The Potential Role of Auxin and Abscisic Acid Balance and FtARF2 in the Final Size Determination of Tartary Buckwheat Fruit

 ,
,
Abstract
:
1. Introduction
2. Results
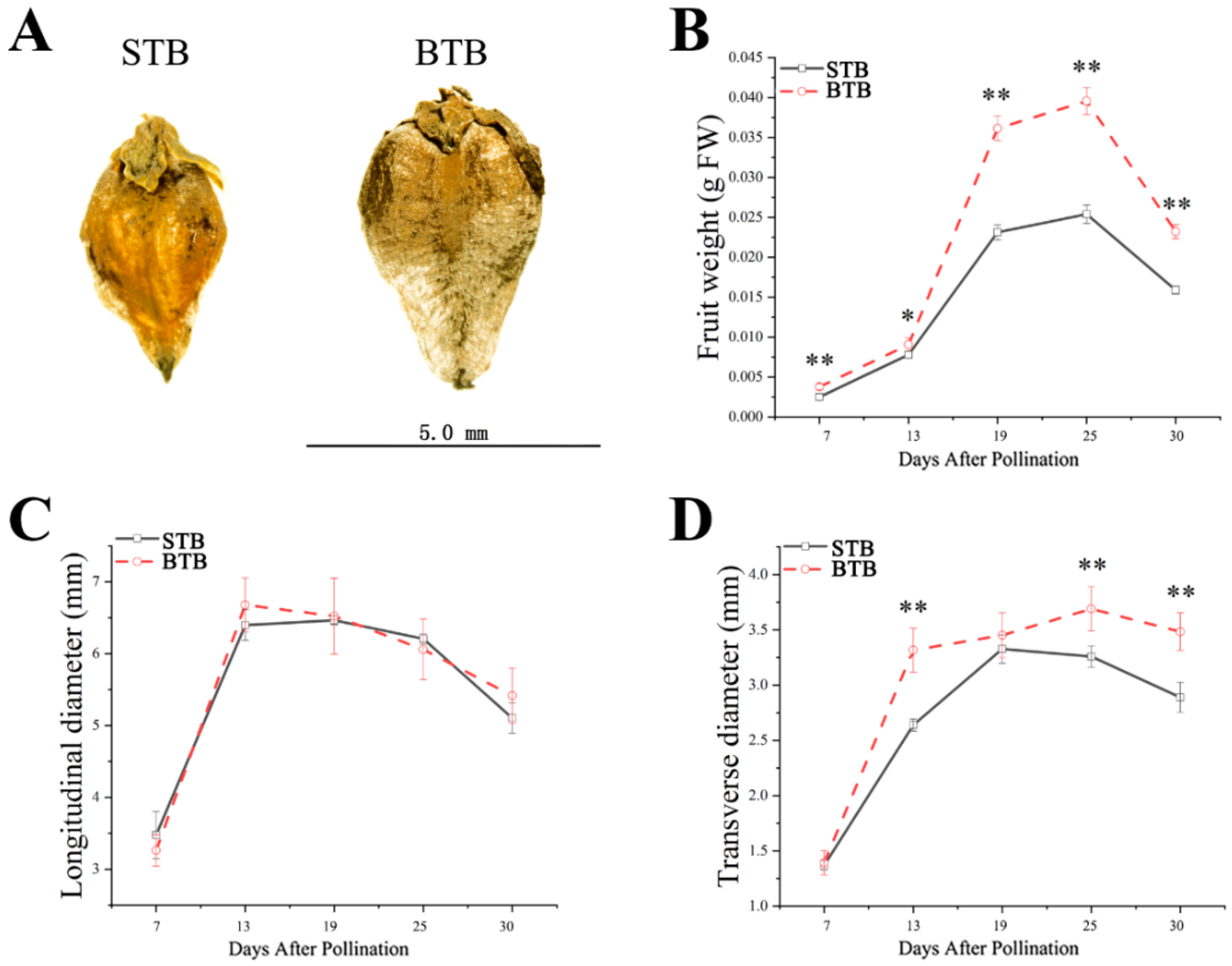
2.1. Morphological Characteristics of Tartary Buckwheat with Different Types of Fruits and Different Growth Patterns
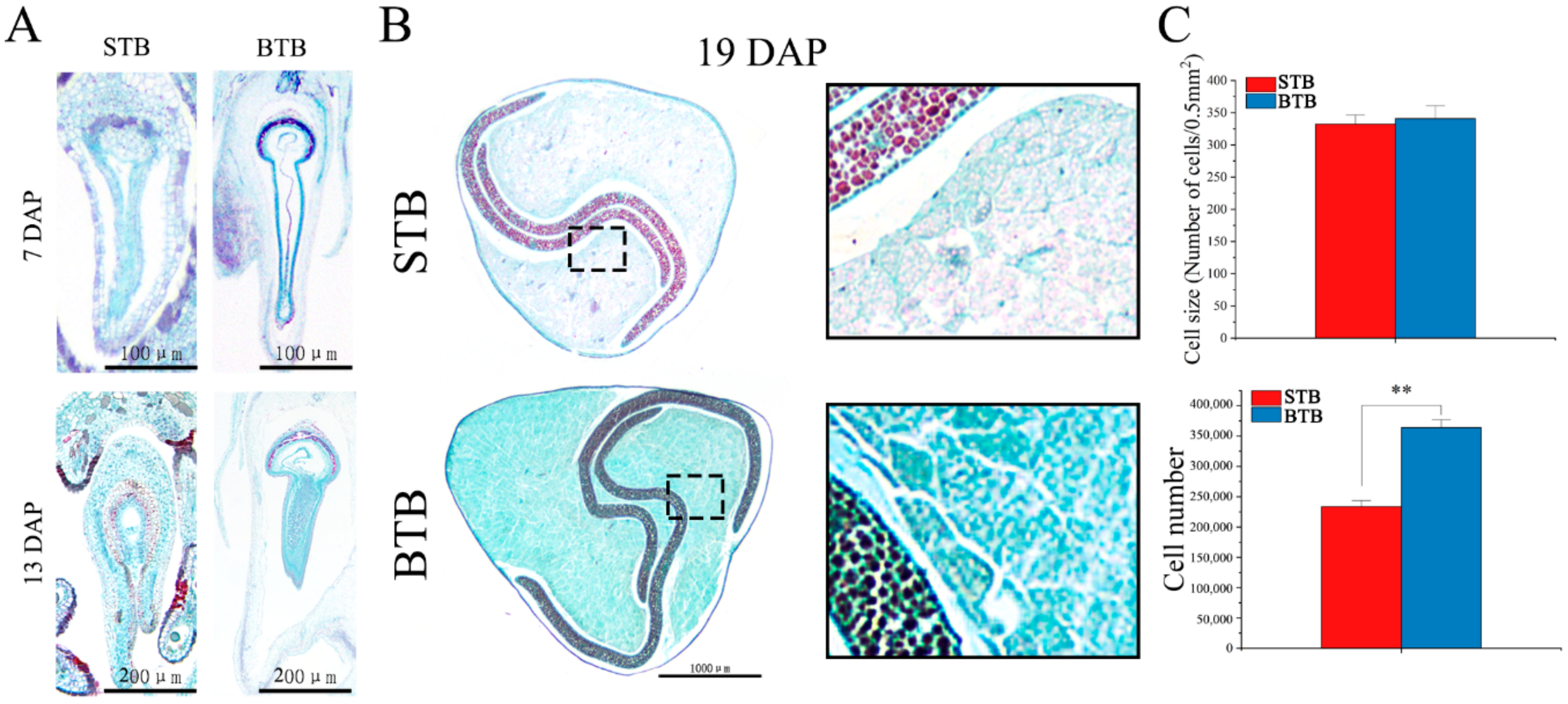
2.2. Anatomical Structure of Tartary Buckwheat with Different Fruit Types
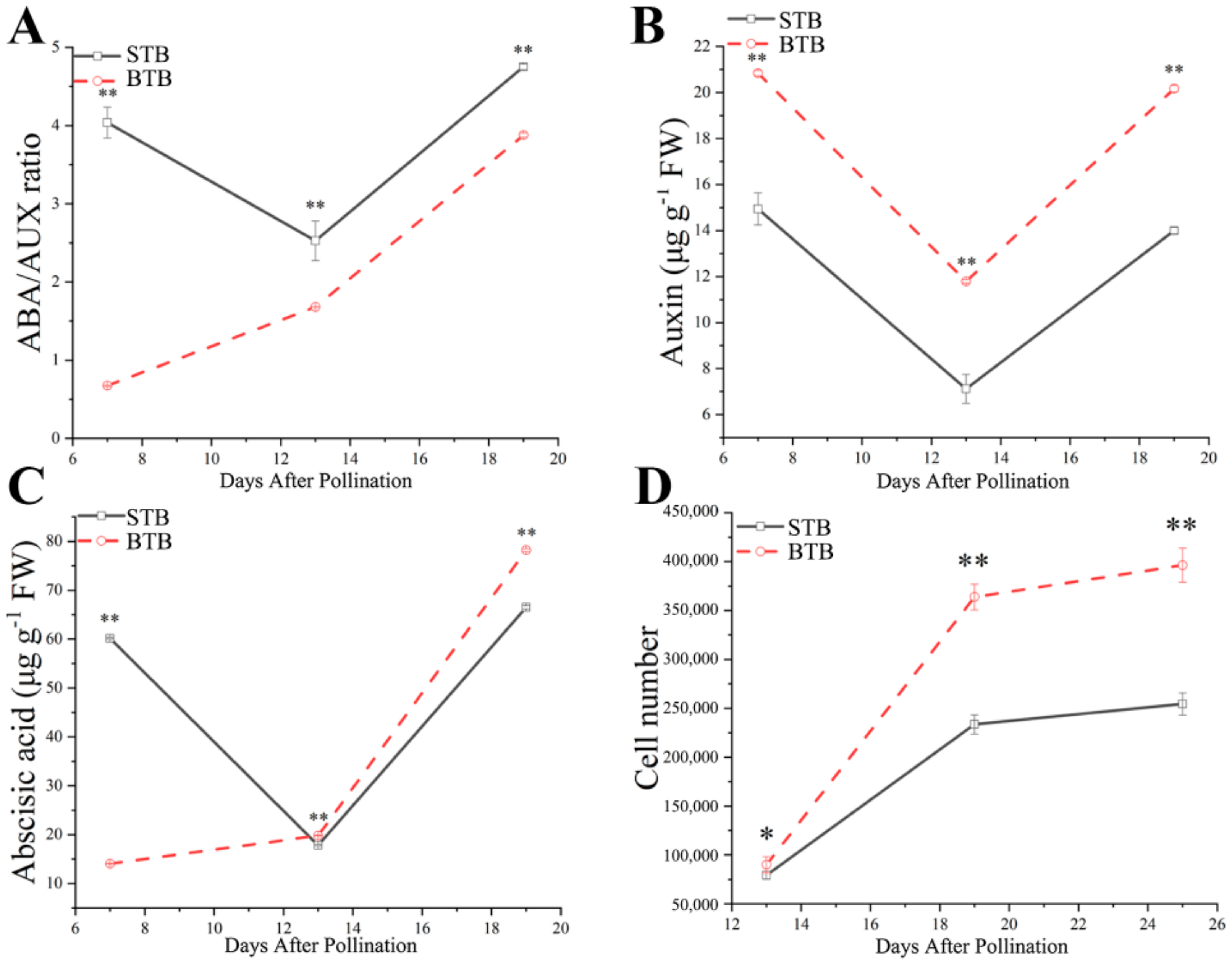
2.3. AUX and ABA Levels Correlate with the Variation in Fruit Size
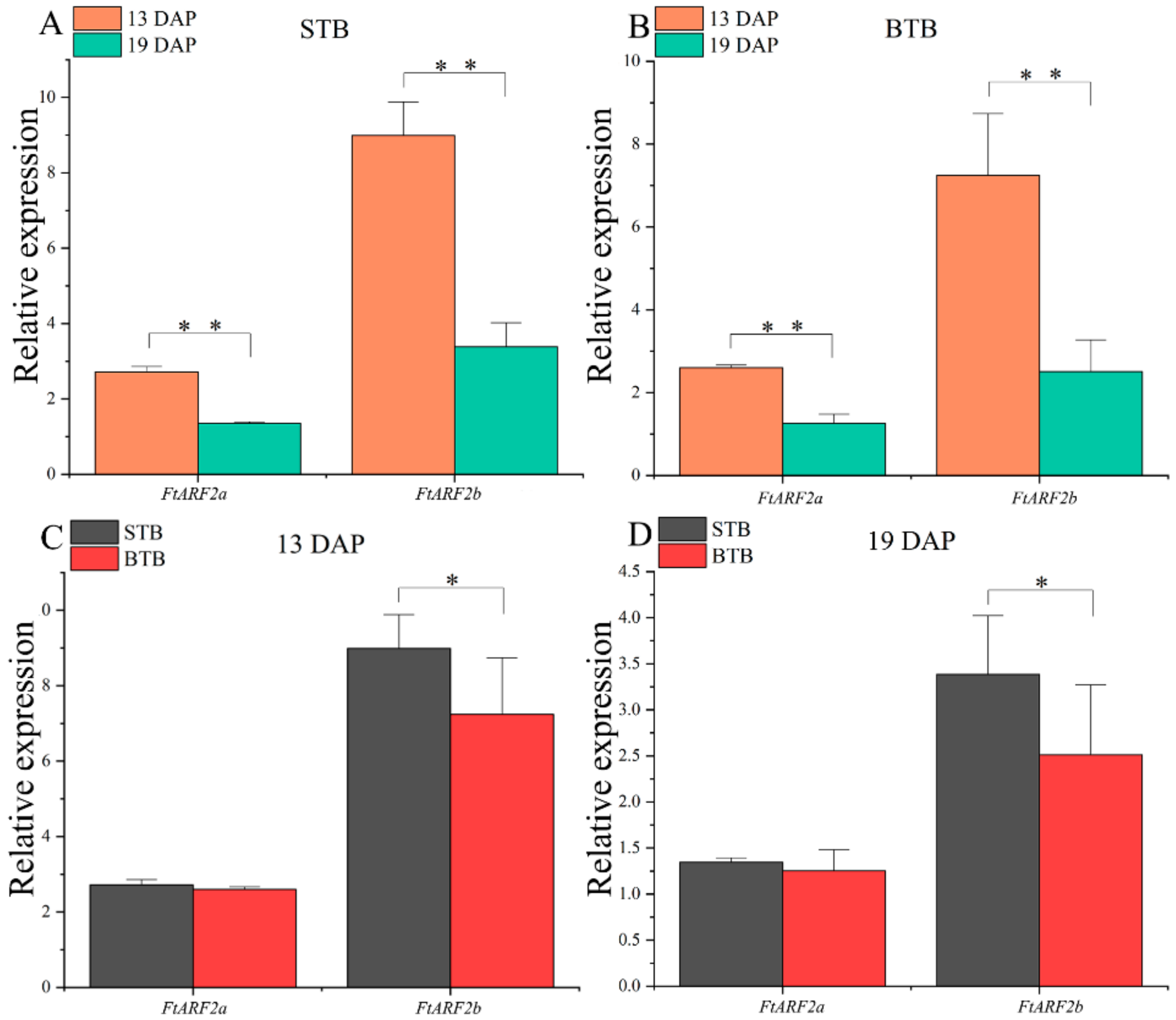
2.4. Expression of FtARF2 Genes of Tartary Buckwheat with Different Fruit Types from 13 to 19 DAP
2.5. Weight Gain of STB Fruit Induced by AUX
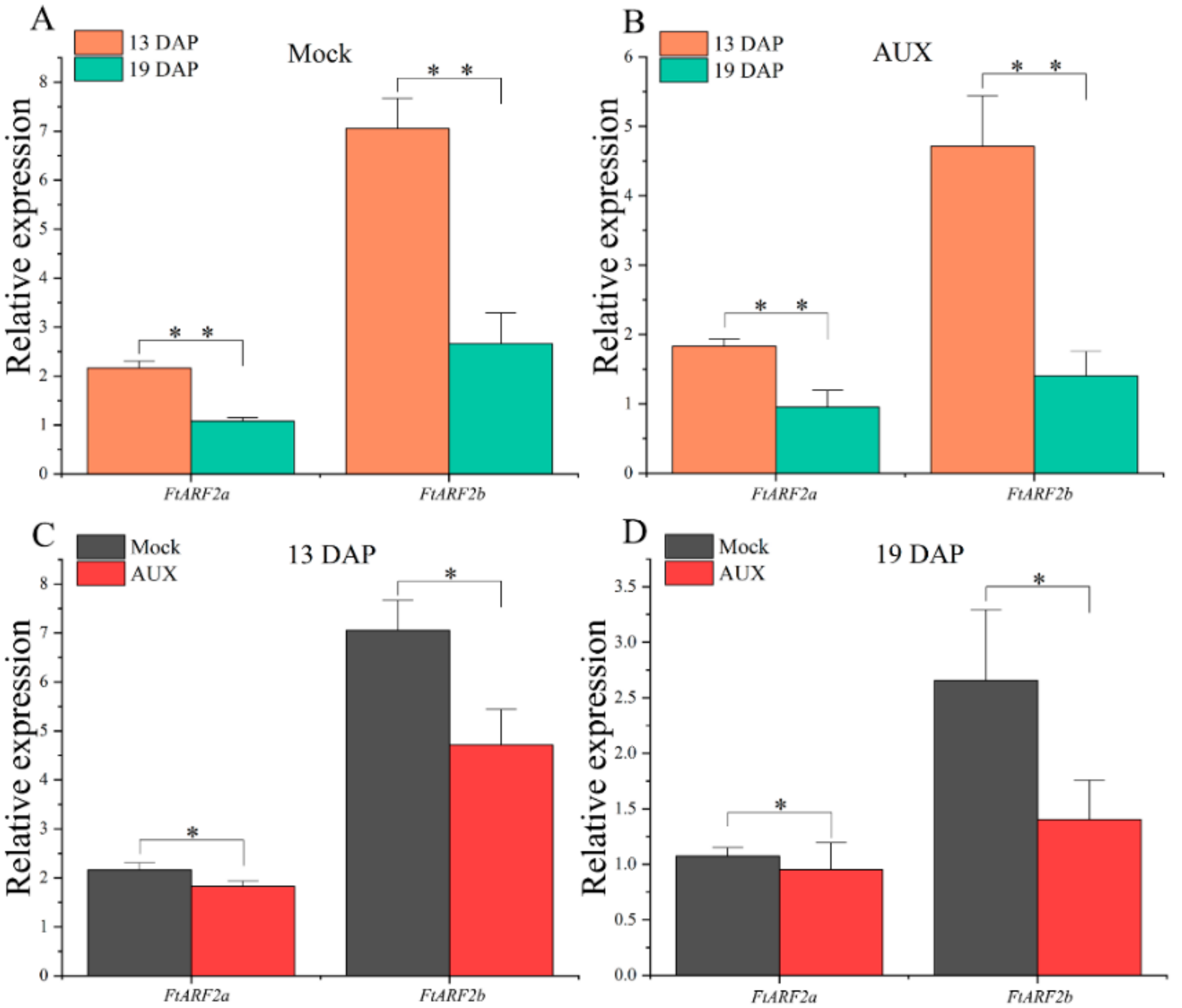
2.6. Expression of the FtARF2 Genes of STB Under AUX Treatment Conditions
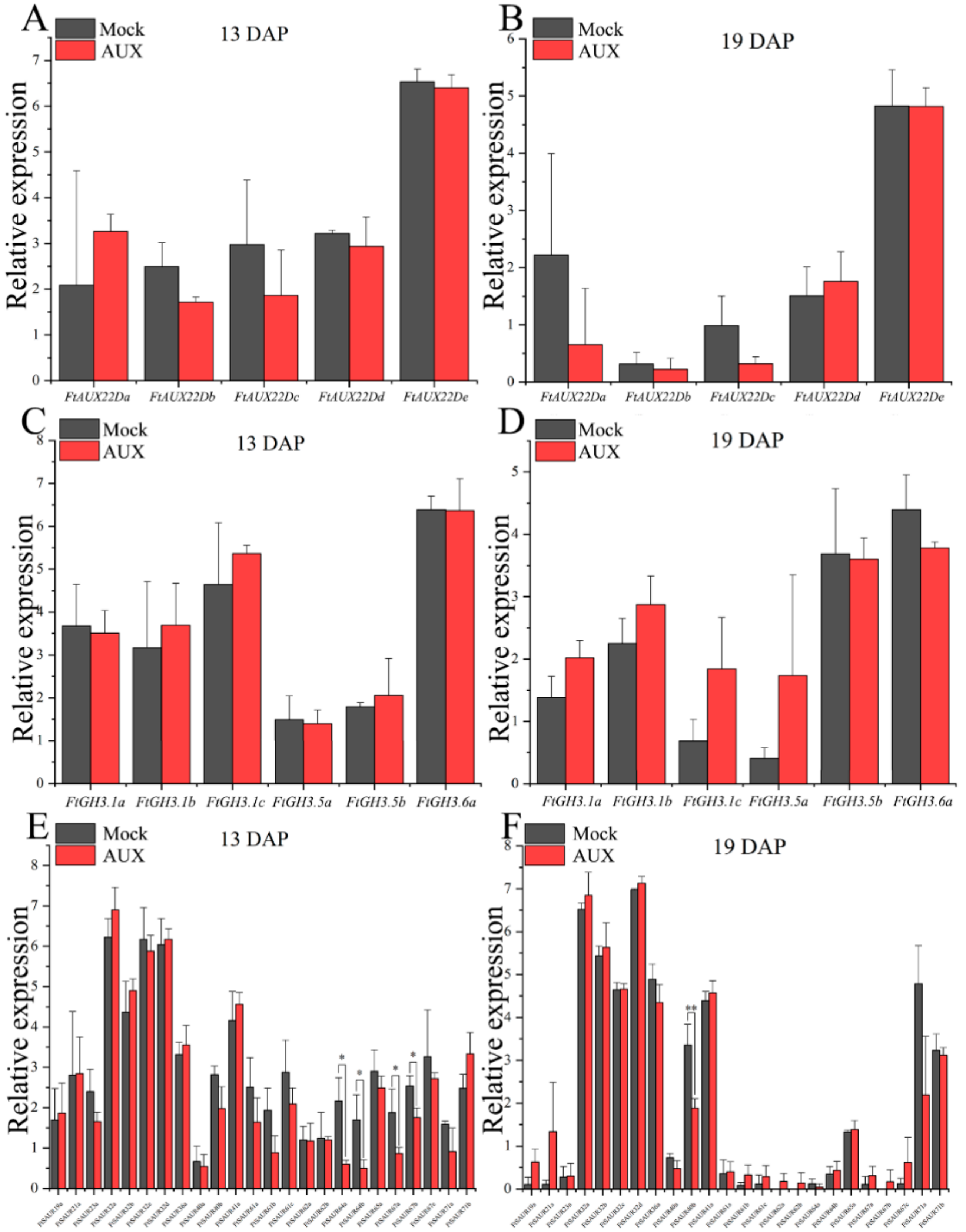
2.7. FtARF2 Acts Upstream of Fruit Development Genes
3. Discussion
3.1. Increased Cell Division During Embryogenesis Leads to an Increase in Fruit Size
3.2. Regulation of Plant Hormones on Cell Division
3.3. ARF2s and Their Downstream SAURs Regulate the Fruit Volume of Tartary Buckwheat at the End of Embryo Maturation
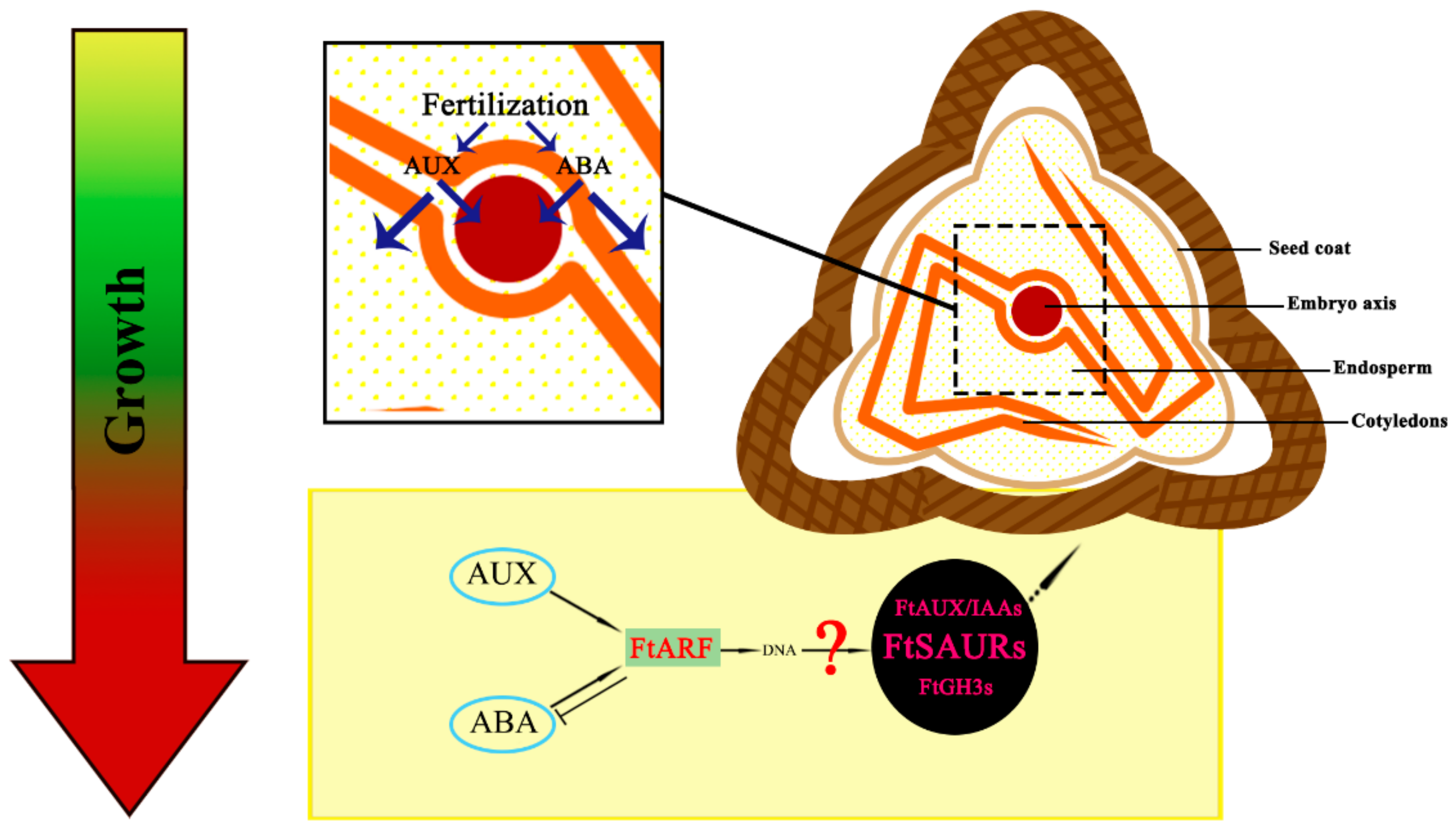
3.4. Model of Fruit Expansion of Tartary Buckwheat Development: A Time Consideration
4. Materials and Methods
4.1. Plant Material
4.2. Light Microscopy
4.3. Counting Total Number of Cells in Mature Fruits
4.4. Counting Number of Cells in a Given Area (Cell Size)
4.5. Phytohormone Analysis
4.6. AUX Treatment of STB
4.7. Gene Identification and Phylogenetic Analysis
4.8. Real-Time PCR Confirmation of Differentially Expressed Genes
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| ARF2 | auxin response factor 2 |
| AUX | auxin |
| Aux/IAAs | auxin/indole acetic acid |
| BTB | big Tartary buckwheat |
| DAP | days after pollination |
| HPLC | high performance liquid chromatography |
| GH3s | Gretchen Hagen 3 |
| IAA | indoleacetic acid |
| ICK1 | cyclin dependent kinase inhibitor |
| RT-qPCR | reverse transcription-quantitative PCR |
| SAURs | small auxin up RNAs |
| STB | small Tartary buckwheat |
References
- Tweddle, J.C.; Dickie, J.B.; Webb, C.O.; Moles, A.T.; Ackerly, D.D.; Westoby, M. A Brief History of Seed Size. Science 2005, 307, 576–580. [Google Scholar]
