Zebrafish as a Model System for Investigating the Compensatory Regulation of Ionic Balance during Metabolic Acidosis


Department of Biology, York University, Toronto, ON M3J 1P3, Canada
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(4), 1087; https://doi.org/10.3390/ijms19041087
Submission received: 12 March 2018
/
Revised: 25 March 2018
/
Accepted: 2 April 2018
/
Published: 5 April 2018
(This article belongs to the Special Issue Ion Transporters and Channels in Physiology and Pathophysiology)
Abstract
:Zebrafish (Danio rerio) have become an important model for integrative physiological research. Zebrafish inhabit a hypo-osmotic environment; to maintain ionic and acid-base homeostasis, they must actively take up ions and secrete acid to the water. The gills in the adult and the skin at larval stage are the primary sites of ionic regulation in zebrafish. The uptake of ions in zebrafish is mediated by specific ion transporting cells termed ionocytes. Similarly, in mammals, ion reabsorption and acid excretion occur in specific cell types in the terminal region of the renal tubules (distal convoluted tubule and collecting duct). Previous studies have suggested that functional regulation of several ion transporters/channels in the zebrafish ionocytes resembles that in the mammalian renal cells. Additionally, several mechanisms involved in regulating the epithelial ion transport during metabolic acidosis are found to be similar between zebrafish and mammals. In this article, we systemically review the similarities and differences in ionic regulation between zebrafish and mammals during metabolic acidosis. We summarize the available information on the regulation of epithelial ion transporters during acidosis, with a focus on epithelial Na+, Cl− and Ca2+ transporters in zebrafish ionocytes and mammalian renal cells. We also discuss the neuroendocrine responses to acid exposure, and their potential role in ionic compensation. Finally, we identify several knowledge gaps that would benefit from further study.
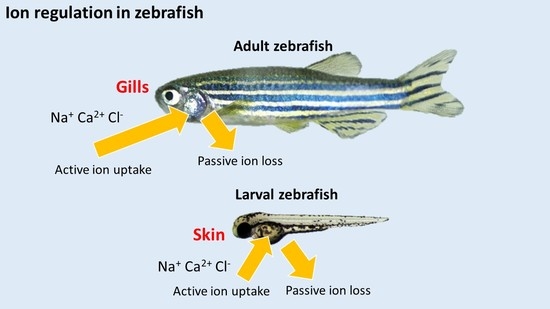
1. Introduction
Maintaining ionic and acid-base homeostasis is essential for all organisms. In recent years, the zebrafish (Danio rerio) has become an important model for understanding the mechanisms underlying the regulation of ionic and acid-base balance. Most fish species can only live in a narrow range of pH near neutrality, primarily owing to the disruption of ionic balance [1]. Zebrafish are highly tolerant to acid exposure (as low as pH 4.0), thus they can be useful for investigating the mechanisms underlying acid tolerance. Additionally, the zebrafish model has several advantages over other vertebrate models, including external and rapid development, short generation time, and applicability of many cellular and molecular physiological techniques in vivo (e.g., non-invasive measurements of ion and acid/base movement, whole-animal fluorescence imaging, transgenics, gene knockdown and knockout).
Zebrafish inhabit a hypo-osmotic freshwater (FW) environment. To maintain ionic and acid-base homeostasis, they have to actively absorb ions and secrete acid to the environment. In adult zebrafish, the gills are the predominant site for regulating ion uptake through specific ion-transporting cells termed ionocytes. During larval stages before the gills are fully developed, regulation of ion transport is mediated by ionocytes found in the skin of the yolk sac. The transepithelial transport of major ions, such as Na+, Ca2+ and Cl−, occurs through three subtypes of ionocytes: H+-ATPase-rich cells (HRCs) for Na+ uptake, and for H+ and ammonia excretion; Na+-K+-ATPase-rich cells (NaRCs) for Ca2+ uptake; Na+-Cl−-cotransporter expressing cells (NCCCs) for both Na+ and Cl− uptake. In the mammalian kidney, ion reabsorption and acid-base regulation are also mediated by specific cell types (Table 1). In mammals, most transepithelial H+ secretion occurs through the apical Na+/H+ exchanger 3 (NHE3) in the proximal tubule. The collecting duct also contributes significantly to H+ secretion through V type H+-ATPase expressed in Type A intercalated cells [2,3,4]. The expression (Table 1) and functional regulation (Table 2) of ion transporters/channels in the zebrafish ionocytes appear to be markedly similar to that in the mammalian renal cells. Therefore, the zebrafish gill or larval skin may serve as a useful in vivo model for investigating the mechanisms regulating ionic and acid-base balance during acid stress in vertebrates.
The processes in the uptake and acid-base regulation in zebrafish have been comprehensively discussed in several previous reviews [1,5,6,7,8,9,10,11]. However, no article has systematically reviewed the similarities and differences in the regulation of epithelial ion transporters between zebrafish and mammals during metabolic acidosis. In this review, we summarize the available information on the regulation of ion transporters in zebrafish ionocytes and mammalian renal cells during acid stress. We discuss the neuroendocrine responses to acid exposure, and their potential role in ionic compensation. Finally, we identify areas that would benefit from future investigations with the zebrafish model.
2. Physiological Responses and Compensatory Regulation of Ion Transport during Acidosis
2.1. Overview of the Effects of Acid Exposure on Freshwater Fish
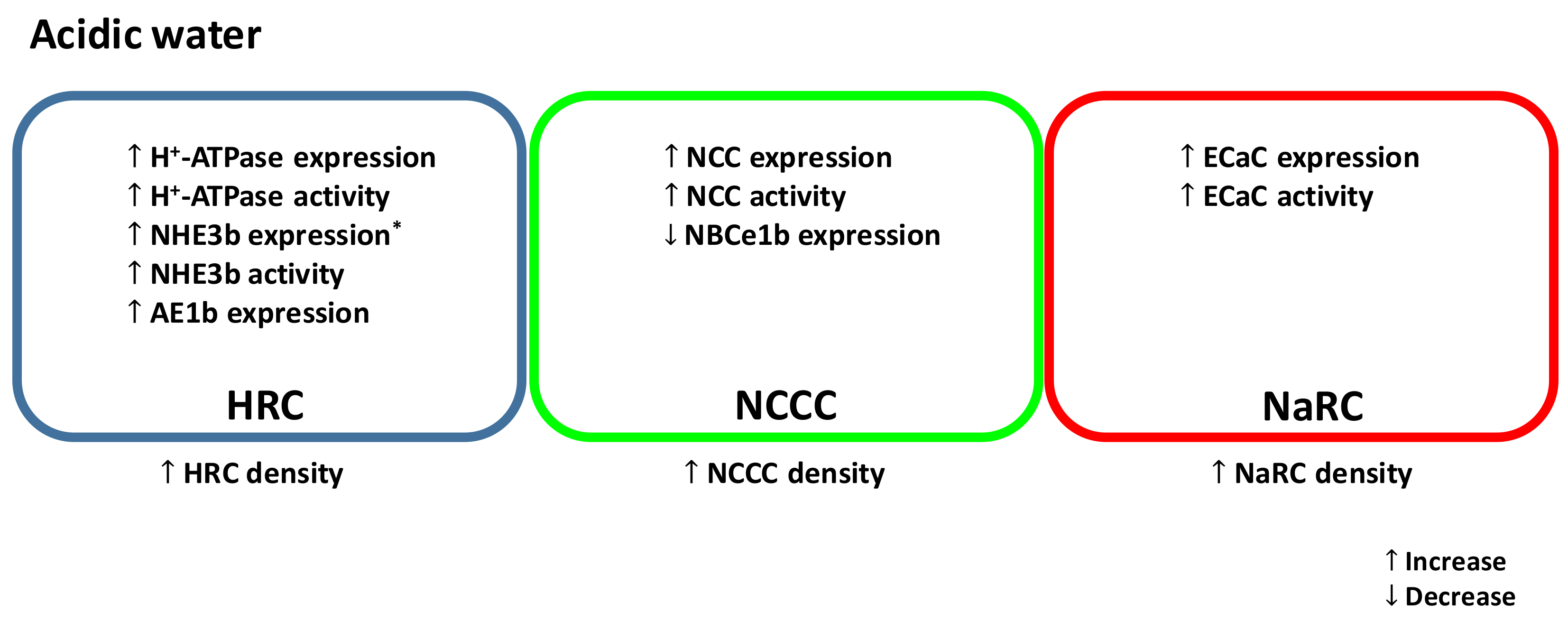
In most FW fish species studied, the primary effects of acid exposure are inhibition of active Na+ uptake coupled with stimulation of passive Na+ losses (for a review, see [1]). This disruption in Na+ regulation by acid exposure may result in a decrease in plasma Na+ level, ultimately leading to cardiovascular failure and mortality [51]. The reduced Na+ uptake by acid exposure is thought to occur through competitive inhibition with an elevated water H+ level [52]. Additionally, Na+ uptake in FW fish is predominately linked to H+ secretion through the actions of the Na+/H+ exchanger (NHE) and H+-ATPase at the apical membrane [53,54,55,56]. Therefore, the elevated level of H+ in the environment may reduce the gradient to drive Na+ influx. On the other hand, the increased Na+ efflux during acid exposure is believed to be associated with the disruption in the integrity of epithelial tight junction, thereby increasing the passive loss of Na+ through paracellular route [1]. Zebrafish, however, are able to increase active Na+ uptake and reduce paracellular Na+ loss following acid exposure. Several recent studies have demonstrated that zebrafish can also increase their capacity for Ca2+ and Cl− uptake after acid exposure (discussed below). These compensatory responses are associated with an activation of a complex network of regulatory mechanisms, including modulation of the expression of ion transporters, stimulation of hormone production and thereby activation of its downstream signaling pathways, changes in the abundance of epithelial tight junction proteins, remodeling of ionocyte differentiation and proliferation, and post-translational regulation of ion transporters. A simplified model of the effects of acid exposure on zebrafish ionocytes is illustrated in Figure 1.
