The Long Non-Coding RNA MIR503HG Enhances Proliferation of Human ALK-Negative Anaplastic Large-Cell Lymphoma


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
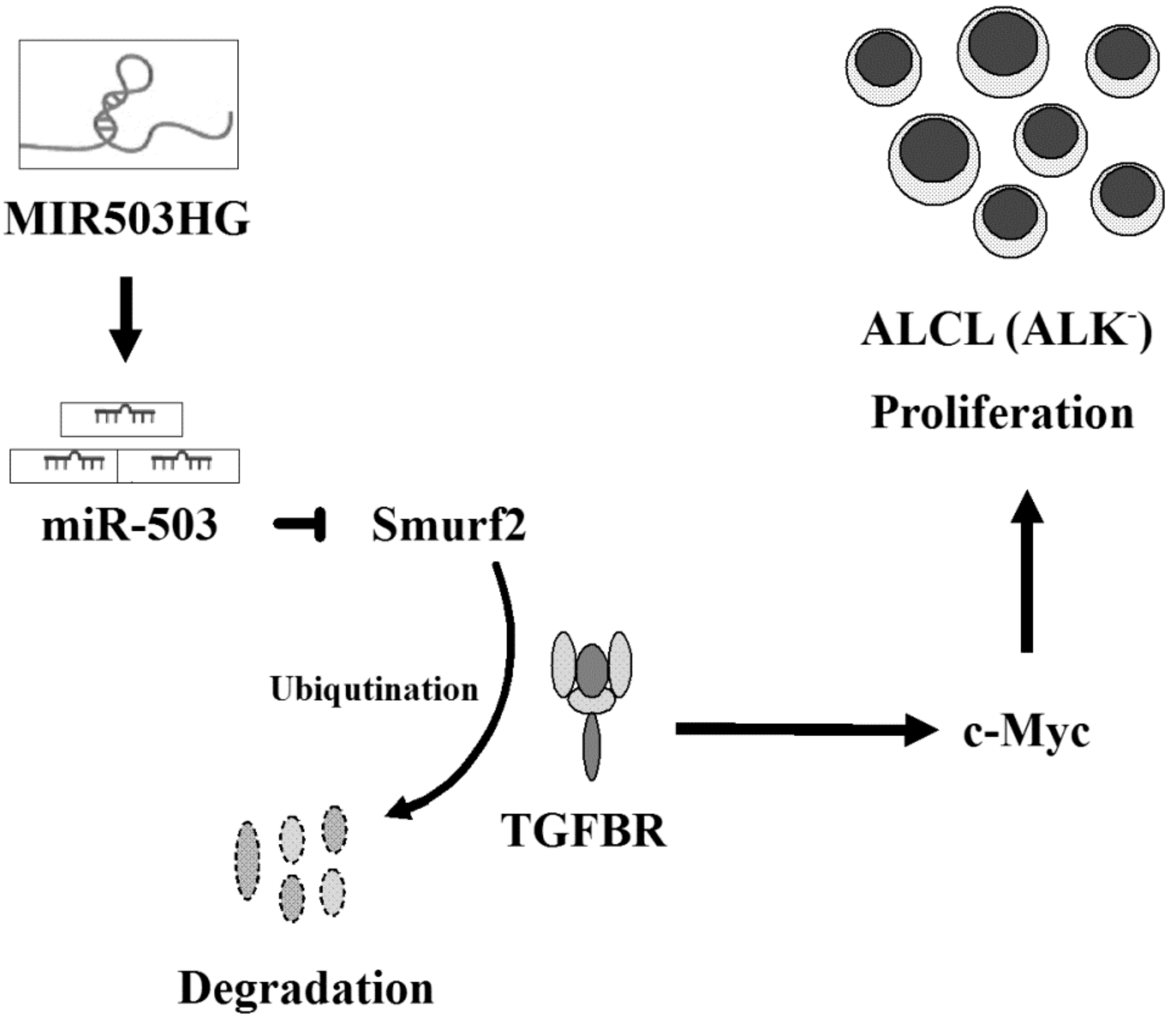
Abstract
:1. Introduction
2. Results
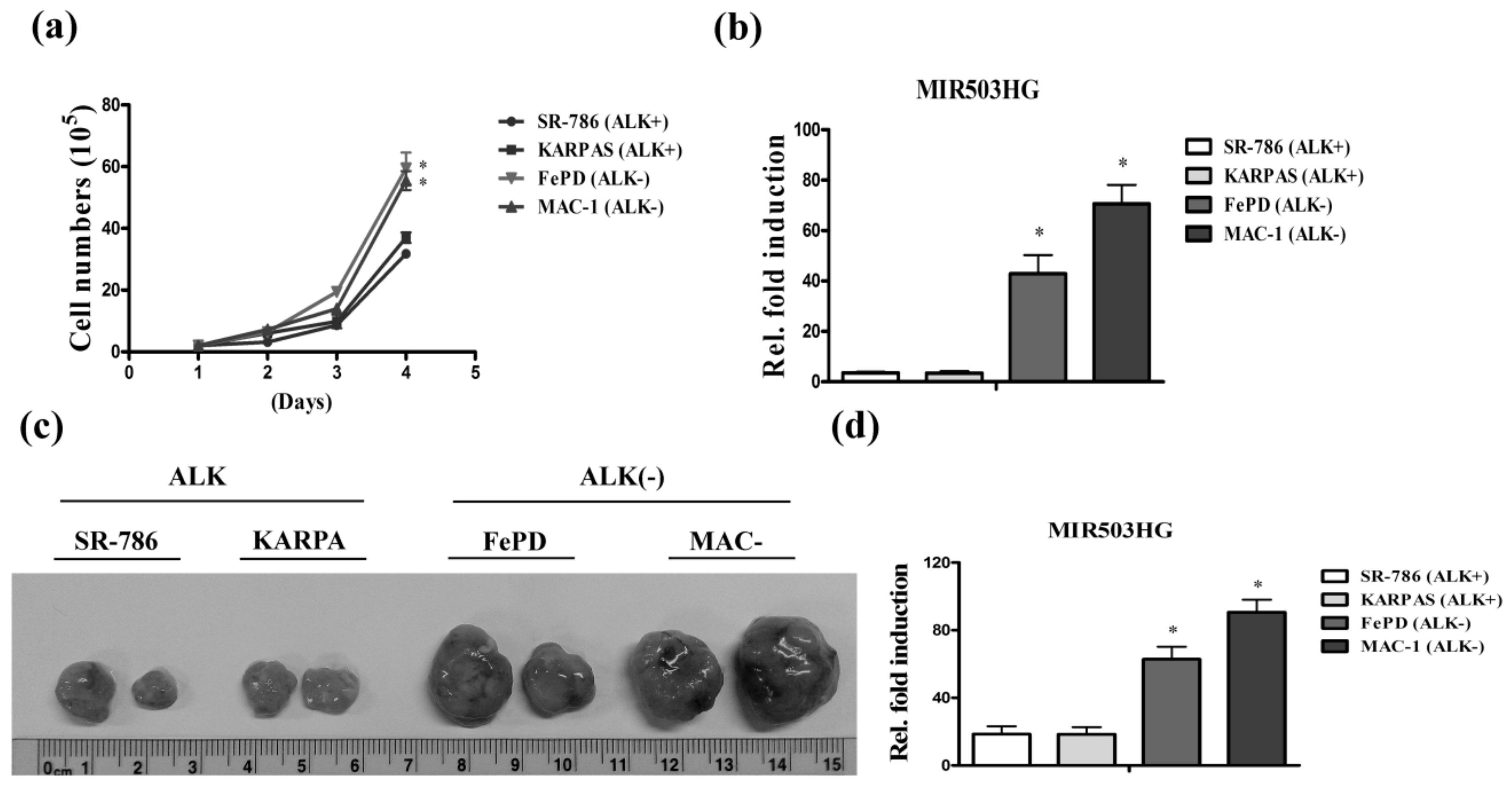
2.1. High Expression of MIR503HG in ALK-Negative ALCL Is Associated with Tumor Proliferation
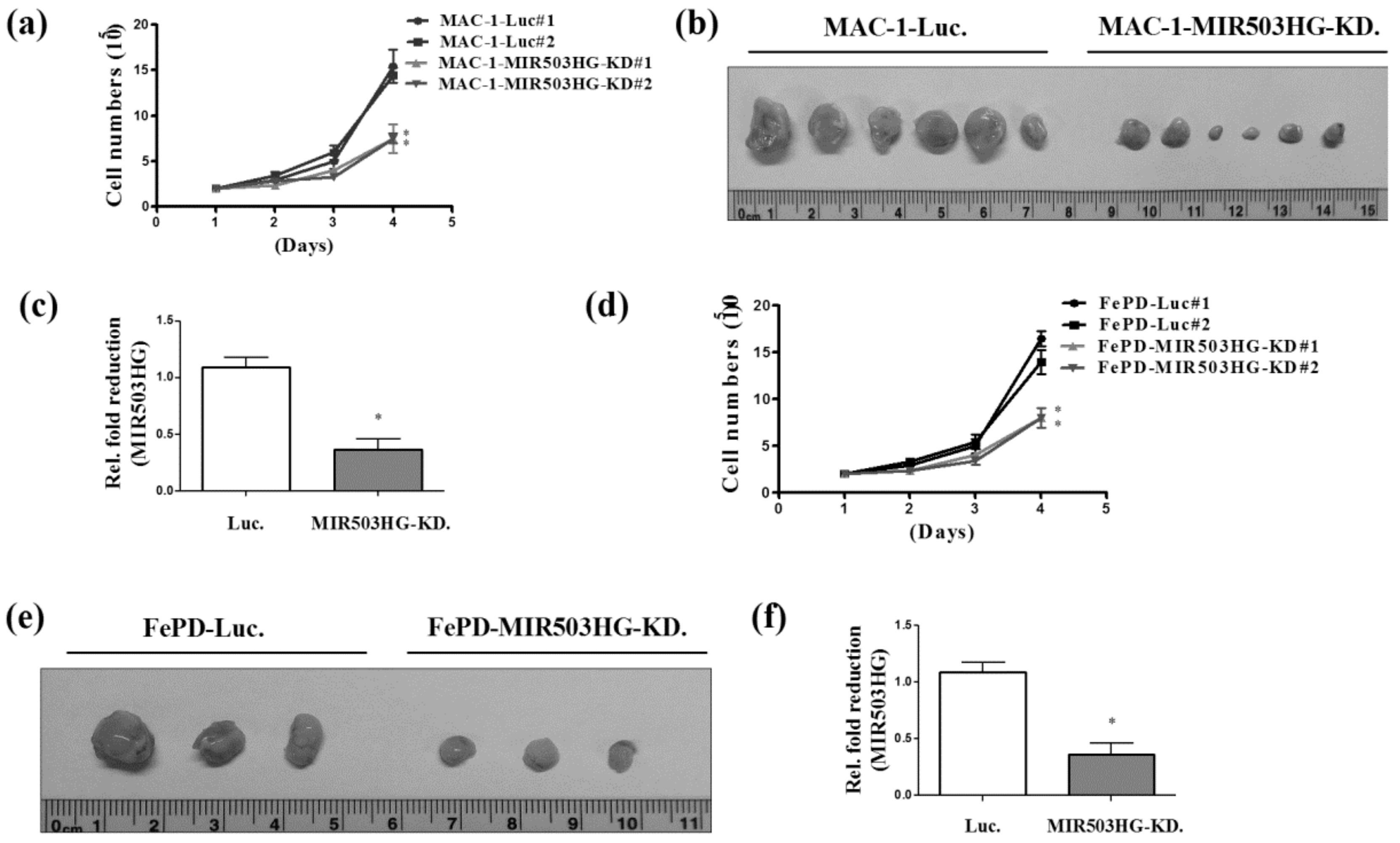
2.2. MIR503HG Depletion Suppresses ALK-Negative ALCL Proliferation In Vitro and In Vivo
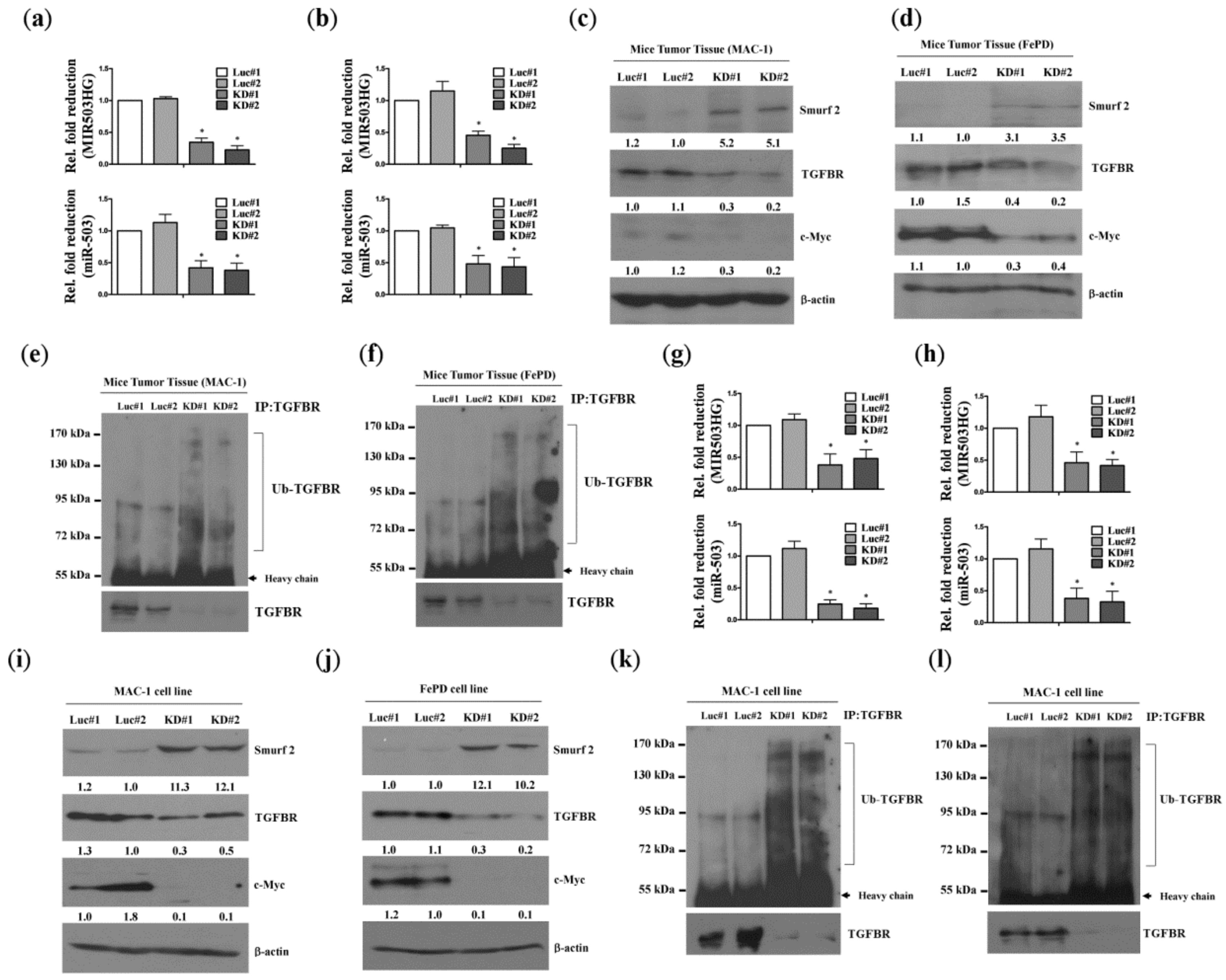
2.3. MIR503HG Depletion Acts through Suppression of miR-503 to Promote Degradation of the TGF-β Receptor In Vitro and In Vivo
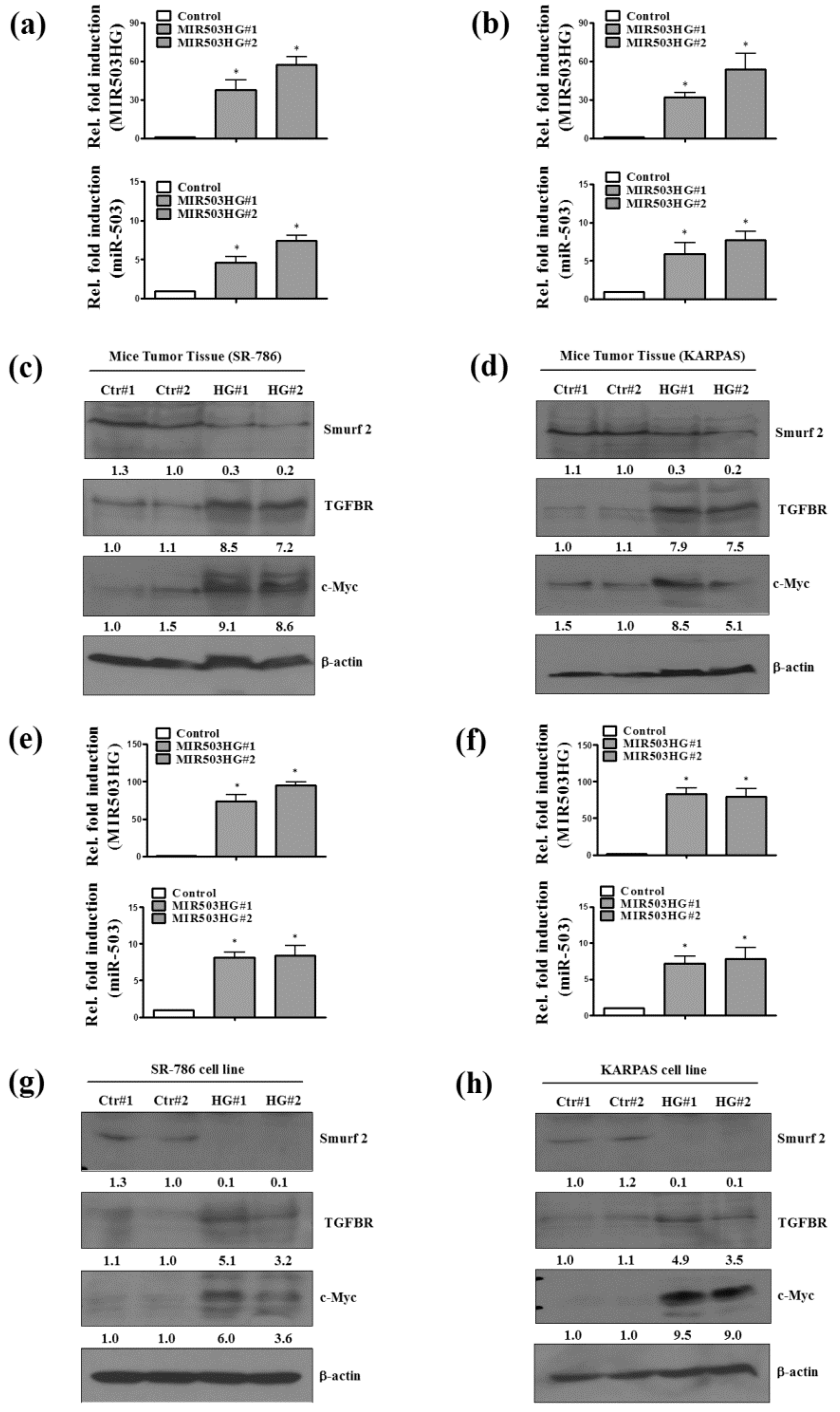
2.4. MIR503HG Overexpression Promotes ALK-Positive ALCL Growth In Vitro and In Vivo
2.5. MIR503HG Overexpression Acts through Induction of miR-503 to Stabilize TGFBR and Enhance Cell Proliferation In Vitro and In Vivo
3. Discussion
4. Materials and Methods
4.1. Mouse Xenograft Model
4.2. Quantitative Reverse Transcription-Polymerase Chain Reaction
4.3. Cell Culture
4.4. Immunoblotting
4.5. Establishment of Stable MIR503HG-Knockdown Cell Lines
4.6. Establishment of Stable MIR503HG-Overexpressing Cell Lines
4.7. Proliferation Assay
4.8. Ubiquitination Assay