- Li, N.; Li, Y. Ubiquitin-mediated control of seed size in plants. Front. Plant Sci. 2014, 5, 332. [Google Scholar] [CrossRef] [PubMed]
- Sundaresan, V. Control of seed size in plants. Proc. Natl. Acad. Sci. USA 2005, 102, 17887–17888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, R.B.; Paiva, G.D.; Yadegari, R. Plant Embryogenesis: Zygote to Seed. Science 1994, 266, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, U.; Xu, W.; Liu, A. Transcriptome profiling identifies ABA mediated regulatory changes towards storage filling in developing seeds of castor bean (Ricinus communis L.). Cell Biosci. 2014, 4, 33. [Google Scholar] [CrossRef] [PubMed]
- Schussler, J.R.; Brenner, M.L.; Brun, W.A. Abscisic Acid and its relationship to seed filling in soybeans. Plant Physiol. 1984, 76, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Möller, B.; Weijers, D. Auxin Control of Embryo Patterning. Cold Spring Harb. Perspect. Biol. 2009, 1, a001545. [Google Scholar] [CrossRef] [PubMed]
- Venglat, P.; Xiang, D.; Wang, E.; Datla, R. Genomics of seed development: Challenges and opportunities for genetic improvement of seed traits in crop plants. Biocatal. Agric. Biotechnol. 2014, 3, 24–30. [Google Scholar] [CrossRef]
- Mizutani, M.; Naganuma, T.; Tsutsumi, K.; Saitoh, Y. The syncytium-specific expression of the Orysa;KRP3 CDK inhibitor: Implication of its involvement in the cell cycle control in the rice (Oryza sativa L.) syncytial endosperm. J. Exp. Bot. 2010, 61, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Jong, A.D.; Koerselmankooij, J.W.; Schuurmans, J.; Borstlap, A.C. The Mechanism of Amino Acid Efflux from Seed Coats of Developing Pea Seeds as Revealed by Uptake Experiments. Plant Physiol. 1997, 114, 731–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, H.; Borisjuk, L.; Wobus, U. Molecular physiology of legume seed development. Annu. Rev. Plant Biol. 2005, 56, 253–279. [Google Scholar] [CrossRef] [PubMed]
- Guilfoyle, T.J.; Hagen, G. Auxin response factors. Curr. Opin. Plant Biol. 2007, 10, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Zouine, M.; Fu, Y.; Chateignerboutin, A.L.; Mila, I.; Frasse, P.; Wang, H.; Audran, C.; Roustan, J.P.; Bouzayen, M. Characterization of the Tomato ARF Gene Family Uncovers a Multi-Levels Post-Transcriptional Regulation Including Alternative Splicing. PLoS ONE 2014, 9, e84203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, S.B.; Hagen, G.; Guilfoyle, T. The