2.2. Functional Regulation of Ion Transporters and Their Involvement in Ionic Compensation during Acidosis
2.2.1. Na+/H+ Exchanger (NHE)
In mammals, NHE3 (SLC9A3) is the major transporter regulating Na+ reabsorption in the proximal tubules of the kidney [57]. In the zebrafish gills/skin, NHE3 (i.e., NHE3b), which is specifically expressed in the apical membrane of HRCs, is also thought to be the predominant transporter for regulating Na+ uptake [17,42,58,59]. In the mammalian kidney, metabolic acidosis was reported to increase the abundance of NHE3 and its activity in the transport of Na+ [43]. In larval zebrafish, mRNA expression of nhe3b was found to increase following acid exposure [38]. However, its expression was decreased in the gills of adult zebrafish exposed to acidic water [17,48]. In acidic environments, elevated level of H+ in the water is expected to significantly impair Na+ uptake via NHE. Interestingly, NHE3b appears to play a role in increasing Na+ uptake after acid exposure. It has been suggested that NHE3b-mediated Na+ uptake in acidic water is overcome through its interaction with the ammonia transporter Rhcg1 [42]. Diffusion of NH3 through Rhcg1 creates a microenvironment where H+ level is lowered by its combination with NH3 to form NH4+. The resultant H+ gradient is believed to drive Na+ uptake via electroneutral NHE3b. However, recent findings have suggested that in acidic environments, the relative role of NHE3b is reduced and NCC may become the predominant pathway for the absorption of Na+ (discussed below).
2.2.2. Na+-Cl− Cotransporter (NCC)
In the distal convoluted tubules of the mammalian kidney, NCC (SLC12A3) is responsible for fine-tuning the reabsorption of filtered Na+ (5%–7%) [60]. Several studies have investigated the effects of metabolic acidosis on the abundance and function of NCC. However, the findings are currently inconclusive. Following 7 days of metabolic acidosis, it was shown that the expression of NCC, as well as its activity in Na+ transport, were decreased in the rat kidney [61,62]. It was also demonstrated that the expression of NCC was increased significantly after 5 days of metabolic acidosis in both rat and mice kidney [47,63]. Clearly, further study is required to resolve this inconsistency regarding NCC expression during metabolic acidosis.
In zebrafish, NCC was found to be involved in the compensatory regulation of Na+ uptake following acid exposure [26,48]. In zebrafish, NCC (SLC12A10.2; an NCC-like member distinct from the mammalian orthologous of SLC12A3) is expressed in a specific subtype of ionocyte, the NCCCs [27]. Under normal condition, NHE3b plays a more critical role in Na+ regulation. However, recent studies have demonstrated that NCC could become important under physiological stressors, such as acidosis [26,48] or during periods of increased paracellular ion losses [64]. After acid exposure, the number of NCCCs in larval zebrafish skin as well as in the adult gills was markedly increased [48]. Whole body Na+ levels in NCC-deficient fish was also found to be lower than that in control fish following acid exposure [48]. Additionally, it was demonstrated that NCC was involved in the rapid regulation of both Na+ and Cl− uptake after acute acidosis [26]. These responses appeared to compensate for the reduced Na+ and Cl− uptake during acute acid exposure. Interestingly, this rapid compensation of Na+ and Cl- uptake was found to be solely mediated by NCCCs, but not HRCs [26]. How NCC is acutely activated by acid exposure has remained unclear, but it has been proposed to occur via post-translational modification (e.g., phosphorylation) and/or by membrane trafficking [26]. A recent mammalian study has shown that post-translational activation of NCC is associated with the WNK4-SPAK signalling pathway [63]. Whether a similar mechanism also occurs in zebrafish NCC awaits further investigation.
2.2.3. Anion Exchanger (AE)
In mammals experiencing metabolic acidosis, bicarbonate excretion from the kidney is reduced to preserve base equivalents in the body. Such reduction is facilitated by two anion exchangers: pendrin and AE1. In the kidney, pendrin (SLC26A4; Cl−/HCO3− exchanger) is expressed in the apical membrane of Type B intercalated cells. During metabolic acidosis, the expression of pendrin is significantly reduced [65,66]. Acid loading also appears to induce endocytosis of pendrin into the cytosol [46]. These responses subsequently decrease bicarbonate excretion into the lumen. SLC26A4 is also found to be expressed in zebrafish, and the knockdown of SLC26A4 significantly decreases Cl- uptake after acclimation to low Cl− water [67]. Reduced base excretion (i.e., titratable base) is also observed in SLC26A4-deficient zebrafish [67]. These findings indicate the physiological significance of SLC26A4 in Cl− uptake and HCO3− secretion in zebrafish. However, how acidosis affects the expression and function of SLC26A4 in zebrafish are yet to be explored.
AE1 (SLC4A), which is expressed in the basolateral membrane of Type A intercalated cells, exhibits opposite response during acidosis. Expression of AE1 is increased during metabolic acidosis, thereby enhancing bicarbonate absorption into the interstitium [22,68]. In zebrafish, AE1 (i.e., AE1b) is expressed in the basolateral membrane of HRCs [21]. It has been shown that acid exposure increases mRNA expression of ae1b in larval zebrafish [21]. Additionally, larval zebrafish experiencing AE1b knockdown have been found to exhibit a reduction in whole body Na+ level and H+ secretion [21]. These findings suggest that AE1b is involved in regulating Na+ balance and acid secretion in zebrafish.
2.2.4. Sodium-Bicarbonate Cotransporter (NBCe)
In mammals, the NBCe1 (“e” stands for electrogenic) plays a crucial role in maintaining normal blood pH by regulating bicarbonate absorption in the proximal tubules. The mammalian NBCe1 has multiple transcript variants, including NBCe1-A (kidney-specific), NBCe1-B (intestine/pancreas-specific) and NBCe1-C (brain-specific). Only NBCe1-A, which is expressed in the basolateral membrane of the proximal tubular cells, is thought to be involved in regulating bicarbonate absorption in the kidney [24,25]. It has been proposed that movement of HCO3− via NBCe1-A is coupled to apical NHE3 and the activity of carbonic anhydrase (CA) [69]. Specifically, luminal bicarbonate is initially protonated by NHE3 and is converted to CO2 via a dehydration reaction that is accelerated by CA4. Dissolved CO2 permeates into the cell and undergoes a reverse hydration reaction catalyzed by CA2 which converts CO32− to HCO3−. Subsequently, the basolateral Na+/K+-ATPase generates the driving force for NBCe1-A mediated electrogenic Na+ and HCO3- efflux. In rat, inducing metabolic acidosis by NH4Cl loading did not affect mRNA expression of NBCe1 in the kidney [70]. However, in isolated basolateral membrane vesicles from rabbit kidney, HCO3−-dependent Na+ uptake was increased during incubation in acidic media [71]. These results suggest that activation of the NBCe1-A by acid exposure is likely a post-transcriptional event. On the other hand, it has been documented that loss-of-function mutation in NBCe1-A results in proximal renal tubular acidosis [72,73,74]. NBCe1 null mice (NBCe1−/−) have also been found to exhibit severe metabolic acidosis and death before weaning [75].
In zebrafish, the NBCe1 protein sequence is more similar to the mammalian NBCe1-B (78%) than NBCe1-A (73%) [21,23]. Zebrafish NBCe1-B is expressed in the pronephros, eyes, brain, and ionocytes (i.e., NCCCs) in larvae, as well as the gills in adults [21,23]. Using a Xenopus oocyte expression system, it was demonstrated that zebrafish NBCe1-B exhibited electrogenic Na+ and HCO3− cotransport activity [23]. As discussed previously, the transport function of NBCe1 is believed to be coupled with the activity of NHE3 and CA2. In zebrafish, these proteins (i.e., NHE3b, CA2 like-a) are expressed specifically in the HRCs [18,59,76]. Interestingly, NBCe1-B was shown to be expressed in NCCCs [21]. In the gills of zebrafish, mRNA level of NBCe1-B was also found to decrease following acid exposure [21]. These results suggest that NBCe1-B is unlikely involved in bicarbonate absorption in zebrafish. The functional role of NBCe1-B in the basolateral membrane of NCCCs remains to be explored.
2.2.5. Epithelial Ca2+ Channel (ECaC)
In mammals, transepithelial transport of Ca2+ is mediated by the transient receptor potential channels (TRPV): TRPV5 (also called ECaC) and TRPV6. TRPV5 contributes to Ca2+ transport in the kidney, while TRPV6 is more ubiquitously expressed [77]. In the mammalian kidney, the majority of Ca2+ reabsorption occurs in the proximal tubules (~60–70%) and the thick ascending limb of Henle (~10%) through the paracellular route [78]. This passive movement of Ca2+ is driven by local Na+ reabsorption, and thus Ca2+ reabsorption in these regions is not directly regulated. Although only 5–10% of total Ca2+ reabsorption occurs in the distal convoluted tubule and the collecting duct, these are the major sites for active regulation of Ca2+ transport by TRPV5 [78,79].
The uptake of Ca2+ by ECaC in the mammalian kidney is highly influenced by luminal pH. For example, it was shown that a decrease in extracellular pH significantly inhibited Ca2+ uptake in primary cultures from rabbit cortical collecting duct [80]. Additionally, reduced pH was also demonstrated to inhibit Ca2+ uptake in a human embryonic kidney 293 cell line (HEK 293) expressing ECaC [81].
