Genome-wide Identification, Classification, Molecular Evolution and Expression Analysis of Malate Dehydrogenases in Apple


Abstract
:1. Introduction
2. Results
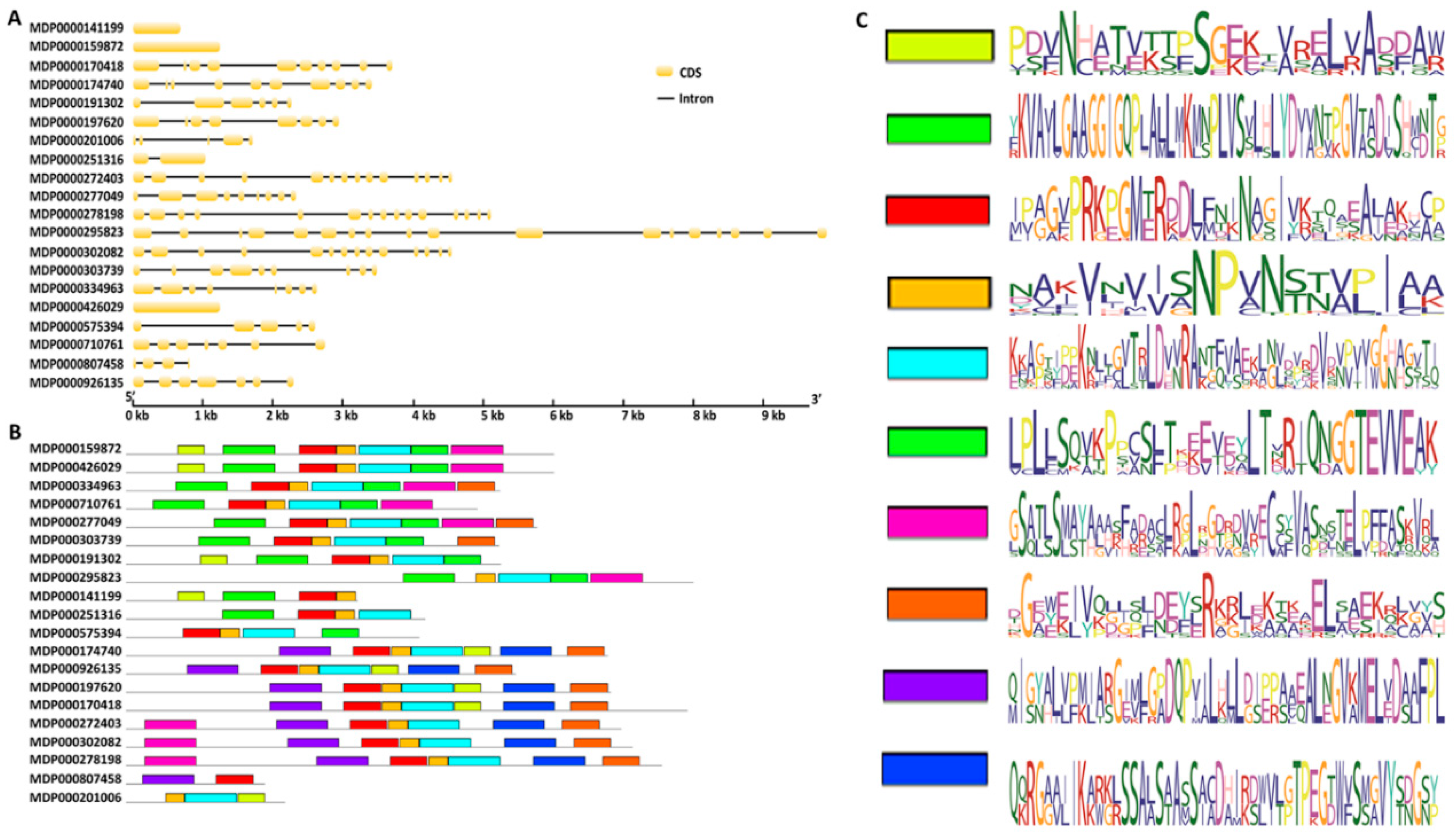
2.1. Identification and Characterization of 20 MDH Genes in Apple
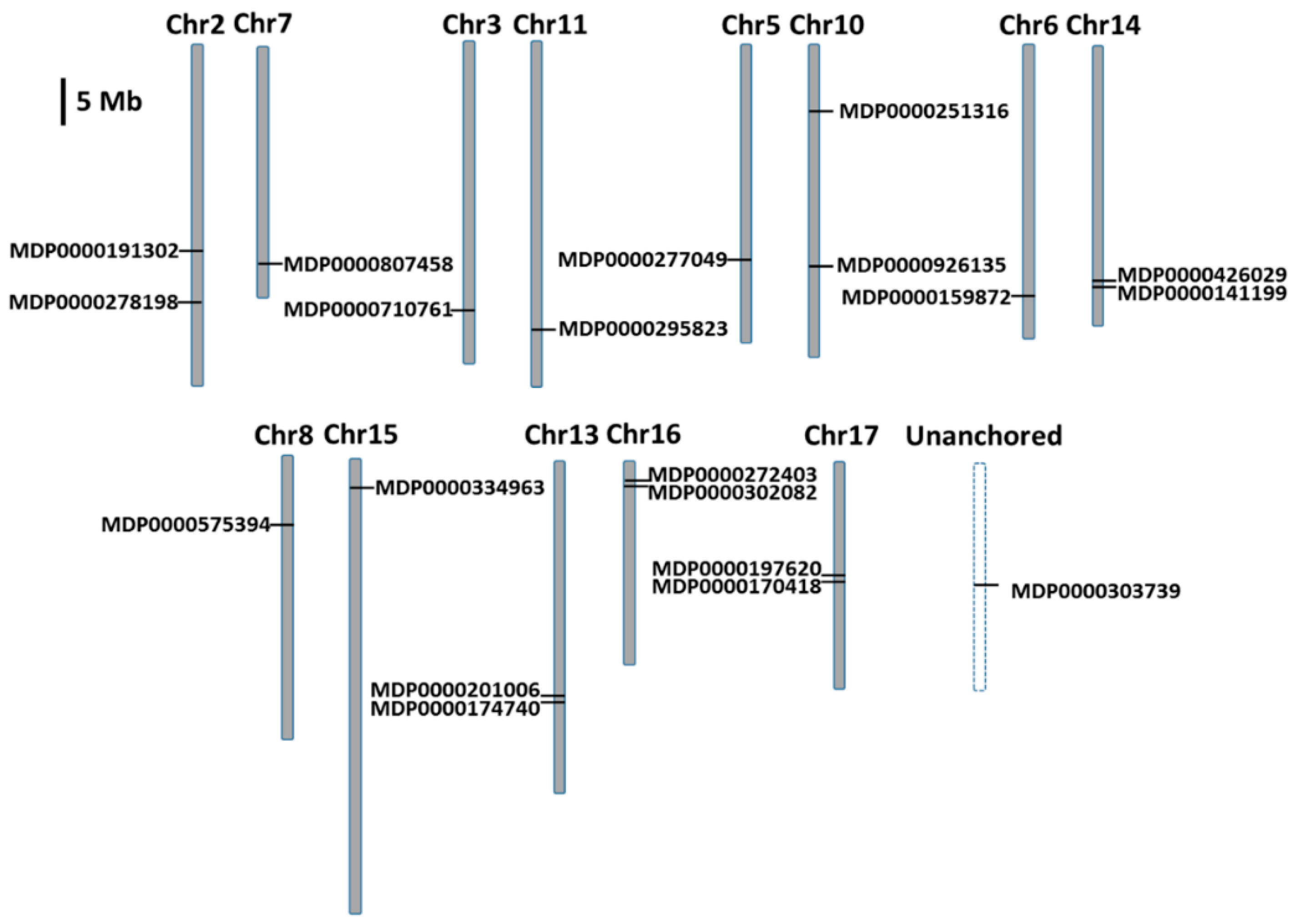
2.2. Chromosomal Localization and Duplication of MdMDH Genes
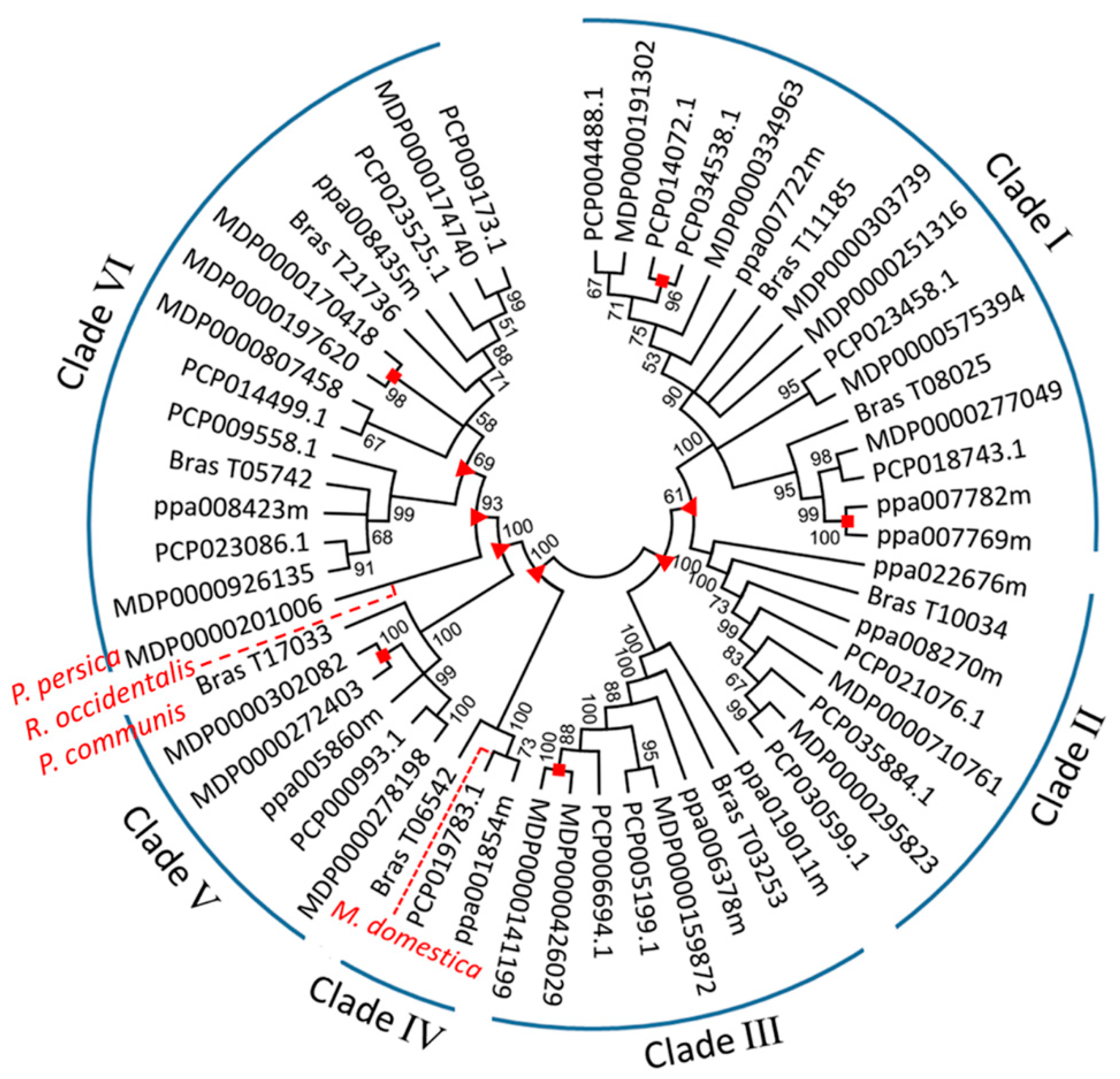
2.3. Phylogenetic Analysis, Gene Duplication and Loss of MDH Genes in Four Rosaceae Species
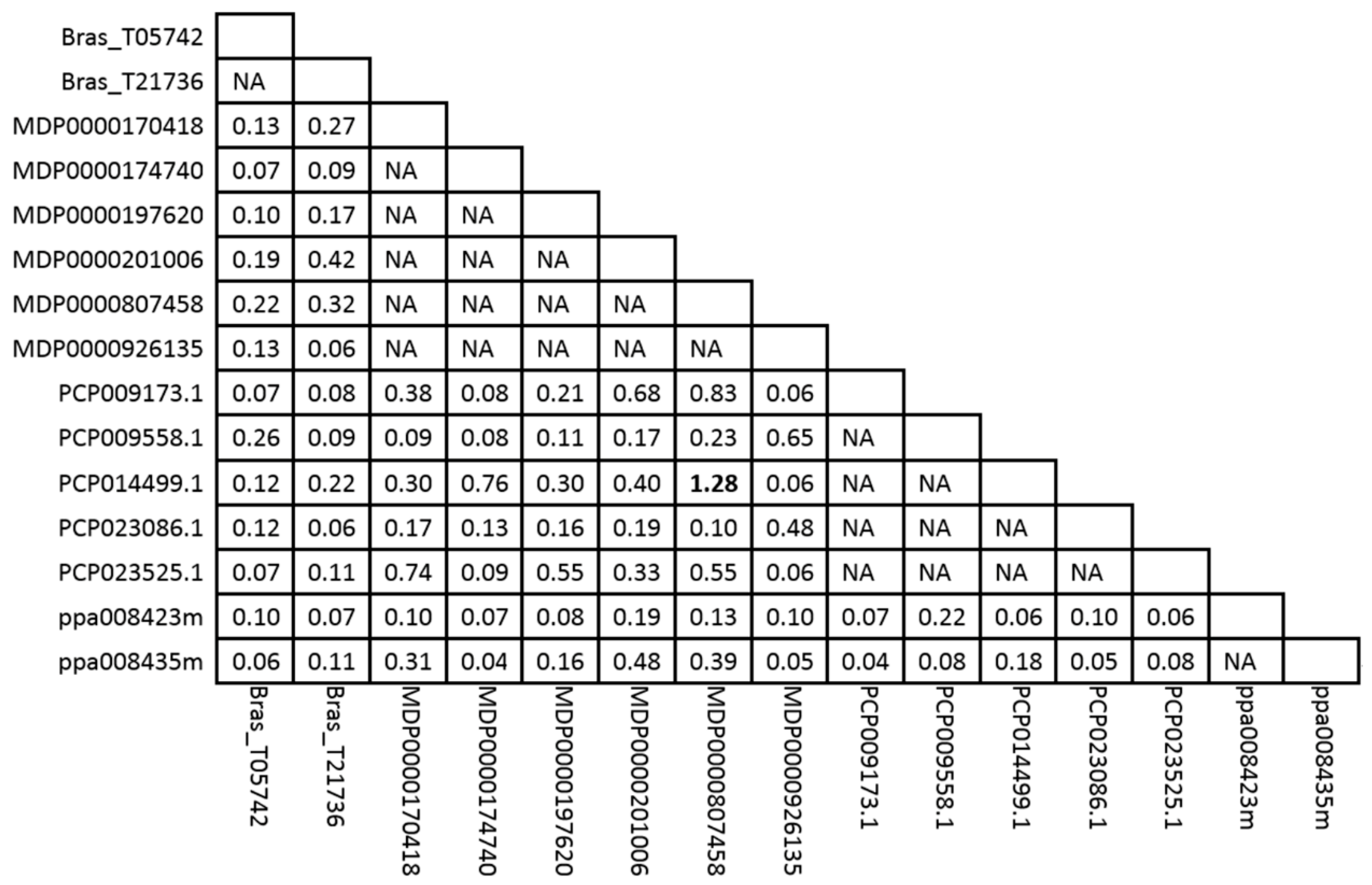
2.4. Estimation of the Positive Selection of MDH Genes in Four Rosaceae Species
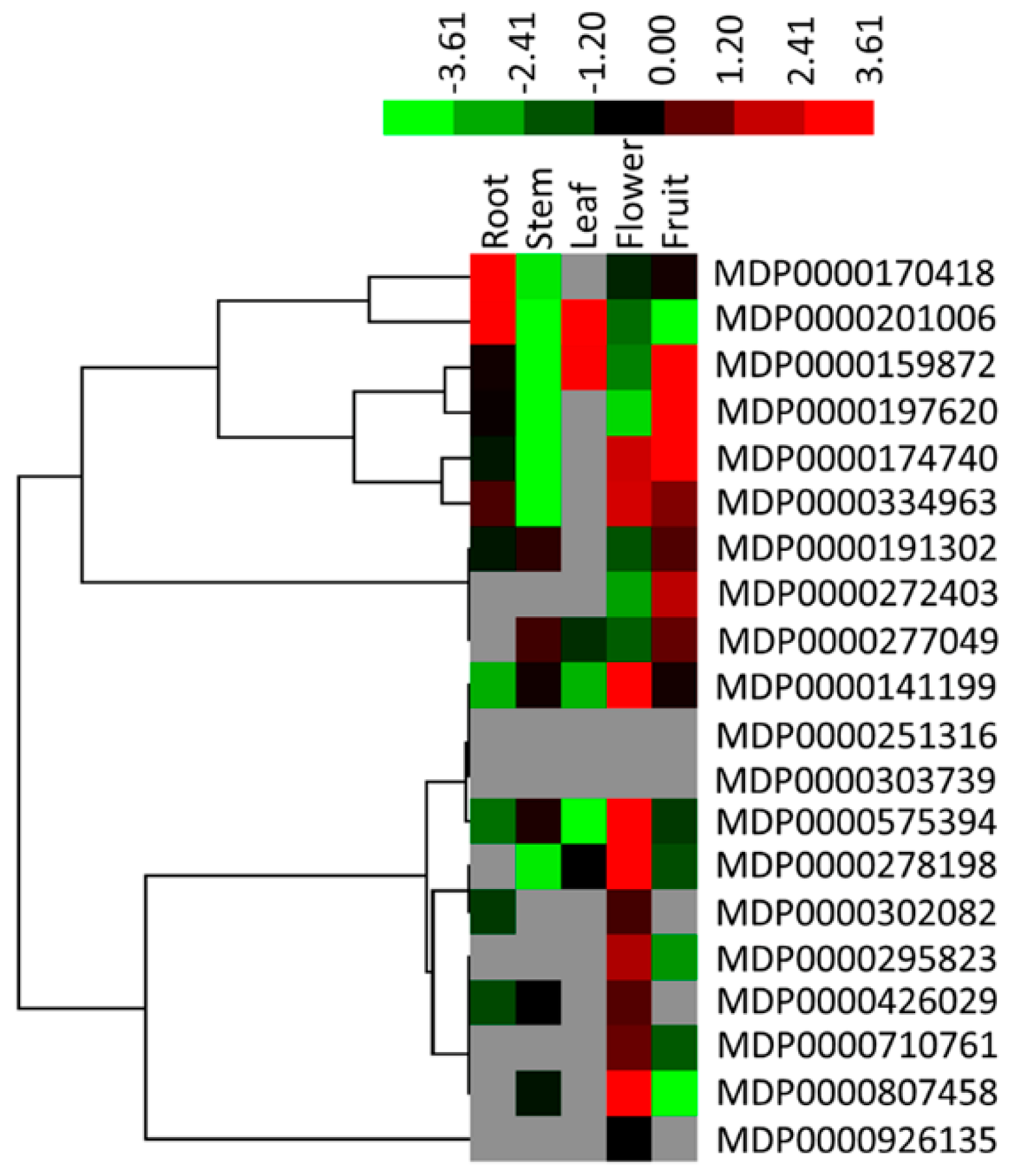
2.5. Expression Analysis of the MdMDH Genes in Different Tissues of Apple Plants
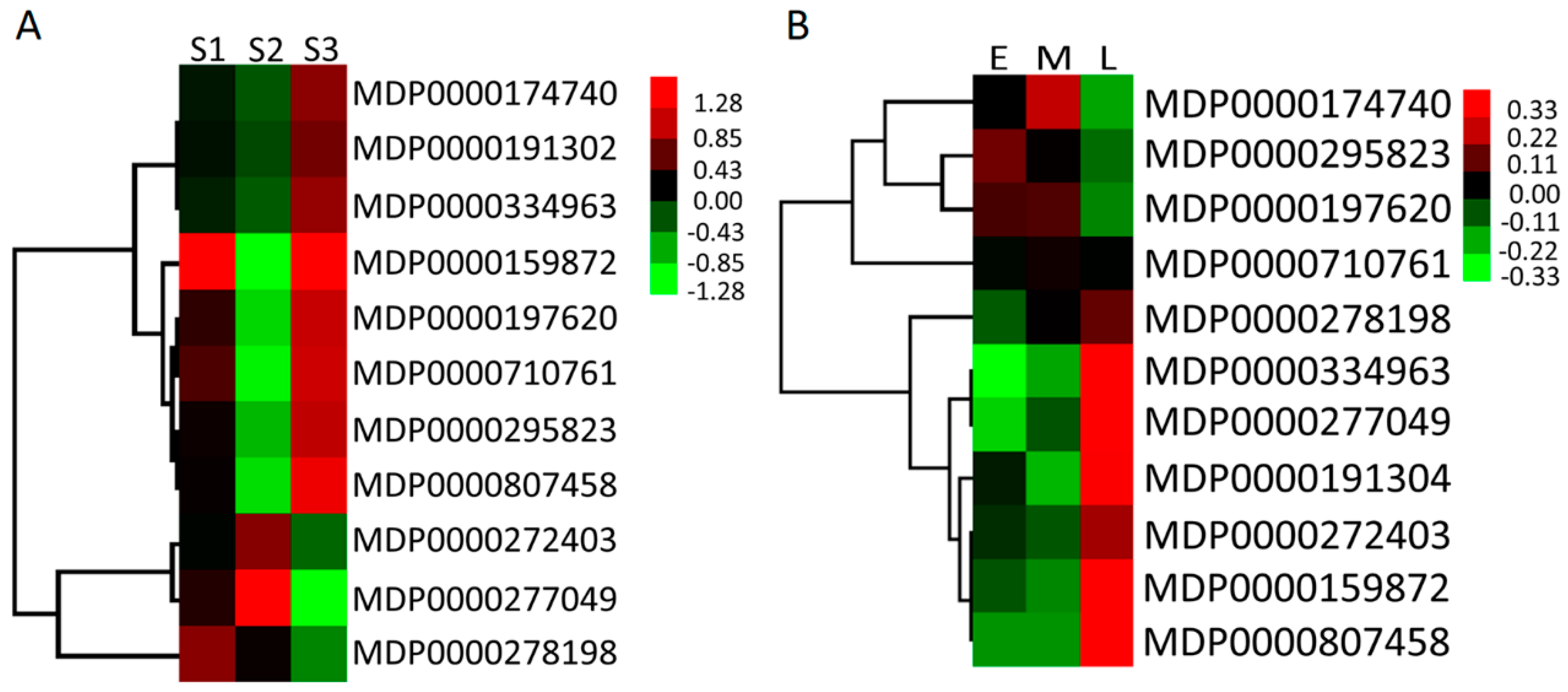
2.6. Expression Profiles of MdMDH Genes During Apple Flower Bud Differentiation and in Fruit at Different Developmental Stages
3. Discussion
4. Materials and Methods
4.1. Identification of MDHs and Phylogenetic Analyses
4.2. Gene Structure and Conserved Motif Analysis of MDH Genes
4.3. Estimations of Gene Duplication and Gene Loss
4.4. Positive Selection Analysis
4.5. RNA Isolation and Quantitative RT-PCR (qRT-PCR) Analysis
4.6. Plant Material and Transcriptome Analysis
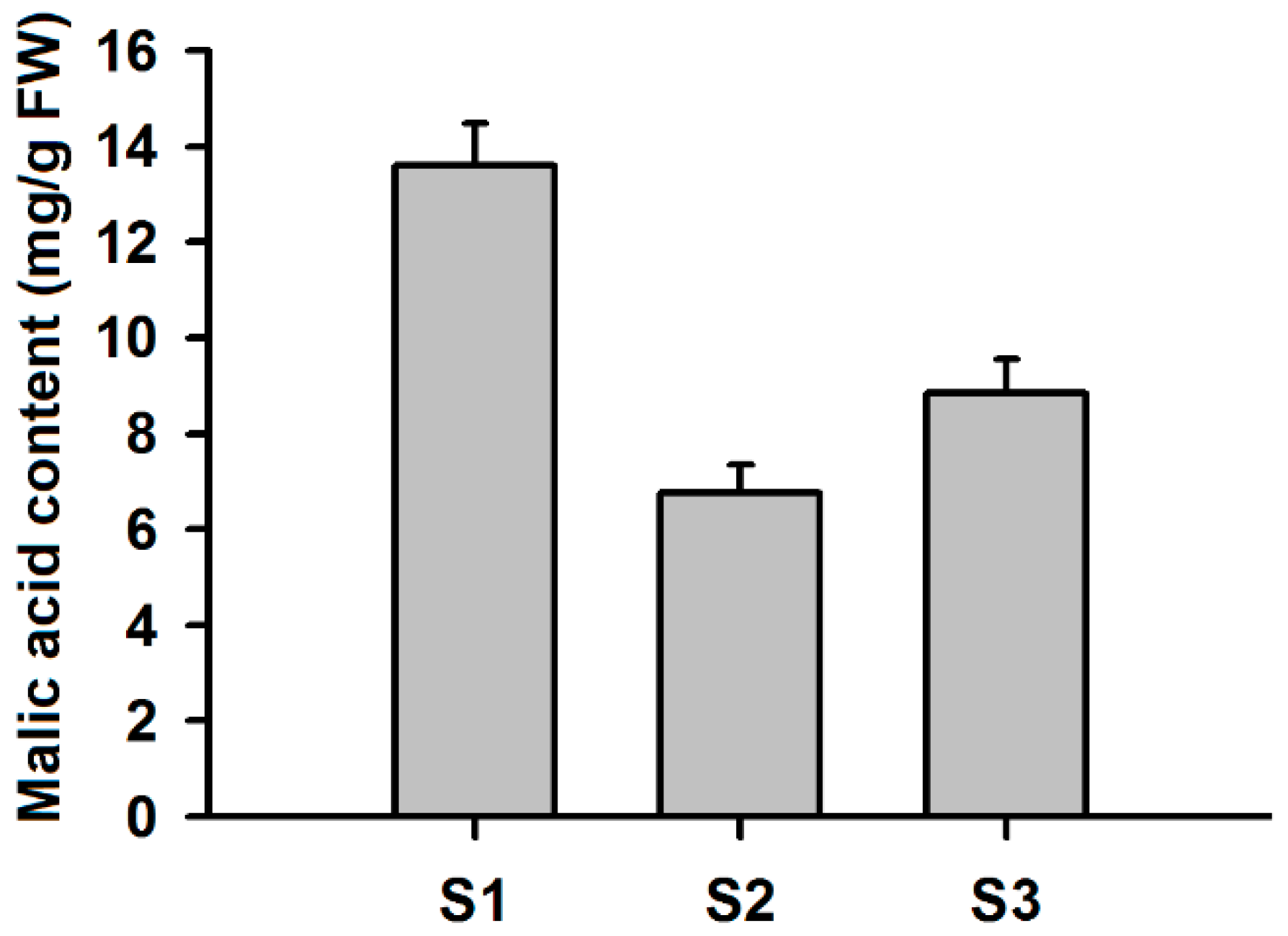
4.7. Measurements of Malic Acid Contents in Apple Fruits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ocheretina, O.; Scheibe, R. Cloning and sequence analysis of cDNAs encoding plant cytosolic malate dehydrogenase. Gene 1997, 199, 145–148. [Google Scholar] [CrossRef]
