Administration of Downstream ApoE Attenuates the Adverse Effect of Brain ABCA1 Deficiency on Stroke

 and
and 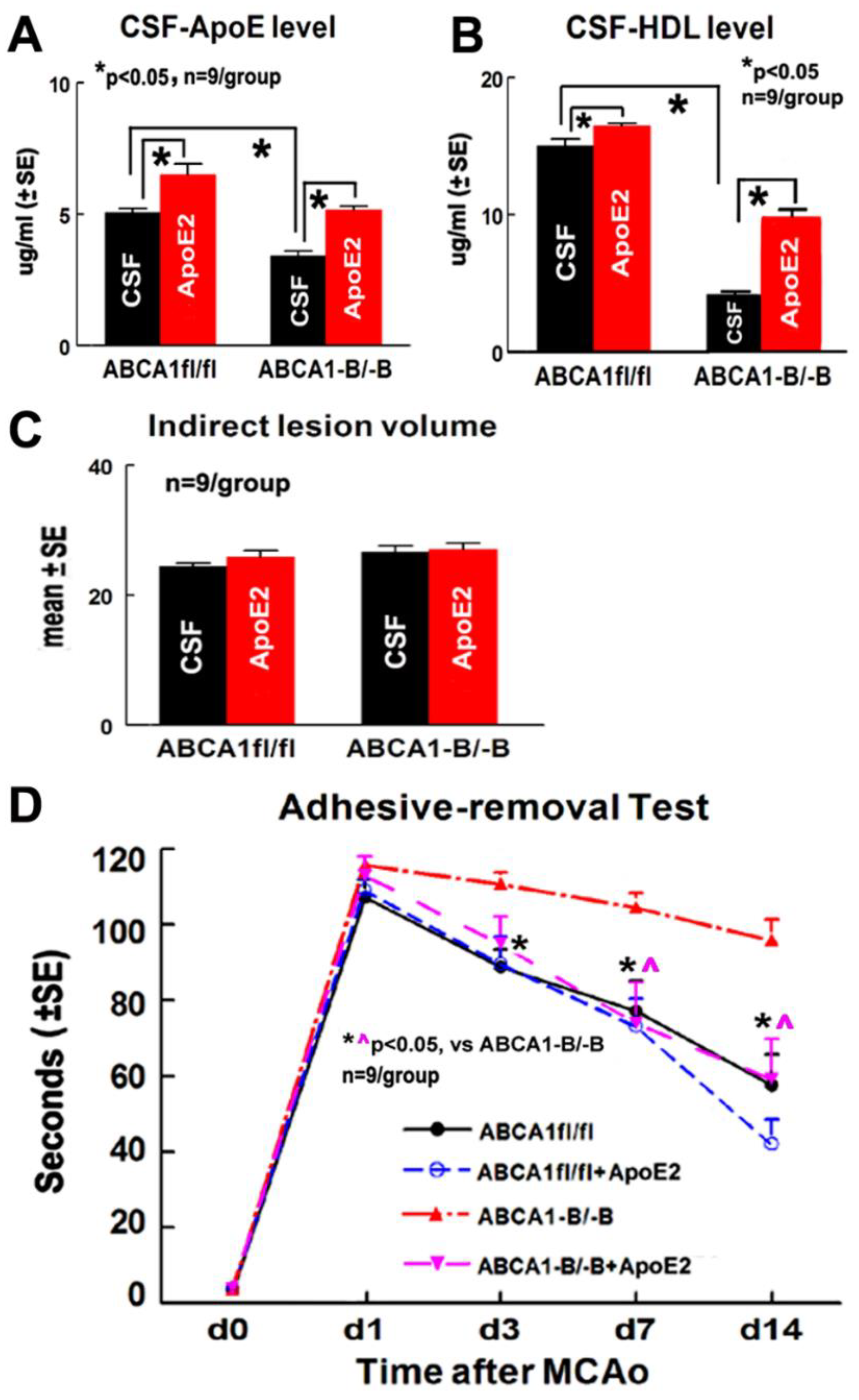{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. ApoE2 Increases ApoE/HDL Level in the CSF and Improves Functional Outcome in ABCA1−B/−B-Stroke Mice
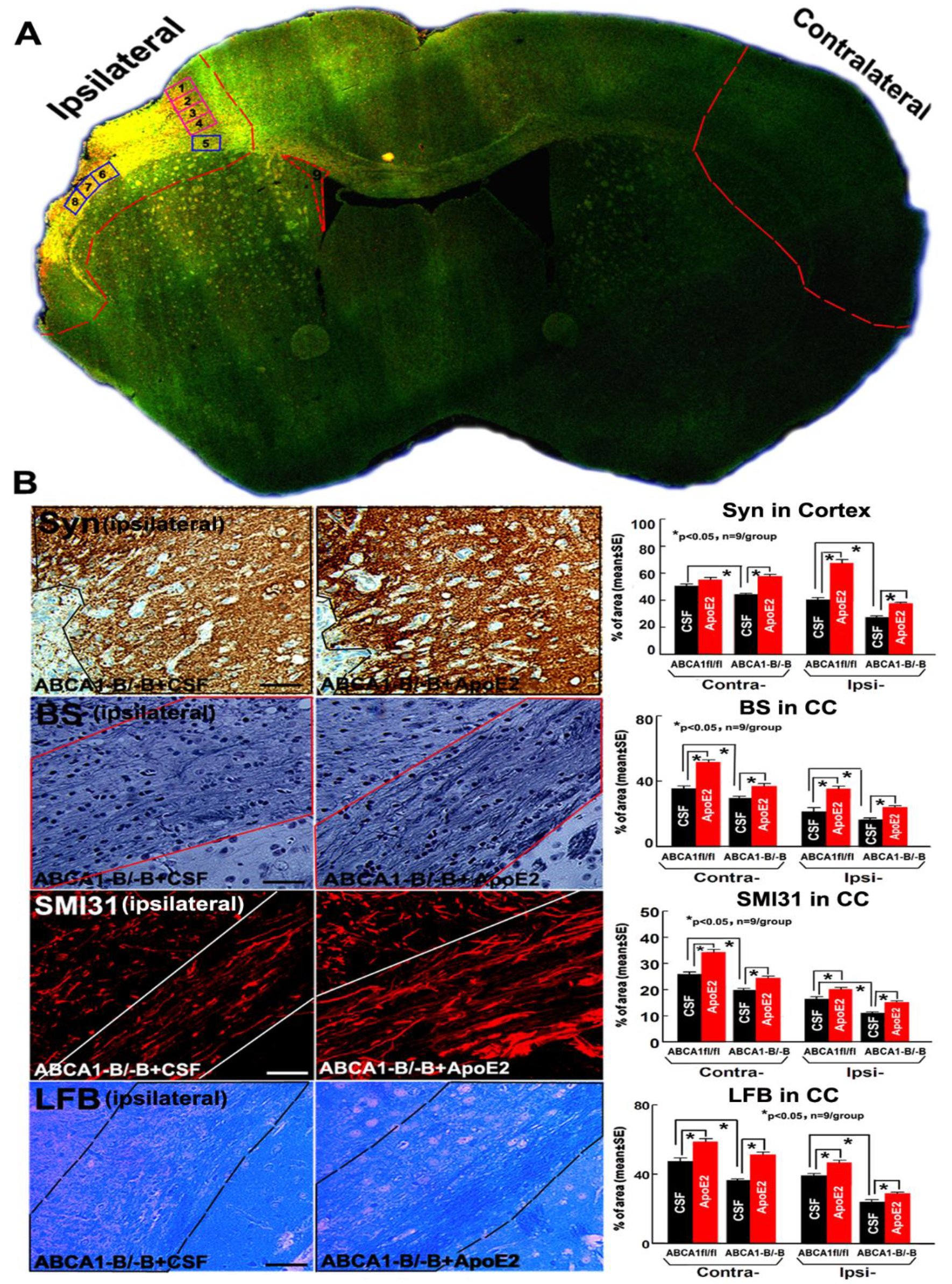
2.2. ApoE2 Increases GM and WM Densities in ABCA1−B/−B-Stroke Mice
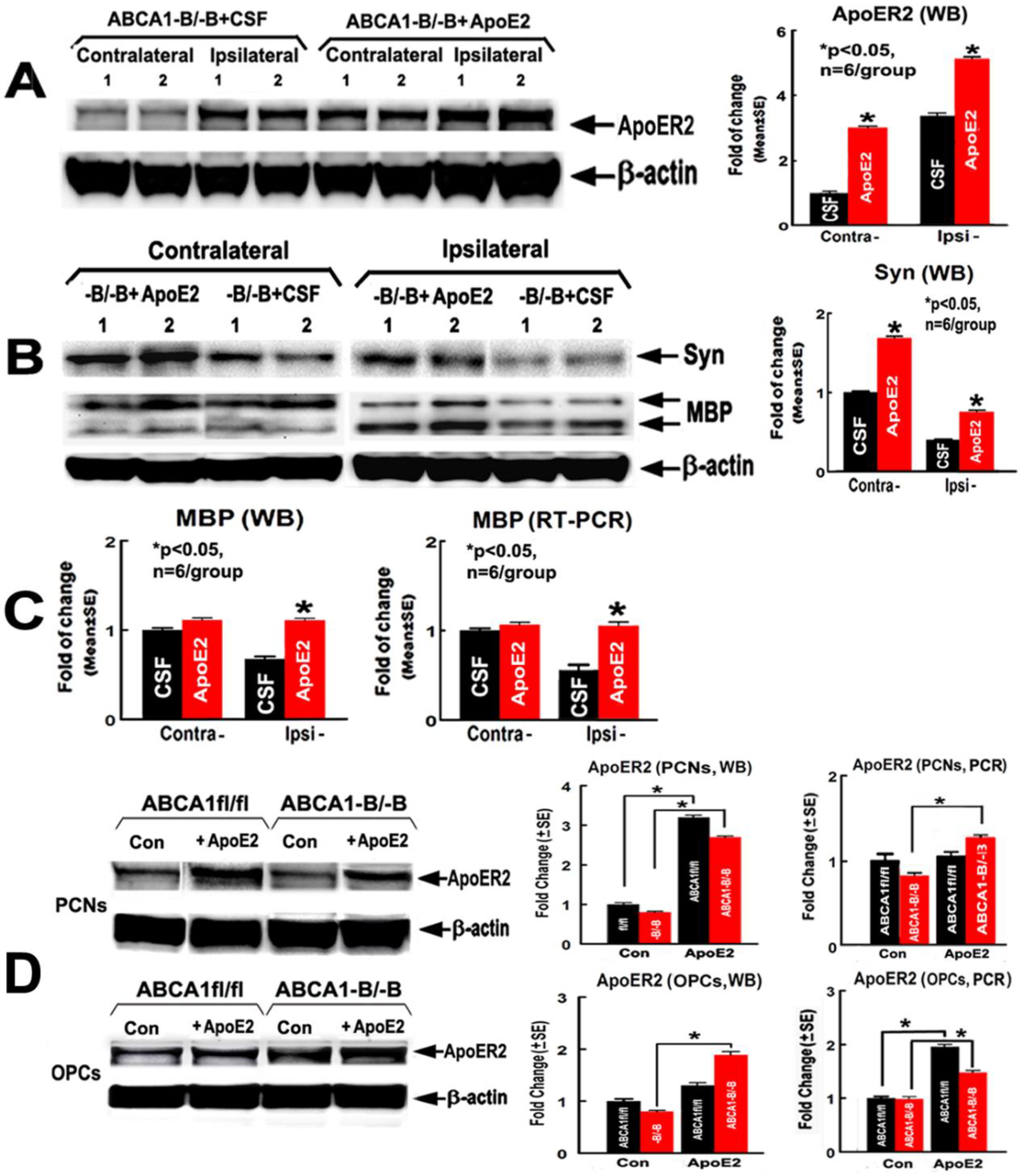
2.3. ApoE2 Increases ApoER2, Synaptic Protein and Myelin Protein Level in the Ischemic Brain
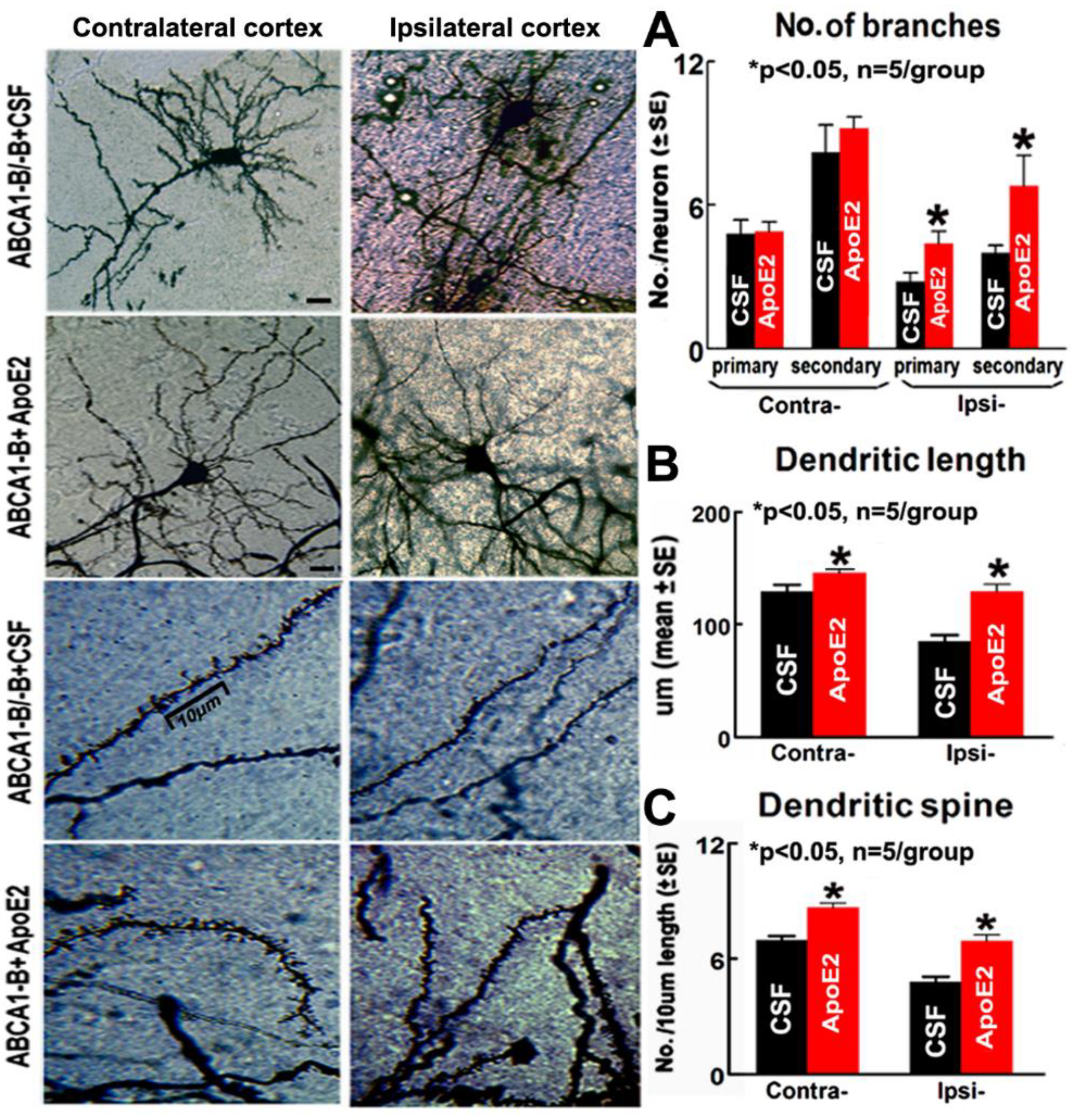
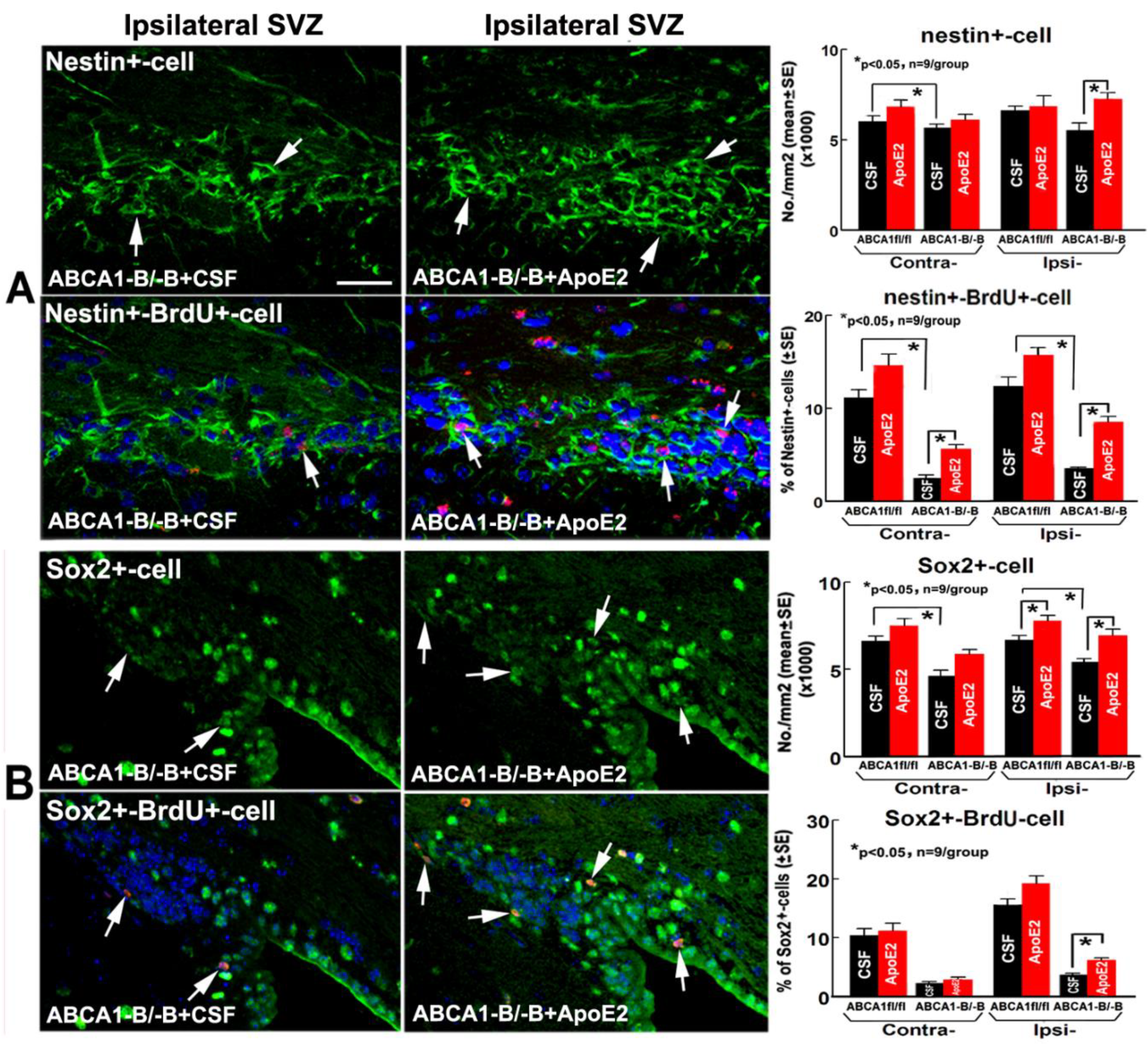
2.4. ApoE2 Enhances Neurogenesis in the Brain of ABCA1−B/−B Stroke Mice
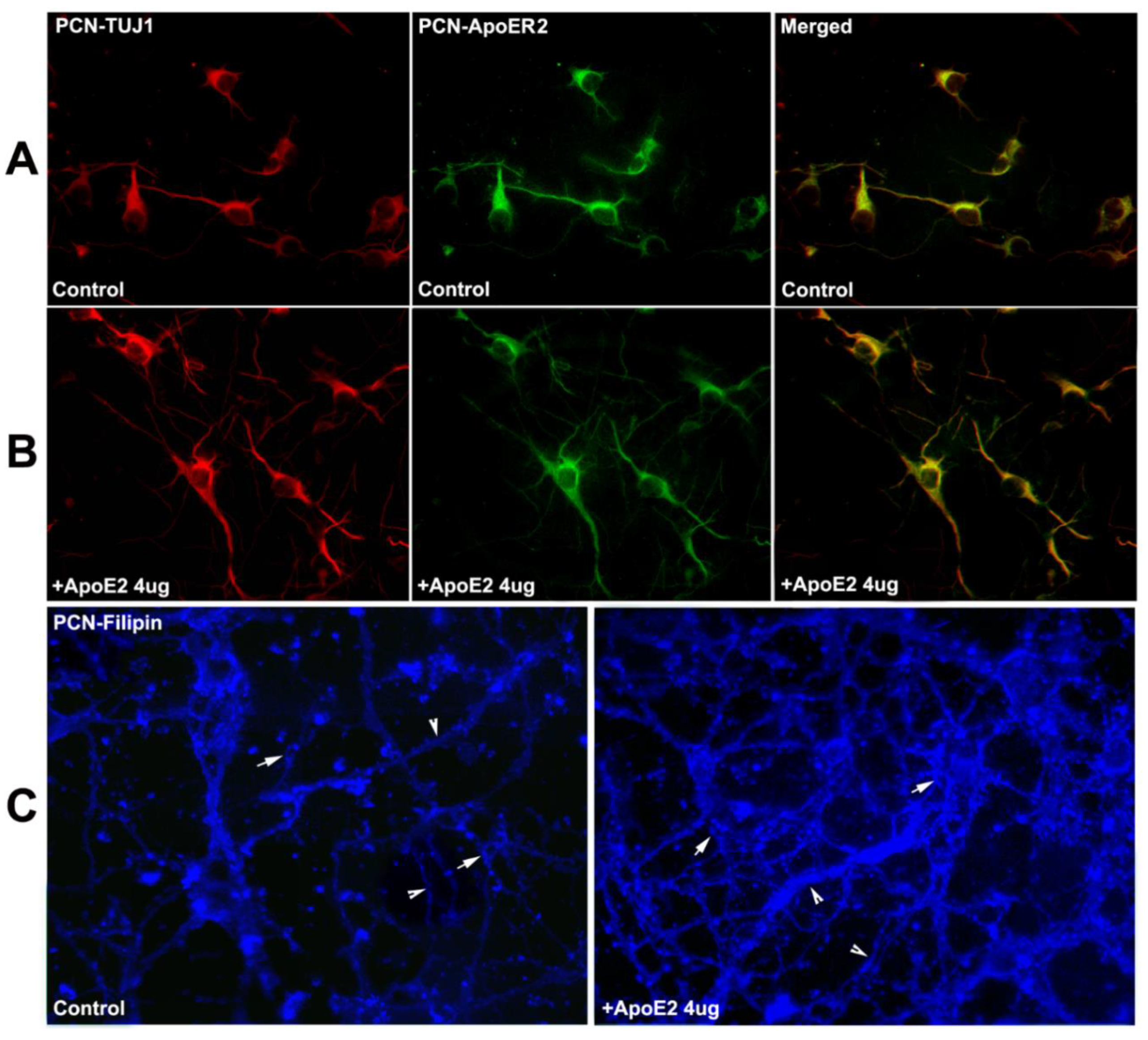
2.5. ApoER2 Co-Localizes in PCNs, and Supplementation with ApoE2 Increases Cholesterol Uptake in PCNs
2.6. ApoE2 Increases Neurite and Axonal Outgrowth in ABCA1−B/−B-PCNs
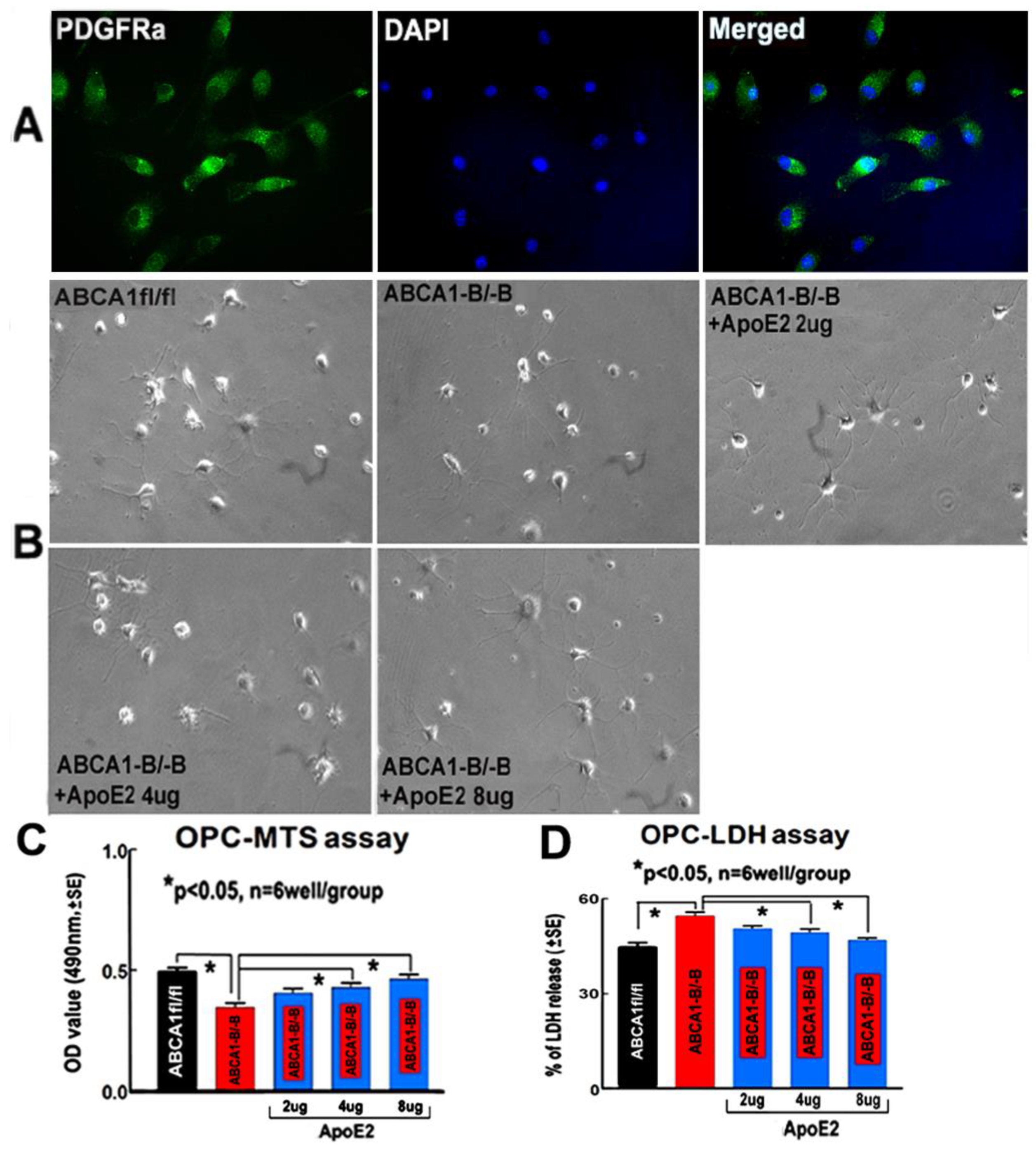
2.7. ApoE2 Increases Proliferation and Survival in ABCA1−B/−B-OPCs
3. Discussion
4. Materials and Methods
4.1. Animal Stroke Model
4.2. Experimental Groups
4.3. Functional Test
4.4. CSF Sampling and Laboratory Investigations
4.5. Lesion Volume Measurement
4.6. Histochemical and Immunohistochemical Staining Assessment
4.7. Image Acquisition and Quantification for Immunostaining Analysis
4.8. Real-Time RT-PCR and WB Assay
4.9. PCN Culture, ApoR2 Expression, and Cholesterol Distribution Analysis
4.10. PCN Neurite and Axonal Outgrowth Measurements
4.11. Primary OPC Culture and Purity Analysis
4.12. OPC Proliferation and Death Measurement
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fitzgerald, J.; Fawcett, J. Repair in the central nervous system. J. Bone Jt. Surg. 2007, 89, 1413–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westlye, L.T.; Reinvang, I.; Rootwelt, H.; Espeseth, T. Effects of APOE on brain white matter microstructure in healthy adults. Neurology 2012, 79, 1961–1969. [Google Scholar] [CrossRef] [PubMed]