4.9. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Blum, K.A. CD30: Seeing is not always believing. Blood 2015, 125, 1358–1359. [Google Scholar] [CrossRef] [PubMed]
- Chiarle, R.; Voena, C.; Ambrogio, C.; Piva, R.; Inghirami, G. The anaplastic lymphoma kinase in the pathogenesis of cancer. Nat. Rev. Cancer 2008, 8, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Cillessen, S.A.; Hijmering, N.J.; Moesbergen, L.M.; Vos, W.; Verbrugge, S.E.; Jansen, G.; Visser, O.J.; Oudejans, J.J.; Meijer, C.J. ALK-negative anaplastic large cell lymphoma is sensitive to bortezomib through Noxa upregulation and release of Bax from Bcl-2. Haematologica 2015, 100, e365–e368. [Google Scholar] [CrossRef] [PubMed]
- Ceccon, M.; Merlo, M.E.; Mologni, L.; Poggio, T.; Varesio, L.M.; Menotti, M.; Bombelli, S.; Rigolio, R.; Manazza, A.D.; Di Giacomo, F.; et al. Excess of NPM-ALK oncogenic signaling promotes cellular apoptosis and drug dependency. Oncogene 2015, 35, 3854–3865. [Google Scholar] [CrossRef] [PubMed]
- Choudhari, R.; Minero, V.G.; Menotti, M.; Pulito, R.; Brakebusch, C.; Compagno, M.; Voena, C.; Ambrogio, C.; Chiarle, R. Redundant and nonredundant roles for Cdc42 and Rac1 in lymphomas developed in NPM-ALK transgenic mice. Blood 2016, 127, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Crescenzo, R.; Abate, F.; Lasorsa, E.; Tabbo, F.; Gaudiano, M.; Chiesa, N.; Di Giacomo, F.; Spaccarotella, E.; Barbarossa, L.; Ercole, E.; et al. Convergent mutations and kinase fusions lead to oncogenic STAT3 activation in anaplastic large cell lymphoma. Cancer Cell 2015, 27, 516–532. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, A.J.; Govi, S.; Pileri, S.A.; Savage, K.J. Anaplastic large cell lymphoma, ALK-negative. Crit. Rev. Oncol. Hematol. 2013, 85, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Mereu, E.; Pellegrino, E.; Scarfo, I.; Inghirami, G.; Piva, R. The heterogeneous landscape of ALK negative ALCL. Oncotarget 2017, 8, 18,525–18,536. [Google Scholar] [CrossRef] [PubMed]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Lau, E. Non-coding RNA: Zooming in on lncRNA functions. Nat. Rev. Genet. 2014, 15, 574–575. [Google Scholar] [CrossRef] [PubMed]
- Seton-Rogers, S. Non-coding RNA: Stressed to bits. Nat. Rev. Cancer 2015, 15, 320. [Google Scholar] [CrossRef] [PubMed]
- Fatima, F.; Nawaz, M. Vesiculated Long Non-Coding RNAs: Offshore Packages Deciphering Trans-Regulation between Cells, Cancer Progression and Resistance to Therapies. Non-Coding RNA 2017, 3, 10. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.H.; Lu, P.H.; Lin, Y.H.; Tsai, M.M.; Lin, Y.W.; Yeh, C.T.; Lin, K.H. The long non-coding RNA LINC01013 enhances invasion of human anaplastic large-cell lymphoma. Sci. Rep. 2017, 7, 295. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K.J.; Blobe, G.C. Role of transforming growth factor-β superfamily signaling pathways in human disease. Biochim. Biophys. Acta 2008, 1782, 197–228. [Google Scholar] [CrossRef] [PubMed]
- Uchida, D.; Kawamata, H.; Omotehara, F.; Miwa, Y.; Horiuchi, H.; Furihata, T.; Tachibana, M.; Fujimori, T. Overexpression of TSC-22 (transforming growth factor-β-stimulated clone-22) causes marked obesity, splenic abnormality and B cell lymphoma in transgenic mice. Oncotarget 2016, 7, 14,310–14,323. [Google Scholar] [CrossRef] [PubMed]
- Bizet, A.A.; Liu, K.; Tran-Khanh, N.; Saksena, A.; Vorstenbosch, J.; Finnson, K.W.; Buschmann, M.D.; Philip, A. The TGF-β co-receptor, CD109, promotes internalization and degradation of TGF-β receptors. Biochim. Biophys. Acta 2011, 1813, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Xiao, L.; Rao, J.N.; Zou, T.; Liu, L.; Zhang, D.; Turner, D.J.; Gorospe, M.; Wang, J.Y. Inhibition of Smurf2 translation by miR-322/503 modulates TGF-β/Smad2 signaling and intestinal