Molecular Characterization, Expression and Functional Analysis of Chicken STING

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chicken, Virus and Tissue Sample Collection
2.2. Newcastle Disease Virus (NDV) Challenge and Sample Collection
2.3. Cell Culture, STING Knockdown, Virus Infection and Transfection of Poly (I:C) and Poly (dA:dT)
2.4. Total RNA Extraction and Reverse Transcription
2.5. Cloning and Sequencing of the Full-Length cDNA for Chicken’s STING Gene
2.6. Quantitative Polymerase Chain Reaction (qPCR) Analysis of Chicken STING, IRF7, IFN-α and IFN-β mRNA
2.7. Sequence Comparison, Multiple Sequences Alignment, and Statistical Analysis
3. Results
3.1. Bioinformatics Analysis of STING
3.1.1. Cloning and Sequence Analysis of the Chicken STING
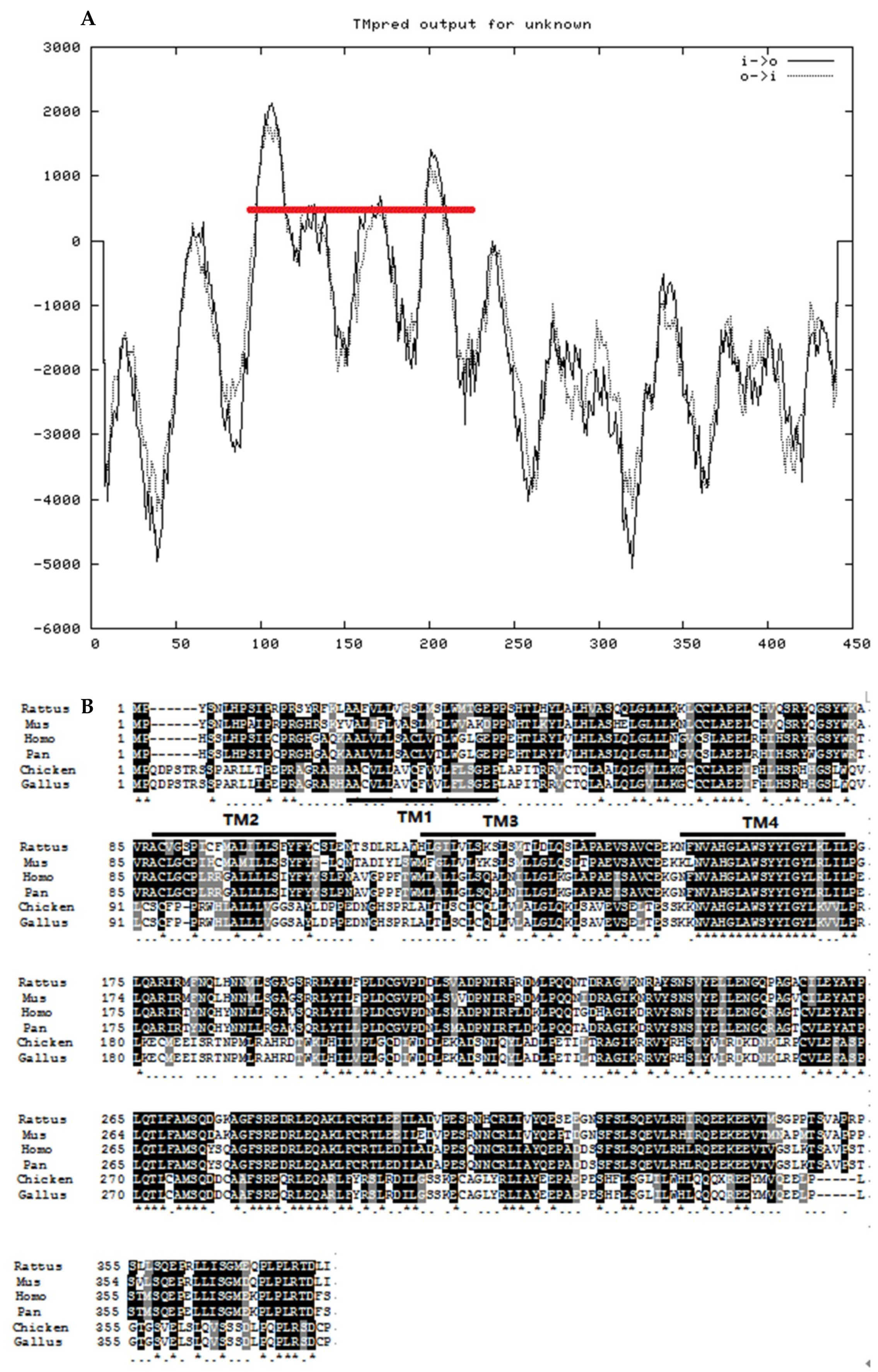
3.1.2. Structure Analysis of Chicken STING
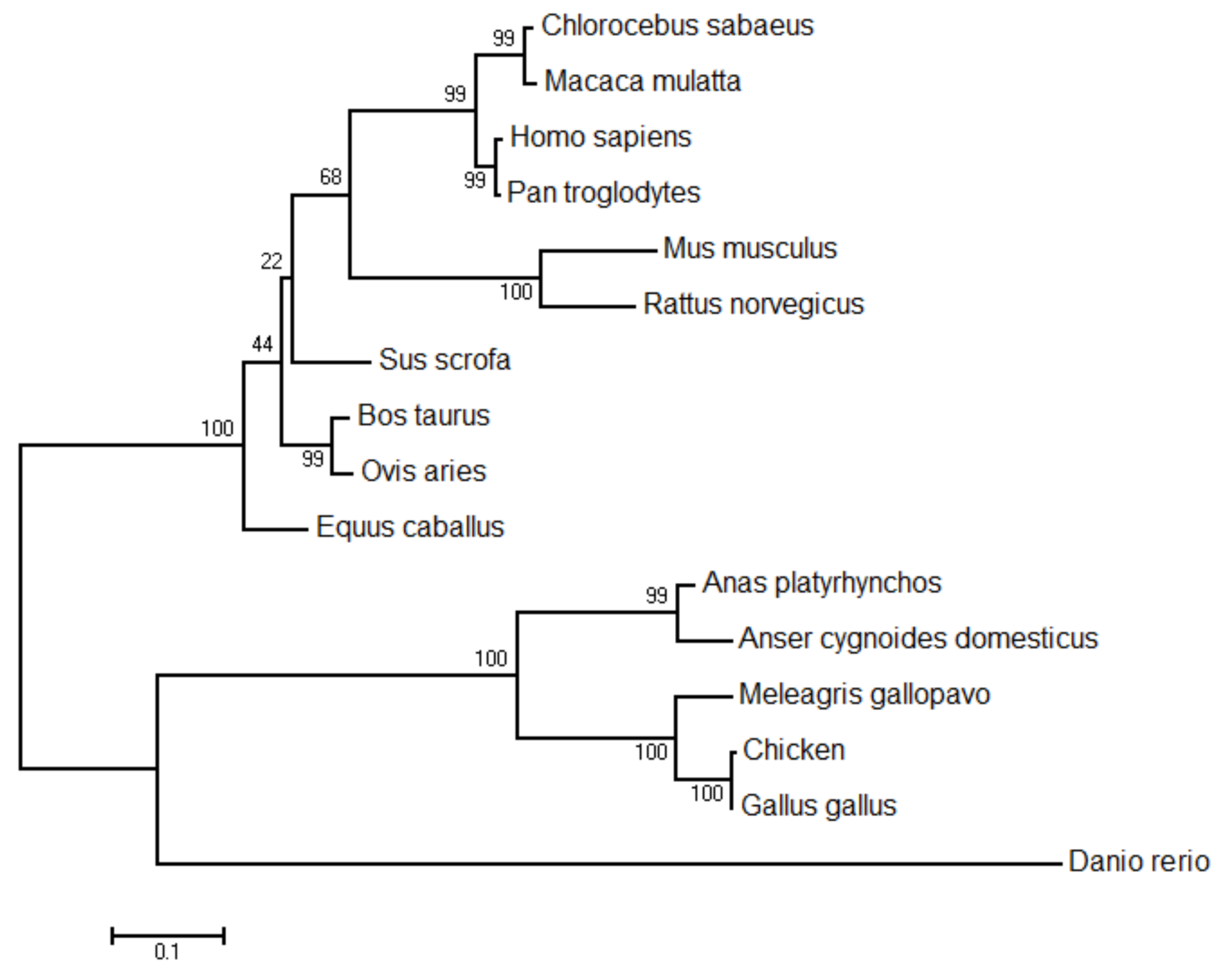
3.1.3. Comparison and Evolutionary Analyses of Chicken STING
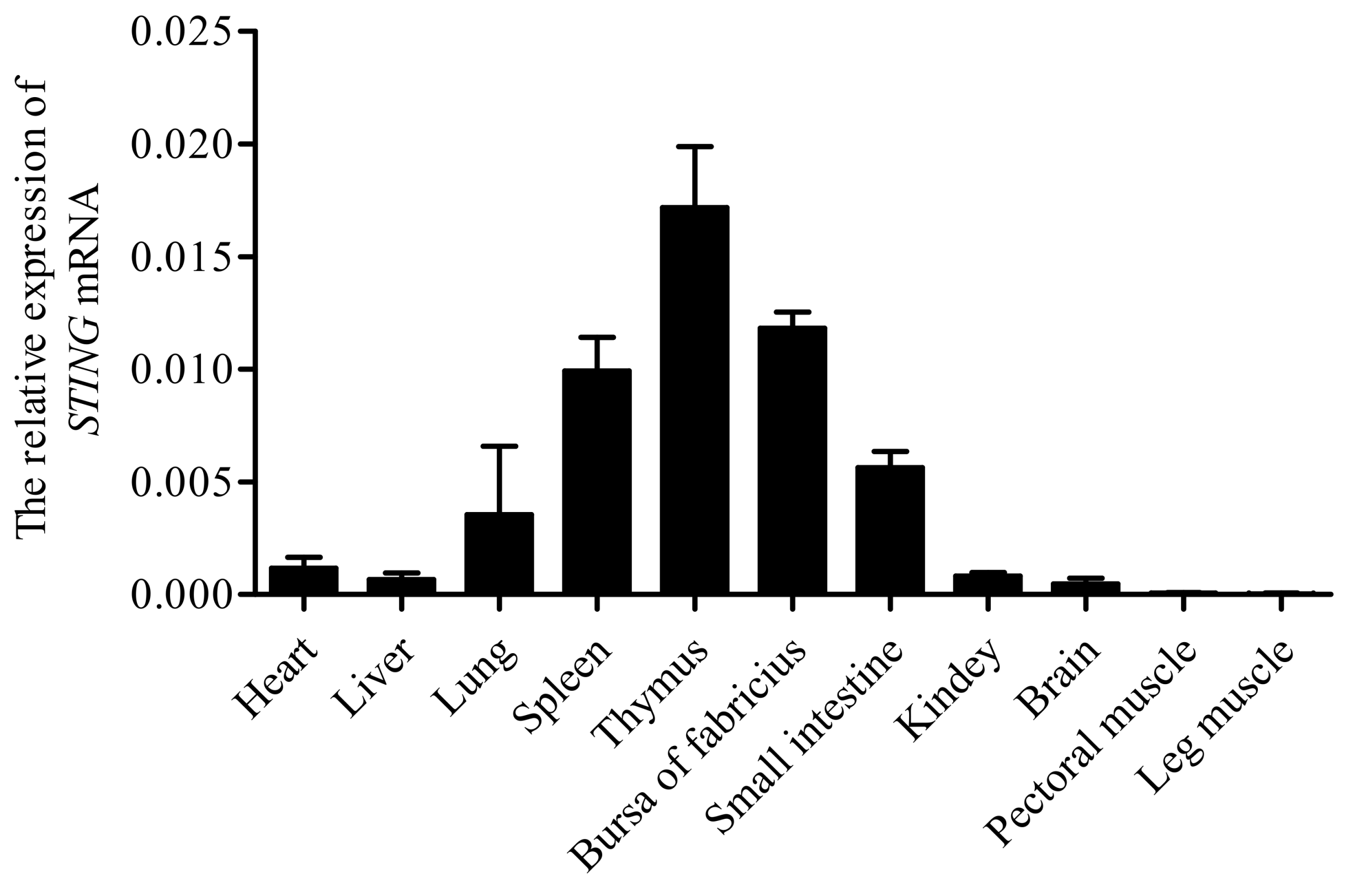
3.2. Tissue Distribution of STING mRNA
3.3. Antiviral Function Analysis of STING
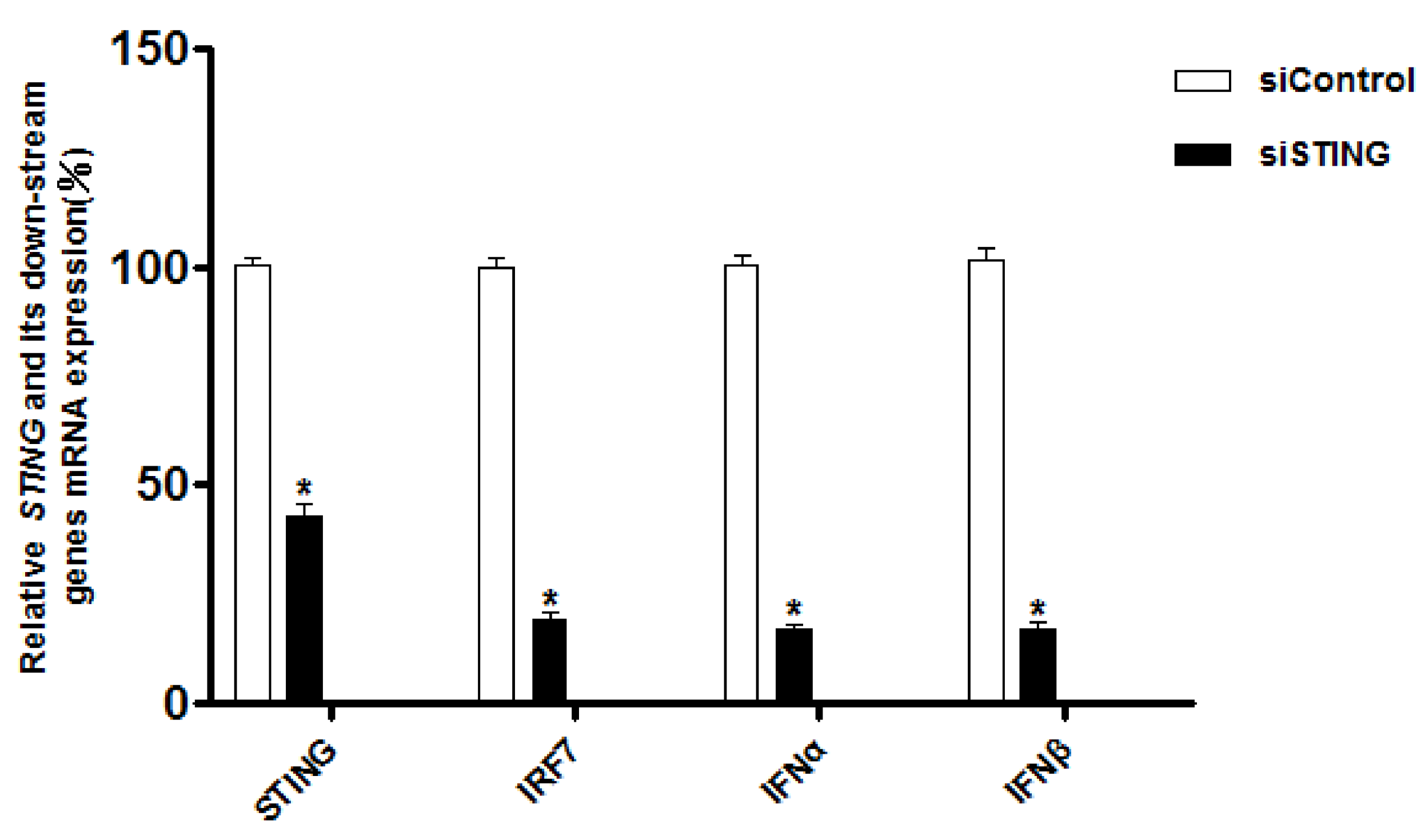
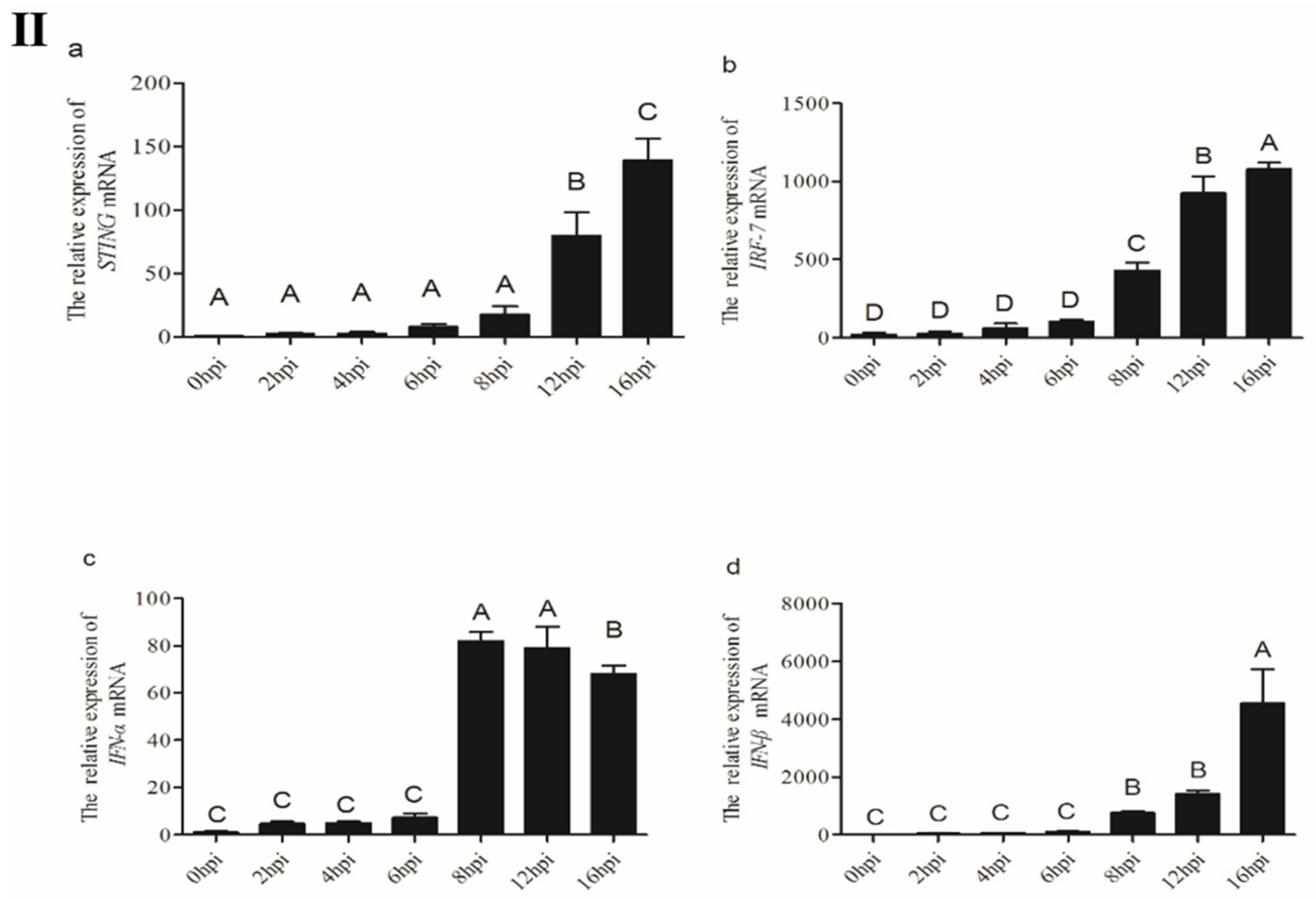
3.3.1. Interference with STING Gene Inhibits the Expression of IRF7, IFN-α and IFN-β Genes in CEFs
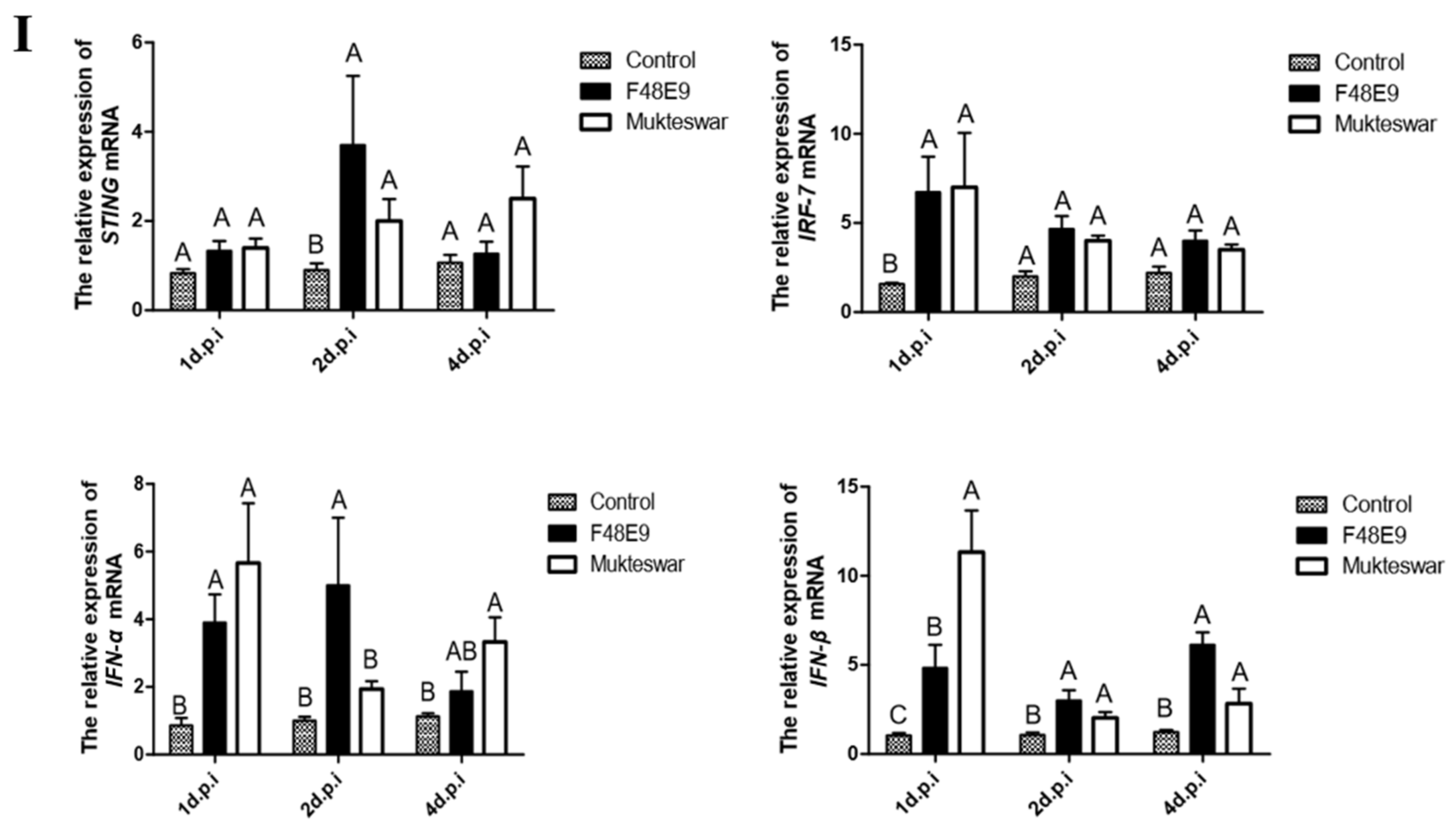
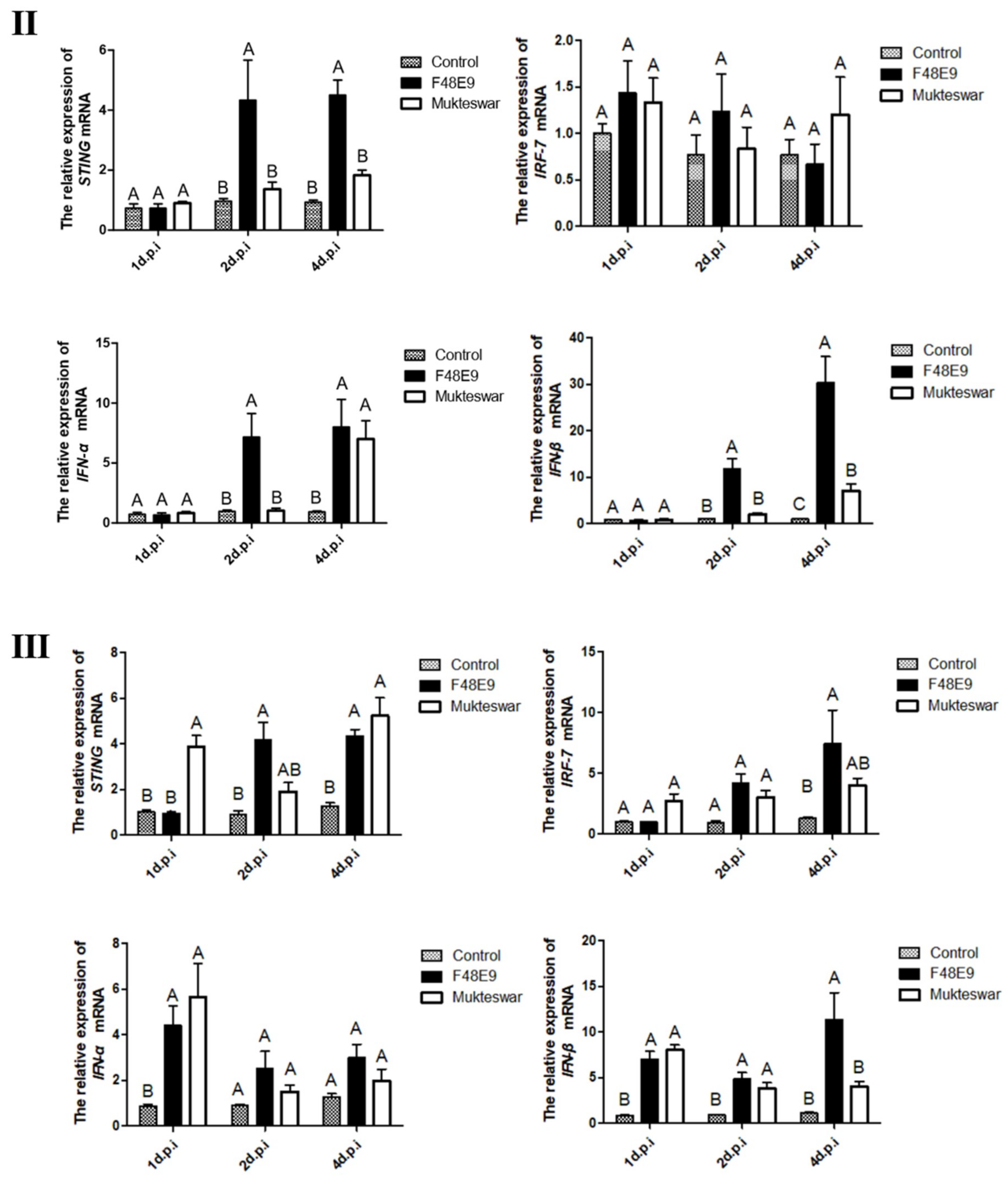