- Crisby, M.; Bronge, L.; Wahlund, L.O. Low levels of high density lipoprotein increase the severity of cerebral white matter changes: Implications for prevention and treatment of cerebrovascular diseases. Curr. Alzheimer Res. 2010, 7, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Ward, M.A.; Bendlin, B.B.; McLaren, D.G.; Hess, T.M.; Gallagher, C.L.; Kastman, E.K.; Rowley, H.A.; Asthana, S.; Carlsson, C.M.; Sager, M.A.; et al. Low HDL Cholesterol is Associated with Lower Gray Matter Volume in Cognitively Healthy Adults. Front. Aging Neurosci. 2010, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Li, S.; Lv, S.H.; Piao, H.; Zhang, Y.H.; Zhang, Y.M.; Ma, H.; Zhang, J.; Sun, C.K.; Li, A.P. Hypoxia-ischemia brain damage disrupts brain cholesterol homeostasis in neonatal rats. Neuropediatrics 2009, 40, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Rust, S.; Rosier, M.; Funke, H.; Real, J.; Amoura, Z.; Piette, J.C.; Deleuze, J.F.; Brewer, H.B.; Duverger, N.; Denefle, P.; et al. Tangier disease is caused by mutations in the gene encoding ATP-binding cassette transporter 1. Nat. Genet. 1999, 22, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Dietschy, J.M.; Turley, S.D. Cholesterol metabolism in the brain. Curr. Opin. Lipidol. 2001, 12, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Woo, M.S.; Qin, L.; Ma, T.; Beltran, C.D.; Bao, Y.; Bailey, J.A.; Corbett, D.; Ratan, R.R.; Lahiri, D.K.; et al. Daidzein Augments Cholesterol Homeostasis via ApoE to Promote Functional Recovery in Chronic Stroke. J. Neurosci. 2015, 35, 15113–15126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danik, M.; Champagne, D.; Petit-Turcotte, C.; Beffert, U.; Poirier, J. Brain lipoprotein metabolism and its relation to neurodegenerative disease. Crit. Rev. Neurobiol. 1999, 13, 357–407. [Google Scholar] [CrossRef] [PubMed]