epithelial homeostasis. Mol. Biol. Cell 2014, 25, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Zlotorynski, E. Non-Coding RNA: A class of their own. Nat. Rev. Mol. Cell Biol. 2017, 18, 71. [Google Scholar] [CrossRef] [PubMed]
- Zlotorynski, E. Non-coding RNA: Circular RNAs promote transcription. Nat. Rev. Mol. Cell Biol. 2015, 16, 206. [Google Scholar] [CrossRef] [PubMed]
- Zlotorynski, E. Non-coding RNA: The cancer link(RNA) between PIP3 and AKT. Nat. Rev. Mol. Cell Biol. 2017, 18, 212–213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, H.Y.; Liu, X.; Wasik, M.A. STAT5A is epigenetically silenced by the tyrosine kinase NPM1-ALK and acts as a tumor suppressor by reciprocally inhibiting NPM1-ALK expression. Nat. Med. 2007, 13, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Molavi, O.; Samadi, N.; Wu, C.; Lavasanifar, A.; Lai, R. Silibinin suppresses NPM-ALK, potently induces apoptosis and enhances chemosensitivity in ALK-positive anaplastic large cell lymphoma. Leuk Lymphoma 2015, 57, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Heuckmann, J.M.; Balke-Want, H.; Malchers, F.; Peifer, M.; Sos, M.L.; Koker, M.; Meder, L.; Lovly, C.M.; Heukamp, L.C.; Pao, W.; et al. Differential protein stability and ALK inhibitor sensitivity of EML4-ALK fusion variants. Clin. Cancer Res. 2012, 18, 4682–4690. [Google Scholar] [CrossRef] [PubMed]