3.3.2. Expression of STING and Its Down-Stream Genes after In Vivo Infection by NDV Strains
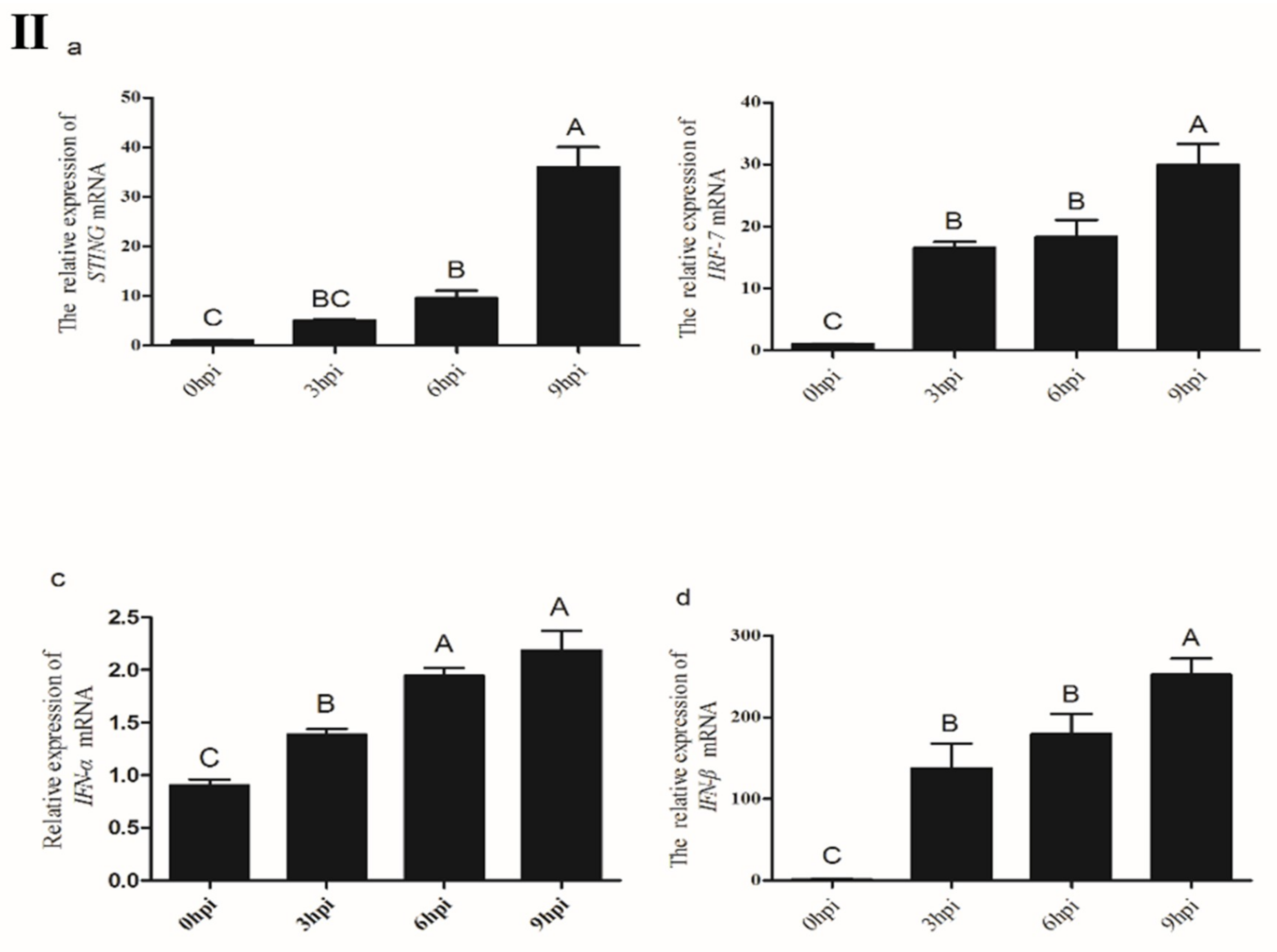
3.3.3. Expression of STING and its Down-Stream Genes Following CEFs’ Infection with NDV
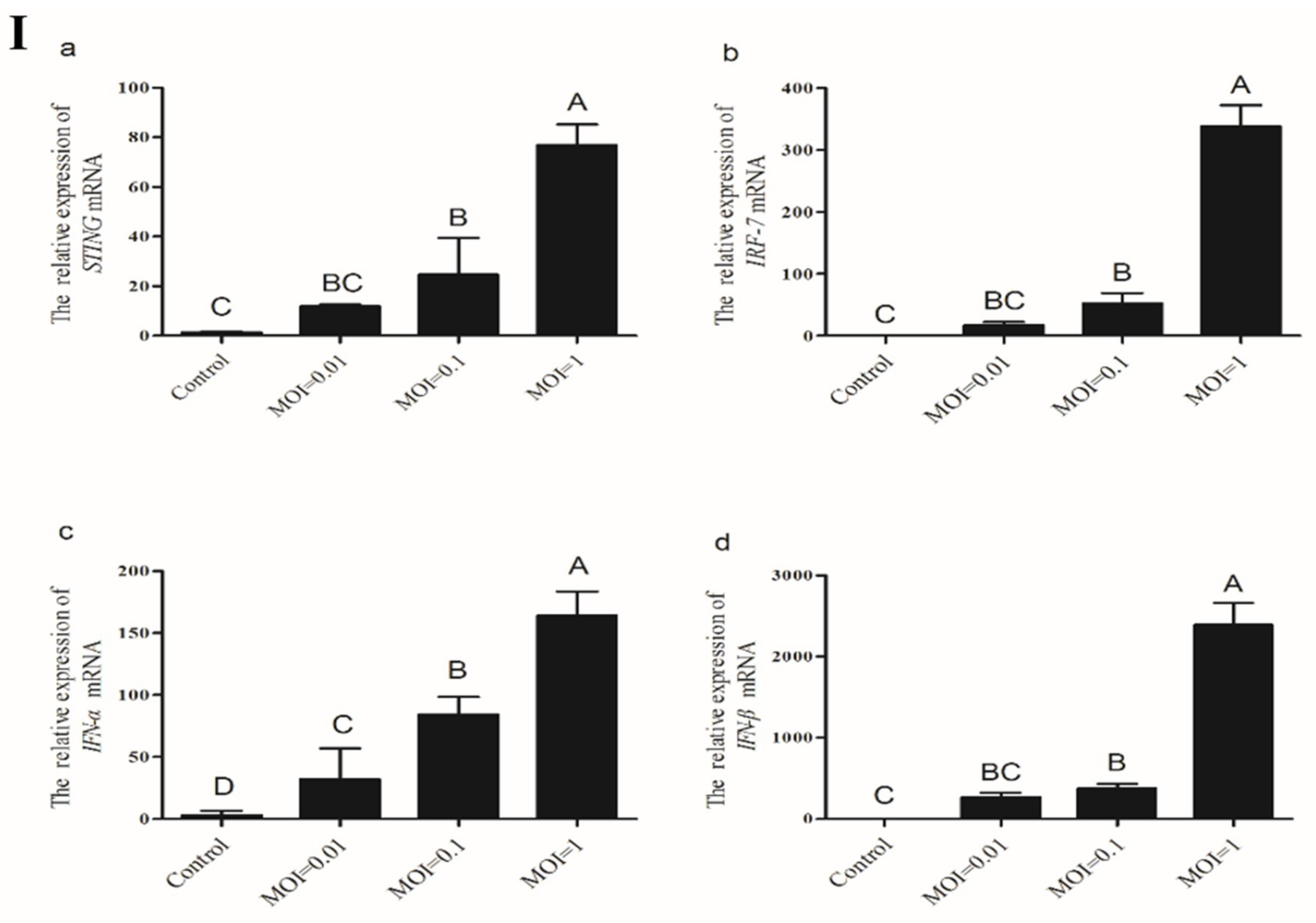
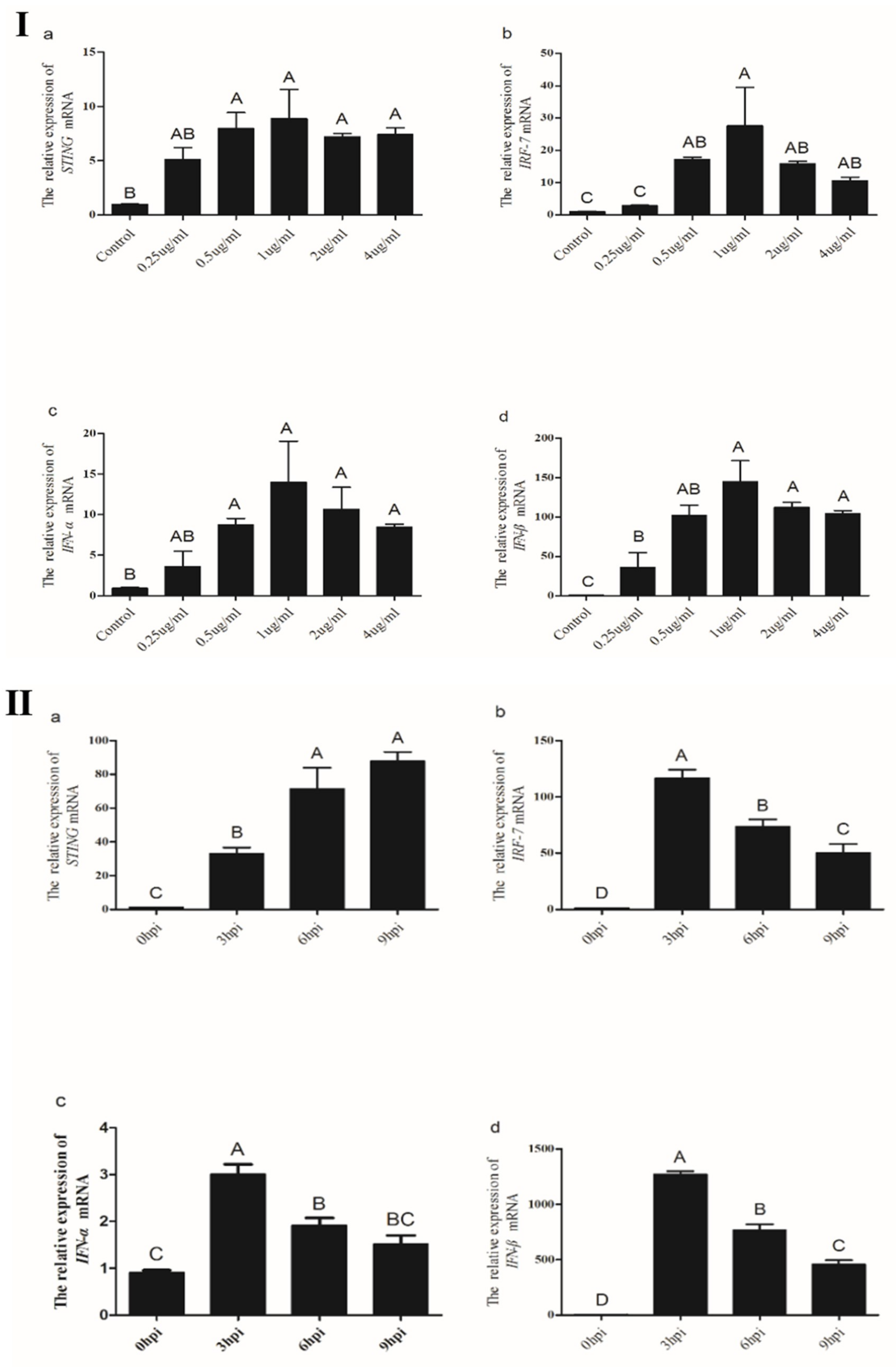
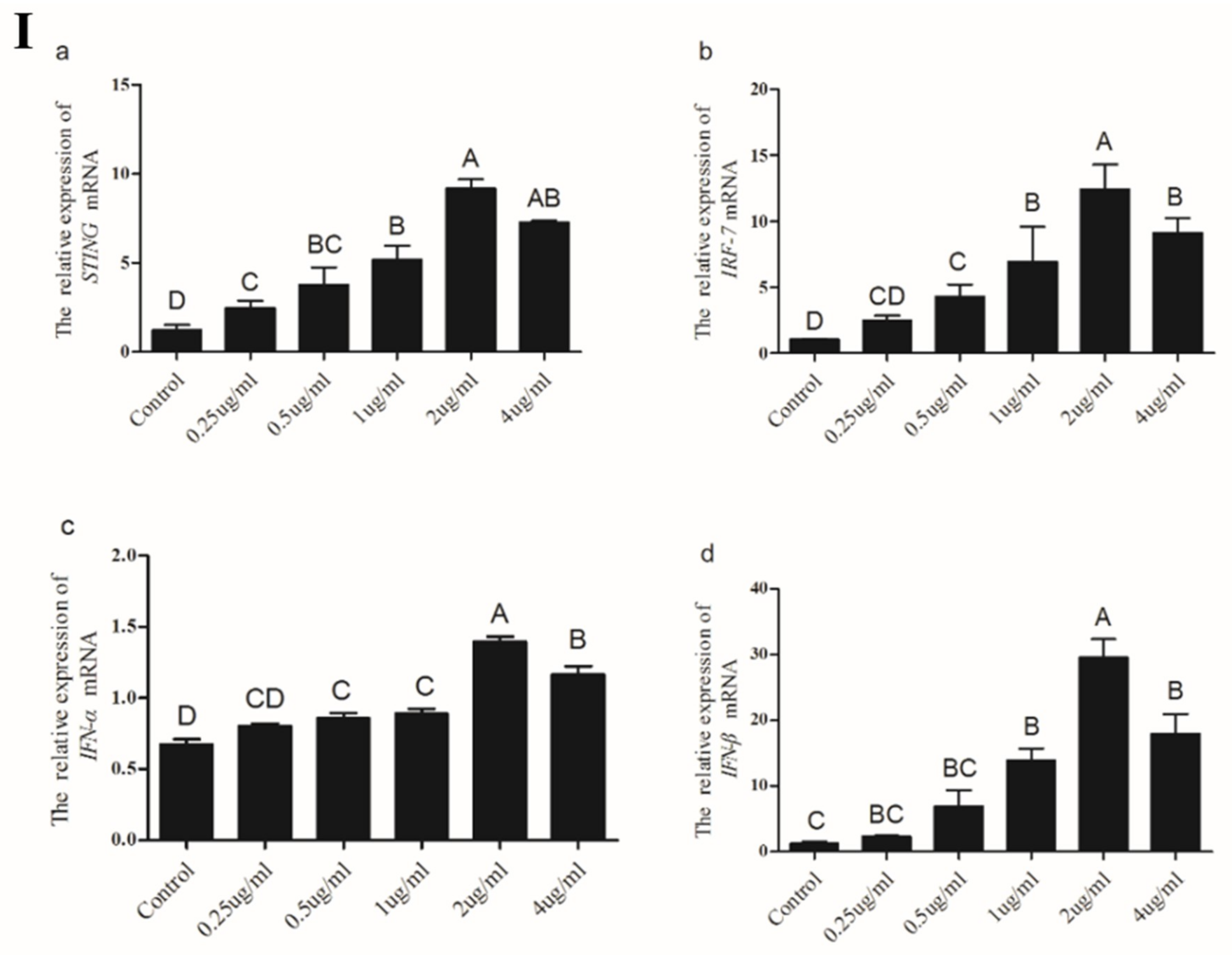
3.3.4. Induction of Chicken STING, IRF7, IFN-α and IFN-β in CEFs Transfected with Poly(I:C)
3.3.5. Induction of Chicken STING, IRF7, IFN-α and IFN-β in CEFs Transfected with Poly(dA:dT)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, S.; Song, X.; Wang, Y. Structural analysis of the STING adaptor protein reveals a hydrophobic dimer interface and mode of cyclic di-GMP binding. Immunity 2012, 36, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yang, Y. Innate immune recognition of viruses and viral vectors. Hum. Gene. Ther. 2009, 20, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 2004, 5, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Andrejeva, J.; Childs, K.S.; Young, D.F. The V proteins of paramyxoviruses bind the IFN-inducible RNA helicase, mda-5, and inhibit its activation of the IFN-β promoter. Proc. Natl. Acad. Sci. USA 2004, 101, 17264–17269. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Barber, G.N. STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 2008, 455, 674–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, B.; Yang, Y.; Li, S.; Wang, Y.Y.; Li, Y.; Diao, F.; Lei, C.; He, X.; Zhang, L.; Tien, P.; et al. The adaptor protein MITA links virus-sensing receptors to IRF3 transcription factor activation. Immunity 2008, 29, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Li, Y.; Chen, L. ERIS, an endoplasmic reticulum IFN stimulator, activates innate immune signaling through dimerization. Proc. Natl. Acad. Sci. USA 2009, 106, 8653–8658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, Y.; Shu, H.B.; Wang, Y.Y. MITA/STING: A central and multifaceted mediator in innate immune response. Cytokine Growth Factor Rev. 2014, 25, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, S.; Yu, H. ER Adaptor SCAP Translocates and Recruits IRF3 to Perinuclear Microsome Induced by Cytosolic Microbial DNAs. PLoS pathog. 2016, 12, e1005462. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; Paulson, M.; Holko, M.; Rice, C.M.; Williams, B.R.; Marié, I.; Levy, D.E. Induction of interferon-stimulated gene expression and antiviral responses require protein deacetylase activity. Proc. Natl. Acad. Sci. USA 2004, 101, 9578–9583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, J.; Corrigall, H.; Angus, N.J. Home versus hospital-based cardiac rehabilitation: A systematic review. Rural Remote Health 2011, 11, 1–17. [Google Scholar]
