Genome-Wide Analyses of the NAC Transcription Factor Gene Family in Pepper (Capsicum annuum L.): Chromosome Location, Phylogeny, Structure, Expression Patterns, Cis-Elements in the Promoter, and Interaction Network

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Identification of NAC Family Members in the Capsicum Annuum Genome
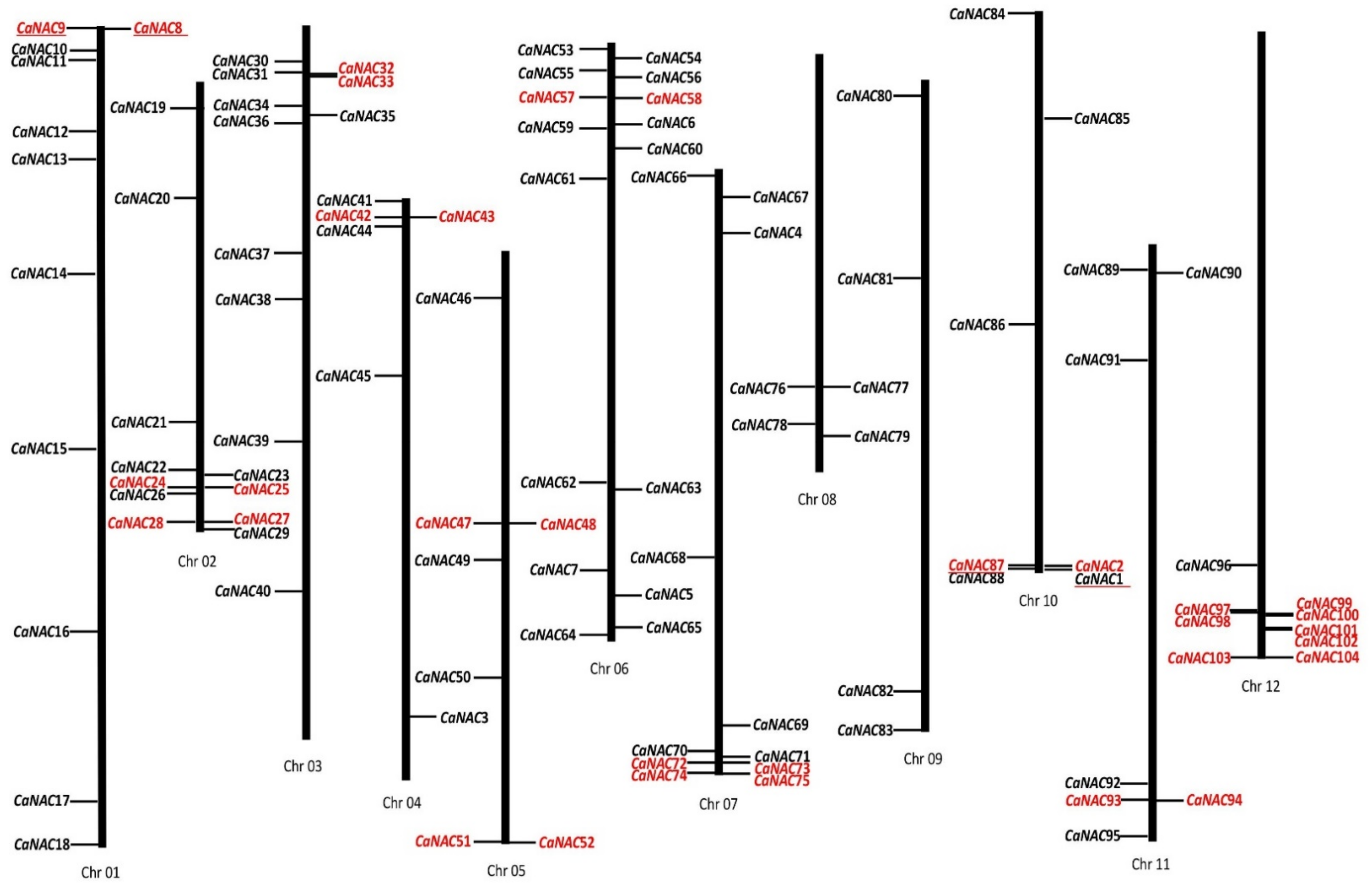
2.2. Chromosomal Distribution and Duplication of CaNAC Genes
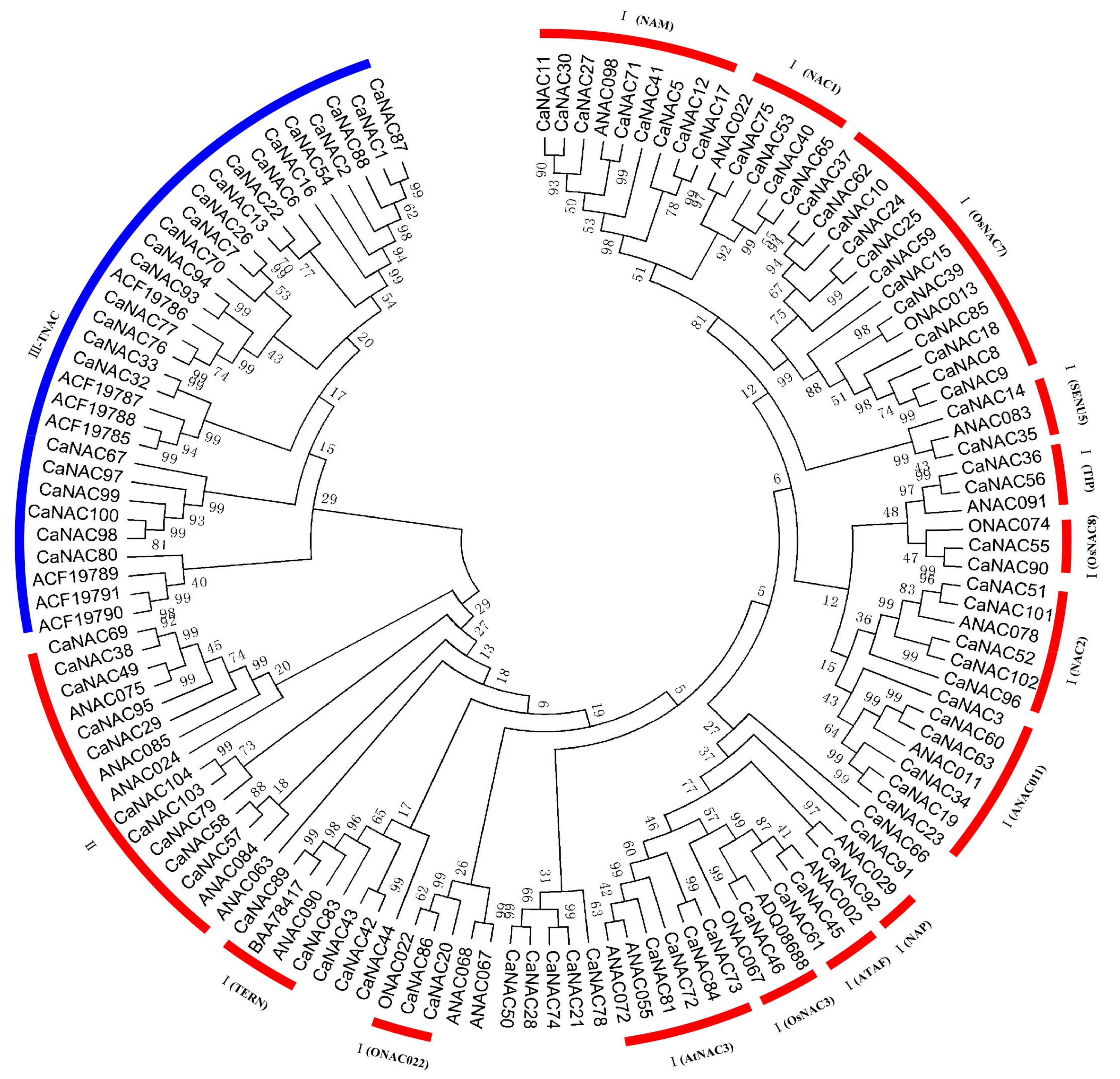
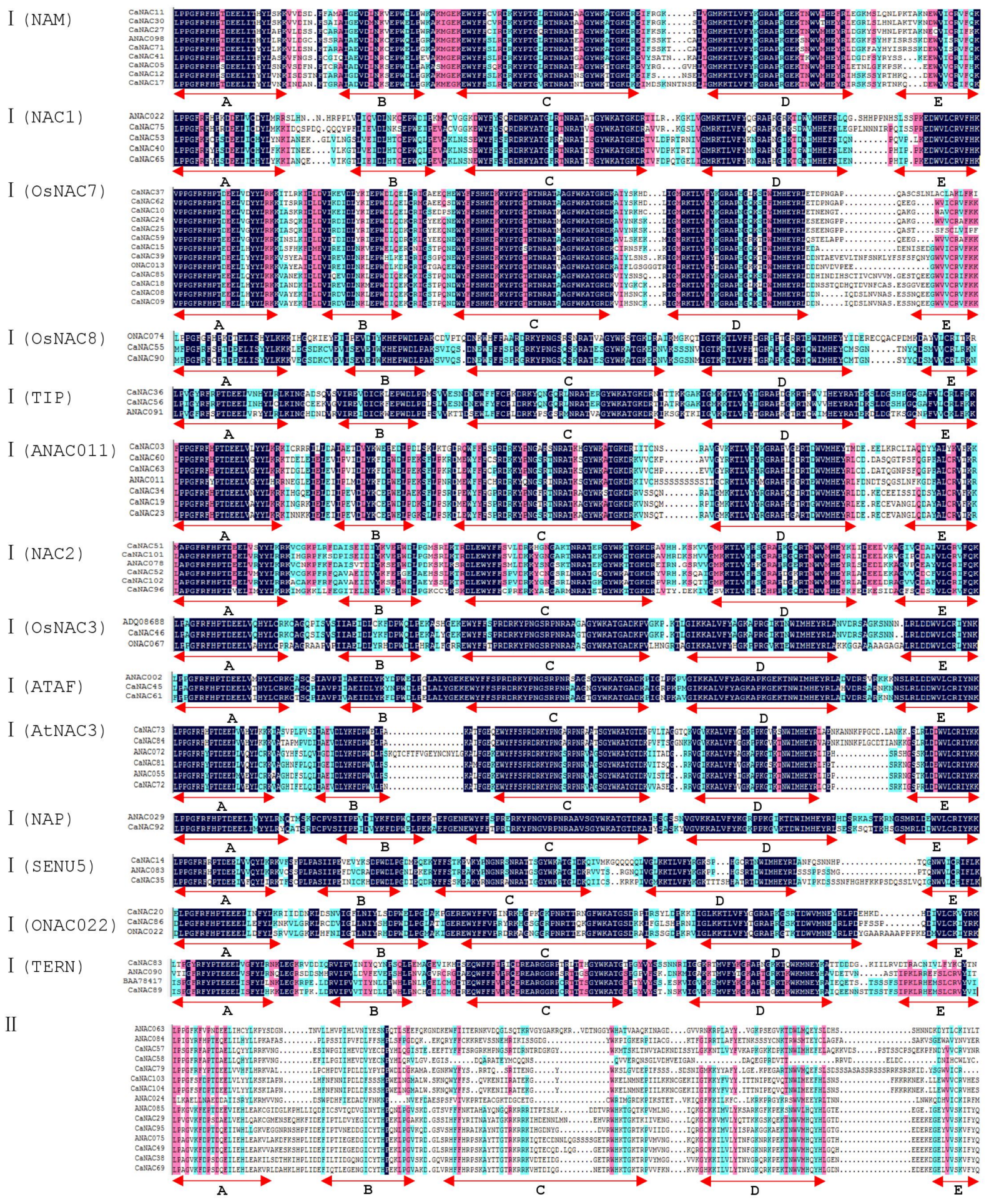
2.3. Classification of CaNACs
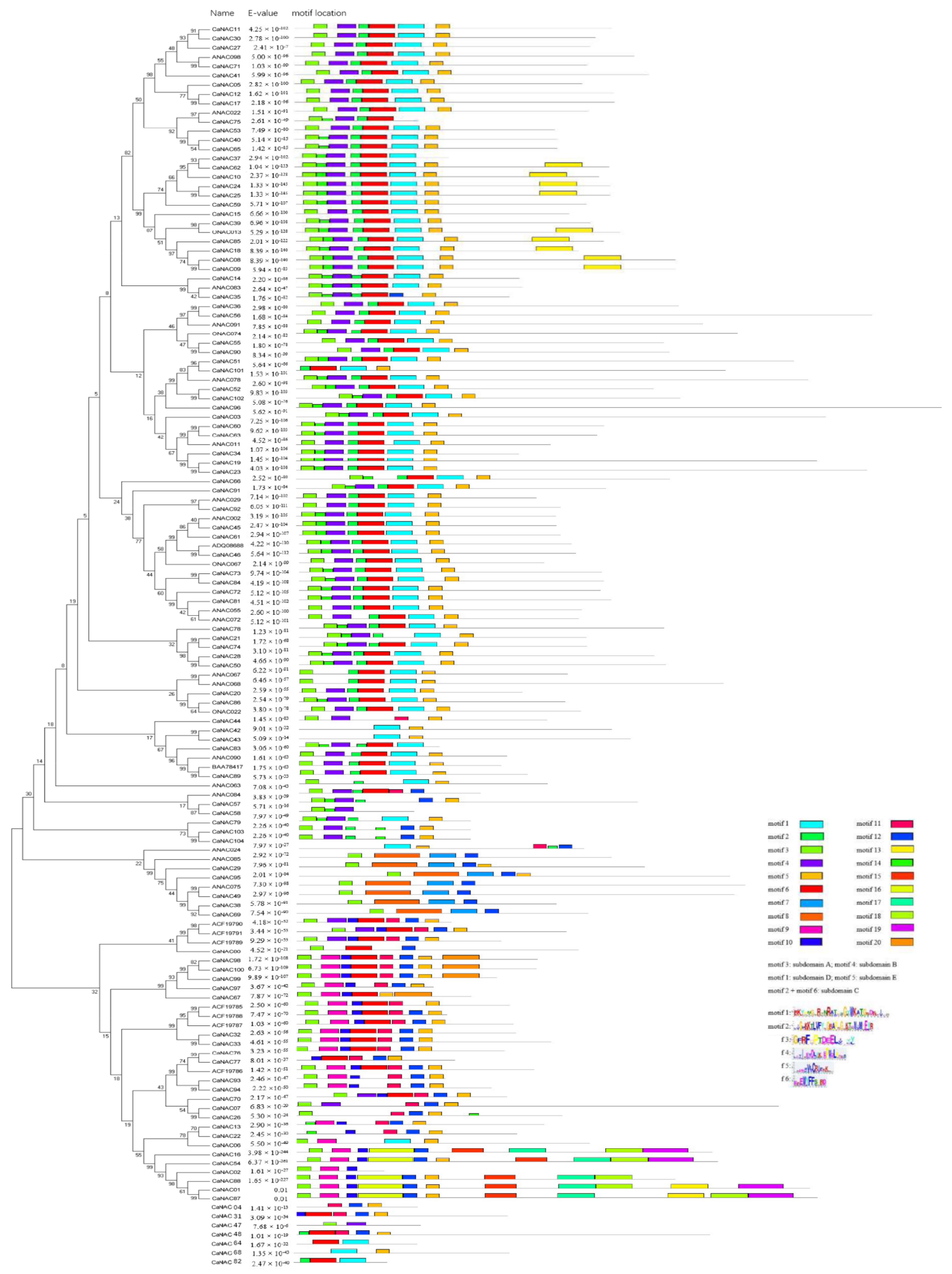
2.4. Gene Structure and Motif Composition Analysis of CaNAC Proteins
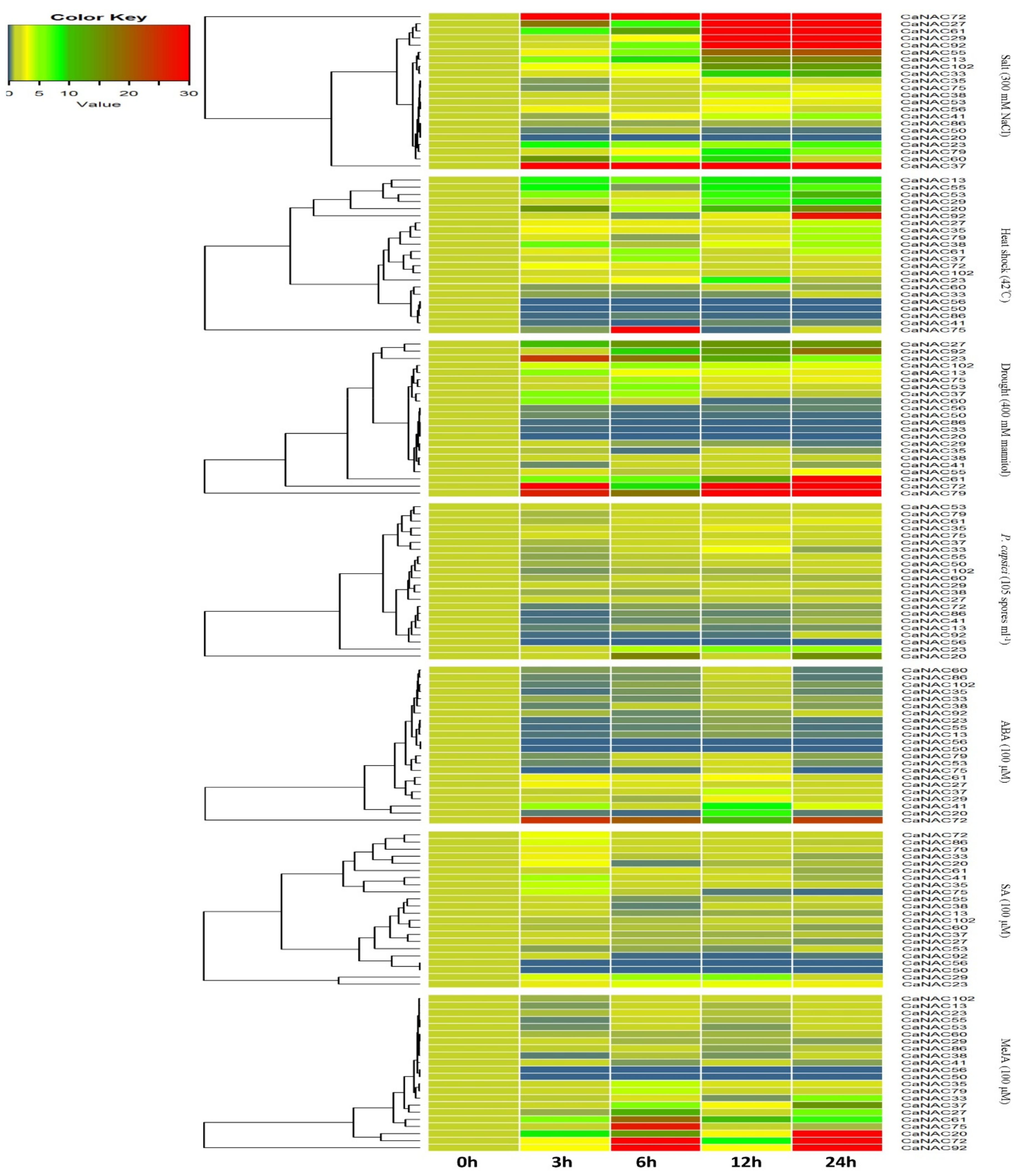
2.5. Expression Profiling of CaNAC Genes under Biotic and Abiotic Stresses
2.5.1. Expression of Patterns of CaNAC Genes under Salt, Heat, and Drought Treatment
2.5.2. Expression Patterns of CaNAC Genes under P. capsici Inoculation
2.5.3. Expression Patterns of CaNAC Genes under Hormone Treatment
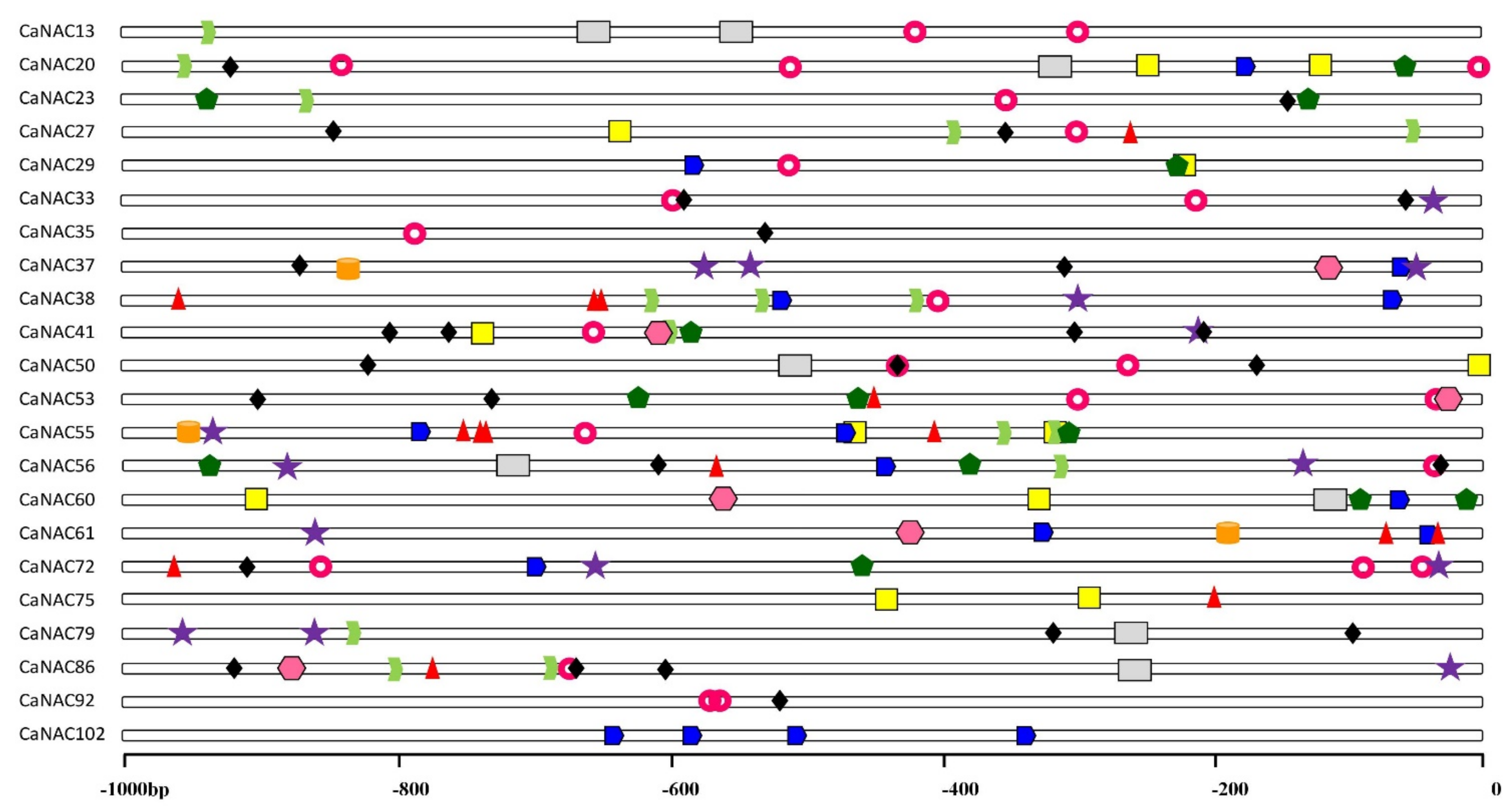
2.6. Analysis of Stress-Related Cis-Elements in the CaNAC Promoters
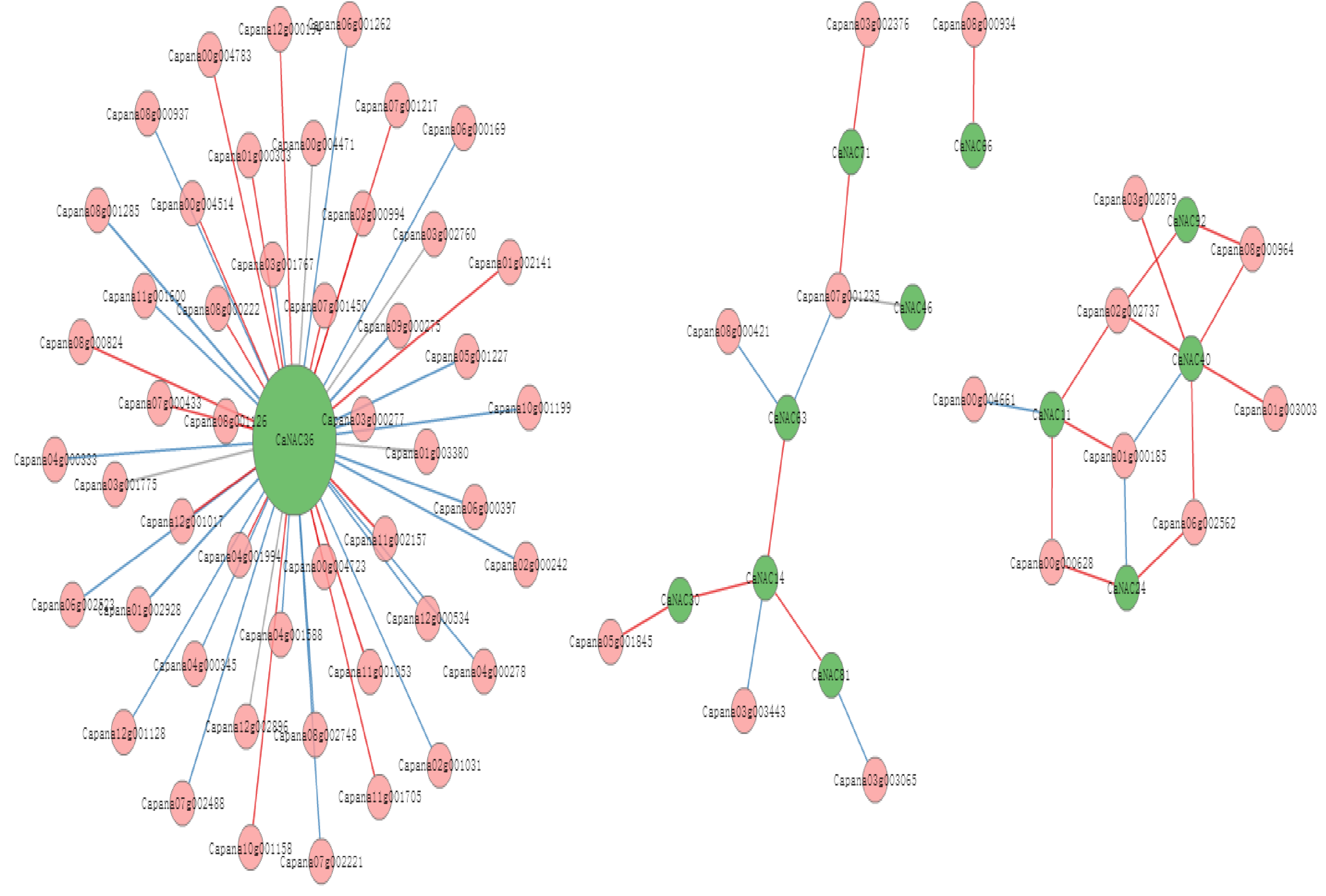
2.7. The Interaction Network of CaNAC Genes
3. Discussion
3.1. NAC Gene Expansion and Evolution in Pepper
3.2. CaNAC Proteins Play Important Roles in Various Biological Processes
4. Materials and Methods
4.1. Sequence Database Searches
4.2. Multiple Sequence Alignment, Gene Chromosomal Location, and Phylogenetics Analysis
4.3. Motif Composition Analysis of CaNAC Proteins
4.4. Treatments of Pepper Plants with Various Biotic and Abiotic Stresses
4.5. Real-Time Quantitative RT-PCR
4.6. Search for Cis-Acting Elements in the Promoters of CaNAC Genes
4.7. Prediction of CaNACs Protein–Protein Interaction Network
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Roychoudhury, A. WRKY Proteins: Signaling and regulation of expression during abiotic stress responses. Sci. World J. 2015, 2015, 807560. [Google Scholar] [CrossRef] [PubMed]
- Hisako, O.; Kouji, S.; Koji, D.; Toshifumi, N.; Yasuhiro, O.; Kazuo, M.; Kenichi, M.; Naoki, O.; Jun, K.; Piero, C.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar]
- Souer, E.; Houwelingen, A.; Kloos, D.; Mol, J.; Koes, R. The No Apical Meristem gene of Petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, N.; Ramaswamy, M.; Akhter, M.S.; Kouji, S.; Hiroaki, K.; Hisako, O.; Shoshi, K. Genome-wide analysis of NAC transcription factor family in rice. Gene 2010, 465, 30–44. [Google Scholar]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Mochida, K.; Yamaguchi, S.K.; Shinozaki, K.; Tran, L.S.P. Genome-wide survey and expression analysis of the plant-specific NAC transcription factor family in soybean during development and dehydration stress. DNA Res. 2011, 18, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Su, H.Y.; Zhang, S.Z.; Yuan, X.W.; Chen, C.T.; Wang, X.F.; Hao, Y.J. Genome-wide analysis and identification of stress-responsive genes of the NAM-ATAF1, 2-CUC2 transcription factor family in apple. Plant Physiol. Biochem. 