Discovery of lahS as a Global Regulator of Environmental Adaptation and Virulence in Aeromonas hydrophila

Abstract
:1. Introduction
2. Results
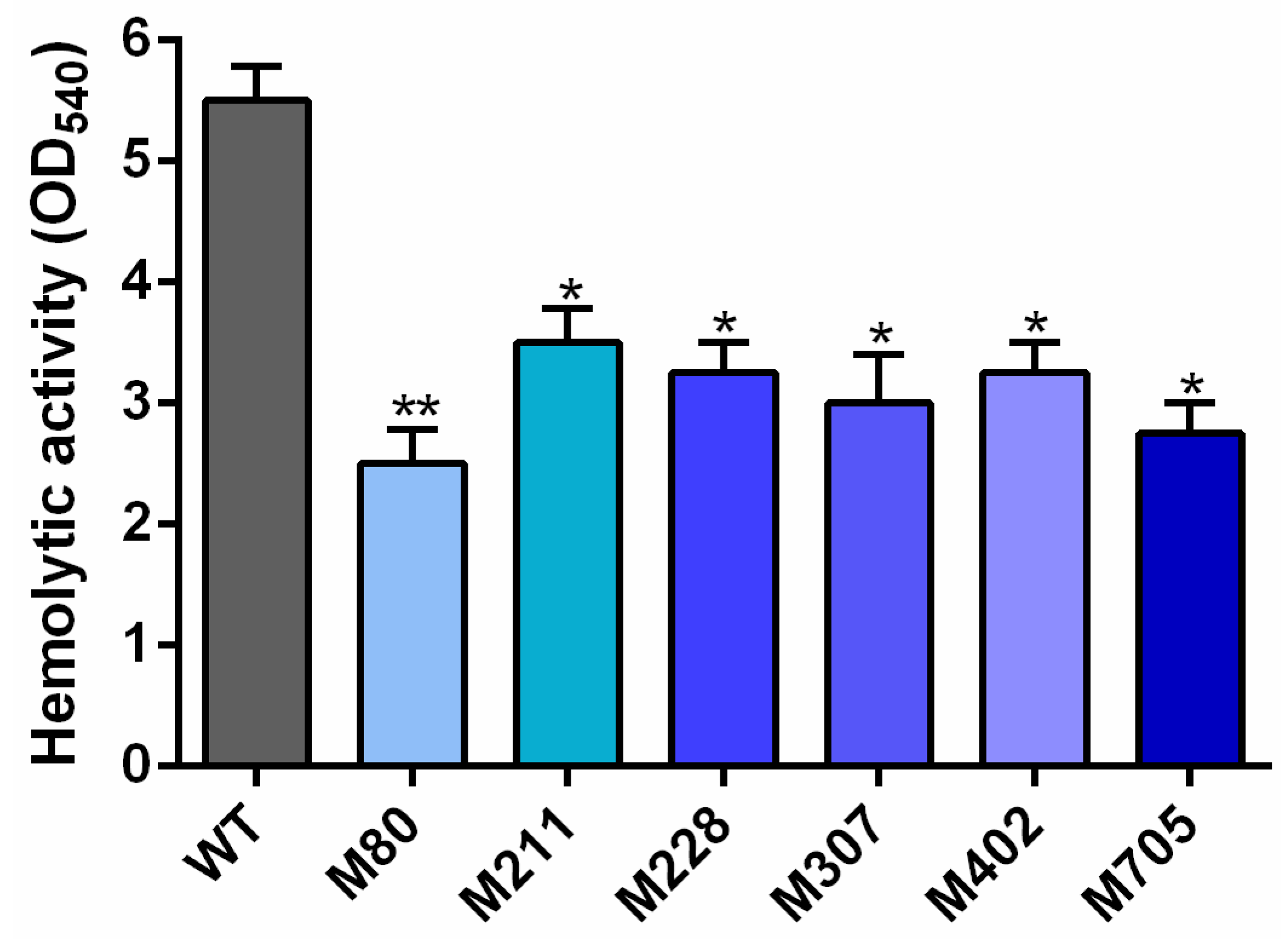
2.1. Isolation of Transposon Mutants with Reduced Hemolytic Activity
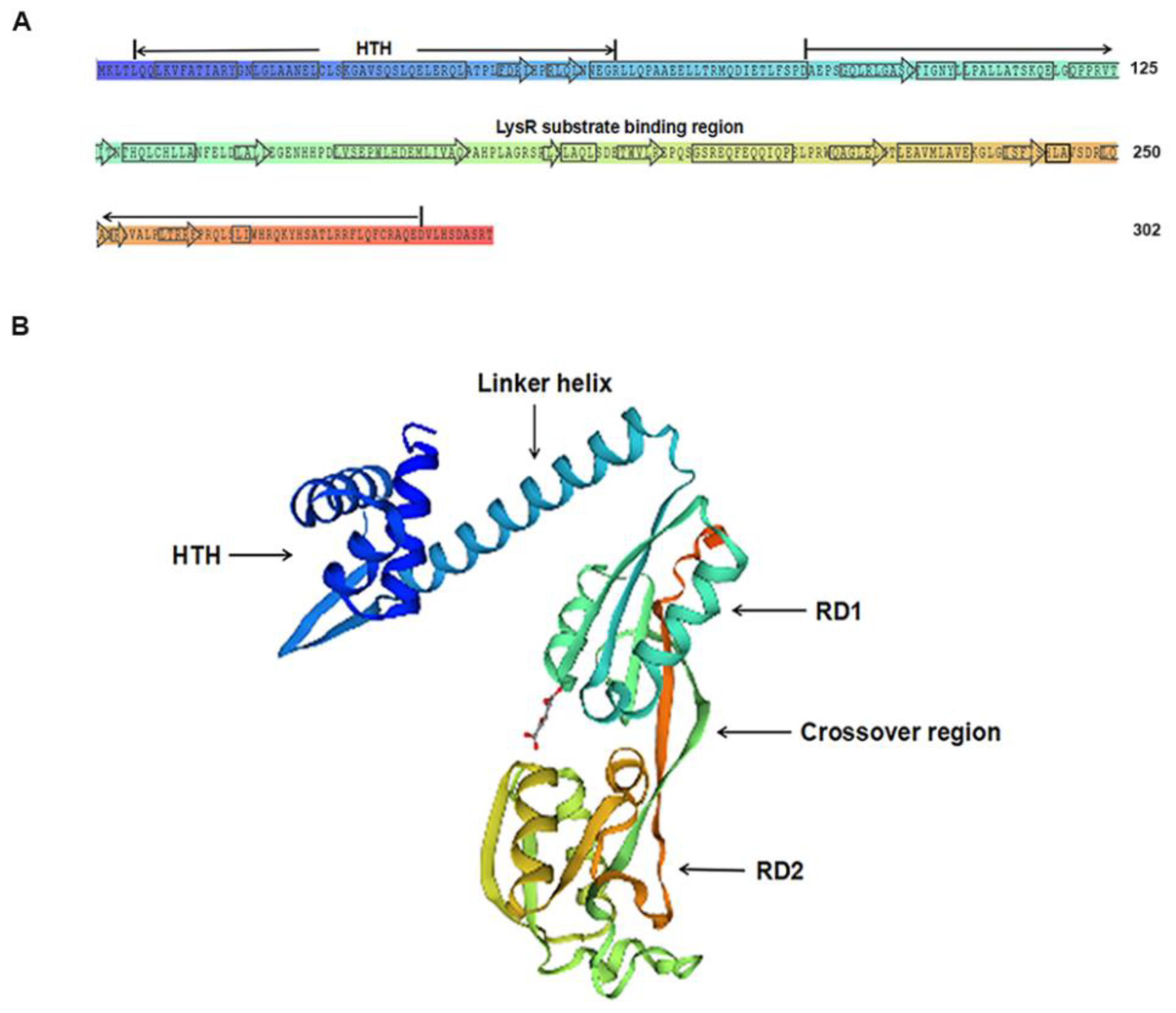
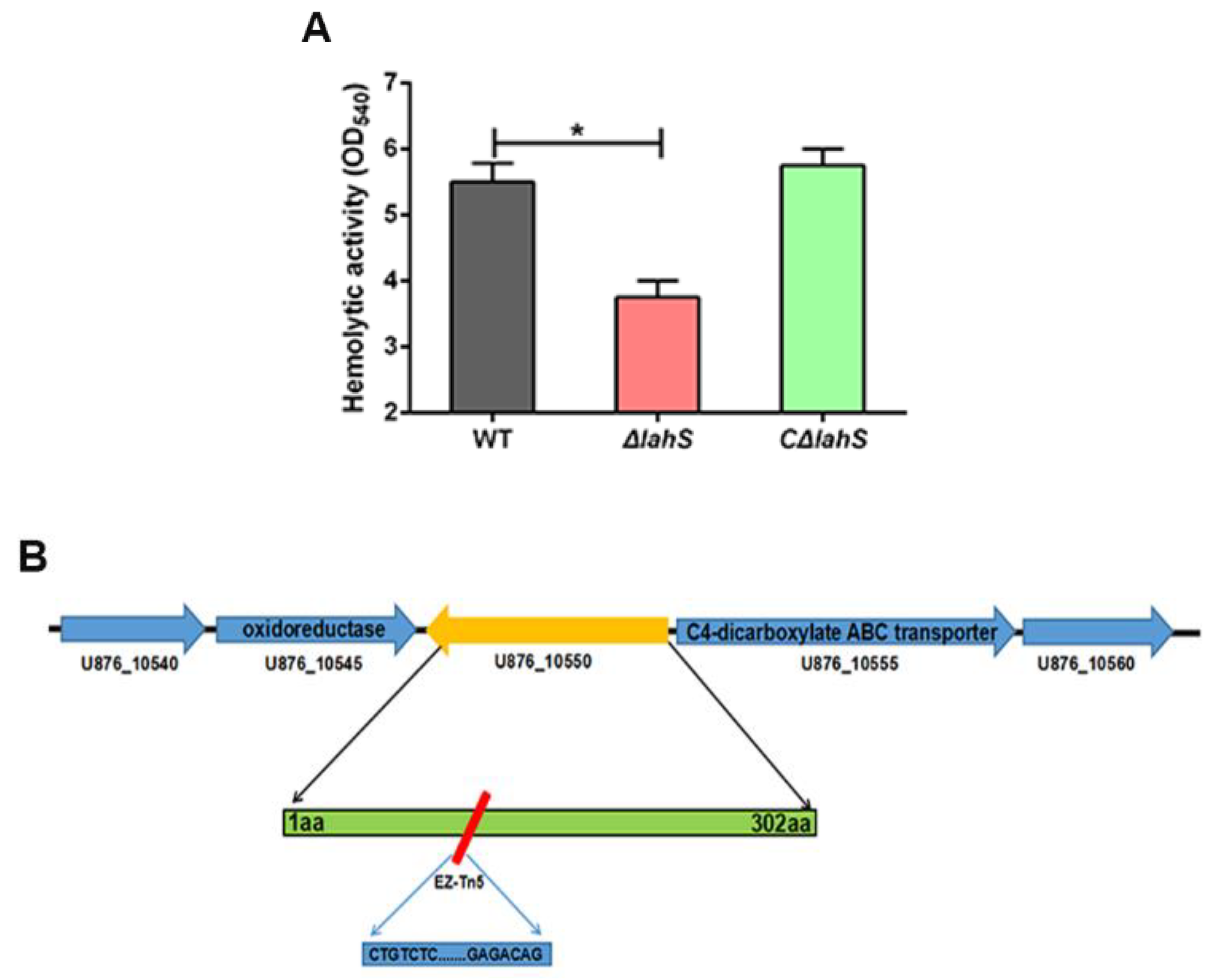
2.2. Effect of the LysR-type Transcriptional Regulator on A. hydrophila Hemolytic Activity
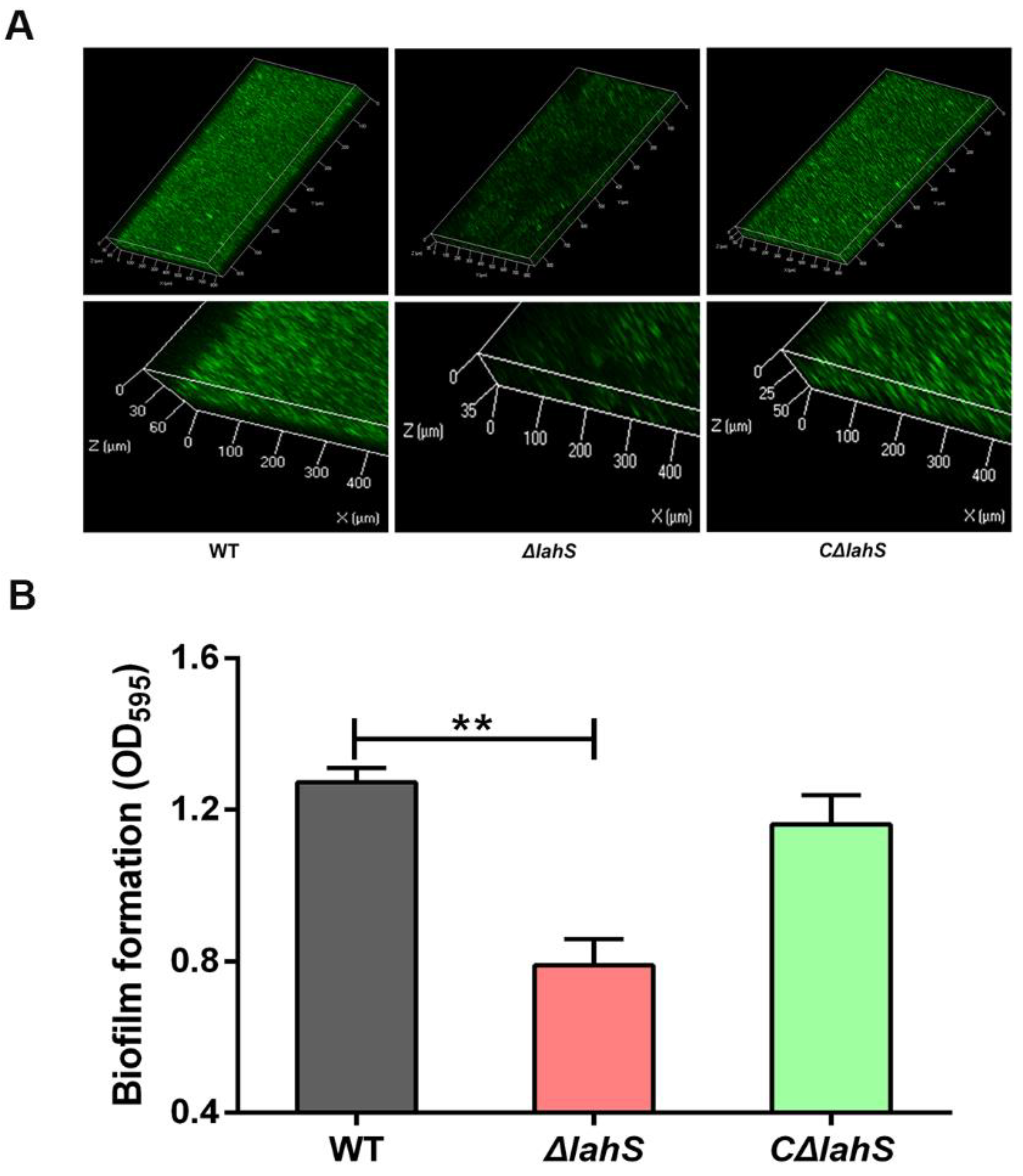
2.3. LahS Is Involved in Biofilm Formation of A. hydrophila NJ-35
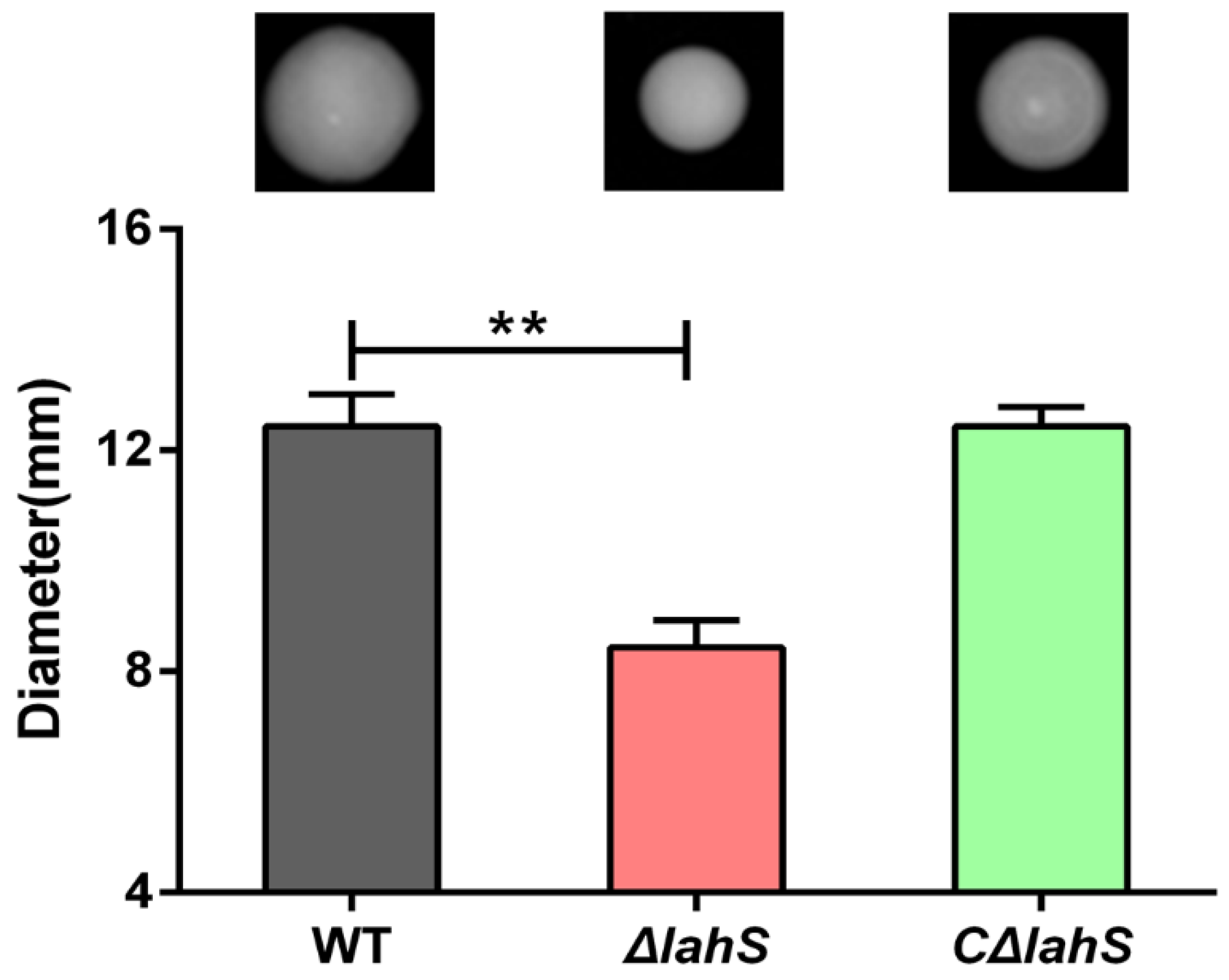
2.4. LahS Influences Motility in A. hydrophila NJ-35
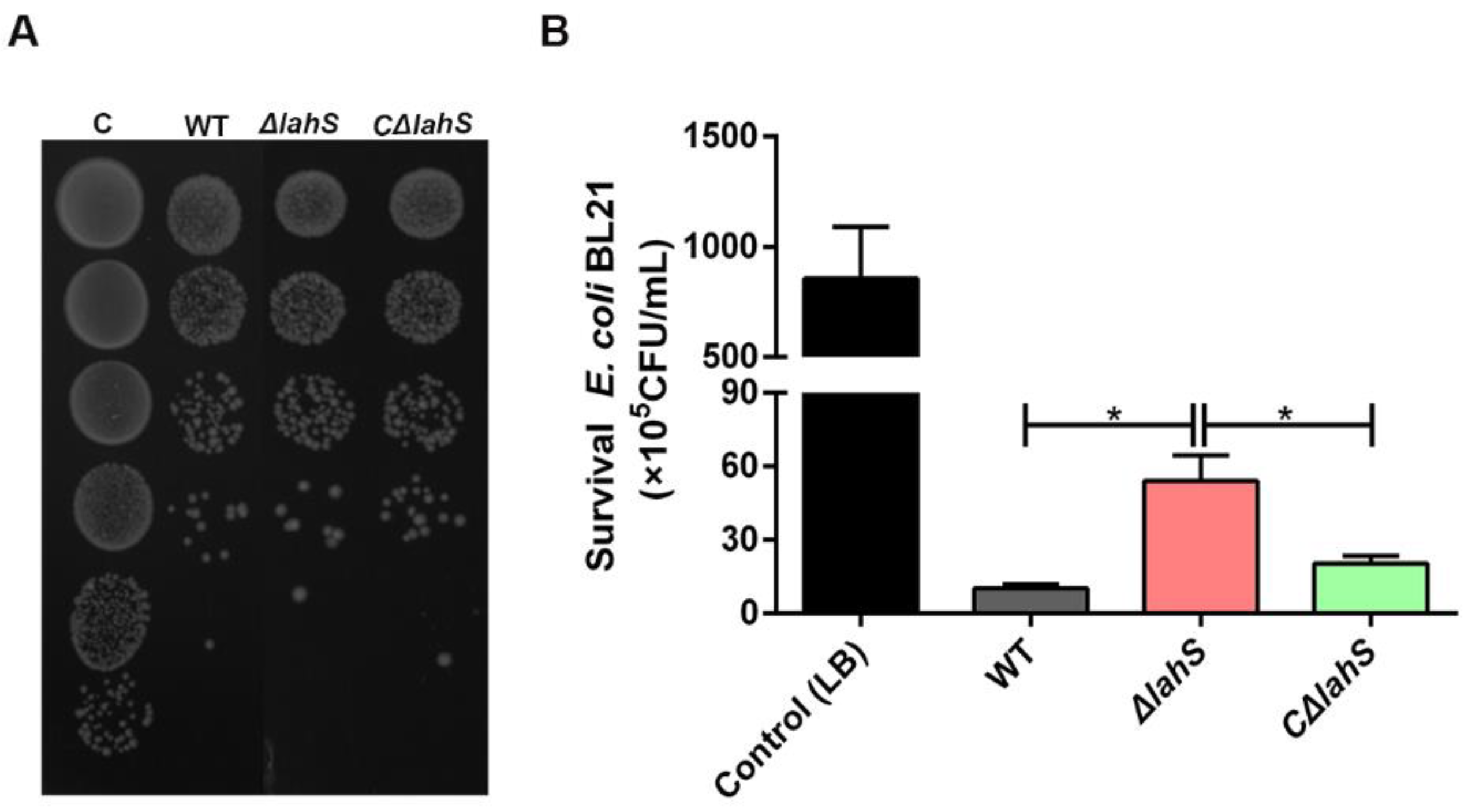
2.5. LahS Contributes to the Antibacterial Activity of A. hydrophila NJ-35
2.6. LahS Plays a Role in the Resistance of A. hydrophila NJ-35 to Oxidative Stress
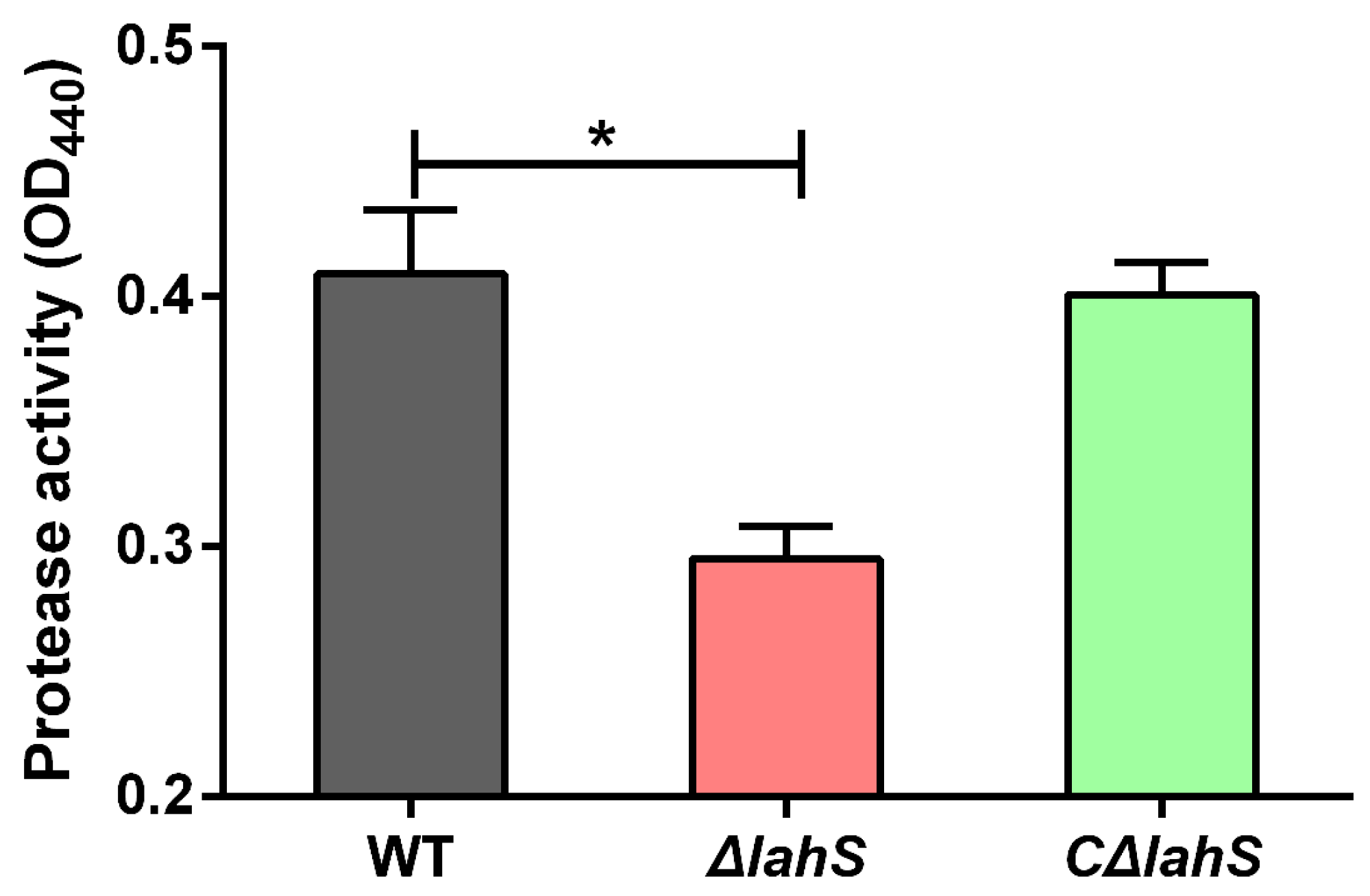
2.7. Deletion of lahS Reduced the Protease Activity of A. hydrophila NJ-35
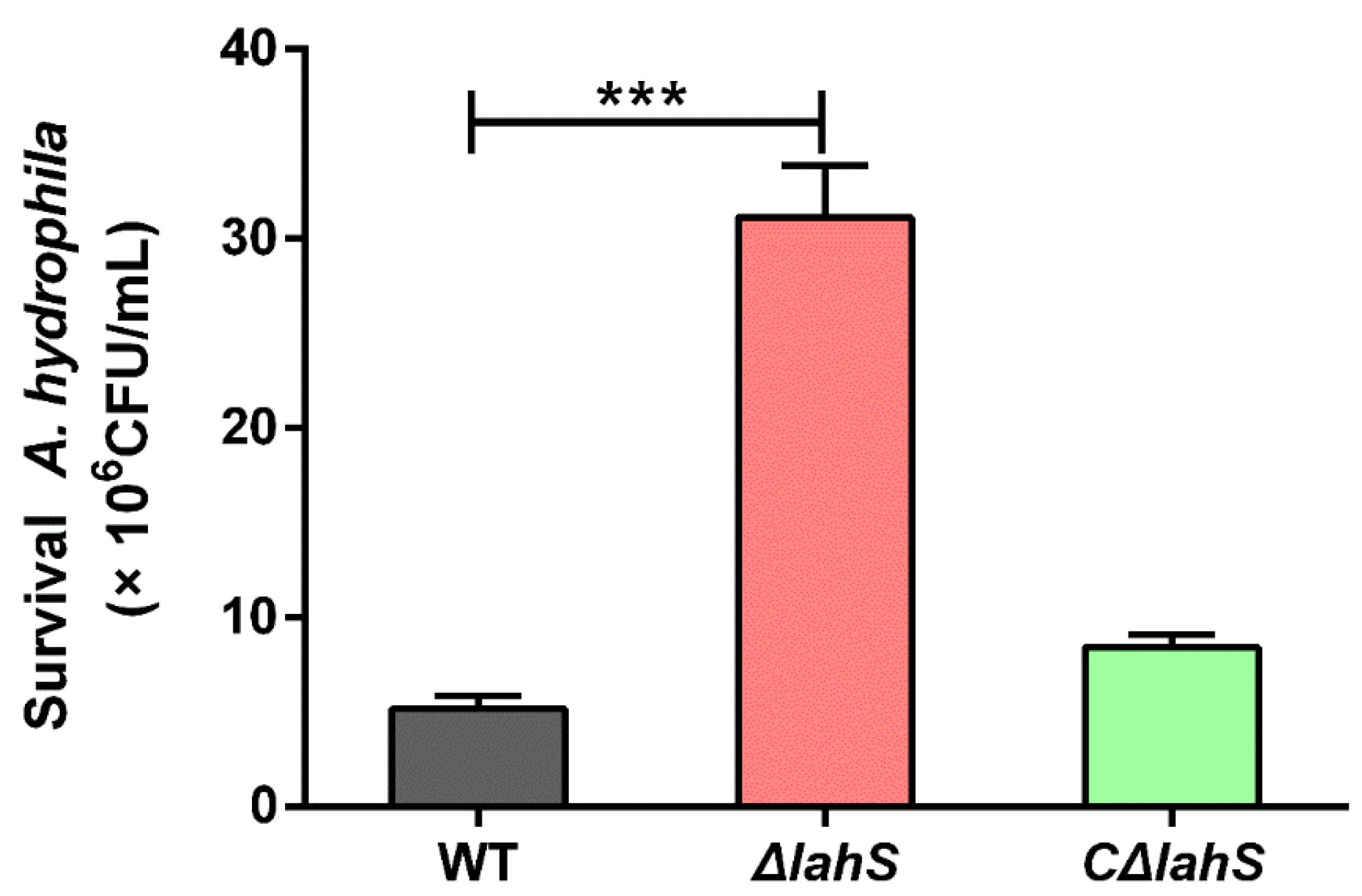
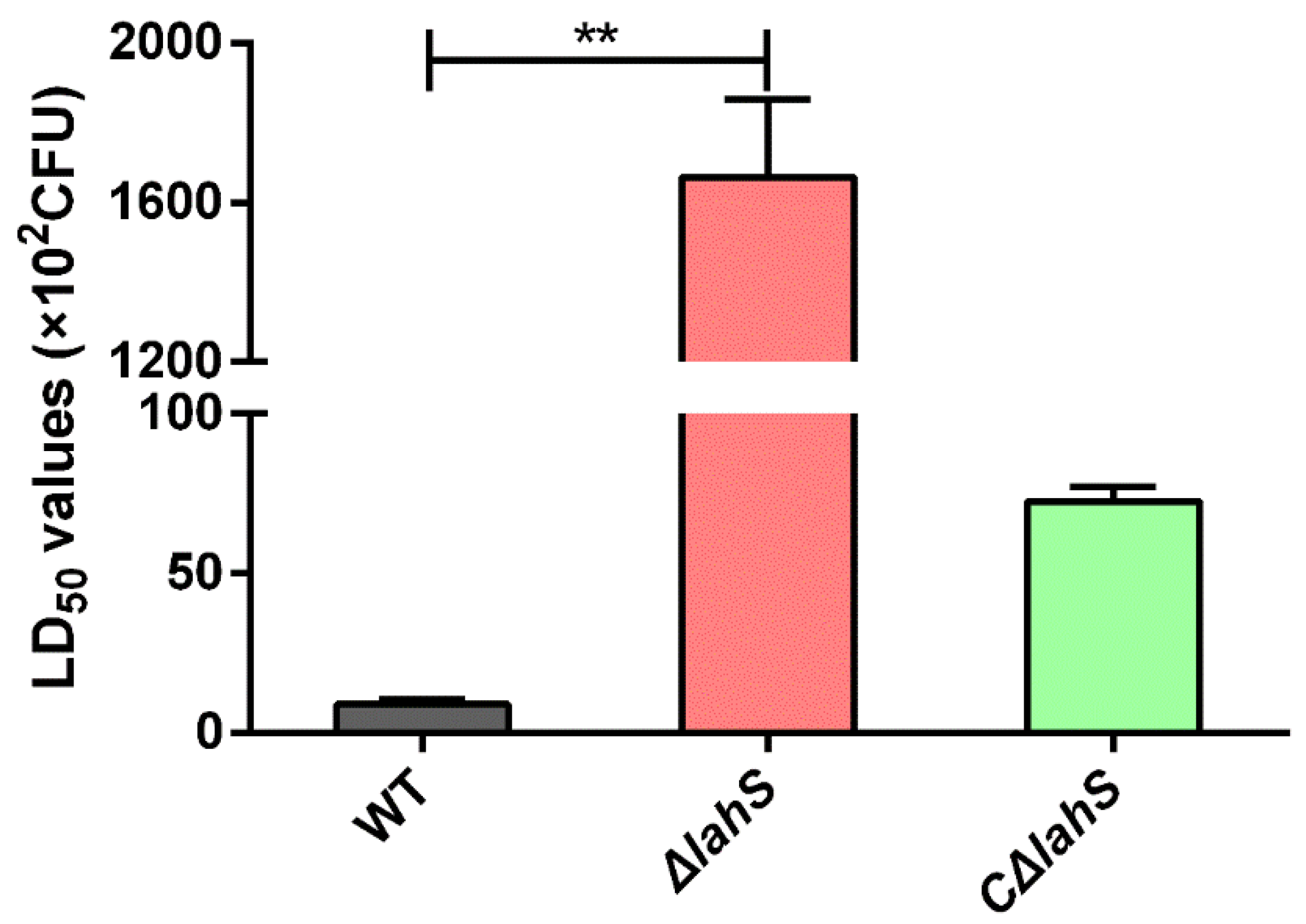
2.8. LahS Is Essential for the Virulence of A. hydrophila in Zebrafish
2.9. Comparative Proteomic Analysis