Unlike mammals, zebrafish only have one TRPV5/6 orthologue (ECaC) [31]. In zebrafish, ECaC is expressed in a subset of NaRCs, which plays an essential role in Ca2+ absorption [31,82]. Previous studies have shown that larval zebrafish exposed to acidic water exhibit a significant reduction in whole body Ca2+ level [39,49]. Such reduction is likely due to a concurrent decrease in Ca2+ uptake and an increase in Ca2+ efflux [49]. Unlike Na+ uptake, which is stimulated in zebrafish following acid acclimation, Ca2+ uptake remains significantly inhibited in acidic water [49]. Interestingly, when the acid-exposed fish are returned to control pH water, the capacity for Ca2+ uptake is substantially increased. The increased Ca2+ uptake appears to be associated with an increased mRNA expression of ecac and density of ecac-expressing NaRCs [49]. This finding appears to be different from a previous study with mice where acid-loading (NH4Cl) resulted in a reduction in ecac expression in the kidney [50]. The mechanisms underlying the increased ecac-expressing NaRCs by acid exposure in zebrafish remain unknown. Nevertheless, acid exposure has been shown to increase whole body cortisol level in zebrafish [49,83], and that cortisol is involved in promoting the proliferation of NaRCs [84,85]. Therefore, it is possible that the increased ecac-expressing NaRCs is due to the increased cortisol level during acid exposure. On the other hand, it has been reported that exposure to acidic water increases whole body 3′,5′-cyclic adenosine monophosphate (cAMP) levels in zebrafish [86], and that treatment with cAMP-elevating agents (i.e., 8-Br-cAMP, forskolin) significantly stimulates Ca2+ influx [87]. Future experiments should address whether cAMP plays a role in stimulating Ca2+ uptake following acid exposure. It is also noteworthy that zebrafish exposed to acidic water for 4 days exhibited over a 5-fold reduction in whole body Ca2+ level [39,49]. The increased Ca2+ uptake by acid exposure was only observed when the fish were returned back to normal pH water [49]. How zebrafish compensate for the loss of Ca2+ (e.g., regulation of paracellular permeability to Ca2+) during long-term acid exposure requires further investigation.
2.2.6. Effects of Acidosis on the Regulation of Other Epithelial Ion Transporters
In mammals, several other epithelial ion transporters are known to be involved in ionic compensation during metabolic acidosis, including Na+/phosphate cotransporter (NaPi; Pi stands for inorganic phosphate), Cl−/HCO3− exchanger, and H+-K+-ATPase (HKA). However, whether these transporters are involved in ion regulation in zebrafish exposed to acidic water are virtually unknown. In mammals, NaPi has three different protein families, NaPi-I, NaPi-II and NaPi-III, with NaPi-II thought to be the major player in Pi regulation in the kidney [88]. In the mammalian kidney, reabsorption of Pi occurs predominantly in the proximal tubule [89]. In rats experiencing 10 days of metabolic acidosis, renal mRNA and protein expression of NaPi (NaPi-II) were significantly decreased [90]. The transport activity (e.g., Jmax) of NaPi was also found to decrease [90]. In another study with the mouse kidney, however, it was demonstrated that NaPi (NaPi-IIa and NaPi-IIc) protein abundance was increased after 2 days or 10 days of acidosis [91]. The activity of NaPi was found to increase after 2 days of acidosis, but then returned to normal level after 7 days [91]. These findings suggest possible species-specific differences in the regulation of NaPi during acid exposure. In zebrafish, two paralogues of NaPi-IIb, NaPi-IIb1 and NaPi-IIb2, were found to express in the apical membrane of renal tubular cells [92]. NaPi-IIb1 and NaPi-IIb2 appeared to exhibit a significant difference in their functional properties. For example, the transport activity of zebrafish NaPi-IIb2 was shown to have a much higher affinity to Pi than NaPi-IIb1 when expressed in Xenopus oocytes [92]. NaPi-IIb2 was also found to be insensitive to extracellular pH as opposed to NaPi-IIb1 [92]. To date, no study has yet identified the expression of NaPi in zebrafish gill/ionocytes. [93,94]
In mammals, excretion of inorganic sulfate (Si) is primarily regulated by the Na-Si cotransporter in the proximal tubule [95]. During metabolic acidosis, reabsorption of Si is decreased [96], which appears to be associated with decreased NaSi-1 mRNA and protein abundance, as well as reduced Na+-Si cotransport activity [96]. In zebrafish, NaSi1 mRNA is expressed in the kidney [97,98]. Functional characterization of the zebrafish NaSi1 in Xenopus oocytes suggested that the zebrafish transporter closely resembles the mammalian counterpart in terms of both affinity and transport capacity [97]. However, NaSi1 was not found to be expressed in the zebrafish gill [97].
In the mammalian kidney, K+ reabsorption and acid excretion can be mediated by HKA, which is predominantly expressed in the apical membrane of intercalated cells (both type-A and type-B) and principal cells [99]. Mice experiencing metabolic acidosis exhibited an increased mRNA expression of HKA (α-subunits) [100]. The activity of HKA was also found to increase during metabolic acidosis [101,102]. The protein sequence of the mammalian H+-K+-ATPase α-subunit is most similar to the zebrafish Na+/K+-ATPase α-subunit isoforms 1, 2, 3, 5 and 8 (~63%–64%). Currently, however, no physiological evidence has demonstrated that these zebrafish Na+/K+-ATPases can function as HKA.
3. Neuroendocrine Responses Following Acid Exposure
In zebrafish, several neuroendocrine factors are found to be involved in ionic and acid-base regulation during acid exposure, including cortisol [38,83], endothelin 1 [103], angiotensin II [104], stanniocalcin [49,105], catecholamines [106] and oestrogen-related receptor [107]. In other FW fish species, mRNA expression and/or plasma levels of thyroid hormone [108], prolactin [109], growth hormone [110] and somatolactin [110,111] are found to be modulated by acid exposure. Whether these hormones are affected by acid exposure in zebrafish remain unexplored.
3.1. Cortisol
In mammals, the corticosteroid is a class of steroid hormones that include cortisol (glucocorticoid) and aldosterone (mineralocorticoid). Both of these hormones are synthesized in the adrenal cortex of mammals, with the aldosterone thought to be the primary factor in regulating ion reabsorption [112]. In mammals, activation of the mineralocorticoid receptor (MR) stimulates Na+ reabsorption and H+ excretion in the kidney [113]. Teleosts lack aldosterone synthase to produce aldosterone [114]. However, several in vitro studies have demonstrated that cortisol is able to activate both glucocorticoid receptor (GR) and MR [114,115,116].
In zebrafish larvae, GR was found to be expressed in all three ionocytes discussed above, including NaRCs [85], HRCs [83,117] and NCCCs [118]. Treatment with cortisol was shown to increase mRNA expression of genes that are involved in ion uptake or H+ secretion, including ecac, ncc, nhe3b, atp6v1a (encodes for H+-ATPase) and ae1b [38,117]. On the other hand, mRNA level of 11β-hydroxylase (cyp11b; encodes for a cortisol-synthesis enzyme) was found to increase following acid exposure [38]. Consistently, whole body level of cortisol was markedly increased in larval zebrafish exposed to acidic water [49,83]. The increased whole body cortisol level by acid exposure appears to be important in increasing acid secretion and active Na+ uptake in the HRCs [38,83,119]. During acid exposure, cortisol was also shown to be involved in reducing diffusive Na+ losses through paracellular pathways, likely by increasing the abundance of epithelial tight junctions [119]. Interestingly, the cortisol-induced effects on Na+ regulation and H+ secretion in zebrafish appear to be mediated by GR only.
3.2. Endothelin
In mammals, endothelin is involved in increasing acid excretion in the kidney during metabolic acid stress [120]. During acid loading, expression of endothelin is increased in the kidney [121], which subsequently stimulates acid secretion by enhancing the activity of NHE3 [122,123] and H+-ATPase [124]. A recent study with zebrafish has also suggested that endothelin plays a critical role in regulating acid secretion [103]. The mRNA expression levels of endothelin-1 (edn1) and its receptors (ednraa) were found to increase in zebrafish exposed to acidic water. EDN1 was also shown to promote acid secretion, primarily by post-translational activation of the H+-ATPase in the HRCs of zebrafish [103]. These findings clearly demonstrate that endothelin is involved in regulating acid excretion in zebrafish.
3.3. Oestrogen-Related Receptor
The oestrogen-related receptor (ERR) belongs to orphan nuclear receptors which control the transcription of several genes essential in energy metabolism in mammals [125]. In zebrafish, it was recently demonstrated that ERRa is involved in promoting acid secretion in zebrafish, partially by increasing the density of HRCs [107]. erra mRNA was found to be expressed in HRCs, and that acid exposure increased its abundance [107]. ERRa was also demonstrated to positively regulate the mRNA expressions of nhe3b, atp6v1a (encodes for H+-ATPase) and ae1b [107]. This is a novel finding on the role of ERRa in acid-base regulation in vertebrates.
3.4. Catecholamines
In mammals, it is well documented that catecholamines are involved in regulating Na+ reabsorption in the kidney [126]. For example, treatment with catecholamines was found to stimulate Na+/K+-ATPase activity in the basolateral membrane of proximal tubules isolated from rats [127,128]. The activity and abundance of NHE3 in the proximal tubule of rats were also shown to increase following stimulation of the α-adrenergic receptor [129,130,131]. It was also demonstrated that activation of the β adrenergic receptor increased NHE3 activity by promoting the association of the NHE regulatory factor (NHERF) with NHE3 [132]. At present, the involvement of the adrenergic system in the kidney during metabolic acidosis is not fully understood.
In larval zebrafish, both HRCs and NaRCs are innervated, and β receptors appear to be present in both cell types [7,106,133]. Whether NCCCs are innervated is currently unknown. It has been demonstrated that knockdown of specific β adrenergic receptor reduces the magnitude of increased Na+ uptake in larval zebrafish exposed to acidic water [106]. The underlying mechanism of how β adrenergic receptor elicits its effect on Na+ transporters remains unclear. The downstream signalling cascade of β adrenergic receptor is mediated by cAMP, and it has been shown that treatment with cAMP-elevating agents increases Na+ uptake in larval zebrafish [86]. Whole body cAMP level has also been found to increase in larval zebrafish acutely exposed to acidic water [86]. These findings suggest that activation of the β adrenergic system during acid exposure increases whole body cAMP level, which subsequently increases Na+ uptake. Using a morpholino knockdown approach, it has also been demonstrated that cAMP-stimulated Na+ uptake is likely mediated by both NHE3b and NCC [86].