- Hirsch-Reinshagen, V.; Zhou, S.; Burgess, B.L.; Bernier, L.; McIsaac, S.A.; Chan, J.Y.; Tansley, G.H.; Cohn, J.S.; Hayden, M.R.; Wellington, C.L. Deficiency of ABCA1 impairs apolipoprotein E metabolism in brain. J. Biol. Chem. 2004, 279, 41197–41207. [Google Scholar] [CrossRef] [PubMed]
- Seitz, A.; Kragol, M.; Aglow, E.; Showe, L.; Heber-Katz, E. Apolipoprotein E expression after spinal cord injury in the mouse. J. Neurosci. Res. 2003, 71, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, M.; Crespo, P.; Berciano, M.T.; Andres, M.A.; Leon, J. Apolipoprotein E expression in the cerebellum of normal and hypercholesterolemic rabbits, Brain research. Mol. Brain Res. 1994, 21, 115–123. [Google Scholar] [CrossRef]
- Whitney, K.D.; Watson, M.A.; Collins, J.L.; Benson, W.G.; Stone, T.M.; Numerick, M.J.; Tippin, T.K.; Wilson, J.G.; Winegar, D.A.; Kliewer, S.A. Regulation of cholesterol homeostasis by the liver X receptors in the central nervous system. Mol. Endocrinol. (Baltim. MD) 2002, 16, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- Karasinska, J.M.; Rinninger, F.; Lutjohann, D.; Ruddle, P.; Franciosi, S.; Kruit, J.K.; Singaraja, R.R.; Hirsch-Reinshagen, V.; Fan, J.; Brunham, L.R.; et al. Specific loss of brain ABCA1 increases brain cholesterol uptake and influences neuronal structure and function. J. Neurosci. 2009, 29, 3579–3589. [Google Scholar] [CrossRef] [PubMed]
- Karasinska, J.M.; de Haan, W.; Franciosi, S.; Ruddle, P.; Fan, J.; Kruit, J.K.; Stukas, S.; Lutjohann, D.; Gutmann, D.H.; Wellington, C.L.; et al. ABCA1 influences neuroinflammation and neuronal death. Neurobiol. Dis. 2013, 54, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Chopp, M.; Zhang, Z.; Li, R.; Zacharek, A.; Landschoot-Ward, J.; Venkat, P.; Chen, J. ABCA1/ApoE/HDL Pathway Mediates GW3965-Induced Neurorestoration after Stroke. Stroke 2017, 48, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Chopp, M.; Zacharek, A.; Karasinska, J.M.; Cui, Y.; Ning, R.; Zhang, Y.; Wang, Y.; Chen, J. Deficiency