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids and Growth Conditions
4.2. Screening Transposon Insertion Mutants for Hemolytic Activity
4.3. Identification of Insertion Sites by Tail-PCR
4.4. Construction of a lahS Mutant and Complemented Strains
4.5. Biofilm Formation Assay
4.6. Motility Assay
4.7. Anti-Bacterial Competition Assay
4.8. Oxidative Stress Resistance Test
4.9. Protease Activity
4.10. Determination of LD50 in Zebrafish
4.11. Comparative Proteomic Analysis
4.12. Quantitative Reverse Transcription-PCR (qRT-PCR)
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rasmussen-Ivey, C.R.; Hossain, M.J.; Odom, S.E.; Terhune, J.S.; Hemstreet, W.G.; Shoemaker, C.A.; Zhang, D.; Xu, D.H.; Griffin, M.J.; Liu, Y.J.; et al. Classification of a hypervirulent Aeromonas hydrophila pathotype responsible for epidemic outbreaks in warm-water fishes. Front. Microbiol. 2016, 7, 1615. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.L.; Shaw, J.G. Aeromonas spp. clinical microbiology and disease. J. Infect. 2011, 62, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Tomas, J.M. The main Aeromonas pathogenic factors. ISRN Microbiol. 2012, 2012, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Hamid, R.; Ahmad, A.; Usup, G. Pathogenicity of Aeromonas hydrophila isolated from the Malaysian Sea against coral (Turbinaria sp.) and sea bass (Lates calcarifer). Environ. Sci. Pollut. Res. Int. 2016, 23, 17269–17276. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, D.; Schneper, L.; Kumari, H.; Mathee, K. A dynamic and intricate regulatory network determines Pseudomonas aeruginosa virulence. Nucleic Acids. Res. 2013, 41, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Mou, X.; Spinard, E.J.; Driscoll, M.V.; Zhao, W.; Nelson, D.R. H-NS is a negative regulator of the two hemolysin/cytotoxin gene clusters in Vibrio anguillarum. Infect. Immun. 2013, 81, 3566–3576. [Google Scholar] [CrossRef] [PubMed]
- Rattanama, P.; Thompson, J.R.; Kongkerd, N.; Srinitiwarawong, K.; Vuddhakul, V.; Mekalanos, J.J. Sigma E regulators control hemolytic activity and virulence in a shrimp pathogenic Vibrio harveyi. PLoS ONE 2012, 7, e32523. [Google Scholar] [CrossRef]