- Hall, M.D.; Levitt, D.G.; Banaszak, L.J. Crystal structure of Escherichia coli malate dehydrogenase: A complex of the apoenzyme and citrate at 1·87 Å resolution. J. Mol. Biol. 1992, 226, 867–882. [Google Scholar] [CrossRef]
- Minárik, P.; Tomásková, N.; Kollárová, M.; Antalík, M. Malate dehydrogenases—Structure and function. Gen. Physiol. Biophys. 2002, 21, 257–265. [Google Scholar] [PubMed]
- Hatch, M.D.; Slack, C.R. NADP-specific malate dehydrogenase and glycerate kinase in leaves and evidence for their location in chloroplasts. Biochem. Bioph. Res. Commun. 1969, 34, 589–593. [Google Scholar] [CrossRef]
- Kandoi, D.; Mohanty, S.; Tripathy, B.C. Overexpression of plastidic maize NADP-malate dehydrogenase (ZmNADP-MDH) in Arabidopsis thaliana confers tolerance to salt stress. Protoplasma 2018, 255, 547–563. [Google Scholar] [CrossRef] [PubMed]
- Beeler, S.; Liu, H.C.; Stadler, M.; Schreier, T.; Eicke, S.; Lue, W.L.; Truernit, E.; Zeeman, S.C.; Chen, J.; Kötting, O. Plastidial NAD-dependent malate dehydrogenase is critical for embryo development and heterotrophic metabolism in Arabidopsis. Plant Physiol. 2014, 164, 1175–1190. [Google Scholar] [CrossRef] [PubMed]
- Selinski, J.; König, N.; Wellmeyer, B.; Hanke, G.T.; Linke, V.; Neuhaus, H.E.; Scheibe, R. The plastid-localized NAD-dependent malate dehydrogenase is crucial for energy homeostasis in developing Arabidopsis thaliana seeds. Mol. Plant 2014, 7, 170–186. [Google Scholar] [CrossRef] [PubMed]