3.5. Angiotensin II
In mammals, the renin-angiotensin system (RAS) plays a critical role in regulating ion and fluid reabsorption in the kidney [134]. Angiotensinogen, which is primarily released from the liver, is first cleaved to angiotensin-I (ANG-I) by the enzyme renin secreted in the kidney. ANG-I is then converted to the biologically active form ANG-II by angiotensin-converting enzyme (ACE). In the mammalian kidney, ANG-II is found to stimulate NHE3, H+-ATPase and NCC [135,136,137,138,139,140]. A previous study with larval zebrafish has demonstrated that whole body ANG-II level is elevated following acid exposure [104]. It has also been shown that translational gene knockdown of renin significantly attenuated the increased Na+ uptake by acute acid exposure [104]. These findings suggest that the RAS is involved in promoting Na+ uptake under acidic environment. On the other hand, treatment with exogenous ANG-II was found to increase the mRNA expression of ncc, but not nhe3b or atp6v1a (encodes for H+-ATPase) [104]. Interestingly, NCC appears to play a critical role in the rapid compensatory regulation of Na+ uptake following acute acid exposure [26]. In mammals, ANG-II is proposed to activate NCC through post-translational regulation [141]. Whether similar mechanism occurs in zebrafish NCC has yet to be explored. The expression of ANG-II receptors in ionocytes of zebrafish also awaits further study.
3.6. Stanniocalcin
In mammals, the principal cells in the collecting duct are one of the primary sources of stanniocalcin, which acts as a hypocalcemic factor to decrease Ca2+ uptake in the intestine and kidney [142,143]. In zebrafish, stanniocalcin (i.e., STC1) is known to negatively regulate ecac expression and thereby reducing Ca2+ uptake [144]. Recently, this hormone has also been proposed to regulate acid excretion and Ca2+ transport following acid exposure [49,105]. Exposure to acidic water appears to specifically reduce mRNA expression of stc1, but not other Ca2+-regulatory hormones (e.g., parathyroid hormone, calcitonin) [49,105]. The reduced expression of stc1 may potentially promote Ca2+ uptake by increasing ecac expression and density of ecac-expressing NaRCs during acid exposure [49,105]. Additionally, acid excretion is significantly reduced in zebrafish overexpressing STC1, possibly owing to its inhibitory effects on HRCs differentiation from epidermal stem cells [105]. These novel findings from the zebrafish model provide new understanding of the role of STC1 during metabolic acidosis.
4. Conclusions and Perspectives
The zebrafish has emerged as a useful in vivo model to investigate the cellular and molecular mechanisms underlying the regulation of ionic and acid-base homeostasis in vertebrates. Previous research efforts with the gill/skin of zebrafish have greatly improved our fundamental understanding of the compensatory mechanisms, ion transport function, and neuroendocrine responses during metabolic acidosis. However, it is also important to recognize the differences in the expression pattern and functional regulation of some of the ion transporters between zebrafish and higher vertebrates. For examples, (i) H+-ATPase is expressed in both the apical and basolateral membranes of mammalian intercalated cells, whereas it is expressed only in the apical membrane of zebrafish HRCs; (ii) NaPi and NaSi are absent in the zebrafish gill/ionocytes; (iii) compared to zebrafish, mammals do not appear to compensate for the increased Ca2+ loss during acid loading. These differences may ultimately contribute to different strategies employed by zebrafish and mammals in regulating systemic acid-base and ion homeostasis during metabolic acidosis.
Currently, studies on the ionic and acid-base regulation in zebrafish are mostly limited to early developmental stages. Ion regulation occurs in the skin of the yolk sac in larvae, as opposed to the gills, intestine, and kidney in adults. Therefore, future studies should also address the regulation of ionic and acid-base balance in adult zebrafish. Additionally, a number of areas still remain largely unknown, and should be addressed in future investigations: (i) identification of the localization of several hormone receptors (e.g., receptors for STC1 and ANG-II) in zebrafish ionocytes; (ii) mechanism of environmental and intracellular acid sensing; (iii) responses of other neuroendocrine factors that are thought to be important in ionic and acid-base regulation in FW fish (e.g., thyroid hormone, prolactin, growth hormone, and somatolactin); (iv) post-translational regulation of ion transporters/channels; (v) potential crosstalk among neuroendocrine factors; and (vi) role of transcription factors and hormones in ionocyte differentiation/proliferation during acidosis. Furthermore, previous research on the effects of acidosis in zebrafish is limited to Na+ and Ca2+, and to a lesser extent Cl−. Future studies should also address the regulation of other major ions (e.g., Mg2+ and K+) during acid exposure.
Acknowledgments
Our original research was supported by the Canada Research Chairs Program and a Discovery Grant from the Natural Sciences and Engineering Research Council of Canada (NSERC) to Raymond W. M. Kwong. Processing charge for this article was supported by the York University Libraries Open Access Author Fund.
Author Contributions
Lletta Lewis and Raymond W. M. Kwong drafted the manuscript; Lletta Lewis and Raymond W. M. Kwong edited and revised the manuscript; Lletta Lewis and Raymond W. M. Kwong approved the final version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kwong, R.W.M.; Kumai, Y.; Perry, S.F. The physiology of fish at low pH: The zebrafish as a model system. J. Exp. Biol. 2014, 217, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.A.; Finberg, K.E.; Breton, S.; Marshansky, V.; Brown, D.; Geibel, J.P. Renal Vacuolar H+-ATPase. Physiol. Rev. 2004, 84, 1263–1314. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.A.; Devuyst, O.; Bourgeois, S.; Mohebbi, N. Regulated acid-base transport in the collecting duct. Pflügers Arch. 2009, 458, 137–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobulescu, I.A.; Moe, O.W. Na+/H+ exchangers in renal regulation of acid-base balance. Semin. Nephrol. 2006, 26, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Guh, Y.-J.; Lin, C.-H.; Hwang, P.-P. Osmoregulation in zebrafish: Ion transport mechanisms and functional regulation. EXCLI J. 2015, 14, 627–659. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Hwang, P.-P. The Control of Calcium Metabolism in Zebrafish (Danio rerio). Int. J. Mol. Sci. 2016, 17, 1783. [Google Scholar] [CrossRef] [PubMed]
- Kwong, R.W.M.; Kumai, Y.; Perry, S.F. Neuroendocrine control of ionic balance in zebrafish. Gen. Comp. Endocrinol. 2016, 234, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.-P. Ion uptake and acid secretion in zebrafish (Danio rerio). J. Exp. Biol. 2009, 212, 1745–1752. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.; Kumai, Y.; Porteus, C.S.; Tzaneva, V.; Kwong, R.W.M. An emerging role for gasotransmitters in the control of breathing and ionic regulation in fish. J. Comp. Physiol. B 2016, 186, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Guh, Y.-J.; Hwang, P.-P. Insights into molecular and cellular mechanisms of hormonal actions on fish ion regulation derived from the zebrafish model. Gen. Comp. Endocrinol. 2017, 251, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.-P.; Chou, M.-Y. Zebrafish as an animal model to study ion homeostasis. Pflugers Arch. 2013, 465, 1233–1247. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-Y.; Horng, J.-L.; Kunkel, J.G.; Hwang, P.-P. Proton pump-rich cell secretes acid in skin of zebrafish larvae. Am. J. Physiol. Cell Physiol. 2005, 290, C371–C378. [Google Scholar] [CrossRef] [PubMed]
- Horng, J.-L.; Lin, L.-Y.; Huang, C.-J.; Katoh, F.; Kaneko, T.; Hwang, P.-P. Knockdown of V-ATPase subunit A (atp6v1a) impairs acid secretion and ion balance in zebrafish (Danio rerio). Am. J. Physiol. Integr. Comp. Physiol. 2007, 292, R2068–R2076. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Hirsch, S.; Gluck, S. Localization of a proton-pumping ATPase in rat kidney. J. Clin. Investig. 1988, 82, 2114–2126. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Hirscht, S.; Gluck, S. An H+-ATPase in opposite plasma membrane domains in kidney epithelial cell subpopulations. Nature 1988, 331, 622–624. [Google Scholar] [CrossRef] [PubMed]