- Gao, D.; Li, Y.; Xu, Z.; Sheng, A.; Zheng, E.; Shao, Z.; Liu, N.; Lu, C. The role of regulator Eha in Edwardsiella tarda pathogenesis and virulence gene transcription. Microb. Pathog. 2016, 95, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Heroven, A.K.; Dersch, P. RovM, a novel LysR-type regulator of the virulence activator gene rovA, controls cell invasion, virulence and motility of Yersinia pseudotuberculosis. Mol. Microbiol. 2006, 62, 1469–1483. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Lucas, I.; Gallego-Hernandez, A.L.; Encarnacion, S.; Fernandez-Mora, M.; Martinez-Batallar, A.G.; Salgado, H.; Oropeza, R.; Calva, E. The LysR-type transcriptional regulator LeuO controls expression of several genes in Salmonella enterica serovar Typhi. J. Bacteriol. 2008, 190, 1658–1670. [Google Scholar] [CrossRef] [PubMed]
- Stratmann, T.; Madhusudan, S.; Schnetz, K. Regulation of the yjjQ-bglJ operon, encoding LuxR-type transcription factors, and the divergent yjjP gene by H-NS and LeuO. J. Bacteriol. 2008, 190, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Blumer, C.; Kleefeld, A.; Lehnen, D.; Heintz, M.; Dobrindt, U.; Nagy, G.; Michaelis, K.; Emody, L.; Polen, T.; Rachel, R.; et al. Regulation of type 1 fimbriae synthesis and biofilm formation by the transcriptional regulator LrhA of Escherichia coli. Microbiology 2005, 151, 3287–3298. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Shi, M.; Gao, H. Loss of OxyR reduces efficacy of oxygen respiration in Shewanella oneidensis. Sci. Rep. 2017, 7, 42609. [Google Scholar] [CrossRef] [PubMed]
- Kovacikova, G.; Lin, W.; Skorupski, K. The LysR-type virulence activator AphB regulates the expression of genes in Vibrio cholerae in response to low pH and anaerobiosis. J. Bacteriol. 2010, 192, 4181–4191. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, R.R.; Mooij, M.J.; Reen, F.J.; Lesouhaitier, O.; O′Gara, F. A new regulator of pathogenicity (bvlR) is required for full virulence and tight microcolony formation in Pseudomonas aeruginosa. Microbiology 2014, 160, 1488–1500. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Yamamoto, K.; Ishihama, A. Involvement of the leucine response transcription factor LeuO in regulation of the genes for sulfa drug efflux. J. Bacteriol. 2009, 191, 4562–4571. [Google Scholar] [CrossRef] [PubMed]
- Honda, N.; Iyoda, S.; Yamamoto, S.; Terajima, J.; Watanabe, H. LrhA positively controls the expression of the locus of enterocyte effacement genes in enterohemorrhagic Escherichia coli by differential regulation of their master regulators PchA and PchB. Mol. Microbiol. 2009, 74, 1393–1411. [Google Scholar] [CrossRef] [PubMed]
- Bina, X.R.; Howard, M.F.; Ante, V.M.; Bina, J.E. Vibrio cholerae LeuO links the ToxR regulon to expression of lipid A remodeling genes. Infect. Immun. 2016, 84, 3161–3171. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.P.; Wen, Y.P.; Wen, X.T.; Cao, S.J.; Huang, X.B.; Wu, R.; Zhao, Q.; Liu, M.F.; Huang, Y.; Yan, Q.G.; et al. OxyR of Haemophilus parasuis is a global transcriptional regulator important in oxidative stress resistance and growth. Gene 2018, 643, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.D.; Xie, X.; Dong, Y.H.; Du, H.C.; Wang, N.N.; Lu, C.P.; Liu, Y.J. Identification of novel virulence-related genes in Aeromonas hydrophila by screening transposon mutants in a Tetrahymena infection model. Vet. Microbiol. 2017, 199, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. BLAST: Basic Local Alignment Search Tool. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cg (accessed on 11 May 2018).