- Sew, Y.S.; Ströher, E.; Fenske, R.; Millar, A.H. Loss of Mitochondrial Malate Dehydrogenase Activity Alters Seed Metabolism Impairing Seed Maturation and Post-Germination Growth in Arabidopsis. Plant Physiol. 2016, 171, 849–863. [Google Scholar] [PubMed]
- Yudina, R.S. Malate dehydrogenase in plants: Its genetics, structure, localization and use as a marker. Adv. Biosci. Biotechnol. 2012, 3, 370–377. [Google Scholar] [CrossRef]
- Wang, Q.J.; Sun, H.; Dong, Q.L.; Sun, T.Y.; Jin, Z.X.; Hao, Y.J.; Yao, Y.X. The enhancement of tolerance to salt and cold stresses by modifying the redox state and salicylic acid content via the cytosolic malate dehydrogenase gene in transgenic apple plants. Plant Biotechnol. J. 2016, 14, 1986–1997. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.X.; Dong, Q.L.; Zhai, H.; You, C.X.; Hao, Y.J. The functions of an apple cytosolic malate dehydrogenase gene in growth and tolerance to cold and salt stresses. Plant Physiol. Bioch. 2011, 49, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rensing, S.A. Gene duplication as a driver of plant morphogenetic evolution. Curr. Opin. Plant Biol. 2014, 17, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Blanc, G.; Wolfe, K.H. Functional divergence of duplicated genes formed by polyploidy during Arabidopsis evolution. Plant Cell 2004, 16, 1679–1691. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of gene duplication in plant. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.Q.; Yuan, Y.Y.; Gao, M.; Qi, T.H.; Li, M.J.; Ma, F.W. Genome-Wide Identification, Molecular Evolution, and Expression Divergence of Aluminum-Activated Malate Transporters in Apples. Int. J. Mol. Sci. 2018, 19, 2807. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.S.; Raes, J. Duplication and divergence: The evolution of new genes and old ideas. Annu. Rev. Genet. 2004, 38, 615–643. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, H.; Ma, B.Q.; Owiti, A.; Korban, S.S.; Han, Y.P. Divergent evolutionary pattern of sugar transporter genes is associated with the difference in sugar accumulation between grasses and eudicots. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zheng, D.; Vimolmangkang, S.; Khan, M.A.; Beever, J.E.; Korban, S.S. Integration of physical and genetic maps in apple confirms whole-genome and segmental duplications in the apple genome. J. Exp. Bot. 2011, 62, 5117–5130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maere, S.; De Bodt, S.; Raes, J.; Casneuf, T.; Van Montagu, M.; Kuiper, M.; Van de Peer, Y. Modeling gene and genome duplications in eukaryotes. Proc. Nat. Acad. Sci. USA 2005, 102, 5454–5459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Wang, Z.; Shi, Z.; Zhang, S.; Ming, R.; Zhu, S. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2013, 23, 396–408. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Chagné, D.; Crowhurst, R.N.; Pindo, M.; Thrimawithana, A.; Deng, C.; Ireland, H.; Fiers, M.; Dzierzon, H.; Cestaro, A.; Fontana, P.; et al. The Draft Genome Sequence of European Pear (Pyrus communis L. ‘Bartlett’). PLoS ONE 2014, 9, e92644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The International Peach Genome Initiative. The high-quality draft genome of peach (Prunus persica) identifies unique patterns of genetic diversity, domestication and genome evolution. Nat. Genet. 2013, 45, 487–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanBuren, R.; Bryant, D.; Bushakra, J.M.; Vining, K.J.; Edger, P.P.; Rowley, E.R.; Priest, H.D.; Michael, T.P.; Lyons, E.; Filichkin, S.A.; et al. The genome of black raspberry (Rubus occidentalis). Plant J. 2016, 87, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic. Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: A program package for phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinf. 2010, 8, 77–80. [Google Scholar] [CrossRef]
- Ma, B.Q.; Liao, L.; Zheng, H.Y.; Chen, J.; Wu, B.H.; Ogutu, C.; Li, S.H.; Korban, S.S.; Han, Y.P. Genes encoding aluminum-activated malate transporter II and their association with fruit acidity in apple. Plant Genome 2015, 8, 1–14. [Google Scholar] [CrossRef]
- Xing, L.; Zhang, D.; Zhao, C.; Li, Y.; Ma, J.J.; An, N.; Han, M.Y. Shoot bending promotes flower bud formation by miRNA-mediated regulation in apple (Malus domestica Borkh.). Plant Biotechnol. J. 2016, 14, 749–770. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.B.; Zhang, D.; Li, Y.M.; Shen, Y.W.; Zhao, C.P.; Ma, J.J.; An, N.; Han, M.Y. Transcription Profiles Reveal Sugar and Hormone Signaling Pathways Mediating Flower Induction in Apple (Malus domestica Borkh.). Plant Cell Physiol. 2015, 56, 2052–2068. [Google Scholar] [CrossRef] [PubMed]
- Lou, Q.; Liu, Y.; Qi, Y.; Jiao, S.; Tian, F.; Jiang, L.; Wang, Y. Transcriptome sequencing and metabolite analysis reveals the role of delphinidin metabolism in flower colour in grape hyacinth. J. Exp Bot. 2014, 65, 3157–3164. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.Q.; Chen, J.; Zheng, H.Y.; Fang, T.; Ogutu, C.; Li, S.H.; Han, Y.P.; Wu, B.H. Comparative assessment of sugar and malic acid composition in cultivated and wild apples. Food Chem. 2015, 172, 86–91. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Animo acids | MW (kDa) | pI | GRAVY | Instability index | Aliphatic index |
|---|---|---|---|---|---|---|
| MDP0000159872 | 413 | 43.57 | 8.10 | 0.052 | 34.70 | 94.77 |
| MDP0000426029 | 413 | 43.44 | 6.61 | 0.076 | 33.78 | 94.77 |
| MDP0000334963 | 361 | 38.12 | 8.46 | 0.136 | 25.09 | 98.59 |
| MDP0000710761 | 339 | 35.51 | 8.98 | 0.120 | 43.04 | 101.27 |
| MDP0000277049 | 397 | 41.76 | 7.57 | 0.231 | 24.20 | 101.89 |
| MDP0000303739 | 360 | 38.47 | 8.86 | 0.133 | 30.27 | 101.25 |
| MDP0000191302 | 362 | 38.94 | 8.84 | 0.204 | 33.03 | 105.50 |
| MDP0000295823 | 946 | 103.76 | 8.32 | −0.085 | 46.76 | 95.25 |
| MDP0000141199 | 224 | 23.49 | 8.93 | 0.087 | 32.91 | 88.08 |