- Alper, S.L.; Natale, J.; Gluck, S.; Lodish, H.F.; Brown, D. Subtypes of intercalated cells in rat kidney collecting duct defined by antibodies against erythroid band 3 and renal vacuolar H+-ATPase. Proc. Natl. Acad. Sci. USA 1989, 86, 5429–5433. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.-J.; Chou, M.-Y.; Kaneko, T.; Hwang, P.-P. Gene expression of Na+/H+ exchanger in zebrafish H+-ATPase-rich cells during acclimation to low-Na+ and acidic environments. Am. J. Physiol. Cell Physiol. 2007, 293, C1814–C1823. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Kobayashi, S.; Nakamura, N.; Miyagi, H.; Esaki, M.; Hoshijima, K.; Hirose, S. Close Association of Carbonic Anhydrase (CA2a and CA15a), Na(+)/H(+) Exchanger (Nhe3b), and Ammonia Transporter Rhcg1 in Zebrafish Ionocytes Responsible for Na(+) Uptake. Front. Physiol. 2013, 4, 59. [Google Scholar] [CrossRef] [PubMed]
- Biemesderfer, D.; Pizzonia, J.; Abu-Alfa, A.; Exner, M.; Reilly, R.; Igarashi, P.; Aronson, P.S. NHE3: A Na+/H+ exchanger isoform of renal brush border. Am. J. Physiol. 1993, 265, F736–F742. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, M.; Loffing, J.; Lötscher, M.; Kaissling, B.; Alpern, R.J.; Moe, O.W. Expression of NHE-3 in the apical membrane of rat renal proximal tubule and thick ascending limb. Kidney Int. 1995, 48, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-C.; Yan, J.-J.; Cruz, S.A.; Horng, J.-L.; Hwang, P.-P. Anion exchanger 1b, but not sodium-bicarbonate cotransporter 1b, plays a role in transport functions of zebrafish H+-ATPase-rich cells. Am. J. Physiol. Cell Physiol. 2011, 300, C295–C307. [Google Scholar] [CrossRef] [PubMed]
- Fejes-Tóth, G.; Chen, W.R.; Rusvai, E.; Moser, T.; Náray-Fejes-Tóth, A. Differential expression of AE1 in renal HCO3-secreting and -reabsorbing intercalated cells. J. Biol. Chem. 1994, 269, 26717–26721. [Google Scholar] [PubMed]
- Sussman, C.R.; Zhao, J.; Plata, C.; Lu, J.; Daly, C.; Angle, N.; DiPiero, J.; Drummond, I.A.; Liang, J.O.; Boron, W.F.; et al. Cloning, localization, and functional expression of the electrogenic Na+ bicarbonate cotransporter (NBCe1) from zebrafish. Am. J. Physiol. Cell Physiol. 2009, 297, C865–C875. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, B.M.; Biemesderfer, D.; Romero, M.F.; Boulpaep, E.L.; Boron, W.F. Immunolocalization of the electrogenic Na+-HCO3 cotransporter in mammalian and amphibian kidney. Am. J. Physiol. 1999, 276, F27–F38. [Google Scholar] [PubMed]
- Abuladze, N.; Song, M.; Pushkin, A.; Newman, D.; Lee, I.; Nicholas, S.; Kurtz, I. Structural organization of the human NBC1 gene: KNBC1 is transcribed from an alternative promoter in intron 3. Gene 2000, 251, 109–122. [Google Scholar] [CrossRef]
- Kwong, R.W.M.; Perry, S.F. A role for sodium-chloride cotransporters in the rapid regulation of ion uptake following acute environmental acidosis: New insights from the zebrafish model. Am. J. Physiol. Cell Physiol. 2016, 311, C931–C941. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-F.; Tseng, Y.-C.; Yan, J.-J.; Hiroi, J.; Hwang, P.-P. Role of SLC12A10.2, a Na-Cl cotransporter-like protein, in a Cl uptake mechanism in zebrafish (Danio rerio). Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1650–R1660. [Google Scholar] [CrossRef] [PubMed]
- Gamba, G. The thiazide-sensitive Na+-Cl− cotransporter: Molecular biology, functional properties, and regulation by WNKs. Am. J. Physiol. Ren. Physiol. 2009, 297, F838–F848. [Google Scholar] [CrossRef] [PubMed]
- Ellison, D.H.; Velázquez, H.; Wright, F.S. Thiazide-sensitive sodium chloride cotransport in early distal tubule. Am. J. Physiol. 1987, 253, F546–F554. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, M.D.; Kaplan, M.R.; Verlander, J.W.; Lee, W.S.; Brown, D.; Poch, E.; Gullans, S.R.; Hebert, S.C. Localization of the thiazide sensitive Na-Cl cotransporter, rTSC1 in the rat kidney. Kidney Int. 1996, 50, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.-C.; Liao, B.-K.; Huang, C.-J.; Lin, L.-Y.; Hwang, P.-P. Epithelial Ca2+ channel expression and Ca2+ uptake in developing zebrafish. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1202–R1211. [Google Scholar] [CrossRef] [PubMed]
- Liao, B.-K.; Deng, A.-N.; Chen, S.-C.; Chou, M.-Y.; Hwang, P.-P. Expression and water calcium dependence of calcium transporter isoforms in zebrafish gill mitochondrion-rich cells. BMC Genom. 2007, 8, 354. [Google Scholar] [CrossRef] [PubMed]
- Hoenderop, J.G.; van der Kemp, A.W.; Hartog, A.; van de Graaf, S.F.; van Os, C.H.; Willems, P.H.; Bindels, R.J. Molecular identification of the apical Ca2+ channel in 1,25-dihydroxyvitamin D3-responsive epithelia. J. Biol. Chem. 1999, 274, 8375–8378. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.B.; Chen, X.Z.; Berger, U.V.; Vassilev, P.M.; Brown, E.M.; Hediger, M.A. A rat kidney-specific calcium transporter in the distal nephron. J. Biol. Chem. 2000, 275, 28186–28194. [Google Scholar] [CrossRef] [PubMed]
- Hoenderop, J.G.; Hartog, A.; Stuiver, M.; Doucet, A.; Willems, P.H.; Bindels, R.J. Localization of the epithelial Ca(2+) channel in rabbit kidney and intestine. J. Am. Soc. Nephrol. 2000, 11, 1171–1178. [Google Scholar] [PubMed]
- Baxendale-Cox, L.M.; Duncan, R.L.; Liu, X.; Baldwin, K.; Els, W.J.; Helman, S.I. Steroid hormone-dependent expression of blocker-sensitive ENaCs in apical membranes of A6 epithelia. Am. J. Physiol. 1997, 273, C1650–C1656. [Google Scholar] [CrossRef] [PubMed]
- Garty, H.; Palmer, L.G. Epithelial sodium channels: Function, structure, and regulation. Physiol. Rev. 1997, 77, 359–396. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Shih, T.-H.; Liu, S.-T.; Hsu, H.-H.; Hwang, P.-P. Cortisol Regulates Acid Secretion of H(+)-ATPase-rich Ionocytes in Zebrafish (Danio rerio) Embryos. Front. Physiol. 2015, 6, 328. [Google Scholar] [CrossRef] [PubMed]
- Horng, J.-L.; Lin, L.-Y.; Hwang, P.-P. Functional regulation of H+-ATPase-rich cells in zebrafish embryos acclimated to an acidic environment. Am. J. Physiol. Cell Physiol. 2009, 296, C682–C692. [Google Scholar] [CrossRef] [PubMed]
- Fejes-Tóth, G.; Náray-Fejes-Tóth, A. Effect of acid/base balance on H-ATPase 31 kD subunit mRNA levels in collecting duct cells. Kidney Int. 1995, 48, 1420–1426. [Google Scholar] [CrossRef] [PubMed]
- Verlander, J.W.; Madsen, K.M.; Cannon, J.K.; Tisher, C.C. Activation of acid-secreting intercalated cells in rabbit collecting duct with ammonium chloride loading. Am. J. Physiol. 1994, 266, F633–F645. [Google Scholar] [CrossRef] [PubMed]
- Kumai, Y.; Perry, S.F. Ammonia excretion via Rhcg1 facilitates Na+ uptake in larval zebrafish, Danio rerio, in acidic water. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1517–R1528. [Google Scholar] [CrossRef] [PubMed]
- Laghmani, K.; Borensztein, P.; Ambühl, P.; Froissart, M.; Bichara, M.; Moe, O.W.; Alpern, R.J.; Paillard, M. Chronic metabolic acidosis enhances NHE-3 protein abundance and transport activity in the rat thick ascending limb by increasing NHE-3 mRNA. J. Clin. Investig. 1997, 99, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Ambuhl, P.M.; Amemiya, M.; Danczkay, M.; Lotscher, M.; Kaissling, B.; Moe, O.W.; Preisig, P.A.; Alpern, R.J. Chronic metabolic acidosis increases NHE3 protein abundance in rat kidney. Am. J. Physiol. 1996, 271, F917–F925. [Google Scholar] [CrossRef] [PubMed]
- Preisig, P.A.; Alpern, R.J. Chronic metabolic acidosis causes an adaptation in the apical membrane Na/H antiporter and basolateral membrane Na(HCO3)3 symporter in the rat proximal convoluted tubule. J. Clin. Investig. 1988, 82, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Purkerson, J.M.; Tsuruoka, S.; Zachary Suter, D.; Nakamori, A.; Schwartz, G.J. Adaptation to metabolic acidosis and its recovery are associated with changes in anion exchanger distribution and expression in the cortical collecting duct. Kidney Int. 2010, 78, 993–1005. [Google Scholar] [CrossRef] [PubMed]
- Faroqui, S.; Sheriff, S.; Amlal, H. Metabolic acidosis has dual effects on sodium handling by rat kidney. Am. J. Physiol. Ren. Physiol. 2006, 291, F322–F331. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-J.; Wang, Y.-F.; Hu, H.-J.; Wang, J.-H.; Lee, T.-H.; Hwang, P.-P. Compensatory regulation of Na+ absorption by Na+/H+ exchanger and Na+-Cl− cotransporter in zebrafish (Danio rerio). Front. Zool. 2013, 10, 46. [Google Scholar] [CrossRef] [PubMed]
- Kumai, Y.; Kwong, R.W.M.; Perry, S.F. A role for transcription factor glial cell missing 2 in Ca2+ homeostasis in zebrafish, Danio rerio. Pflügers Arch. 2015, 467, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Nijenhuis, T.; Renkema, K.Y.; Hoenderop, J.G.J.; Bindels, R.J.M. Acid-base status determines the renal expression of Ca2+ and Mg2+ transport proteins. J. Am. Soc. Nephrol. 2006, 17, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Milligan, C.L.; Wood, C.M. Disturbances in Haematology, Fluid Volume Distribution and Circulatory Function Associated with Low Environmental pH in the Rainbow Trout, Salmo Gairdneri. J. Exp. Biol. 1982, 99, 397–415. [Google Scholar]