of brain ATP-binding cassette transporter A-1 exacerbates blood-brain barrier and white matter damage after stroke. Stroke 2015, 46, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Arold, S.; Sullivan, P.; Bilousova, T.; Teng, E.; Miller, C.A.; Poon, W.W.; Vinters, H.V.; Cornwell, L.B.; Saing, T.; Cole, G.M.; et al. Apolipoprotein E level and cholesterol are associated with reduced synaptic amyloid beta in Alzheimer’s disease and apoE TR mouse cortex. Acta Neuropathol. 2012, 123, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.M.; Jessberger, S. Adult neurogenesis and its role in neuropsychiatric disease, brain repair and normal brain function. Neuropathol. Appl. Neurobiol. 2014, 40, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Luong, M.; Pooley, A.; Nathan, B.P. Isoform-specific effects of apoE on neurite outgrowth in olfactory epithelium culture. J. Biomed. Sci. 2013, 20, 49. [Google Scholar] [CrossRef] [PubMed]
- Nathan, B.P.; Barsukova, A.G.; Shen, F.; McAsey, M.; Struble, R.G. Estrogen facilitates neurite extension via apolipoprotein E in cultured adult mouse cortical neurons. Endocrinology 2004, 145, 3065–3073. [Google Scholar] [CrossRef] [PubMed]
- Neely, M.D.; Montine, T.J. CSF lipoproteins and Alzheimer’s disease. J. Nutr. Health Aging 2002, 6, 383–391. [Google Scholar] [PubMed]
- Dumanis, S.B.; Cha, H.J.; Song, J.M.; Trotter, J.H.; Spitzer, M.; Lee, J.Y.; Weeber, E.J.; Turner, R.S.; Pak, D.T.; Rebeck, G.W.; et al. ApoE receptor 2 regulates synapse and dendritic spine formation. PLoS ONE 2011, 6, e17203. [Google Scholar] [CrossRef] [PubMed]
- Han, C.W.; Lee, K.H.; Noh, M.G.; Kim, J.M.; Kim, H.S.; Kim, H.S.; Kim, R.G.; Cho, J.; Kim, H.I.; Lee, M.C. An Experimental Infarct Targeting the Internal Capsule: Histopathological and Ultrastructural Changes. J. Pathol. Transl. Med. 2017, 51, 292–305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chopp, M. Neural