Roles of Auxin Response Factor Domains in Auxin-Responsive Transcription. Plant Cell 2003, 15, 533–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okushima, Y.; Mitina, I.; Quach, H.L.; Theologis, A. Auxin response factor 2 (ARF2): A pleiotropic developmental regulator. Plant J. 2010, 43, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.M.; Nagpal, P.; Young, J.C.; Hagen, G.; Guilfoyle, T.J.; Reed, J.W. Auxin response factor 1 and Auxin response factor 2 regulate senescence and floral organ abscission in Arabidopsis thaliana. Development 2005, 132, 4563–4574. [Google Scholar] [CrossRef] [PubMed]
- Schruff, M.C.; Spielman, M.; Tiwari, S.; Adams, S.; Fenby, N.; Scott, R.J. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development 2006, 133, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hua, D.; He, J.; Duan, Y.; Chen, Z.; Hong, X.; Gong, Z. Auxin Response Factor2 (ARF2) and Its Regulated Homeodomain Gene HB33 Mediate Abscisic Acid Response in Arabidopsis. PLoS Genet. 2011, 7, e1002172. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.F. Seed proteins of buckwheat. In Buckwheat Plant Science; Du, M.F., Fan, Y., Eds.; Science Press: Beijing, China, 2012. [Google Scholar]
- Liu, D.S.; Xu, R.Y.; Wang, Q.Q. Analysis of protein content and Amino Acid composition in Buckwheat. Chin. Seed ind. 1997, 2, 26–28. [Google Scholar]
- Lin, R.F. Tartary buckwheat. Agrotechny 2013, 21, 41. [Google Scholar]
- Jain, M.; Kaur, N.; Tyagi, A.K.; Khurana, J.P. The auxin-responsive GH3 gene family in rice (Oryza sativa). Funct. Integr. Genom. 2006, 6, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Hagen, G.; Guilfoyle, T. Auxin-responsive gene expression: Genes, promoters and regulatory factors. Plant Mol. Biol. 2002, 49, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Liscum, E.; Reed, J.W. Genetics of Aux/IAA and ARF action in plant growth and development. Plant Mol. Biol. 2002, 49, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Chojecki, A.J.S.; Bayliss, M.W.; Gale, M.D. Cell Production and DNA Accumulation in the Wheat Endosperm, and their Association with Grain Weight. Ann. Bot. 1986, 58, 809–817. [Google Scholar] [CrossRef]
- Chojecki, A.J.S.; Gale, M.D.; Bayliss, M.W. The Number and Sizes of Starch Granules in the Wheat Endosperm, and their Association with Grain Weight. Ann. Bot. 1986, 58, 819–831. [Google Scholar] [CrossRef]