2013, 71, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Puranik, S.; Sahu, P.P.; Mandal, S.N.; Venkata, S.B.; Parida, S.K.; Prasad, M. Comprehensive genome-wide survey, genomic constitution and expression profiling of the NAC transcription factor family in foxtail millet (Setaria italica L.). PLoS ONE 2013, 8, e64594. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Sharma, V.; Pal, A.K.; Acharya, V.; Ahuja, P.S. Genome-wide organization and expression profiling of the NAC transcription factor family in potato (Solanum tuberosum L.). DNA Res. 2013, 20, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, F.; Li, M.Y.; Jiang, Q.; Tan, G.F.; Xiong, A.S. Genome wide analysis of the NAC transcription factor family in Chinese cabbage of elucidate responses to temperature stress. Sci. Hortic. 2014, 165, 82–90. [Google Scholar] [CrossRef]
- Hu, W.; Wei, Y.X.; Xia, Z.Q.; Yan, Y.; Hou, Z.W.; Zou, M.L.; Lu, C.; Wang, W.Q.; Peng, M. Genome-wide identification and expression analysis of the NAC transcription factor family in cassava. PLoS ONE 2015, 10, e0136993. [Google Scholar] [CrossRef]
- Shiriga, K.; Sharma, R.; Kumar, K.; Yadav, S.K.; Hossain, F.; Thirunavukkarasu, N. Genome-wide identification and expression pattern of drought-responsive members of the NAC family in maize. Meta Gene 2014, 2, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.W.; Gao, L.W.; Zhang, Y.D.; Zhang, F.R.; Yang, X.; Huang, D.F. Genome-wide investigation of the NAC transcription factor family in melon (Cucumis melon) and their expression analysis under salt stress. Plant Cell Rep. 2016, 35, 1827–1839. [Google Scholar] [CrossRef] [PubMed]
- Borrill, P.; Harrington, S.A.; Uauy, C. Genome-wide sequence and expression analysis of the NAC transcription factor family in polyploidy wheat. G3 Genes Genomes Genet. 2017, 7, 3019–3029. [Google Scholar]
- Xu, Z.Y.; Kim, S.Y.; Hyeon, D.Y.; Kim, D.H.; Dong, T.; Park, Y.; Jin, J.B.; Joo, S.; Kim, S.; Hong, J.C.; et al. The Arabidopsis NAC transcription factor ANAC096 cooperates with bZIP-type transcription factors in dehydration and osmotic stress responses. Plant Cell 2013, 25, 4708–4724. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.S.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.J.; Liao, K.F.; Du, H.; Xu, Y.; Song, H.Z.; Li, X.H.; Xiong, L. A stress-responsive NAC transcription factor SNAC3 confers heat and drought tolerance through modulation of reactive oxygen species in rice. J. Exp. Bot. 2013, 66, 6803–6817. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Ouyang, Z.G.; Zhang, Y.F.; Li, X.H.; Hong, Y.B.; Huang, L.; Liu, S.X.; Zhang, H.J.; Li, D.Y.; Song, F.M. Tomato NAC transcription factor SlSRN1 positively regulates defense response against biotic stress but negatively regulates abiotic stress response. PLoS ONE 2014, 9, e102067. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.K.; Lee, S.; Yu, S.H.; Choi, D. Expression of a novel NAC motif-containing transcription factor (CaNAC1) is preferentially associated with incompatible interactions between chili pepper and pathogens. Planta 2005, 222, 876–887. [Google Scholar] [CrossRef] [PubMed]
- Lin, J. The Cloning and Function Characterization of Transcription Factor CaNAC72 of Capsicum annuum. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2012. [Google Scholar]
- Guo, W.L.; Wang, S.B.; Chen, R.G.; Chen, B.H.; Du, X.H.; Yin, Y.X.; Gong, Z.H.; Zhang, Y.Y. Characterization and expression profile of CaNAC2 pepper gene. Front. Plant Sci. 2015, 6, 755. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.S.; Gadnayak, A.; Mandal, S.; Dehury, B. Genome wide screening, identification and characterization of NAC transcription factors from Capsicum Species: An in silico approach. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 527–562. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Sharoni, A.M.; Kikuchi, S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front. Microbiol. 2013, 4, 248. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Park, M. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yu, C.; Shen, Y. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushton, P.J.; Bokowiec, M.T.; Han, S.C. Tobacco transcription factors: Novel insights into transcriptional regulation in the Solanaceae. Plant Physiol. 2008, 147, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Xu, Y.Y.; Wang, X.; Du, C.; Du, J.Z.; Yuan, M.; Xu, Z.H.; Chong, K. OsRAN2, essential for mitosis, enhances cold tolerance in rice by promoting export of intranuclear tubulin and maintaining cell division under cold stress. Plant Cell Environ. 2011, 34, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.L.; Ji, W.; Ding, X.D.; Bai, X.; Cai, H.; Yang, S.S.; Qian, X.; Sun, M.Z.; Zhu, Y.M. GsVAMP72, a novel Glycine soja R-SNARE protein, is involved in regulating plant salt tolerance and ABA sensitivity. Plant Cell Tissue Organ Cult. 2013, 113, 199–215. [Google Scholar] [CrossRef]
- Chen, L.H.; Hu, W.; Tan, S.L.; Wang, M.; Ma, Z.B.; Zhou, S.Y.; Deng, X.M.; Zhang, Y.; Huang, C.; Yang, G.X.; et al. Genome-wide identification and analysis of MAPK and MAPKK gene families in Brachypodium distachyon. PLoS ONE 2012, 7, e46744. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Hong, S.H.; Kim, Y.W.; Lee, I.H.; Jun, J.H.; Phee, B.; Rupak, T. Gene regulatory cascade of senescence-associated NAC transcription factors activated by ETHYLENE- INSENSITIVE2 mediated leaf senescence signaling in Arabidopsis. J. Exp. Bot. 2014, 65, 4023–4036. [Google Scholar] [CrossRef] [PubMed]