| MDP0000251316 | 289 | 31.21 | 8.86 | −0.030 | 34.54 | 99.07 |
| MDP0000575394 | 283 | 30.95 | 6.02 | 0.008 | 43.55 | 92.93 |
| MDP0000174740 | 465 | 50.03 | 6.38 | 0.010 | 30.12 | 97.12 |
| MDP0000926135 | 376 | 41.26 | 7.05 | 0.062 | 30.35 | 102.42 |
| MDP0000197620 | 468 | 51.69 | 7.02 | −0.050 | 36.72 | 95.00 |
| MDP0000170418 | 542 | 60.45 | 8.80 | −0.044 | 43.04 | 93.87 |
| MDP0000272403 | 478 | 52.82 | 5.99 | −0.058 | 31.05 | 94.87 |
| MDP0000302082 | 489 | 54.12 | 6.09 | −0.048 | 31.15 | 96.32 |
| MDP0000278198 | 517 | 57.10 | 6.05 | −0.029 | 29.90 | 97.35 |
| MDP0000807458 | 134 | 14.48 | 9.27 | 0.172 | 32.91 | 99.63 |
| MDP0000201006 | 154 | 16.87 | 9.22 | −0.019 | 34.47 | 99.35 |
| No. | Cis-Element Name | Cis-Element Sequence | Function |
|---|---|---|---|
| 1 | AE-box | AGAAACAA | Light responsive element |
| 2 | ARE | AAACCA | regulatory element essential for the anaerobic induction |
| 3 | As | TGACG | unknown |
| 4 | CAAT-box | CCAAT | cis-acting element in promoter and enhancer regions |
| 5 | CGTCA-motif | CGTCA | MeJA-responsiveness |
| 6 | CTAG-motif | ACTAGCAGAA | unknown |
| 7 | GA-motif | ATAGATAA | Light responsive element |
| 8 | GATA-motif | AAGATAAGATT | Light responsive element |
| 9 | GT1-motif | GGTTAA | Light responsive element |
| 10 | MBSI | AAAAAACSGTTA | MYB binding site involved in flavonoid biosynthesis |
| 11 | MYB | TAACYR | unknown |
| 12 | MYC | TCTCTTA/CAATTG | unknown |
| 13 | STRE | AGGGG | unknown |
| 14 | TATA-box | TATWWAAW | unknown |