- Munier-Jolain, N.G.; Ney, B. Seed growth rate in grain legumes II. Seed growth rate depends on cotyledon cell number. J. Exp. Bot. 1998, 49, 1971–1976. [Google Scholar] [CrossRef] [Green Version]
- Perrot-Rechenmann, C. Cellular responses to auxin: Division versus expansion. Cold Spring Harb. Perspect. Biol. 2010, 2, a001446. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.J.; Estelle, M. Mechanism of auxin-regulated gene expression in plants. Annu. Rev. Genet. 2009, 43, 265–285. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.F.; Peng, H.R.; Guo, X.; Zhao, G.; Chen, Q.F. Differences of Endogenous hormones in two Tartary Buckwheat cultivars at maturity and their relationship with yield and quality. Jiangsu J. Agric. 2013, 29, 28–32. [Google Scholar]
- Qin, P.Y. Evaluation of the Resources and quality of main Buckwheat varieties in China and the effect of processing on the composition and activity of Buckwheat. Chin. Acad. Agric. Sci. 2012, 1, 265–285. [Google Scholar]
- Yang, X.; Ren, Z.X.; Lin, D.B.; Xian, Z.Q.; Li, Z.G. Expression pattern of SLARF2 Gene in Tomato and its function Analysis in Fruit Development. J. Trop. Crop 2014, 35, 1969–1974. [Google Scholar]
- Mei, Z.; Bing, Y.; Ping, L. The role of ABA in triggering ethylene biosynthesis and ripening of tomato fruit. J. Exp. Bot. 2009, 60, 1579–1588. [Google Scholar] [Green Version]
- Ofosu-Anim, J.; Kanayama, Y.; Yamaki, S. Sugar uptake into strawberry fruit is stimulated by abscisic acid and indoleacetic acid. Physiol. Plant. 2010, 97, 169–174. [Google Scholar] [CrossRef]
- Manning, K. Changes in gene expression during strawberry fruit ripening and their regulation by auxin. Planta 1994, 194, 62–68. [Google Scholar] [CrossRef]
- Jiang, Y.; Joyce, D.C. ABA effects on ethylene production, PAL activity, anthocyanin and phenolic contents of strawberry fruit. Plant Growth Regul. 2003, 39, 171–174. [Google Scholar] [CrossRef]
- Kano, Y.; Asahira, T. Roles of Cytokinin and Abscisic Acid in the Maturing of Strawberry Fruits. J. Jpn. Soc. Hort. Sci. 1981, 50, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, R.R.; Gampala, S.S.; Rock, C.D. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002, 14, S15. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qi, Q.; Schorr, P.; Cutler, A.J.; Crosby, W.L.; Fowke, L.C. ICK1, a cyclin-dependent protein kinase inhibitor from Arabidopsis thaliana interacts with both Cdc2a and CycD3, and its expression is induced by abscisic acid. Plant J. 2010, 15, 501–510. [Google Scholar] [CrossRef]