- Cheng, Y.; Sun, Y.; Wang, H. Chicken STING mediates activation of the IFN gene independently of the RIG-I. gene. J. Immunol. 2015, 195, 3922–3936. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Hill, K.K.; Filak, H. MPYS is required for IFN response factor 3 activation and type I IFN production in the response of cultured phagocytes to bacterial second messengers cyclic-di-AMP and cyclic-di-GMP. J. Immunol. 2011, 187, 2595–2601. [Google Scholar] [CrossRef] [PubMed]
- Diner, E.J.; Burdette, D.L.; Wilson, S.C. The innate immune DNA sensor cGAS produces a noncanonical cyclic dinucleotide that activates human STING. Cell Rep. 2013, 3, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.; Burdette, D.L.; Sharma, S. Mouse, but not human STING, binds and signals in response to the vascular disrupting agent 5, 6-dimethylxanthenone-4-acetic acid. J. Immunol. 2013, 190, 5216–5225. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Chen, X.; Zhang, K. Molecular cloning and functional characterization of Tibetan Porcine STING. Int. J. Mol. Sci. 2012, 13, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Chen, X.; Yu, B. Cloning and functional characterization of rat stimulator of interferon genes (STING) regulated by miR-24. Dev. Comp. Immunol. 2012, 37, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Cormican, P.; Lloyd, A.T.; Downing, T. The avian Toll-Like receptor pathway--subtle differences amidst general conformity. Dev. Comp. Immunol. 2009, 33, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Qi, Z.T.; Xu, Z. Global characterization of interferon regulatory factor (IRF) genes in vertebrates: Glimpse of the diversification in evolution. BMC Immunol. 2010, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J.; Decanini, E.L.; Afonso, C.L. Newcastle disease: Evolution of genotypes and the related diagnostic challenges. Infect. Gen. Evol. 2010, 10, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Kopecky-Bromberg, S.A.; Palese, P. Recombinant vectors as influenza vaccines. Curr. Top. Microbiol. Immunol. 2009, 333, 243–267. [Google Scholar] [PubMed]
- Hayashi, T.; Watanabe, C.; Suzuki, Y. Chicken MDA5 senses short double-stranded RNA with implications for antiviral response against avian influenza viruses in chicken. J. Immunol. 2014, 6, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Lorence, R.M.; Scot Roberts, M.; O’Neil, J.D. Phase 1 clinical experience using intravenous administration of PV701, an oncolytic Newcastle disease virus. Curr. Cancer Drug Targets 2007, 7, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Wehmann, E.; Herczeg, J.; Ballagi-Pordány, A. Rapid identification of Newcastle disease virus vaccine strains LaSota and B-1 by restriction site analysis of their matrix gene. Vaccine 1997, 15, 1430–1433. [Google Scholar] [CrossRef]