- Voitsik, A.M.; Muench, S.; Deising, H.B.; Voll, L.M. Two recently duplicated maize NAC transcription factor paralogs are induced in response to Colletotrichum graminicola infection. BMC Plant Biol. 2013, 13, 85. [Google Scholar] [CrossRef] [PubMed]
- Lamesch, P.; Berardini, T.Z.; Li, D.; Swarbreck, D.; Wilks, C.; Sasidharan, R. The Arabidopsis information resource (TAIR): Improved gene annotation and new tools. Nucleic Acids Res. 2012, 40, D1202–D1210. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, S.; Satoh, K.; Nagata, T. Collection, mapping, and annotation of over 28,000 cDNA clones from japonica rice. Science 2003, 310, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Diao, W.P.; Wang, S.B.; Liu, J.B.; Pan, B.G.; Guo, G.J.; Ge, W.; Gao, C.Z. Identification and expression analysis of NAC transcription factor induced by Phytophthora blight in pepper. Mol. Plant Breed. 2018, in press. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. ClustalW and ClustalX version2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhu, Z.L.; Kasahara, M.; Zhang, Z. Segmental duplications in the silkworm genome. BMC Genomics 2013, 14, 521. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Barksdale, T.H.; Papavizas, G.C.; Johnston, S.A. Resistance to foliar blight and crown rot of pepper caused by Phytophthora capsici. Plant Dis. 1984, 68, 506–509. [Google Scholar] [CrossRef]
- Diao, W.P.; Wang, S.B.; Liu, J.B.; Pan, B.G.; Guo, G.J.; Ge, W. Genome-wide identification and expression analysis of WRKY gene family in Capsicum annuum L. Front. Plant Sci. 2016, 7, 211. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.B.; Liu, K.W.; Diao, W.P.; Zhi, L.; Ge, W.; Liu, J.B.; Pan, B.G.; Wan, H.J.; Chen, J.F. Evaluation of appropriate reference genes for gene expression studies in pepper by quantitative real-time PCR. Mol. Breed. 2012, 30, 1393–1400. [Google Scholar]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Peer, Y.V.; Rouze, P.; Rombauts, S. Plant CARE, a data base of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Liu, J.H.; Lu, J.P.; Zhai, Y.F.; Wang, H.; Gong, Z.H.; Wang, S.B.; Lu, M.H. Genome-wide analysis of the CaHsp20 gene family in pepper: Comprehensive sequence and expression profile analysis under heat stress. Front. Plant Sci. 2015, 6, 806. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Annotation ID | Chr | Group | Introns | PI | AA | WT | At Orthologous Gene | e-Value |
|---|---|---|---|---|---|---|---|---|---|
| CaNAC1 | Capana00g000175/Ca10g20640 | chr 10 | II(26) | 1 | 4.21 | 569 | 64.52 | AT2G43000 | 0.35 |
| CaNAC2 | Capana00g001681/Ca10g20530 | chr 10 | II(26) | 0 | 8.47 | 100 | 11.80 | AT2G43000 | 0.097 |
| CaNAC3 | Capana00g001918/Ca04g14740 | chr 04 | I(8) | 1 | 4.56 | 586 | 65.85 | AT1G34190 | 1 × 10−13 |
| CaNAC4 | Capana00g003238/Ca07g03900 | chr 07 | - | 0 | 7.45 | 137 | 16.09 | - | - |