Stem Cells and Ischemic Brain. J. Stroke 2016, 18, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Champagne, D.; Rochford, J.; Poirier, J. Effect of apolipoprotein E deficiency on reactive sprouting in the dentate gyrus of the hippocampus following entorhinal cortex lesion: Role of the astroglial response. Exp. Neurol. 2005, 194, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Weisgraber, K.H. Apolipoprotein E: Structure-function relationships. Adv. Protein Chem. 1994, 45, 249–302. [Google Scholar] [PubMed]
- Horsburgh, K.; McCulloch, J.; Nilsen, M.; McCracken, E.; Large, C.; Roses, A.D.; Nicoll, J.A. Intraventricular infusion of apolipoprotein E ameliorates acute neuronal damage after global cerebral ischemia in mice. J. Cereb. Blood Flow Metab. 2000, 20, 458–462. [Google Scholar] [CrossRef] [PubMed]
- DeMattos, R.B.; Brendza, R.P.; Heuser, J.E.; Kierson, M.; Cirrito, J.R.; Fryer, J.; Sullivan, P.M.; Fagan, A.M.; Han, X.; Holtzman, D.M. Purification and characterization of astrocyte-secreted apolipoprotein E and J-containing lipoproteins from wild-type and human apoE transgenic mice. Neurochem. Int. 2001, 39, 415–425. [Google Scholar] [CrossRef]
- Ito, J.; Zhang, L.Y.; Asai, M.; Yokoyama, S. Differential generation of high-density lipoprotein by endogenous and exogenous apolipoproteins in cultured fetal rat astrocytes. J. Neurochem. 1999, 72, 2362–2369. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, X.; Kusumo, H.; Costa, L.G.; Guizzetti, M. Cholesterol efflux is differentially regulated in neurons and astrocytes: Implications for brain cholesterol homeostasis. Biochim. Biophys. Acta 2013, 1831, 263–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietschy, J.M.; Turley, S.D. Thematic review series: Brain Lipids. Cholesterol metabolism in the central nervous system during early development and in the mature animal. J. Lipid Res. 2004, 45, 1375–1397. [Google Scholar] [CrossRef] [PubMed]