- Guilfoyle, T.J. Auxin-regulated genes and promoters. New Compr. Biochem. 1999, 33, 423–459. [Google Scholar]
- Guilfoyle, T.J.; Gretchen, H. Auxin response factors: Recent advances in auxin biology. J. Plant Growth Regulat. 2001, 3, 317–319. [Google Scholar]
- Guilfoyle, T.J. Aux/IAA proteins and auxin signal transduction. Trends Plant Sci. 1998, 3, 205–207. [Google Scholar] [CrossRef]
- Jameson, P.E.; Song, J. Cytokinin: A key driver of seed yield. J. Exp. Bot. 2015, 67, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Gil, P.; Liu, Y.; Poff, K.L.; Green, P.J. Characterization of the Auxin-Inducible SAUR-AC1 Gene for Use as a Molecular Genetic Tool in Arabidopsis. Plant Physiol. 1994, 104, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Roux, C.; Bilang, J.; Theunissen, B.H.; Perrot-Rechenmann, C. Identification of new early auxin markers in tobacco by mRNA differential display. Plant Mol. Biol. 1998, 37, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Guilfoyle, T.J.; Hagen, G.; Li, Y.; Ulmasov, T.; Liu, Z.B.; Strabala, T.; Gee, M. Auxin-Regulated Transcription. Funct. Plant Biol. 1993, 20, 489–502. [Google Scholar] [CrossRef]
- Mcclure, B.A.; Guilfoyle, T. Characterization of a class of small auxin-inducible soybean polyadenylated RNAs. Plant Mol. Biol. 1987, 9, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Zhu, Y.; Gao, C.; She, W.; Lin, W.; Chen, Y.; Han, N.; Bian, H.; Zhu, M.; Wang, J. Tissue-specific expression of SMALL AUXIN UP RNA41 differentially regulates cell expansion and root meristem patterning in Arabidopsis. Plant Cell Physiol. 2013, 54, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Kant, S.; Rothstein, S. Auxin-responsive SAUR39 gene modulates auxin level in rice. Plant Signal. Behav. 2009, 4, 1174–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Q.; Reed, J.W. Arabidopsis SHY2/IAA3 inhibits auxin-regulated gene expression. Plant Cell 2002, 14, 301–319. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Stone, J. Arabidopsis thaliana GH3.9 influences primary root growth. Planta 2007, 226, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Xu, X. The Role of Gibberellin, Abscisic Acid, and Sucrose in the Regulation of Potato Tuber Formation In Vitro. Plant Physiol. 1998, 117, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Baydar, H.; Ulger, S. Correlations Between Changes in the Amount of Endogenous Phytohormones and Flowering in the Safflower (Carthamus tinctorius L.). Turk. J. Biol. 2014, 22, 421–426. [Google Scholar]