| CaNAC5 | Capana00g004670/Ca06g17870 | chr 06 | I(14) | 2 | 6.42 | 317 | 36.12 | AT3G15170 | 4 × 10−21 |
| CaNAC6 | Capana00g004843/Ca06g14960 | chr 06 | II(25) | 0 | 4.88 | 326 | 37.03 | AT5G04400 | 3.3 |
| CaNAC7 | Capana00g004888/Ca06g15350 | chr 06 | II(23) | 0 | 4.36 | 534 | 60.80 | - | - |
| CaNAC8 | Capana01g000033 | chr 01 | I(12) | 2 | 6.79 | 420 | 48.06 | AT1G32770 | 1 × 10−18 |
| CaNAC9 | Capana01g000034 | chr 01 | I(12) | 2 | 6.79 | 420 | 48.06 | AT1G32770 | 1 × 10−18 |
| CaNAC10 | Capana01g000515/Ca01g04740 | chr 01 | I(15) | 2 | 7.34 | 336 | 38.87 | AT1G62700 | 2 × 10−47 |
| CaNAC11 | Capana01g000650/Ca06g18770 | chr 01 | I(14) | 3 | 7.57 | 350 | 38.97 | AT5G39610 | 5 × 10−30 |
| CaNAC12 | Capana01g001228/Ca01g09650 | chr 01 | I(14) | 2 | 7.70 | 352 | 39.20 | AT5G18270 | 7 × 10−15 |
| CaNAC13 | Capana01g001521/Ca01g11960 | chr 01 | II(25) | 0 | 5.36 | 276 | 31.72 | AT1G60340 | 2.7 |
| CaNAC14 | Capana01g002000/Ca00g54690 | chr 01 | I(3) | 2 | 10.12 | 248 | 28.34 | AT5G13180 | 1 × 10−8 |
| CaNAC15 | Capana01g002406/Ca01g17160 | chr 01 | I(12) | 1 | 8.07 | 303 | 34.94 | AT4G10350 | 6 × 10−57 |
| CaNAC16 | Capana01g003266/Ca01g26660 | chr 01 | II(26) | 0 | 4.08 | 461 | 52.28 | AT2G43000 | 0.29 |
| CaNAC17 | Capana01g004120/Ca01g30550 | chr 01 | I(14) | 2 | 9.15 | 353 | 39.92 | AT2G24430 | 1 × 10−21 |
| CaNAC18 | Capana01g004489/Ca01g34750 | chr 01 | I(12) | 1 | 6.57 | 313 | 35.95 | AT1G32770 | 3 × 10−25 |
| CaNAC19 | Capana02g000057/Ca02g06450 | chr 02 | I(9) | 5 | 4.92 | 577 | 66.03 | AT3G03200 | 2 × 10−9 |
| CaNAC20 | Capana02g000302/Ca02g03060 | chr 02 | I(2) | 1 | 6.52 | 252 | 29.08 | AT1G62700 | 4 × 10−5 |
| CaNAC21 | Capana02g001393/Ca02g04820 | chr 02 | I(16) | 2 | 6.26 | 322 | 36.74 | AT1G34190 | 6 × 10−11 |
| CaNAC22 | Capana02g002153/Ca02g17910 | chr 02 | II(25) | 0 | 8.56 | 246 | 28.72 | AT5G39690 | 8.5 |
| CaNAC23 | Capana02g002277/Ca02g20290 | chr 02 | I(9) | 5 | 6.27 | 632 | 72.58 | AT3G03200 | 1 × 10−16 |
| CaNAC24 | Capana02g002611/Ca00g89790 | chr 02 | I(15) | 0 | 5.17 | 348 | 39.97 | AT2G18060 | 6 × 10−85 |
| CaNAC25 | Capana02g002612/Ca00g89790 | chr 02 | I(15) | 1 | 5.17 | 348 | 39.97 | AT2G18060 | 1 × 10−75 |
| CaNAC26 | Capana02g002682/Ca02g22730 | chr 02 | II(23) | 0 | 7.09 | 296 | 33.97 | - | - |
| CaNAC27 | Capana02g003352/Ca02g28070 | chr 02 | I(14) | 2 | 7.74 | 326 | 37.31 | AT5G07680 | 3 × 10−37 |
| CaNAC28 | Capana02g003374/Ca02g28290 | chr 02 | I(16) | 1 | 6.52 | 397 | 44.85 | AT5G39820 | 2 × 10−11 |
| CaNAC29 | Capana02g003557/Ca02g30090 | chr 02 | II(19) | 5 | 4.81 | 387 | 43.11 | AT5G18270 | 2 × 10−4 |
| CaNAC30 | Capana03g000802/Ca12g13470 | chr 03 | I(14) | 2 | 7.53 | 332 | 38.10 | AT3G29035 | 3 × 10−34 |
| CaNAC31 | Capana03g000991/Ca03g28100 | chr 03 | - | 0 | 4.38 | 236 | 26.39 | - | - |
| CaNAC32 | Capana03g001014/Ca03g27130 | chr 03 | II(23) | 0 | 4.94 | 245 | 27.96 | - | - |