- Mauch, D.H.; Nagler, K.; Schumacher, S.; Goritz, C.; Muller, E.C.; Otto, A.; Pfrieger, F.W. CNS synaptogenesis promoted by glia-derived cholesterol. Science 2001, 294, 1354–1357. [Google Scholar] [CrossRef] [PubMed]
- Donkin, J.J.; Stukas, S.; Hirsch-Reinshagen, V.; Namjoshi, D.; Wilkinson, A.; May, S.; Chan, J.; Fan, J.; Collins, J.; Wellington, C.L. ATP-binding cassette transporter A1 mediates the beneficial effects of the liver X receptor agonist GW3965 on object recognition memory and amyloid burden in amyloid precursor protein/presenilin 1 mice. J. Biol. Chem. 2010, 285, 34144–34154. [Google Scholar] [CrossRef] [PubMed]
- Brooks-Wilson, A.; Marcil, M.; Clee, S.M.; Zhang, L.H.; Roomp, K.; van Dam, M.; Yu, L.; Brewer, C.; Collins, J.A.; Molhuizen, H.O.; et al. Mutations in ABC1 in Tangier disease and familial high-density lipoprotein deficiency. Nat. Genet. 1999, 22, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Pfrieger, F.W.; Ungerer, N. Cholesterol metabolism in neurons and astrocytes. Prog. Lipid Res. 2011, 50, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Wahrle, S.E.; Jiang, H.; Parsadanian, M.; Legleiter, J.; Han, X.; Fryer, J.D.; Kowalewski, T.; Holtzman, D.M. ABCA1 is required for normal central nervous system ApoE levels and for lipidation of astrocyte-secreted apoE. J. Biol. Chem. 2004, 279, 40987–40993. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Campenot, R.B.; Vance, D.E.; Vance, J.E. Apolipoprotein E-containing lipoproteins protect neurons from apoptosis via a signaling pathway involving low-density lipoprotein receptor-related protein-1. J. Neurosci. 2007, 27, 1933–1941. [Google Scholar] [CrossRef] [PubMed]
- Valastro, B.; Ghribi, O.; Poirier, J.; Krzywkowski, P.; Massicotte, G. AMPA receptor regulation and LTP in the hippocampus of young and aged apolipoprotein E-deficient mice. Neurobiol. Aging 2001, 22, 9–15. [Google Scholar] [CrossRef]