- Parrilla Castellar, E.R.; Jaffe, E.S.; Said, J.W.; Swerdlow, S.H.; Ketterling, R.P.; Knudson, R.A.; Sidhu, J.S.; Hsi, E.D.; Karikehalli, S.; Jiang, L.; et al. ALK-negative anaplastic large cell lymphoma is a genetically heterogeneous disease with widely disparate clinical outcomes. Blood 2014, 124, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Muys, B.R.; Lorenzi, J.C.; Zanette, D.L.; Lima e Bueno Rde, B.; de Araujo, L.F.; Dinarte-Santos, A.R.; Alves, C.P.; Ramao, A.; de Molfetta, G.A.; Vidal, D.O.; et al. Placenta-Enriched LincRNAs MIR503HG and LINC00629 Decrease Migration and Invasion Potential of JEG-3 Cell Line. PLoS ONE 2016, 11, e0151560. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, J.; Breckwoldt, K.; Remmele, C.W.; Hartmann, D.; Dittrich, M.; Pfanne, A.; Just, A.; Xiao, K.; Kunz, M.; Muller, T.; et al. Development of Long Noncoding RNA-Based Strategies to Modulate Tissue Vascularization. J. Am. Coll. Cardiol. 2015, 66, 2005–2015. [Google Scholar] [CrossRef] [PubMed]