| 15 | TGACG-motif | TGACG | MeJA-responsiveness |
| 16 | TGA-element | AACGAC | auxin-responsive element |
| 17 | WUN-motif | AAATTTCCT | wound-responsive element |
| Clade | Null Hypothesis | Alternative Hypothesis | LRT | ||||
|---|---|---|---|---|---|---|---|
| −In L | ω | −In L | ω1 | ω2 | Statistic | p | |
| I+II | 1777.25 | 0.12 | 1776.71 | 0.11 | 0.08 | 1.08 | > 0.05 |
| (I + II) + III | 2305.32 | 0.15 | 2299.96 | 0.10 | 0.28 | 10.7 | < 0.01 |
| (V + VI) + IV | 1991.21 | 0.13 | 1987.85 | 0.12 | 0.55 | 6.7 | < 0.01 |
| V + VI | 1555.13 | 0.12 | 1553.69 | 0.13 | 0.07 | 2.9 | > 0.05 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, B.; Yuan, Y.; Gao, M.; Xing, L.; Li, C.; Li, M.; Ma, F. Genome-wide Identification, Classification, Molecular Evolution and Expression Analysis of Malate Dehydrogenases in Apple. Int. J. Mol. Sci. 2018, 19, 3312. https://doi.org/10.3390/ijms19113312
Ma B, Yuan Y, Gao M, Xing L, Li C, Li M, Ma F. Genome-wide Identification, Classification, Molecular Evolution and Expression Analysis of Malate Dehydrogenases in Apple. International Journal of Molecular Sciences. 2018; 19(11):3312. https://doi.org/10.3390/ijms19113312
Chicago/Turabian StyleMa, Baiquan, Yangyang Yuan, Meng Gao, Libo Xing, Cuiying Li, Mingjun Li, and Fengwang Ma. 2018. "Genome-wide Identification, Classification, Molecular Evolution and Expression Analysis of Malate Dehydrogenases in Apple" International Journal of Molecular Sciences 19, no. 11: 3312. https://doi.org/10.3390/ijms19113312