- Llovera, G.; Roth, S.; Plesnila, N.; Veltkamp, R.; Liesz, A. Modeling stroke in mice: Permanent coagulation of the distal middle cerebral artery. J. Vis. Exp. JoVE 2014, e51729. [Google Scholar] [CrossRef] [PubMed]
- Pitas, R.E.; Boyles, J.K.; Lee, S.H.; Hui, D.; Weisgraber, K.H. Lipoproteins and their receptors in the central nervous system. Characterization of the lipoproteins in cerebrospinal fluid and identification of apolipoprotein B,E(LDL) receptors in the brain. J. Biol. Chem. 1987, 262, 14352–14360. [Google Scholar] [PubMed]
- Wahrle, S.E.; Shah, A.R.; Fagan, A.M.; Smemo, S.; Kauwe, J.S.; Grupe, A.; Hinrichs, A.; Mayo, K.; Jiang, H.; Thal, L.J.; et al. Apolipoprotein E levels in cerebrospinal fluid and the effects of ABCA1 polymorphisms. Mol. Neurodegener. 2007, 2, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freret, T.; Bouet, V.; Leconte, C.; Roussel, S.; Chazalviel, L.; Divoux, D.; Schumann-Bard, P.; Boulouard, M. Behavioral deficits after distal focal cerebral ischemia in mice: Usefulness of adhesive removal test. Behav. Neurosci. 2009, 123, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Swanson, R.A.; Morton, M.T.; Tsao-Wu, G.; Savalos, R.A.; Davidson, C.; Sharp, F.R. A semiautomated method for measuring brain infarct volume. J. Cereb. Blood Flow Metab. 1990, 10, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chopp, M.; Zhang, Y.; Xiong, Y.; Li, C.; Sadry, N.; Rhaleb, I.; Lu, M.; Zhang, Z.G. Diabetes Mellitus Impairs Cognitive Function in Middle-Aged Rats and Neurological Recovery in Middle-Aged Rats After Stroke. Stroke 2016, 47, 2112–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkat, P.; Chopp, M.; Zacharek, A.; Cui, C.; Zhang, L.; Li, Q.; Lu, M.; Zhang, T.; Liu, A.; Chen, J. White matter damage and glymphatic dysfunction in a model of vascular dementia in rats with no prior vascular pathologies. Neurobiol. Aging 2017, 50, 