- Wood, C.M. The physiological problems of fish in acid waters. In Acid Toxicity and Aquatic Animals; Morris, R., Taylor, E.W., Brown, D.J.A., Brown, J.A., Eds.; Cambridge University Press: Cambridge, UK, 1989; pp. 125–152. [Google Scholar]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.A.; Wood, C.M. Seven things fish know about ammonia and we don’t. Respir. Physiol. Neurobiol. 2012, 184, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Kumai, Y.; Perry, S.F. Mechanisms and regulation of Na+ uptake by freshwater fish. Respir. Physiol. Neurobiol. 2012, 184, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.-P.; Lee, T.-H.; Lin, L.-Y. Ion regulation in fish gills: Recent progress in the cellular and molecular mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R28–R47. [Google Scholar] [CrossRef] [PubMed]
- Meneton, P. Comparative roles of the renal apical sodium transport systems in blood pressure control. J. Am. Soc. Nephrol. 2000, 11 (Suppl. S1), S135–S139. [Google Scholar] [PubMed]
- Shih, T.-H.; Horng, J.-L.; Liu, S.-T.; Hwang, P.-P.; Lin, L.-Y. Rhcg1 and NHE3b are involved in ammonium-dependent sodium uptake by zebrafish larvae acclimated to low-sodium water. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R84–R93. [Google Scholar] [CrossRef] [PubMed]
- Esaki, M.; Hoshijima, K.; Kobayashi, S.; Fukuda, H.; Kawakami, K.; Hirose, S. Visualization in zebrafish larvae of Na+ uptake in mitochondria-rich cells whose differentiation is dependent on foxi3a. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 292, R470–R480. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, S.; Bostanjoglo, M.; Schmitt, R.; Ellison, D.H. Sodium transport-related proteins in the mammalian distal nephron—Distribution, ontogeny and functional aspects. Anat. Embryol. 1999, 200, 447–468. [Google Scholar] [CrossRef] [PubMed]
- Fanestil, D.D.; Vaughn, D.A.; Blakely, P. Metabolic acid-base influences on renal thiazide receptor density. Am. J. Physiol. 1997, 272, R2004–R2008. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Martin, S.W.; Fernández-Llama, P.; Masilamani, S.; Packer, R.K.; Knepper, M.A. Long-term regulation of renal Na-dependent cotransporters and ENaC: Response to altered acid-base intake. Am. J. Physiol. Ren. Physiol. 2000, 279, F459–F467. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.-W.; Yang, S.-S.; Cheng, C.-J.; Tseng, M.-H.; Hsu, H.-M.; Lin, S.-H. Chronic Metabolic Acidosis Activates Renal Tubular Sodium Chloride Cotransporter through Angiotension II-dependent WNK4-SPAK Phosphorylation Pathway. Sci. Rep. 2016, 6, 18360. [Google Scholar] [CrossRef] [PubMed]
- Kwong, R.W.M.; Perry, S.F. The tight junction protein claudin-b regulates epithelial permeability and sodium handling in larval zebrafish, Danio rerio. Am. J. Physiol. Integr. Comp. Physiol. 2013, 304, R504–R513. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, S.; Wang, Z.; Ma, L.; Soleimani, M. Regulation of the apical Cl−/HCO exchanger pendrin in rat cortical collecting duct in metabolic acidosis. Am. J. Physiol. Ren. Physiol. 2003, 284, F103–F112. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.A.; Finberg, K.E.; Stehberger, P.A.; Lifton, R.P.; Giebisch, G.H.; Aronson, P.S.; Geibel, J.P. Regulation of the expression of the Cl−/anion exchanger pendrin in mouse kidney by acid-base status. Kidney Int. 2002, 62, 2109–2117. [Google Scholar] [CrossRef] [PubMed]
- Bayaa, M.; Vulesevic, B.; Esbaugh, A.; Braun, M.; Ekker, M.E.; Grosell, M.; Perry, S.F. The involvement of SLC26 anion transporters in chloride uptake in zebrafish (Danio rerio) larvae. J. Exp. Biol. 2009, 212, 3283–3295. [Google Scholar] [CrossRef] [PubMed]
- Huber, S.; Asan, E.; Jöns, T.; Kerscher, C.; Püschel, B.; Drenckhahn, D. Expression of rat kidney anion exchanger 1 in type A intercalated cells in metabolic acidosis and alkalosis. Am. J. Physiol. 1999, 277, F841–F849. [Google Scholar] [CrossRef] [PubMed]
- Boron, W.F. Acid-Base Transport by the Renal Proximal Tubule. J. Am. Soc. Nephrol. 2006, 17, 2368–2382. [Google Scholar] [CrossRef] [PubMed]
- Burnham, C.E.; Flagella, M.; Wang, Z.; Amlal, H.; Shull, G.E.; Soleimani, M. Cloning, renal distribution, and regulation of the rat Na+-HCO3− cotransporter. Am. J. Physiol. 1998, 274, F1119–F1126. [Google Scholar] [PubMed]
- Soleimani, M.; Bizal, G.L.; McKinney, T.D.; Hattabaugh, Y.J. Effect of in vitro metabolic acidosis on luminal Na+/H+ exchange and basolateral Na+:HCO3− cotransport in rabbit kidney proximal tubules. J. Clin. Investig. 1992, 90, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, T.; Inatomi, J.; Sekine, T.; Cha, S.H.; Kanai, Y.; Kunimi, M.; Tsukamoto, K.; Satoh, H.; Shimadzu, M.; Tozawa, F.; et al. Mutations in SLC4A4 cause permanent isolated proximal renal tubular acidosis with ocular abnormalities. Nat. Genet. 1999, 23, 264–266. [Google Scholar] [CrossRef] [PubMed]
- Dinour, D.; Chang, M.-H.; Satoh, J.; Smith, B.L.; Angle, N.; Knecht, A.; Serban, I.; Holtzman, E.J.; Romero, M.F. A Novel Missense Mutation in the Sodium Bicarbonate Cotransporter (NBCe1/SLC4A4) Causes Proximal Tubular Acidosis and Glaucoma through Ion Transport Defects. J. Biol. Chem. 2004, 279, 52238–52246. [Google Scholar] [CrossRef] [PubMed]
- Myers, E.J.; Yuan, L.; Felmlee, M.A.; Lin, Y.-Y.; Jiang, Y.; Pei, Y.; Wang, O.; Li, M.; Xing, X.-P.; Marshall, A.; et al. A novel mutant Na+/HCO3− cotransporter NBCe1 in a case of compound-heterozygous inheritance of proximal renal tubular acidosis. J. Physiol. 2016, 594, 6267–6286. [Google Scholar] [CrossRef] [PubMed]
- Gawenis, L.R.; Bradford, E.M.; Prasad, V.; Lorenz, J.N.; Simpson, J.E.; Clarke, L.L.; Woo, A.L.; Grisham, C.; Sanford, L.P.; Doetschman, T.; et al. Colonic anion secretory defects and metabolic acidosis in mice lacking the NBC1 Na+/HCO3− cotransporter. J. Biol. Chem. 2007, 282, 9042–9052. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, C.-D.; You, M.-S.; Guh, Y.-J.; Ma, M.; Jiang, Y.-J.; Hwang, P.-P. A Positive Regulatory Loop between foxi3a and foxi3b Is Essential for Specification and Differentiation of Zebrafish Epidermal Ionocytes. PLoS ONE 2007, 2, e302. [Google Scholar] [CrossRef] [PubMed]
- Van de Graaf, S.F.J.; Bindels, R.J.M.; Hoenderop, J.G.J. Physiology of epithelial Ca2+ and Mg2+ transport. Rev. Physiol. Biochem. Pharmacol. 2007, 158, 77–160. [Google Scholar] [PubMed]
- Blaine, J.; Chonchol, M.; Levi, M. Renal control of calcium, phosphate, and magnesium homeostasis. Clin. J. Am. Soc. Nephrol. 2015, 10, 1257–1272. [Google Scholar] [CrossRef] [PubMed]
- Dimke, H.; Hoenderop, J.G.J.; Bindels, R.J.M. Molecular basis of epithelial Ca2+ and Mg2+ transport: Insights from the TRP channel family. J. Physiol. 2011, 589, 1535–1542. [Google Scholar] [CrossRef] [PubMed]
- Bindels, R.J.; Hartog, A.; Abrahamse, S.L.; Van Os, C.H. Effects of pH on apical calcium entry and active calcium transport in rabbit cortical collecting system. Am. J. Physiol. 1994, 266, F620–F627. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.; Hoenderop, J.G.J.; van Os, C.H.; Bindels, R.J.M. The epithelial calcium channel, ECaC1: Molecular details of a novel player in renal calcium handling. Nephrol. Dial. Transplant. 2001, 16, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Vanoevelen, J.; Janssens, A.; Huitema, L.F.A.; Hammond, C.L.; Metz, J.R.; Flik, G.; Voets, T.; Schulte-Merker, S. Trpv5/6 is vital for epithelial calcium uptake and bone formation. FASEB J. 2011, 25, 3197–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumai, Y.; Nesan, D.; Vijayan, M.M.; Perry, S.F. Cortisol regulates Na+ uptake in zebrafish, Danio rerio, larvae via the glucocorticoid receptor. Mol. Cell. Endocrinol. 2012, 364, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Cruz, S.A.; Lin, C.-H.; Chao, P.-L.; Hwang, P.-P. Glucocorticoid receptor, but not mineralocorticoid receptor, mediates cortisol regulation of epidermal ionocyte development and ion transport in zebrafish (Danio rerio). PLoS ONE 2013, 8, e77997. [Google Scholar] [CrossRef] [PubMed]
- Cruz, S.A.; Chao, P.-L.; Hwang, P.-P. Cortisol promotes differentiation of epidermal ionocytes through Foxi3 transcription factors in zebrafish (Danio rerio). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 164, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Kumai, Y.; Kwong, R.W.M.; Perry, S.F. The role of cAMP-mediated intracellular signaling in regulating Na+ uptake in zebrafish larvae. Am. J. Physiol. Integr. Comp. Physiol. 2014, 306, R51–R60. [Google Scholar] [CrossRef] [PubMed]
- Kwong, R.W.M.; Perry, S.F. Hydrogen sulfide promotes calcium uptake in larval zebrafish. Am. J. Physiol. Cell Physiol. 2015, 309, C60–C69. [Google Scholar] [CrossRef] [PubMed]
- Werner, A.; Dehmelt, L.; Nalbant, P. Na+-dependent phosphate cotransporters: The NaPi protein families. J. Exp. Biol. 1998, 201, 3135–3142. [Google Scholar] [PubMed]
- Biber, J.; Hernando, N.; Forster, I.; Murer, H. Regulation of phosphate transport in proximal tubules. Pflügers Arch. 2009, 458, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Ambühl, P.M.; Zajicek, H.K.; Wang, H.; Puttaparthi, K.; Levi, M. Regulation of renal phosphate transport by acute and chronic metabolic acidosis in the rat. Kidney Int. 1998, 53, 1288–1298. [Google Scholar] [CrossRef] [PubMed]