| CaNAC33 | Capana03g001027/Ca03g27300 | chr 03 | II(23) | 0 | 4.67 | 284 | 32.05 | - | - |
| CaNAC34 | Capana03g001525/Ca03g23090 | chr 03 | I(9) | 1 | 4.77 | 248 | 28.75 | AT3G17730 | 9 × 10−34 |
| CaNAC35 | Capana03g001657/Ca03g22350 | chr 03 | I(3) | 2 | 6.64 | 237 | 27.06 | AT4G17980 | 4 × 10-5 |
| CaNAC36 | Capana03g001780/Ca03g21000 | chr 03 | I(10) | 5 | 4.65 | 632 | 69.87 | AT4G35580 | 0.002 |
| CaNAC37 | Capang03g002541/Ca03g15410 | chr 03 | I(15) | 1 | 9.14 | 170 | 19.75 | AT1G12260 | 5 × 10−19 |
| CaNAC38 | Capana03g002678/Ca02g15520 | chr 03 | II(20) | 2 | 8.46 | 290 | 33.07 | AT1G28470 | 8 × 10−7 |
| CaNAC39 | Capana03g003003/Ca03g14470 | chr 03 | I(12) | 2 | 7.24 | 326 | 37.00 | AT1G79580 | 3 × 10−19 |
| CaNAC40 | Capang03g003315/Ca00g46270 | chr 03 | I(13) | 2 | 6.24 | 290 | 33.35 | AT4G28530 | 3 × 10−6 |
| CaNAC41 | Capana04g000051/Ca04g23340 | chr 04 | - | 2 | 6.71 | 390 | 44.12 | AT5G18270 | 5 × 10−21 |
| CaNAC42 | Capana04g000414 | chr 04 | II(18) | 4 | 4.67 | 350 | 38.80 | - | - |
| CaNAC43 | Capana04g000417/Ca04g20070 | chr 04 | II(18) | 4 | 4.42 | 371 | 40.96 | - | - |
| CaNAC44 | Capana04g000587 | chr 04 | I(13) | 2 | 6.04 | 279 | 31.13 | - | - |
| CaNAC45 | Capana04g001537/Ca00g03050 | chr 04 | I(6) | 2 | 8.33 | 290 | 33.39 | AT1G01720 | 9 × 10−16 |
| CaNAC46 | Capana05g000569/Ca05g04410 | chr 05 | I(6) | 2 | 7.45 | 306 | 34.95 | AT5G08790 | 1 × 10−39 |
| CaNAC47 | Capana05g001291 | chr 05 | - | 0 | 4.26 | 140 | 16.32 | - | - |
| CaNAC48 | Capana05g001292 | chr 05 | - | 2 | 6.16 | 459 | 51.28 | - | - |
| CaNAC49 | Capana05g001365/Ca05g09500 | chr 05 | II(20) | 2 | 6.94 | 486 | 53.97 | AT4G29230 | 3 × 10−47 |
| CaNAC50 | Capana05g001593/Ca11g04580 | chr 05 | I(16) | 2 | 7.13 | 410 | 46.22 | AT1G26870 | 3 × 10−25 |
| CaNAC51 | Capana05g002476/Ca00g86410 | chr 05 | I(8) | 5 | 4.44 | 551 | 61.62 | AT3G10500 | 3 × 10−8 |
| CaNAC52 | Capana05g002477/Ca00g86400 | chr 05 | I(8) | 3 | 5.26 | 396 | 44.64 | AT5G07680 | 4 × 10-12 |
| CaNAC53 | Capana06g000136/Ca06g27940 | chr 06 | I(13) | 2 | 6.14 | 287 | 33.46 | AT4G28530 | 2 × 10−10 |
| CaNAC54 | Capana06g000341/Ca06g25420 | chr 06 | II(26) | 0 | 4.40 | 467 | 53.68 | AT1G34180 | 0.073 |
| CaNAC55 | Capana06g000625/Ca06g22780 | chr 06 | I(11) | 3 | 9.14 | 408 | 45.91 | AT3G44290 | 3 × 10−16 |
| CaNAC56 | Capana06g000752/Ca06g21590 | chr 06 | I(10) | 5 | 4.56 | 637 | 70.12 | AT3G10490 | 0.006 |
| CaNAC57 | Capana06g001075/Ca06g17290 | chr 06 | II(17) | 2 | 5.08 | 379 | 42.94 | AT3G15170 | 0.93 |
| CaNAC58 | Capana06g001093 | chr 06 | II(17) | 1 | 4.54 | 133 | 15.37 | AT3G15170 | 0.08 |
| CaNAC59 | Capana06g001387/Ca06g14050 | chr 06 | I(15) | 2 | 6.24 | 121 | 37.60 | AT4G36160 | 1 × 10−18 |
| CaNAC60 | Capang06g001560/Ca06g13110 | chr 06 | I(9) | 3 | 5.45 | 342 | 38.48 | AT1G32510 | 2 × 10−10 |