- Mwange, K.N.; Hou, H.W.; Cui, K.M. Relationship between endogenous indole-3-acetic acid and abscisic acid changes and bark recovery in Eucommia ulmoides Oliv. after girdling. J. Exp. Bot. 2003, 54, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Tyagi, A.K.; Sharma, A.K. Genome-wide analysis of auxin response factor (ARF) gene family from tomato and analysis of their role in flower and fruit development. Mol. Genet. Genom. 2011, 285, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Kenneth, J.L.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using RT-qPCR. Methods 2001, 25, 402–408. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | Transverse Diameter (mm) | Longitudinal Diameter (mm) | Weight (g) |
|---|---|---|---|
| BTB | 3.48 (±0.17) | 5.42 (±0.38) | 0.023 (±0.00 087) |
| STB | 2.89 (±0.14) | 5.10 (±0.21) | 0.016 (±0.00 051) |
| BTB/STB ratio | 120% | 106% | 143% |
| Gene | Gene ID | Protein Length | Mw (kDa) | pI | Location | Domain | Homologous | E-Value | Similarity | Localization |
|---|---|---|---|---|---|---|---|---|---|---|
| FtARF2.01 | FtPinG0002469700.01 | 766 | 81 | 6.03 | Ft1 | DBD, ARF, CTD | AtARF2 | 8 × 10−50 | 79 | Nucleus |
| FtARF2.02 | FtPinG0005575800.01 | 721 | 85 | 6.03 | Ft2 | DBD, ARF, CTD | AtARF2 | 1 × 10−42 | 79 | Nucleus |
| Materials | IAA Concentration (mg L−1) | Weight (g) |
|---|---|---|
| STB | 0 (Mock) | 0.015 90 (±0.00 051) ad |
| 40 | 0.015 94 (±0.00 041) ab | |
| 70 | 0.014 82 (±0.00 075) cd | |
| 100 | 0.016 44 (±0.00 055) a | |
| 130 | 0.015 38 (±0.00 059) bc | |
| 160 | 0.014 56 (±0.00 060) d |
| No. | Cultivar | Size | Shape | Weight (g) |
|---|---|---|---|---|
| 1 | 87-23 (F6080) | Big | Peach shape | 0.0257 |
| 2 | HEI LI MI QIAO | Big | Long shape | 0.0244 |
| 3 | 87-26 (F6245) | Big | Long cone | 0.0232 |
| 4 | LIAO QIAO75 HUAO (KU) | Big | Long shape | 0.0231 |
| 5 | XI QIAO | Big | Long shape | 0.0230 |
| 6 | 87-27 (F6273) | Big | Long cone | 0.0230 |
| 7 | HEI ZI QIAO | Medium | Wide | 0.0228 |
| 8 | Mai QIAO | Medium | Long shape | 0.0218 |
| 9 | 87-14 (F3091) | Medium | Long cone | 0.0218 |
| 10 | YUAN ZI QIAO | Medium | Long shape | 0.0214 |