- Nowik, M.; Picard, N.; Stange, G.; Capuano, P.; Tenenhouse, H.S.; Biber, J.; Murer, H.; Wagner, C.A. Renal phosphaturia during metabolic acidosis revisited: Molecular mechanisms for decreased renal phosphate reabsorption. Pflügers Arch. 2008, 457, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, C.; Nalbant, P.; Schölermann, B.; Hentschel, H.; Kinne, R.K.H.; Werner, A. Characterization of a type IIb sodium-phosphate cotransporter from zebrafish (Danio rerio) kidney. Am. J. Physiol. Ren. Physiol. 2003, 284, F727–F736. [Google Scholar] [CrossRef] [PubMed]
- Nalbant, P.; Boehmer, C.; Dehmelt, L.; Wehner, F.; Werner, A. Functional characterization of a Na+-phosphate cotransporter (NaPi-II) from zebrafish and identification of related transcripts. J. Physiol. 1999, 520 Pt 1, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Beene, L.C.; Halluer, J.; Yoshinaga, M.; Hamdi, M.; Liu, Z. Pentavalent arsenate transport by zebrafish phosphate transporter NaPi-IIb1. Zebrafish 2011, 8, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, K.J.; Murer, H. Sulphate and phosphate transport in the renal proximal tubule. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1982, 299, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Puttaparthi, K.; Markovich, D.; Halaihel, N.; Wilson, P.; Zajicek, H.K.; Wang, H.; Biber, J.; Murer, H.; Rogers, T.; Levi, M. Metabolic acidosis regulates rat renal Na-Si cotransport activity. Am. J. Physiol. 1999, 276, C1398–C1404. [Google Scholar] [CrossRef] [PubMed]
- Markovich, D.; Romano, A.; Storelli, C.; Verri, T. Functional and structural characterization of the zebrafish Na+-sulfate cotransporter 1 (NaS1) cDNA and gene (slc13a1). Physiol. Genom. 2008, 34, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Wingert, R.A.; Selleck, R.; Yu, J.; Song, H.-D.; Chen, Z.; Song, A.; Zhou, Y.; Thisse, B.; Thisse, C.; McMahon, A.P.; et al. The cdx Genes and Retinoic Acid Control the Positioning and Segmentation of the Zebrafish Pronephros. PLoS Genet. 2007, 3, e189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumz, M.L.; Lynch, I.J.; Greenlee, M.M.; Cain, B.D.; Wingo, C.S. The renal H+-K+-ATPases: Physiology, regulation, and structure. Am. J. Physiol. Renal Physiol. 2010, 298, F12–F21. [Google Scholar] [CrossRef] [PubMed]
- Cheval, L.; Morla, L.; Elalouf, J.-M.; Doucet, A. Kidney collecting duct acid-base “regulon”. Physiol. Genom. 2006, 27, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Silver, R.B.; Mennitt, P.A.; Satlin, L.M. Stimulation of apical H-K-ATPase in intercalated cells of cortical collecting duct with chronic metabolic acidosis. Am. J. Physiol. 1996, 270, F539–F547. [Google Scholar] [CrossRef] [PubMed]
- Silver, R.B.; Frindt, G.; Mennitt, P.; Satlin, L.M. Characterization and regulation of H-K-ATPase in intercalated cells of rabbit cortical collecting duct. J. Exp. Zool. 1997, 279, 443–455. [Google Scholar] [CrossRef]
- Guh, Y.-J.; Tseng, Y.-C.; Yang, C.-Y.; Hwang, P.-P. Endothelin-1 regulates H+-ATPase-dependent transepithelial H+ secretion in zebrafish. Endocrinology 2014, 155, 1728–1737. [Google Scholar] [CrossRef] [PubMed]
- Kumai, Y.; Bernier, N.J.; Perry, S.F. Angiotensin-II promotes Na+ uptake in larval zebrafish, Danio rerio, in acidic and ion-poor water. J. Endocrinol. 2014, 220, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Chou, M.-Y.; Lin, C.-H.; Chao, P.-L.; Hung, J.-C.; Cruz, S.A.; Hwang, P.-P. Stanniocalcin-1 controls ion regulation functions of ion-transporting epithelium other than calcium balance. Int. J. Biol. Sci. 2015, 11, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Kumai, Y.; Ward, M.A.R.; Perry, S.F. β-Adrenergic regulation of Na+ uptake by larval zebrafish Danio rerio in acidic and ion-poor environments. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R1031–R1041. [Google Scholar] [CrossRef] [PubMed]
- Guh, Y.-J.; Yang, C.-Y.; Liu, S.-T.; Huang, C.-J.; Hwang, P.-P. Oestrogen-related receptor α is required for transepithelial H+ secretion in zebrafish. Proc. Biol. Sci. 2016, 283, 20152582. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.A.; Edwards, D.; Whitehead, C. Cortisol and thyroid hormone responses to acid stress in the brown trout, Salmo trutta L. J. Fish Biol. 1989, 35, 73–84. [Google Scholar] [CrossRef]
- Flik, G.; van der Velden, J.A.; Seegers, H.C.; Kolar, Z.; Wendelaar Bonga, S.E. Prolactin cell activity and sodium fluxes in tilapia (Oreochromis mossambicus) after long-term acclimation to acid water. Gen. Comp. Endocrinol. 1989, 75, 39–45. [Google Scholar] [CrossRef]
- Furukawa, F.; Watanabe, S.; Kaneko, T.; Uchida, K. Changes in gene expression levels of somatolactin in the pituitary and morphology of gill mitochondria-rich cells in Mozambique tilapia after transfer to acidic freshwater (pH 3.5). Gen. Comp. Endocrinol. 2010, 166, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Kakizawa, S.; Kaneko, T.; Hirano, T. Elevation of plasma somatolactin concentrations during acidosis in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol. 1996, 199, 1043–1051. [Google Scholar]
- Briet, M.; Schiffrin, E.L. Aldosterone: Effects on the kidney and cardiovascular system. Nat. Rev. Nephrol. 2010, 6, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.A. Effect of Mineralocorticoids on Acid-Base Balance. Nephron Physiol. 2014, 128, 26–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prunet, P.; Sturm, A.; Milla, S. Multiple corticosteroid receptors in fish: From old ideas to new concepts. Gen. Comp. Endocrinol. 2006, 147, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, A.K.; Butler, P.C.; White, R.B.; DeMarco, U.; Pearce, D.; Fernald, R.D. Multiple Corticosteroid Receptors in a Teleost Fish: Distinct Sequences, Expression Patterns, and Transcriptional Activities. Endocrinology 2003, 144, 4226–4236. [Google Scholar] [CrossRef] [PubMed]
- Colombe, L.; Fostier, A.; Bury, N.; Pakdel, F.; Guiguen, Y. A mineralocorticoid-like receptor in the rainbow trout, Oncorhynchus mykiss: Cloning and characterization of its steroid binding domain. Steroids 2000, 65, 319–328. [Google Scholar] [CrossRef]
- Lin, C.-H.; Tsai, I.-L.; Su, C.-H.; Tseng, D.-Y.; Hwang, P.-P. Reverse Effect of Mammalian Hypocalcemic Cortisol in Fish: Cortisol Stimulates Ca2+ Uptake via Glucocorticoid Receptor-Mediated Vitamin D3 Metabolism. PLoS ONE 2011, 6, e23689. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Hu, H.-J.; Hwang, P.-P. Cortisol regulates sodium homeostasis by stimulating the transcription of sodium-chloride transporter (NCC) in zebrafish (Danio rerio). Mol. Cell. Endocrinol. 2016, 422, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Kwong, R.W.M.; Perry, S.F. Cortisol regulates epithelial permeability and sodium losses in zebrafish exposed to acidic water. J. Endocrinol. 2013, 217, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Kohan, D.E.; Inscho, E.W.; Wesson, D.; Pollock, D.M. Physiology of endothelin and the kidney. Compr. Physiol. 2011, 1, 883–919. [Google Scholar] [CrossRef] [PubMed]
- Licht, C.; Laghmani, K.; Yanagisawa, M.; Preisig, P.A.; Alpern, R.J. An autocrine role for endothelin-1 in the regulation of proximal tubule NHE3. Kidney Int. 2004, 65, 1320–1326. [Google Scholar] [CrossRef] [PubMed]
- Laghmani, K.; Preisig, P.A.; Moe, O.W.; Yanagisawa, M.; Alpern, R.J. Endothelin-1/endothelin-B receptor-mediated increases in NHE3 activity in chronic metabolic acidosis. J. Clin. Investig. 2001, 107, 1563–1569. [Google Scholar] [CrossRef] [PubMed]
- Chu, T.S.; Peng, Y.; Cano, A.; Yanagisawa, M.; Alpern, R.J. Endothelin(B) receptor activates NHE-3 by a Ca2+-dependent pathway in OKP cells. J. Clin. Investig. 1996, 97, 1454–1462. [Google Scholar] [CrossRef] [PubMed]
- Tsuruoka, S.; Watanabe, S.; Purkerson, J.M.; Fujimura, A.; Schwartz, G.J. Endothelin and nitric oxide mediate adaptation of the cortical collecting duct to metabolic acidosis. Am. J. Physiol. Ren. Physiol. 2006, 291, F866–F873. [Google Scholar] [CrossRef] [PubMed]
- Ranhotra, H.S. The orphan estrogen-related receptor α and metabolic regulation: New frontiers. J. Recept. Signal Transduct. 2015, 35, 565–568. [Google Scholar] [CrossRef] [PubMed]
- Johns, E.J.; Kopp, U.C.; DiBona, G.F.; Johns, E.J.; Kopp, U.C.; DiBona, G.F. Neural Control of Renal Function. In Comprehensive Physiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; ISBN 9780470650714. [Google Scholar]
- Aperia, A.; Ibarra, F.; Svensson, L.B.; Klee, C.; Greengard, P. Calcineurin mediates α-adrenergic stimulation of Na+, K+-ATPase activity in renal tubule cells. Proc. Natl. Acad. Sci. USA 1992, 89, 7394–7397. [Google Scholar] [CrossRef] [PubMed]