| CaNAC61 | Capana06g001739/Ca06g11310 | chr 06 | I(6) | 2 | 6.72 | 294 | 33.67 | AT5G08790 | 3 × 10−12 |
| CaNAC62 | Capana06g002411/Ca06g06410 | chr 06 | I(15) | 2 | 6.63 | 347 | 40.20 | AT1G12260 | 8 × 10−29 |
| CaNAC63 | Capana06g002441/Ca00g77790 | chr 06 | I(9) | 3 | 5.24 | 334 | 37.91 | AT5G63790 | 1 × 10−5 |
| CaNAC64 | Capana06g002936 | chr 06 | - | 1 | 9.83 | 136 | 15.88 | AT3G10490 | 0.005 |
| CaNAC65 | Capana06g003012/Ca06g01680 | chr 06 | I(13) | 2 | 6.24 | 290 | 33.39 | AT4G28530 | 3 × 10−12 |
| CaNAC66 | Capana07g000058/Ca07g00500 | chr 07 | II(17) | 2 | 5.42 | 415 | 47.22 | AT3G15500 | 4 × 10−9 |
| CaNAC67 | Capana07g000215/Ca07g02120 | chr 07 | II(22) | 0 | 4.42 | 196 | 22.85 | AT1G34180 | 5 × 10−4 |
| CaNAC68 | Capana07g001015 | chr 07 | - | 1 | 10.03 | 238 | 26.98 | AT5G39820 | 1 × 10−8 |
| CaNAC69 | Capana07g001769/Ca00g84200 | chr 07 | II(20) | 3 | 7.65 | 325 | 36.58 | AT4G28500 | 1 × 10−21 |
| CaNAC70 | Capana07g002072/Ca07g16790 | chr 07 | II(25) | 0 | 9.70 | 235 | 27.48 | AT1G34190 | 1.5 |
| CaNAC71 | Capana07g002159/Ca07g17460 | chr 07 | I(14) | 1 | 8.85 | 323 | 36.23 | AT5G53950 | 5 × 10−70 |
| CaNAC72 | Capana07g002219/Ca07g18020 | chr 07 | I(5) | 2 | 7.44 | 338 | 37.67 | AT3G15500 | 5 × 10−27 |
| CaNAC73 | Capana07g002220/Ca07g02180 | chr 07 | I(7) | 8 | 8.70 | 340 | 38.00 | - | - |
| CaNAC74 | Capana07g002471/Ca07g21320 | chr 07 | I(16) | 2 | 7.54 | 323 | 37.20 | AT2G43000 | 3 × 10−9 |
| CaNAC75 | Capana07g002485/Ca07g21470 | chr 07 | I(13) | 2 | 8.61 | 136 | 15.53 | AT1G56010 | 3 × 10−18 |
| CaNAC76 | Capana08g000884/Ca08g06780 | chr 08 | II(24) | 0 | 7.30 | 232 | 26.86 | - | - |
| CaNAC77 | Capana08g000885/Ca08g06790 | chr 08 | II(24) | 0 | 6.79 | 178 | 20.64 | - | - |
| CaNAC78 | Capana08g001727/Ca08g13660 | chr 08 | I(2) | 1 | 7.16 | 408 | 46.32 | AT2G02450 | 3 × 10−75 |
| CaNAC79 | Capana08g001999/Ca08g15460 | chr 08 | II(21) | 2 | 4.48 | 195 | 22.63 | AT5G64530 | 5 × 10−10 |
| CaNAC80 | Capana09g000105/Ca09g18600 | chr 09 | II(21) | 1 | 4.92 | 314 | 36.51 | - | - |
| CaNAC81 | Capana09g000936/Ca09g12970 | chr 09 | I(5) | 2 | 9.45 | 350 | 39.62 | AT4G27410 | 9 × 10−44 |
| CaNAC82 | Capana09g002022/Ca03g21000 | chr 09 | - | 0 | 10.32 | 103 | 12.00 | AT4G35580 | 3 × 10−4 |
| CaNAC83 | Capana09g002444/Ca09g00160 | chr 09 | I(1) | 1 | 9.83 | 161 | 18.66 | AT5G39610 | 0.002 |
| CaNAC84 | Capana10g000063/Ca10g00460 | chr 10 | I(7) | 4 | 8.41 | 342 | 39.05 | AT1G61110 | 2 × 10−1 |
| CaNAC85 | Capana10g000683/Ca11g08290 | chr 10 | I(12) | 2 | 6.46 | 341 | 39.09 | AT2G46770 | 2 × 10−51 |
| CaNAC86 | Capana10g001138/Ca10g09910 | chr 10 | I(2) | 1 | 6.68 | 299 | 34.40 | AT3G12977 | 0.047 |
| CaNAC87 | Capana10g002356/Ca10g20640 | chr 10 | II(26) | 0 | 4.17 | 578 | 65.18 | AT2G43000 | 0.023 |
| CaNAC88 | Capana10g002363/Ca10g20640 | chr 10 | II(26) | 1 | 5.72 | 421 | 48.33 | AT2G43000 | 0.26 |