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART. Available online: http://smart.embl-heidelberg.de/ (accessed on 28 July 2018).
- Guex, N.; Peitsch, M.C. SWISS-MODEL. Available online: https://www.swissmodel.expasy.org/ (accessed on 7 September 2018).
- Ayala, J.C.; Wang, H.; Benitez, J.A.; Silva, A.J. Molecular basis for the differential expression of the global regulator VieA in Vibrio cholerae biotypes directed by H-NS, LeuO and quorum sensing. Mol. Microbiol. 2018, 107, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Lehnen, D.; Blumer, C.; Polen, T.; Wackwitz, B.; Wendisch, V.F.; Unden, G. LrhA as a new transcriptional key regulator of flagella, motility and chemotaxis genes in Escherichia coli. Mol. Microbiol. 2002, 45, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Canals, R.; Ramirez, S.; Vilches, S.; Horsburgh, G.; Shaw, J.G.; Tomas, J.M.; Merino, S. Polar flagellum biogenesis in Aeromonas hydrophila. J. Bacteriol. 2006, 188, 542–555. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm formation as microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef] [PubMed]
- Casper-Lindley, C.; Yildiz, F.H. VpsT is a transcriptional regulator required for expression of vps biosynthesis genes and the development of rugose colonial morphology in Vibrio cholerae O1 EI Tor. J. Bacteriol. 2006, 188, 6044. [Google Scholar] [CrossRef]
- Wang, K.; Liu, E.; Song, S.; Wang, X.; Zhu, Y.; Ye, J.; Zhang, H. Characterization of Edwardsiella tarda rpoN: Roles in σ70 family regulation, growth, stress adaption and virulence toward fish. Arch. Microbiol. 2012, 194, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Kirov, S.M.; Castrisios, M.; Shaw, J.G. Aeromonas flagella (polar and lateral) are enterocyte adhesins that contribute to biofilm formation on surfaces. Infect. Immun. 2004, 72, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xie, J.P. Role and regulation of bacterial LuxR-like regulators. J. Cell. Biochem. 2011, 112, 2694–2702. [Google Scholar] [CrossRef] [PubMed]
- Bi, Z.X.; Liu, Y.J.; Lu, C.P. Contribution of AhyR to virulence of Aeromonas hydrophila J-1. Res. Vet. Sci. 2007, 83, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.T.; Wang, X.T.; Mao, Q.Q.; Xu, R.J.; Zhou, X.H.; Ma, Y.; Liu, Q.; Zhang, Y.X.; Wang, Q.Y. VqsA, a novel LysR-type transcriptional regulator, coordinates quorum sensing (QS) and is controlled by QS to regulate virulence in the pathogen Vibrio alginolyticus. Appl. Environ. Microbiol. 2018, 84, e00444-18. [Google Scholar] [CrossRef] [PubMed]
- Jo, I.; Chung, I.Y.; Bae, H.W.; Kim, J.S.; Song, S.; Cho, Y.H.; Ha, N.C. Structural details of the OxyR peroxide-sensing mechanism. Proc. Natl. Acad. Sci. USA 2015, 112, 6443–6448. [Google Scholar] [CrossRef] [PubMed]
- Hennequin, C.; Forestier, C. oxyR, a LysR-type regulator involved in Klebsiella pneumoniae mucosal and abiotic colonization. Infect. Immun. 2009, 77, 5449–5457. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, H.; Inui, M.; Yukawa, H. OxyR acts as a transcriptional repressor of hydrogen peroxide-inducible antioxidant genes in Corynebacterium glutamicum R. FEBS. J. 2013, 280, 3298–3312. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.M.; Schellhorn, H.E. Regulators of oxidative stress response genes in Escherichia coli and their functional conservation in bacteria. Arch. Biochem. Biophys. 2012, 525, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.M.; Dong, Y.Y.; Luo, Q.X.; Li, N.; Wu, G.F.; Gao, H.C. Protection from oxidative stress relies mainly on derepression of OxyR-dependent KatB and Dps in Shewanella oneidensis. J. Bacteriol. 2014, 196, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.D.; Jiang, J.W.; Xie, X.; Wu, Y.F.; Dong, Y.H.; Kwok, A.H.Y.; Zhang, W.; Yao, H.C.; Lu, C.P.; Leung, F.C.; et al. Novel insights into the pathogenicity of epidemic Aeromonas hydrophila ST251 clones from comparative genomics. Sci. Rep. 2015, 5, 9833. [Google Scholar] [CrossRef] [PubMed]
- Melton-Witt, J.A.; McKay, S.L.; Portnoy, D.A. Development of a single-gene, signature-tag-based approach in combination with alanine mutagenesis to identify listeriolysin O residues critical for the in vivo survival of Listeria monocytogenes. Infect. Immun. 2012, 80, 2221–2230. [Google Scholar] [CrossRef] [PubMed]
- Abolghait, S.K. Suicide plasmid-dependent IS1-element untargeted integration into Aeromonas veronii bv. sobria generates brown pigment-producing and spontaneous pelleting mutant. Curr. Microbiol. 2013, 67, 91–99. [Google Scholar] [PubMed]
- Morales, V.M.; Backman, A.; Bagdasarian, M. A series of wide-host-range low-copy-number vectors that allow direct screening for recombinants. Gene 1991, 97, 39–47. [Google Scholar] [CrossRef]
- Khajanchi, B.K.; Kozlova, E.V.; Sha, J.; Popov, V.L.; Chopra, A.K. The two-component QseBC signalling system regulates in vitro and in vivo virulence of Aeromonas hydrophila. Microbiology 2012, 158, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Du, H.C.; Pang, M.D.; Dong, Y.H.; Wu, Y.F.; Wang, N.N.; Liu, J.; Awan, F.; Lu, C.P.; Liu, Y.J. Identification and characterization of an Aeromonas hydrophila oligopeptidase gene pepF negatively related to biofilm formation. Front. Microbiol. 2016, 7, 1497. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A.; Pratt, L.A.; Watnick, P.I.; Newman, D.K.; Weaver, V.B.; Kolter, R. Genetic approaches to study of biofilms. Methods Enzymol. 1999, 310, 91–109. [Google Scholar] [PubMed]