- Beach, R.E.; Dubose, T.D. Adrenergic regulation of (Na+, K+)-ATPase activity in proximal tubules of spontaneously hypertensive rats. Kidney Int. 1990, 38, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Gesek, F.A. α1-Adrenergic receptors activate NHE1 and NHE3 through distinct signaling pathways in epithelial cells. Am. J. Physiol. Ren. Physiol. 2001, 280, F415–F425. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Nesbitt, T.; Drezner, M.K.; Friedman, P.A.; Gesek, F.A. Proximal nephron Na+/H+ exchange is regulated by α1A- and α1B-adrenergic receptor subtypes. Mol. Pharmacol. 1997, 52, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Healy, V.; Thompson, C.; Johns, E.J. The adrenergic regulation of proximal tubular Na+/H+ exchanger 3 in the rat. Acta Physiol. 2014, 210, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.A.; Premont, R.T.; Chow, C.-W.; Blitzer, J.T.; Pitcher, J.A.; Claing, A.; Stoffel, R.H.; Barak, L.S.; Shenolikar, S.; Weinman, E.J.; et al. The β2-adrenergic receptor interacts with the Na+/H+-exchanger regulatory factor to control Na+/H+ exchange. Nature 1998, 392, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Jonz, M.G.; Nurse, C.A. Epithelial mitochondria-rich cells and associated innervation in adult and developing zebrafish. J. Comp. Neurol. 2006, 497, 817–832. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.D.; Coffman, T.M. Recent advances involving the renin–angiotensin system. Exp. Cell Res. 2012, 318, 1049–1056. [Google Scholar] [CrossRef] [PubMed]
- Dixit, M.P.; Xu, L.; Xu, H.; Bai, L.; Collins, J.F.; Ghishan, F.K. Effect of angiotensin-II on renal Na+/H+ exchanger-NHE3 and NHE2. Biochim. Biophys. Acta 2004, 1664, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Houillier, P.; Chambrey, R.; Achard, J.M.; Froissart, M.; Poggioli, J.; Paillard, M. Signaling pathways in the biphasic effect of angiotensin II on apical Na/H antiport activity in proximal tubule. Kidney Int. 1996, 50, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Schuster, V.L.; Kokko, J.P.; Jacobson, H.R. Angiotensin II directly stimulates sodium transport in rabbit proximal convoluted tubules. J. Clin. Investig. 1984, 73, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Pech, V.; Zheng, W.; Pham, T.D.; Verlander, J.W.; Wall, S.M. Angiotensin II Activates H+-ATPase in Type A Intercalated Cells. J. Am. Soc. Nephrol. 2008, 19, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Talati, G.; Ohta, A.; Rai, T.; Sohara, E.; Naito, S.; Vandewalle, A.; Sasaki, S.; Uchida, S. Effect of angiotensin II on the WNK-OSR1/SPAK-NCC phosphorylation cascade in cultured mpkDCT cells and in vivo mouse kidney. Biochem. Biophys. Res. Commun. 2010, 393, 844–848. [Google Scholar] [CrossRef] [PubMed]
- San-Cristobal, P.; Pacheco-Alvarez, D.; Richardson, C.; Ring, A.M.; Vazquez, N.; Rafiqi, F.H.; Chari, D.; Kahle, K.T.; Leng, Q.; Bobadilla, N.A.; et al. Angiotensin II signaling increases activity of the renal Na-Cl cotransporter through a WNK4-SPAK-dependent pathway. Proc. Natl. Acad. Sci. USA 2009, 106, 4384–4389. [Google Scholar] [CrossRef] [PubMed]
- Ko, B.; Mistry, A.; Hanson, L.; Mallick, R.; Hoover, R.S. Mechanisms of angiotensin II stimulation of NCC are time-dependent in mDCT15 cells. Am. J. Physiol. Ren. Physiol. 2015, 308, F720–F727. [Google Scholar] [CrossRef] [PubMed]
- Sazonova, O.; James, K.A.; McCudden, C.R.; Segal, D.; Talebian, A.; Wagner, G.F. Stanniocalcin-1 secretion and receptor regulation in kidney cells. Am. J. Physiol. Ren. Physiol. 2008, 294, F788–F794. [Google Scholar] [CrossRef] [PubMed]
- Madsen, K.L.; Tavernini, M.M.; Yachimec, C.; Mendrick, D.L.; Alfonso, P.J.; Buergin, M.; Olsen, H.S.; Antonaccio, M.J.; Thomson, A.B.; Fedorak, R.N. Stanniocalcin: A novel protein regulating calcium and phosphate transport across mammalian intestine. Am. J. Physiol. 1998, 274, G96–G102. [Google Scholar] [CrossRef] [PubMed]
- Tseng, D.-Y.; Chou, M.-Y.; Tseng, Y.-C.; Hsiao, C.-D.; Huang, C.-J.; Kaneko, T.; Hwang, P.-P. Effects of stanniocalcin 1 on calcium uptake in zebrafish (Danio rerio) embryo. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 296, R549–R557. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A simplified model illustrating the effects of acute and/or chronic acid exposure on zebrafish ionocytes. NHE, Na+/H+ exchanger; AE, anion exchanger; NCC, Na+-Cl− cotransporter; NBCe, electrogenic Na+-HCO3− cotransporter; ECaC, epithelial Ca2+ channel.HRC, H+-ATPase rich cell (blue); NCCC, NCC expressing cell (green); NaRC, Na+/K+-ATPase rich cell (red). * In acidic water, mRNA encoding NHE3b is increased in larvae but decreased in adult gills.
Figure 1.
A simplified model illustrating the effects of acute and/or chronic acid exposure on zebrafish ionocytes. NHE, Na+/H+ exchanger; AE, anion exchanger; NCC, Na+-Cl− cotransporter; NBCe, electrogenic Na+-HCO3− cotransporter; ECaC, epithelial Ca2+ channel.HRC, H+-ATPase rich cell (blue); NCCC, NCC expressing cell (green); NaRC, Na+/K+-ATPase rich cell (red). * In acidic water, mRNA encoding NHE3b is increased in larvae but decreased in adult gills.


{kind=link}
{kind=link}
Table 1.
Distribution of major epithelial ion transporters in zebrafish ionocytes and mammalian renal cells.
Table 1.
Distribution of major epithelial ion transporters in zebrafish ionocytes and mammalian renal cells.
| Transporters | Gene Name and Cellular Localization of the Protein | Protein Identity between Zebrafish and Humans | |
|---|---|---|---|
| Zebrafish | Mammals | ||
| H+-ATPase | ATP6V1AA. HRCs (apical) [12,13] | ATP6V1A. Proximal tubular cells (apical), Type A (apical) and type B (basolateral) intercalated cells [14,15,16] | 93% |
| NHE3 | NHE3b; SLC9A3.2. HRCs (apical) [17,18] | NHE3; SLC9A3. Proximal tubular cells (apical) [19,20] | 47% |
| AE1 | AE1b; SLC4A1B. HRCs (basolateral) [21] | AE1; SLC4A1. Type A intercalated cells (basolateral) [22] | 53% |
| NBCe1 | NBCe1b; SLC4A4B. NCCCs (basolateral) [21,23] | NBCe1-A ‡; SLC4A4. Proximal tubular cells (basolateral) [24,25] | 78% |
| NCC | NCC like 2; SLC12A10.2. NCCCs (apical) [26,27] | NCC; SLC12A3. Distal convoluted tubular cells (apical) [28,29,30] | 53% |
| ECaC | ECaC; TRPV5. NaRCs (apical) [31,32] | ECaC; TRPV5. Distal convoluted tubular cells and principal cells (apical) [33,34,35] | 48% |
| ENaC | N/A * | ENaC. Principal cells (apical) [36,37] | N/A * |
NHE, Na+/H+ exchanger; AE, anion exchanger; NBCe, electrogenic Na+-HCO3− cotransporter; NCC, Na+-Cl− cotransporter; ECaC, epithelial Ca2+ channel; ENaC, epithelial Na+ channel. HRCs, H+-ATPase-rich cells; NCCCs, Na+-Cl− cotransporter expressing cells; NaRCs, Na+/K+-ATPase-rich cells. TRPV, transient receptor potential cation channel. * The gene encoding for ENaC has not been identified in teleosts. ‡ NBCe1-A is a kidney-specific splice variant of NBCe1.
Table 2.
The in vivo effects of acid exposure on the regulation of major epithelial ion transporters in zebrafish ionocytes and mammalian renal cells.
Table 2.
The in vivo effects of acid exposure on the regulation of major epithelial ion transporters in zebrafish ionocytes and mammalian renal cells.
| Ion Transporters | Expression Levels/Activity | |
|---|---|---|
| Zebrafish * | Mammals | |
| H+-ATPase | Chronic: ↑ H+-ATPase mRNA expression ↑ H+-ATPase activity ↑ HRCs density [12,38,39] | Acute: ↑ H+-ATPase mRNA (rabbit) [40] |
| Chronic: ↑ H+-ATPase protein abundance (rabbit) [41] | ||
| NHE3 | Chronic: ↑ nhe3b mRNA expression [38] ↑ NHE3b activity [42] | Chronic: ↑ NHE3 protein abundance (rat) [43,44] ↑ NHE activity (rat) [45] |
| AE1 | Chronic: ↑ ae1b mRNA expression [38] | Chronic ↑ AE1-expressing cells ↑ AE1 protein abundance (rabbit) [46] |
| NCC | Acute: ↔ ncc mRNA expression ↔ NCCCs density ↑ NCC activity [26] | Acute: ↔ NCC protein abundance (rat) [47] |
| Chronic: ↑ ncc mRNA expression ↑ NCCCs density [48] | Chronic: ↑ NCC protein abundance (rat) [47] | |
| ECaC | Chronic: ↑ ecac-expressing cells ↑ ECaC activity [49] | Chronic: ↓ ecac (TRPV5) mRNA expression ↓ ECaC protein abundance (mice) [50] |
↑ Increase, ↓ decrease, ↔ no change. * Regulation of ion transporters at larval stages (0 to 4 days post-fertilization). Acute: ≤24 h of exposure, chronic: >1 day of exposure.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lewis, L.; Kwong, R.W.M. Zebrafish as a Model System for Investigating the Compensatory Regulation of Ionic Balance during Metabolic Acidosis. Int. J. Mol. Sci. 2018, 19, 1087. https://doi.org/10.3390/ijms19041087
AMA Style
Lewis L, Kwong RWM. Zebrafish as a Model System for Investigating the Compensatory Regulation of Ionic Balance during Metabolic Acidosis. International Journal of Molecular Sciences. 2018; 19(4):1087. https://doi.org/10.3390/ijms19041087
Chicago/Turabian StyleLewis, Lletta, and Raymond W. M. Kwong. 2018. "Zebrafish as a Model System for Investigating the Compensatory Regulation of Ionic Balance during Metabolic Acidosis" International Journal of Molecular Sciences 19, no. 4: 1087. https://doi.org/10.3390/ijms19041087
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.