- Kato, H.; Takeuchi, O.; Sato, S. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Ma, Z.; Barber, G.N. STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 2009, 461, 788–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rue, C.A.; Susta, L.; Cornax, I. Virulent Newcastle disease virus elicits a strong innate immune response in chickens. J. Gen. Virol. 2011, 92, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Ablasser, A.; Bauernfeind, F.; Hartmann, G. RIG-I-dependent sensing of poly (dA: DT) through the induction of an RNA polymerase III–transcribed RNA intermediate. Nat. Immunol. 2009, 10, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lu, C.; Stewart, M.; Xu, H.; Strong, R.K.; Igumenova, T.; Li, P. Structural basis of double-stranded RNA recognition by the RIG-I. like receptor MDA5. Arch. Biochem. Biophys. 2009, 488, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Lopes, S.; Celhar, T.; Sans-Fons, G. The exonuclease Trex1 restrains macrophage proinflammatory activation. J. Immunol. 2013, 191, 6128–6135. [Google Scholar] [CrossRef] [PubMed]
- Karpala, A.J.; Stewart, C.; McKay, J. Characterization of chicken Mda5 activity: Regulation of IFN-β in the absence of RIG-I. functionality. J. Immunol. 2011, 186, 5397–5405. [Google Scholar] [CrossRef] [PubMed]
- Gitlin, L.; Barchet, W.; Gilfillan, S. Essential role of mda-5 in type I IFN responses to polyriboinosinic: Polyribocytidylic acid and encephalomyocarditis picornavirus. Proc. Natl. Acad. Sci. USA 2006, 103, 8459–8464. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhang, Y.B.; Liu, T.K. Fish MITA serves as a mediator for distinct fish IFN gene activation dependent on IRF3 or IRF7. J. Immunol. 2011, 187, 2531–2539. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | Annealing Temperature (°C) | Remarks |
|---|---|---|---|
| STING | F:GGACAGTCGCTGTGACCGAGGAT | 56.5 | Coding region |
| R:GCTTGTTGGACCAGTCCTGATG | amplification | ||
| 5′-RACE | TGCAGACAACTGTCGCAGTAGATC | 55 | 5′GSP |
| 3′-RACE | CCGCCTGGGTAGGAACAGTAGGC | 55 | 3′GSP |
| STING | F:GGACAGTCGCTGTGACCGAGGTT | 56.5 | Real-time PCR |
| R:GCTTGTTGGGCCAGTCCTGATG | |||
| IRF7 | F:TACACTGAGGACTTGCTGGAGGT | 58 | Real-time PCR |
| R:AAGATGGTGGTCTCCTGATCC | |||
| IFN-α | F:CAGGATGCCACCTTCTCTCAC | 57.6 | Real-time PCR |
| R: AGGATGGTGTCGTTGAAGGAG | |||
| IFN-β | F:CCTCAACCAGATCCAGCATTAC | 57.6 | Real-time PCR |
| R:CCCAGGTACAAGCACTGTAGTT | |||
| GAPDH | F: AGGACCAGGTTGTCTCCTGT | 57 | Real-time PCR |
| R: CCATCAAGTCCACAACACGG |
