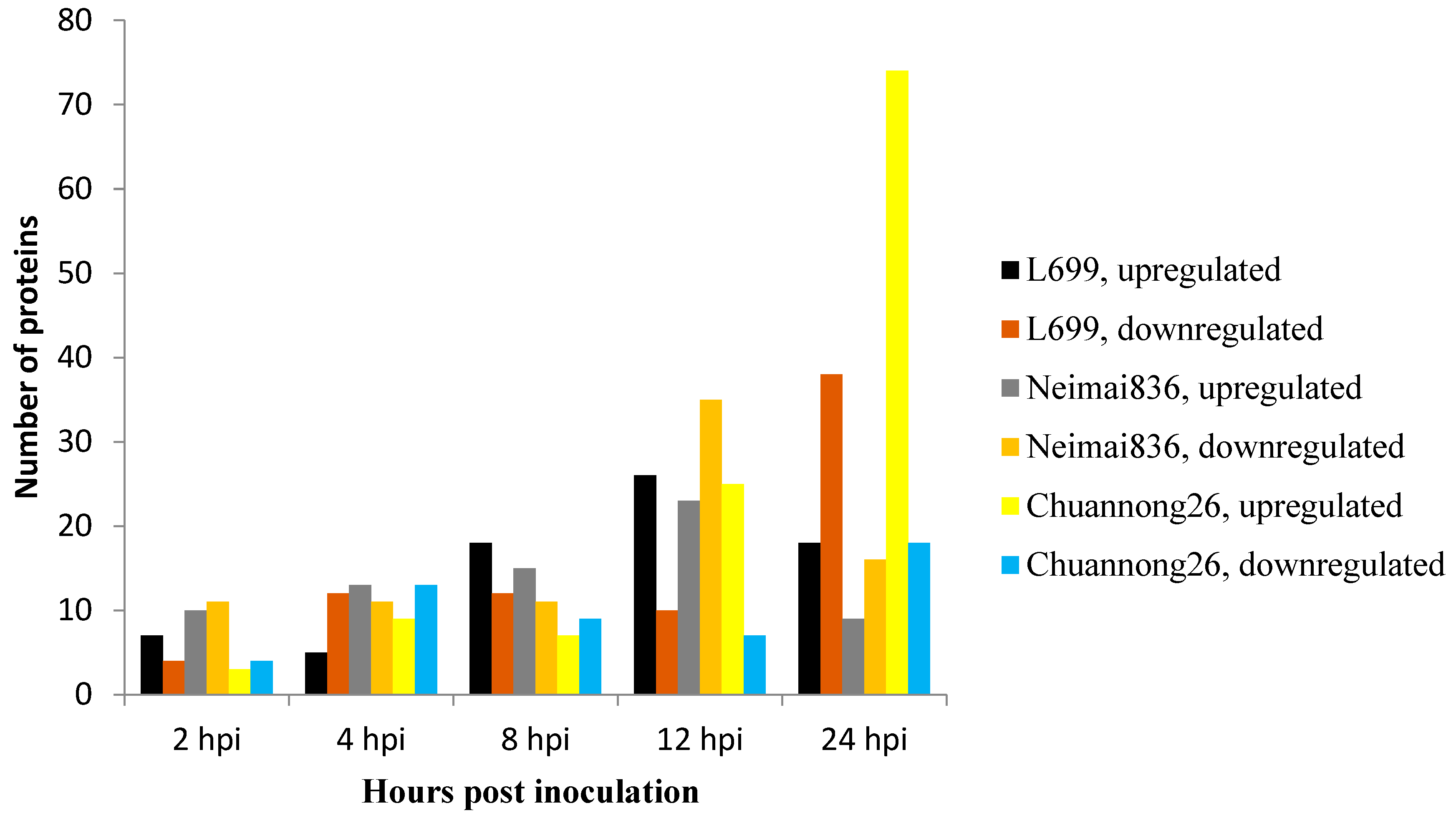{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot | Protein Name | Accession | Matched Peptides | MW/PI | Score | Time (h) |
|---|---|---|---|---|---|---|
| Proteins involved in disease defense response | ||||||
| 3 | Lipoxygenase 2.1, chloroplastic | gi|473948122 | 21 | 105625.33/5.70 | 185 | 12 |
| 4 | Lipoxygenase 2.1, chloroplastic | gi|473948122 | 41 | 105625.33/5.70 | 591 | 12 |
| 5 | Lipoxygenase 2.1, chloroplastic | gi|473948122 | 17 | 105625.33/5.70 | 196 | 12 |
| 29 | Lipoxygenase 1 | gi|474399175 | 16 | 96333.65/5.91 | 299 | 8 |
| 32 | Heat shock cognate 70 kDa protein 1 | gi|474012573 | 37 | 71123.58/5.06 | 641 | 8 |
| 61 | Germin-like protein 8-14 | gi|473963025 | 4 | 21939.25/5.36 | 174 | 24 |
| Photosynthesis-related proteins | ||||||
| 11 | Ribulose-1,5-bisphosphate carboxylase activase, partial | gi|37783283 | 10 | 22336.08/4.98 | 309 | 8, 12 |
| 28 | Ribulose bisphosphate carboxylase small chain, chloroplastic | gi|473882355 | 14 | 18526.35/8.65 | 208 | 12 |
| 33 | RuBisCO large subunit-binding protein subunit alpha, chloroplastic | gi|474113969 | 34 | 65380.60/5.17 | 864 | 8 |
| 36 | Ribulose-1,5-bisphosphate carboxylase activase, partial | gi|37783283 | 10 | 22336.08/4.98 | 330 | 4 |
| 37 | Photosystem II cytochrome b559 alpha subunit (chloroplast) | gi|699976019 | 6 | 9444.60/4.64 | 196 | 4 |
| 44 | Ribulose bisphosphate carboxylase small chain PWS4.3, chloroplastic | gi|132087 | 2 | 19417.36/8.99 | 92 | 4, 8 |
| Proteins involved in Signal transduction | ||||||
| 24 | 14-3-3 protein | gi|431822520 | 16 | 29264.88/4.83 | 434 | 12 |
| Carbohydrate metabolism-related proteins | ||||||
| 6 | Beta-d-glucan exohydrolase | gi|20259685 | 14 | 67301.15/6.87 | 74 | 12 |
| 10 | Glucose-1-phosphate adenylyltransferasesmall subunit, chloroplastic/amyloplastic | gi|474108293 | 23 | 64723.14/7.91 | 266 | 12 |
| 13 | Phosphoglycerate kinase | gi|3293043 | 16 | 49839.53/6.57 | 580 | 8, 12 |
| 14 | Glycerophosphodiester phosphodiesterase GDE1 | gi|473847956 | 13 | 52899.68/5.69 | 43 | 12 |
| 18 | Fructose-bisphosphate aldolase, chloroplastic | gi|473848356 | 15 | 42002.99/5.94 | 358 | 12 |
| 38 | Phosphoribulokinase | gi|5924030 | 22 | 45141.39/5.72 | 587 | 8 |
| 50 | Phosphoglucomutase, cytoplasmic | gi|473763033 | 18 | 63499.68/5.14 | 302 | 2, 24 |