96–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wislet-Gendebien, S.; Hans, G.; Leprince, P.; Rigo, J.M.; Moonen, G.; Rogister, B. Plasticity of cultured mesenchymal stem cells: Switch from nestin-positive to excitable neuron-like phenotype. Stem Cells (Dayton OH) 2005, 23, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.L.; Chopp, M.; Roberts, C.; Jia, L.; Wei, M.; Lu, M.; Wang, X.; Pourabdollah, S.; Zhang, Z.G. Ascl1 lineage cells contribute to ischemia-induced neurogenesis and oligodendrogenesis. J. Cereb. Blood Flow Metab. 2011, 31, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Chopp, M.; Zacharek, A.; Cui, C.; Yan, T.; Ning, R.; Chen, J. D-4F Decreases White Matter Damage After Stroke in Mice. Stroke 2016, 47, 214–220. [Google Scholar] [CrossRef] [PubMed]
- O’Meara, R.W.; Cummings, S.E.; Michalski, J.P.; Kothary, R. A new in vitro mouse oligodendrocyte precursor cell migration assay reveals a role for integrin-linked kinase in cell motility. BMC Neurosci. 2016, 17, 7. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Li, R.; Zacharek, A.; Landschoot-Ward, J.; Wang, F.; Wu, K.-H.H.; Chopp, M.; Chen, J.; Cui, X. Administration of Downstream ApoE Attenuates the Adverse Effect of Brain ABCA1 Deficiency on Stroke. Int. J. Mol. Sci. 2018, 19, 3368. https://doi.org/10.3390/ijms19113368
Wang X, Li R, Zacharek A, Landschoot-Ward J, Wang F, Wu K-HH, Chopp M, Chen J, Cui X. Administration of Downstream ApoE Attenuates the Adverse Effect of Brain ABCA1 Deficiency on Stroke. International Journal of Molecular Sciences. 2018; 19(11):3368. https://doi.org/10.3390/ijms19113368
Chicago/Turabian StyleWang, Xiaohui, Rongwen Li, Alex Zacharek, Julie Landschoot-Ward, Fengjie Wang, Kuan-Han Hank Wu, Michael Chopp, Jieli Chen, and Xu Cui. 2018. "Administration of Downstream ApoE Attenuates the Adverse Effect of Brain ABCA1 Deficiency on Stroke" International Journal of Molecular Sciences 19, no. 11: 3368. https://doi.org/10.3390/ijms19113368