- Koperski, L.; Kotlarek, M.; Swierniak, M.; Kolanowska, M.; Kubiak, A.; Gornicka, B.; Jazdzewski, K.; Wojcicka, A. Next-generation sequencing reveals microRNA markers of adrenocortical tumors malignancy. Oncotarget 2017, 8, 49,191–49,200. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Toiyama, Y.; Kitajima, T.; Imaoka, H.; Hiro, J.; Saigusa, S.; Tanaka, K.; Inoue, Y.; Mohri, Y.; Toden, S.; et al. miRNA-503 Promotes Tumor Progression and Is Associated with Early Recurrence and Poor Prognosis in Human Colorectal Cancer. Oncology 2016, 90, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Ide, S.; Toiyama, Y.; Shimura, T.; Kawamura, M.; Yasuda, H.; Saigusa, S.; Ohi, M.; Tanaka, K.; Mohri, Y.; Kusunoki, M. MicroRNA-503 promotes tumor progression and acts as a novel biomarker for prognosis in oesophageal cancer. Anticancer Res. 2015, 35, 1447–1551. [Google Scholar] [PubMed]
- Kavsak, P.; Rasmussen, R.K.; Causing, C.G.; Bonni, S.; Zhu, H.; Thomsen, G.H.; Wrana, J.L. Smad7 binds to Smurf2 to form an E3 ubiquitin ligase that targets the TGF β receptor for degradation. Mol. Cell 2000, 6, 1365–1375. [Google Scholar] [CrossRef]
- Inoue, Y.; Imamura, T. Regulation of TGF-β family signaling by E3 ubiquitin ligases. Cancer Sci. 2008, 99, 2107–2112. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, W.; Ying, Z.; Tian, H.; Zhu, X.; Li, J.; Li, M. Metastatic heterogeneity of breast cancer cells is associated with expression of a heterogeneous TGFβ-activating miR424-503 gene cluster. Cancer Res. 2014, 74, 6107–6118. [Google Scholar] [CrossRef] [PubMed]