- Grim, C.J.; Kozlova, E.V.; Sha, J.; Fitts, E.C.; van Lier, C.J.; Kirtley, M.L.; Joseph, S.J.; Read, T.D.; Burd, E.M.; Tall, B.D.; et al. Characterization of Aeromonas hydrophila wound pathotypes by comparative genomic and functional analyses of virulence genes. mBio 2013, 4, e00064-13. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Dong, Y.H.; Wang, N.N.; Li, S.G.; Yang, Y.Y.; Wang, Y.; Awan, F.; Lu, C.P.; Liu, Y.J. Tetrahymena thermophila predation enhances environmental adaptation of the carp pathogenic strain Aeromonas hydrophila NJ-35. Front. Cell. Infect. Microbiol. 2018, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.D.; Lin, X.Q.; Hu, M.; Li, J.; Lu, C.P.; Liu, Y.J. Tetrahymena: An alternative model host for evaluating virulence of Aeromonas strains. PLoS ONE 2012, 7, e48922. [Google Scholar] [CrossRef] [PubMed]
- Stanic, M. A simplification of the estimation of the 50 percent endpoints according to the Reed and Muench method. Pathol. Microbiol. (Basel) 1963, 26, 298–302. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutant | Locus Tag | Insertion Site/Gene Length (bp) a | Function b |
|---|---|---|---|
| M80 | U876_21575 | 2/480 | hypothetical protein |
| M211 | U876_23300 | 125/1587 | PTS alpha-glucoside transporter subunit IIBC |
| M228 | U876_22535 | 102/246 | hypothetical protein |
| M307 | U876_10550 | 440/909 | LysR family transcriptional regulator |
| M402 | U876_02250 | 330/516 | hypothetical protein |
| M705 | U876_01475 | 711/857 | Arac family transcriptional regulator |
| Locus Tag | Predicted Function | Fold Change |
|---|---|---|
| Up-regulated proteins | ||
| U876_17600 | Hypothetical protein | 28.34991961 |
| U876_01465 | Catalase | 11.31448355 |
| U876_16540 | Peroxidase | 2.09906885 |
| U876_04415 | Cytochrome C biogenesis protein CcsA | 2.043010753 |
| U876_04895 | Triose-phosphate isomerase | 2.939215876 |
| U876_07195 | Fructose-6-phosphate aldolase | 2.67060636 |
| U876_05460 | Penicillin-sensitive transpeptidase | 6.074663579 |
| U876_13305 | Transcriptional regulator | 2.236916529 |
| U876_10585 | Prolyl-tRNA synthetase | 2.143335753 |
| U876_23535 | Chromosome partitioning protein ParA | 2.1254663 |
| Down-regulated proteins | ||
| U876_07270 | Flagellar hook protein FlgE | 0.123192009 |
| U876_16245 | Flagellar hook-length control protein FliK | 0.396639518 |
| U876_07265 | Flagellar hook capping protein FlgD | 0.446402163 |
| U876_14260 | Flagellar hook-associated protein FliD | 0.49944046 |
| U876_14545 | Phospho-2-dehydro-3-deoxyheptonate aldolase | 0.021640145 |
| U876_09970 | Phospho-2-dehydro-3-deoxyheptonate aldolasee | 0.067411226 |
| U876_11610 | LuxR family transcriptional regulator | 0.279023788 |
| U876_07335 | Fis family transcriptional regulator | 0.366104356 |
| U876_14125 | Glycine cleavage system protein T | 0.014741915 |
| U876_08310 | Hypothetical protein | 0.22058777 |
| U876_14865 | Cytochrome C | 0.243506128 |
| U876_17595 | Hydroxylamine reductase | 0.267712471 |
| U876_13425 | Thioredoxin reductase | 0.280916369 |
| U876_14870 | Nitrate reductase | 0.304735753 |
| U876_22700 | YeeE/YedE family protein | 0.381751269 |
| U876_14250 | Hypothetical protein | 0.401170621 |
| U876_07930 | Cytochrome c biogenesis protein CcmH | 0.426902415 |
| U876_15300 | Hypothetical protein | 0.43795805 |
| U876_13000 | Methionine gamma-lyase | 0.448173659 |
| U876_01200 | Cytochrome C | 0.45592439 |
| U876_16185 | RNA polymerase sigma 70 | 0.457438622 |
| U876_16300 | Peptidase C80 | 0.481165879 |
| U876_01650 | Single-stranded DNA-binding protein | 0.487195726 |
| U876_09765 | Electron transporter HydN | 0.494528201 |
| Strain or Plasmid | Description a | Source or Reference |
|---|---|---|
| Strains | ||
| NJ-35 | Wild-type, isolated from diseased crucian carp, in China | Collected in our laboratory |
| SM10 | E. coli strain, λpir+, Kanr | [41] |
| BL21 | E. coli strain, F−, ompT, hsdS (rB−mB−), gal, dcm (DE3) | Invitrogen |
| ΔlahS | lahS deletion mutant from NJ-35 | This study |
| CΔlahS | ΔlahS complemented with pMMB-lahS | This study |
| Plasmid | ||
| pET 28a (+) | Kanr, F1 origin | Novagen |
| pYAK1 | R6K-ori suicide vector, SacB+, Cmr | [42] |
| pYAK1-lahS | pYAK1 carrying the flanking sequence of lahS, Cmr | This study |
| pMMB207 | Low-copy-number vector, Cmr | [43] |
| pMMB-lahS | Plasmid pMMB207 carrying the complete ORF of lahS | This study |
| Primer | Sequence (5′–3′) |
|---|---|
| lahS-1 | CAGGTCGACTCTAGAGGATCCTGGAGGCAATGAAGGTGA |
| lahS-2 | ATATCCAAACGCCAATGGGATGATCGG |
| lahS-3 | TCCCATTGGCGTTTGGATATCCTGAACGATT |
| lahS-4 | GAGCTCGGTACCCGGGGATCC CGCTCACGGTCAGGTAAC |
| lahS-C-F | GAGCTCGGTACCCGGGGATCCATGAAGCTGACCCTGCAAC |
| lahS-C-R | CAGGTCGACTCTAGAGGATCCTCAGGTGCGGCTGGC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Wang, Y.; Liu, J.; Ma, S.; Awan, F.; Lu, C.; Liu, Y. Discovery of lahS as a Global Regulator of Environmental Adaptation and Virulence in Aeromonas hydrophila. Int. J. Mol. Sci. 2018, 19, 2709. https://doi.org/10.3390/ijms19092709
Dong Y, Wang Y, Liu J, Ma S, Awan F, Lu C, Liu Y. Discovery of lahS as a Global Regulator of Environmental Adaptation and Virulence in Aeromonas hydrophila. International Journal of Molecular Sciences. 2018; 19(9):2709. https://doi.org/10.3390/ijms19092709
Chicago/Turabian StyleDong, Yuhao, Yao Wang, Jin Liu, Shuiyan Ma, Furqan Awan, Chengping Lu, and Yongjie Liu. 2018. "Discovery of lahS as a Global Regulator of Environmental Adaptation and Virulence in Aeromonas hydrophila" International Journal of Molecular Sciences 19, no. 9: 2709. https://doi.org/10.3390/ijms19092709