| 11 | BIAN ZI QIAO | Medium | Long shape | 0.0207 |
| 12 | YANG QU KU QIAO | Medium | Long shape | 0.0203 |
| 13 | KU QIAO (TUO YUAN) | Medium | Long shape | 0.0203 |
| 14 | XI YANG KU QIAO | Medium | Long shape | 0.0199 |
| 15 | SHI CHE E | Medium | Long shape | 0.0196 |
| 16 | JIU JIANG KU QIAO | Medium | Long shape | 0.0195 |
| 17 | PING DING KU QIAO | Medium | Long shape | 0.0194 |
| 18 | MA KU QIAO | Medium | Wide | 0.0193 |
| 19 | E LUO WU QIE | Medium | Long shape | 0.0192 |
| 20 | KU TIAO ZI | Medium | Long shape | 0.0190 |
| 21 | KU BING QIAO | Medium | Wide | 0.0187 |
| 22 | KU QIAO MAI | Small | Long shape | 0.0160 |
| 23 | MI QIAO | Small | Long shape | 0.0160 |
| 24 | ER BAI KU QIAO | Small | Long shape | 0.0158 |
| 25 | 88-33 (IV-171) | Small | Short cone | 0.0154 |
| 26 | LENG FAN TUAN | Small | Long shape | 0.0151 |
| 27 | HEI KU QIAO | Small | Wide | 0.0150 |
| 28 | HONG XI KU QIAO | Small | Long shape | 0.0150 |
| 29 | MA QIAO | Small | Long cone | 0.0149 |
| 30 | HUI CHA KU QIAO | Small | Wide | 0.0148 |
| 31 | 82-4-6 | Small | Long shape | 0.0146 |
| 32 | XI KU QIAO | Small | Long shape | 0.0140 |
| 33 | 82-8-1 | Small | Long shape | 0.0130 |
| 34 | LV QIAO | Small | Long shape | 0.0100 |
| Gene ID | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| FtARF2.01 | ACCTTCCGTTTCTCCACTGACA | GACCCTTGATAATGATAACCCACTT |
| FtARF2.02 | AGACTTGTGGCTGGTGACGCT | GCTAGATATGACTGACGAGGGAACT |
| FtGH3.1a | AAGTTGGTGGACATGGTTGACG | TGCGGAGCAGAGTTGTGGAA |
| FtGH3.1b | TTCCTCCACCTTCATTACCCG | GATACCTTCCTCCCAGTTCTCCTT |
| FtGH3.1c | CGGAACTCGCTGACTTTATCATATC | AGGGAGACCACCGCTGTAGAA |
| FtGH3.5a | CGTTGTGCTTCCTCAAATCGG | TCGCCAACCTCAACCTCAGTC |
| FtGH3.5b | CGGACTCGTACCAAAGCATGT | CTTGATAGCACGAATGAACCCA |
| FtGH3.6a | AGATGCGTCAAGTTCAAGCCA | TGCTTATGACCAGGAACCCACT |
| FtSAUR19a | TCAAAGCCATACTCCTCTTCAGC | TATGGTGGGGCTCGAAGGA |
| FtSAUR21a | GCACTTTGCGGTGTATGTAGGC | TTGTTCATCACAGGGAATGGTTAG |
| FtSAUR23a | GGGTGGATGGGTATTTGGGTG | GCCGCCTTGAGGAGGTTGAT |
| FtSAUR32a | CGATTACGATCCCTTGCCATG | TCAAGCCCTAAAACACCCAACA |
| FtSAUR32b | GAGAGGAGATGCGGCGATTC | CGAAGTACGCCTTTCTGCTGG |
| FtSAUR32c | ATGGCGATTATGCGAAAGCT | ACGATCACCGGAACATACCCT |
| FtSAUR32d | GGTTTCTTATTCCGACTCAGTTCAT | GAATCTCCCTTAAACAATCTGGCT |
| FtSAUR36a | GTCATTCCGAGGTCACCAACAA | AACCCACGGCGGCTTTATG |
| FtSAUR40a | GGACACGAAGCAGAGGAGCA | TTGAGCGACCAAGAACTGAACG |