| CaNAC89 | Capana11g000328 | chr 11 | I(1) | 2 | 8.56 | 256 | 29.67 | AT3G44350 | 7 × 10−13 |
| CaNAC90 | Capana11g000346/Ca11g16490 | chr 11 | I(11) | 3 | 8.94 | 414 | 46.60 | AT3G44290 | 8 × 10−14 |
| CaNAC91 | Capana11g000795/Ca11g12260 | chr 11 | II(17) | 2 | 6.51 | 344 | 40.34 | AT1G34180 | 2× 10−5 |
| CaNAC92 | Capana11g001813/Ca11g04440 | chr 11 | I(4) | 2 | 7.57 | 294 | 33.85 | AT1G69490 | 2 × 10−26 |
| CaNAC93 | Capana11g001932/Ca10g13700 | chr 11 | II(24) | 0 | 10.07 | 167 | 19.90 | - | - |
| CaNAC94 | Capana11g001935/Ca10g13720 | chr 11 | II(24) | 0 | 6.68 | 218 | 25.63 | - | - |
| CaNAC95 | Capana11g002231/Ca11g01550 | chr 11 | II(19) | 3 | 5.12 | 481 | 54.73 | AT1G25580 | 1 × 10−15 |
| CaNAC96 | Capana12g002058/Ca12g07920 | chr 12 | - | 3 | 4.44 | 719 | 79.51 | AT1G33280 | 0.007 |
| CaNAC97 | Capana12g002348/Ca12g05890 | chr 12 | II(22) | 0 | 10.15 | 154 | 18.04 | - | - |
| CaNAC98 | Capana12g002352/Ca12g05910 | chr 12 | II(22) | 0 | 4.75 | 269 | 31.41 | AT5G62380 | 0.011 |
| CaNAC99 | Capana12g002357/Ca12g06210 | chr 12 | II(22) | 0 | 9.35 | 224 | 26.75 | AT1G34180 | 0.14 |
| CaNAC100 | Capana12g002360/Ca12g05910 | chr 12 | II(22) | 0 | 4.63 | 268 | 31.17 | AT5G62380 | 0.011 |
| CaNAC101 | Capana12g002456/Ca12g04970 | chr 12 | I(8) | 5 | 4.35 | 476 | 52.88 | AT3G10500 | 1 × 10−9 |
| CaNAC102 | Capana12g002457/Ca12g04950 | chr 12 | I(8) | 3 | 5.17 | 426 | 47.63 | AT1G34180 | 1 × 10−9 |
| CaNAC103 | Capana12g002917/Ca12g00080 | chr 12 | II(21) | 2 | 4.27 | 195 | 22.09 | - | - |
| CaNAC104 | Capana12g002918/Ca12g00080 | chr 12 | II(21) | 2 | 4.27 | 195 | 22.96 | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diao, W.; Snyder, J.C.; Wang, S.; Liu, J.; Pan, B.; Guo, G.; Ge, W.; Dawood, M.H.S.A. Genome-Wide Analyses of the NAC Transcription Factor Gene Family in Pepper (Capsicum annuum L.): Chromosome Location, Phylogeny, Structure, Expression Patterns, Cis-Elements in the Promoter, and Interaction Network. Int. J. Mol. Sci. 2018, 19, 1028. https://doi.org/10.3390/ijms19041028
Diao W, Snyder JC, Wang S, Liu J, Pan B, Guo G, Ge W, Dawood MHSA. Genome-Wide Analyses of the NAC Transcription Factor Gene Family in Pepper (Capsicum annuum L.): Chromosome Location, Phylogeny, Structure, Expression Patterns, Cis-Elements in the Promoter, and Interaction Network. International Journal of Molecular Sciences. 2018; 19(4):1028. https://doi.org/10.3390/ijms19041028
Chicago/Turabian StyleDiao, Weiping, John C. Snyder, Shubin Wang, Jinbing Liu, Baogui Pan, Guangjun Guo, Wei Ge, and Mohammad Hasan Salman Ali Dawood. 2018. "Genome-Wide Analyses of the NAC Transcription Factor Gene Family in Pepper (Capsicum annuum L.): Chromosome Location, Phylogeny, Structure, Expression Patterns, Cis-Elements in the Promoter, and Interaction Network" International Journal of Molecular Sciences 19, no. 4: 1028. https://doi.org/10.3390/ijms19041028