| 51 | Phosphoglucomutase, partial | gi|18076790 | 15 | 62789.15/5.66 | 218 | 24 |
| 54 | 6-phosphogluconate dehydrogenase, decarboxylating | gi|474379872 | 23 | 81169.95/8.56 | 608 | 24 |
| 55 | Cytosolic 3-phosphoglycerate kinase, partial | gi|28172911 | 16 | 31334.35/4.98 | 291 | 8, 24 |
| Proteins involved in energy pathway | ||||||
| 9 | Vacuolar proton-ATPase subunit A | gi|90025017 | 37 | 68454.90/5.23 | 583 | 12 |
| 17 | ATP synthase CF1 beta subunit (chloroplast) | gi|667669997 | 33 | 53857.48/5.06 | 1200 | 4 |
| 39 | Ferredoxin-NADP(H) oxidoreductase | gi|20302473 | 10 | 40232.03/6.92 | 120 | 8 |
| Proteins involved in gene expression and DNA remodeling | ||||||
| 15 | Guanine nucleotide-binding protein subunit beta-like | gi|473957859 | 6 | 27150.69/6.29 | 211 | 12 |
| Proteins involved in protein turnover | ||||||
| 8 | ATP-dependent zinc metalloprotease FTSH 1, chloroplastic | gi|474350516 | 29 | 54477.49/5.58 | 673 | 12 |
| 27 | 50S Ribosomal protein L12-2, chloroplastic | gi|475532245 | 10 | 21837.90/5.35 | 452 | 12 |
| 30 | Tyrosine phosphorylation protein A | gi|548319365 | 25 | 74252.07/6.61 | 434 | 8 |
| 34 | 5-methyltetrahydropteroyltriglutamate-Homocysteine methyltransferase | gi|473993302 | 14 | 84552.49/5.74 | 423 | 8 |
| 35 | 5, 10-methylene-tetrahydrofolate reductase | gi|115589742 | 12 | 64875.07/5.86 | 83 | 8 |
| 40 | 20 kDa chaperonin, chloroplastic | gi|474407512 | 10 | 29710.03/6.76 | 154 | 8 |
| 47 | Putative alanyl-tRNA synthetase, chloroplastic | gi|474142555 | 12 | 111648.20/5.62 | 198 | 24 |
| 48 | ATP-dependent Clp protease ATP-binding subunit ClpA-like protein CD4B, chloroplastic | gi|474241774 | 33 | 82735.21/5.16 | 513 | 12, 24 |
| 49 | Lysyl-tRNA synthetase | gi|474147702 | 8 | 132545.46/6.28 | 92 | 24 |
| 52 | Putative mitochondrial-processing peptidase subunit beta | gi|474142281 | 30 | 43290.34/5.41 | 486 | 24 |
| 53 | Adenosylhomocysteinase | gi|474154141 | 8 | 45700.84/6.48 | 56 | 24 |
| 56 | Cysteine synthase | gi|474315986 | 13 | 35583.27/5.82 | 216 | 12, 24 |
| 60 | Ribosome-recycling factor, chloroplastic | gi|474043078 | 15 | 24770.60/8.92 | 504 | 24 |
| Cell structure-related proteins | ||||||
| 12 | Actin-3 | gi|474259583 | 18 | 44367.62/5.26 | 376 | 8, 12 |
| Proteins of unknow function | ||||||
| 2 | Hypothetical protein TRIUR3_05354 | gi|473755342 | 27 | 104676.25/5.87 | 368 | 2, 12 |