| FtSAUR40b | ATCTGGCGGTTGATGTGGG | GATACGGGGCAAGGGATAGTG |
| FtSAUR41a | ATTGGACATCCGTCTATTATTGCTC | TAATCTGCCTGAAAGAGTCCACG |
| FtSAUR61a | TTGCCTTCTGTTATCTCAACCACC | CATACCTTTGCTTATCCTCATCGTC |
| FtSAUR61b | TTCCGTTTGCCCTCTGTGAT | GTAAGACAGCGGCACCACATAA |
| FtSAUR61c | TGAGAAGGGCCAGTTTGTCGT | ACTTGCCAGCCCAAACTCTTC |
| FtSAUR62a | TTACACTTCCCTTGTCTTATCTCGG | GGATCAATGCTTGCTCTAGTTCTCT |
| FtSAUR62b | GAGAAGTAACAACACCAAGCAATCA | ATAAGACAAGGGAAGTGTAAAGCGT |
| FtSAUR64a | GAAGAGTTTGGGTTGGTTGGTG | GACACAATCAACGCTTCTTCCAA |
| FtSAUR64b | ATGGCAGAAGAGGAGTTTGGATT | GAGTTCTCTAGCTGCGTTCCGAT |
| FtSAUR65a | ACTTTGTGGTCTACACCGCTGAT | AACTCCTCCTCCGCCATTCTA |
| FtSAUR67a | GCAGTGGCAAGTAGGAAGAGGA | GAGATTTGAGATACACCAAGGGAAG |
| FtSAUR67b | CCAAGGAGCAACAGCACCAA | CAAACTCCTCTTCCGCCATTCT |
| FtSAUR67c | TTGGCAAGAAAGTGGAGAAAGC | CCGTGTAGACCATAAACTGACCCT |
| FtSAUR71a | CATTTCCAAGAAGCAACAACACC | TCCAAACTCCTCTTCTGCCATT |
| FtSAUR71b | CAACAGCACCAACAAATCATCAAC | CAAACTCCTCTTCGGCCATTC |
| FtAUX22Da | GTGTTGGAAAGCATGTTTAAGGTG | CGAAGCCTCTTACACGACGACA |
| FtAUX22Db | AGTGAGCTACTAGAGGCATTGGAAG | CCACGGAACATCACCAACAAG |
| FtAUX22Dc | CAAGGCTTACAAGAGCTACATCGA | AACATTTCCCACGGCACATC |
| FtAUX22Dd | GGATCGACCGACGACCACTT | GCCATCCAACTATTTGTGCCTT |
| FtAUX22De | AGATCAACACTCAACCCTCTGCTT | GCTCCGTCCATGCTTACTTTCAC |
| FtH3 | GAAATTCGCAAGTACCAGAAGAG | CCAACAAGGTATGCCTCAGC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Ma, Z.; Zheng, T.; Wang, J.; Huang, L.; Sun, W.; Zhang, Y.; Jin, W.; Zhan, J.; Cai, Y.; et al. The Potential Role of Auxin and Abscisic Acid Balance and FtARF2 in the Final Size Determination of Tartary Buckwheat Fruit. Int. J. Mol. Sci. 2018, 19, 2755. https://doi.org/10.3390/ijms19092755
Liu M, Ma Z, Zheng T, Wang J, Huang L, Sun W, Zhang Y, Jin W, Zhan J, Cai Y, et al. The Potential Role of Auxin and Abscisic Acid Balance and FtARF2 in the Final Size Determination of Tartary Buckwheat Fruit. International Journal of Molecular Sciences. 2018; 19(9):2755. https://doi.org/10.3390/ijms19092755
Chicago/Turabian StyleLiu, Moyang, Zhaotang Ma, Tianrun Zheng, Jing Wang, Li Huang, Wenjun Sun, Yanjun Zhang, Weiqiong Jin, Junyi Zhan, Yuntao Cai, and et al. 2018. "The Potential Role of Auxin and Abscisic Acid Balance and FtARF2 in the Final Size Determination of Tartary Buckwheat Fruit" International Journal of Molecular Sciences 19, no. 9: 2755. https://doi.org/10.3390/ijms19092755