- Necsulea, A.; Soumillon, M.; Warnefors, M.; Liechti, A.; Daish, T.; Zeller, U.; Baker, J.C.; Grutzner, F.; Kaessmann, H. The evolution of lncRNA repertoires and expression patterns in tetrapods. Nature 2014, 505, 635–640. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, P.-S.; Chung, I.-H.; Lin, Y.-H.; Lin, T.-K.; Chen, W.-J.; Lin, K.-H. The Long Non-Coding RNA MIR503HG Enhances Proliferation of Human ALK-Negative Anaplastic Large-Cell Lymphoma. Int. J. Mol. Sci. 2018, 19, 1463. https://doi.org/10.3390/ijms19051463
Huang P-S, Chung I-H, Lin Y-H, Lin T-K, Chen W-J, Lin K-H. The Long Non-Coding RNA MIR503HG Enhances Proliferation of Human ALK-Negative Anaplastic Large-Cell Lymphoma. International Journal of Molecular Sciences. 2018; 19(5):1463. https://doi.org/10.3390/ijms19051463
Chicago/Turabian StyleHuang, Po-Shuan, I-Hsiao Chung, Yang-Hsiang Lin, Tzu-Kang Lin, Wei-Jan Chen, and Kwang-Huei Lin. 2018. "The Long Non-Coding RNA MIR503HG Enhances Proliferation of Human ALK-Negative Anaplastic Large-Cell Lymphoma" International Journal of Molecular Sciences 19, no. 5: 1463. https://doi.org/10.3390/ijms19051463