| Species | Uniprot Entry | Identity (%) |
|---|---|---|
| Gallus gallus | A0A1D5P7Q9 | 99.4 |
| Anas platyrhynchos | A0A071UC76 | 69.2 |
| Anser cygnoides domesticus | A0A053SKR3 | 67.0 |
| Nipponia nippon | A0A091UQ71 | 64.8 |
| Homo sapiens | A0A091FDC9 | 43.4 |
| Pan troglodytes | A0A087C4K5 | 43.7 |
| Sus scrofa | A0A077C6K8 | 43.5 |
| Mus musculus | A0A093SAR9 | 38.7 |
| Danio rerio | K4Q6R6 | 37.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ran, J.-S.; Jin, J.; Zhang, X.-X.; Wang, Y.; Ren, P.; Li, J.-J.; Yin, L.-Q.; Li, Z.-Q.; Lan, D.; Hu, Y.-D.; et al. Molecular Characterization, Expression and Functional Analysis of Chicken STING. Int. J. Mol. Sci. 2018, 19, 3706. https://doi.org/10.3390/ijms19123706
Ran J-S, Jin J, Zhang X-X, Wang Y, Ren P, Li J-J, Yin L-Q, Li Z-Q, Lan D, Hu Y-D, et al. Molecular Characterization, Expression and Functional Analysis of Chicken STING. International Journal of Molecular Sciences. 2018; 19(12):3706. https://doi.org/10.3390/ijms19123706
Chicago/Turabian StyleRan, Jin-Shan, Jie Jin, Xian-Xian Zhang, Ye Wang, Peng Ren, Jing-Jing Li, Ling-Qian Yin, Zhi-Qiang Li, Dan Lan, Yao-Dong Hu, and et al. 2018. "Molecular Characterization, Expression and Functional Analysis of Chicken STING" International Journal of Molecular Sciences 19, no. 12: 3706. https://doi.org/10.3390/ijms19123706