| 26 | Unnamed protein product | gi|669029445 | 4 | 18152.74/5.60 | 244 | 12 |
| 58 | Hypothetical protein TRIUR3_21449 | gi|474384687 | 14 | 32942.26/9.31 | 129 | 24 |
| 59 | Unnamed protein product | gi|669027704 | 13 | 26818.76/5.57 | 335 | 24 |
| 63 | Unnamed protein product | gi|669029445 | 5 | 18152.74/5.60 | 269 | 24 |
| Spot | Protein Name | Accession No. | Primer Sequence 5′-3′ |
|---|---|---|---|
| Reference gene | 18S rRNA | AY049040 | Sense: 5′-GTGACGGGTGACGGAGAATT-3′ |
| Antisense: 5′-GACACTAATGCGCCCGGTAT-3′ | |||
| 9 | Vacuolar proton-ATPase | ABD85016 | Sense: 5′-TATGAACGTGCTGGGAAGGT-3′ |
| Antisense: 5′-GGGTTGCAGAGGTAACAGGA-3′ | |||
| 18 | Fructose-bisphosphate aldolase | EMS47455 | Sense: 5′-TCTTGTCTGGTGGTCAGTCG-3′ |
| Antisense: 5′-CGTCTTGAGGCAGGTGTTCT-3′ | |||
| 13 | Phosphoglycerate kinase | CAA51931 | Sense: 5′-AATGGTGCTGTTTTGCTCCT-3′ |
| Antisense: 5′-TGTTCCGAATGCATCGTTTA-3′ | |||
| 36 | Ribulose-1,5-bisphosphate carboxylase activase | AAP72270 | Sense: 5′-ACGGACCAGTGACCTTTGAG-3′ |
| Antisense: 5′-ACCAGTCTTCATCGCATCCT-3′ | |||
| 34 | 5-methyltetrahydropteroyltriglutamate-homocysteine methyltransferase | EMS51950 | Sense: 5′-TGTGTTCTGGTCCAAGATGG-3′ |
| Antisense: 5′-CTCAAACCTCGGTTGGTCAT-3′ | |||
| 61 | Germin-like protein 8-14 | EMS51159 | Sense: 5′-TGCAGATCACCGACTACGC-3′ |
| Antisense: 5′-CACGGACTTGAGCTTCTTGAC-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Y.; Xia, Y.; Chang, X.; Gong, G.; Yang, J.; Hu, Y.; Cahill, M.; Luo, L.; Li, T.; He, L.; et al. Comparative Proteomic Analysis of Wheat Carrying Pm40 Response to Blumeria graminis f. sp. tritici Using Two-Dimensional Electrophoresis. Int. J. Mol. Sci. 2019, 20, 933. https://doi.org/10.3390/ijms20040933
Liang Y, Xia Y, Chang X, Gong G, Yang J, Hu Y, Cahill M, Luo L, Li T, He L, et al. Comparative Proteomic Analysis of Wheat Carrying Pm40 Response to Blumeria graminis f. sp. tritici Using Two-Dimensional Electrophoresis. International Journal of Molecular Sciences. 2019; 20(4):933. https://doi.org/10.3390/ijms20040933
Chicago/Turabian StyleLiang, Yinping, Ye Xia, Xiaoli Chang, Guoshu Gong, Jizhi Yang, Yuting Hu, Madison Cahill, Liya Luo, Tao Li, Lu He, and et al. 2019. "Comparative Proteomic Analysis of Wheat Carrying Pm40 Response to Blumeria graminis f. sp. tritici Using Two-Dimensional Electrophoresis" International Journal of Molecular Sciences 20, no. 4: 933. https://doi.org/10.3390/ijms20040933