
Acteoside and Isoacteoside Protect Amyloid β Peptide Induced Cytotoxicity, Cognitive Deficit and Neurochemical Disturbances In Vitro and In Vivo


Abstract
:
1. Introduction
2. Results
2.1. In Vivo Aβ 1-42-Infusion Model
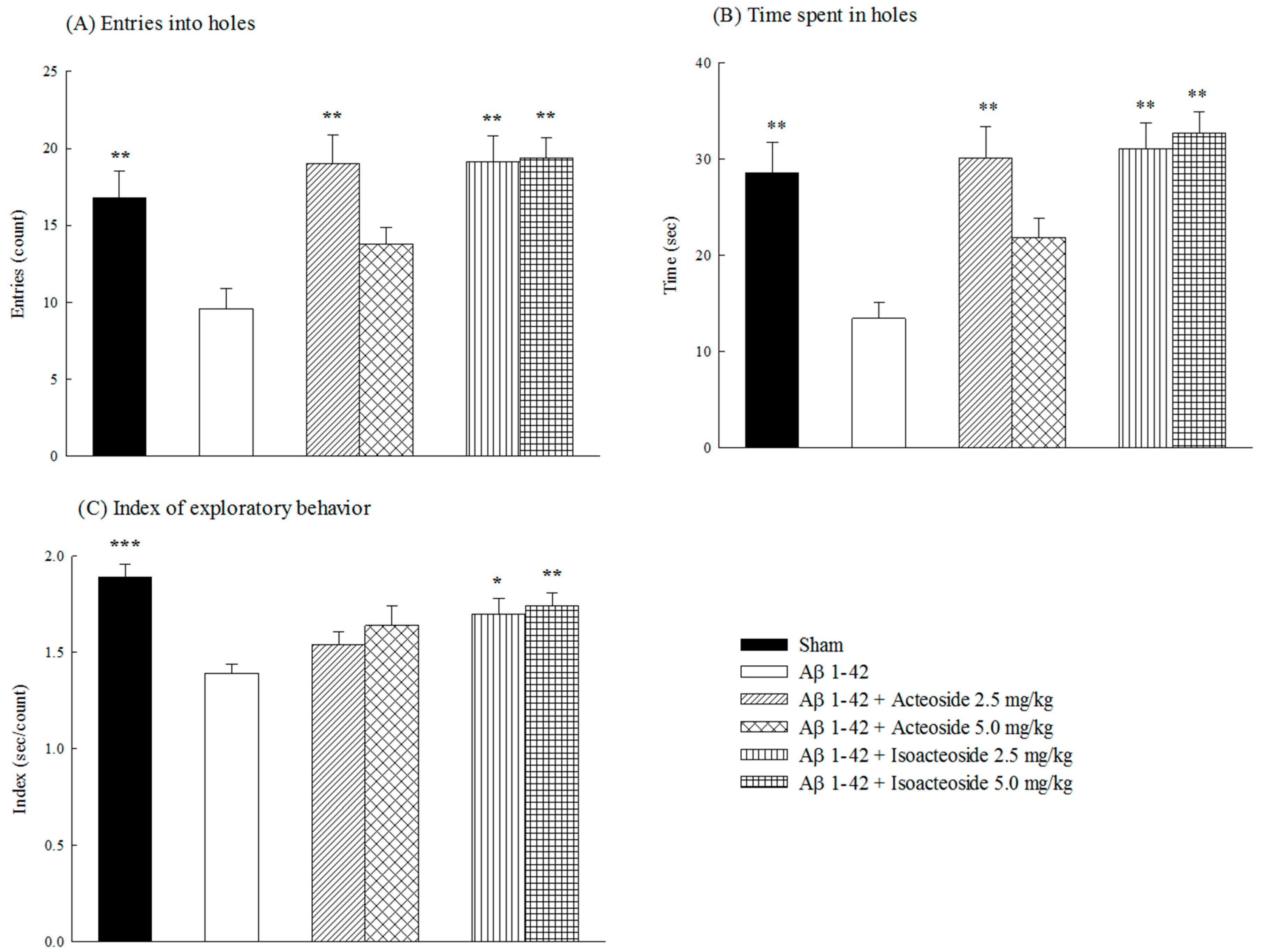
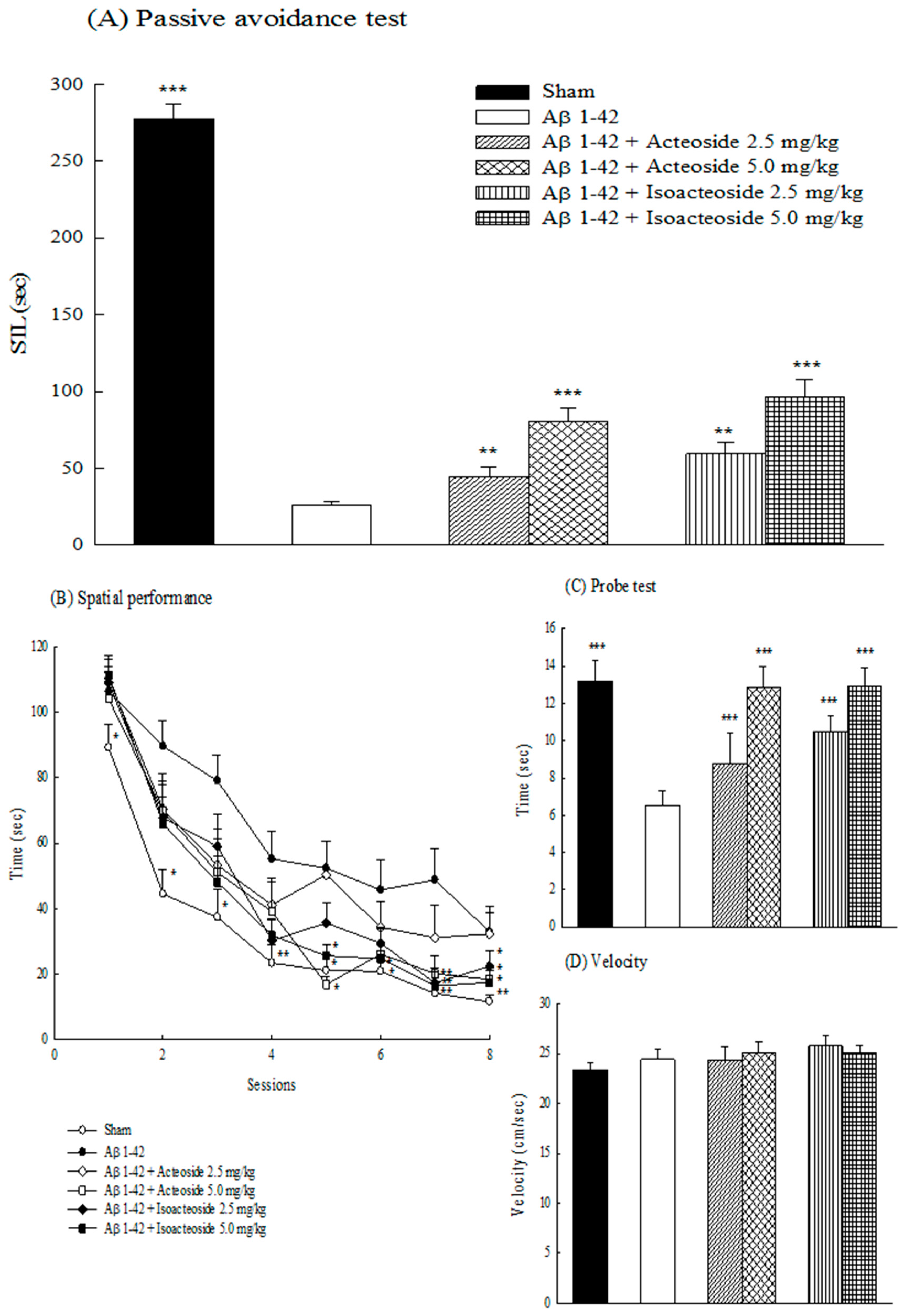
2.1.1. Effects of Acteoside and Isoacteoside on Behavioral Dysfunction Induced by Aβ 1-42 in Rats
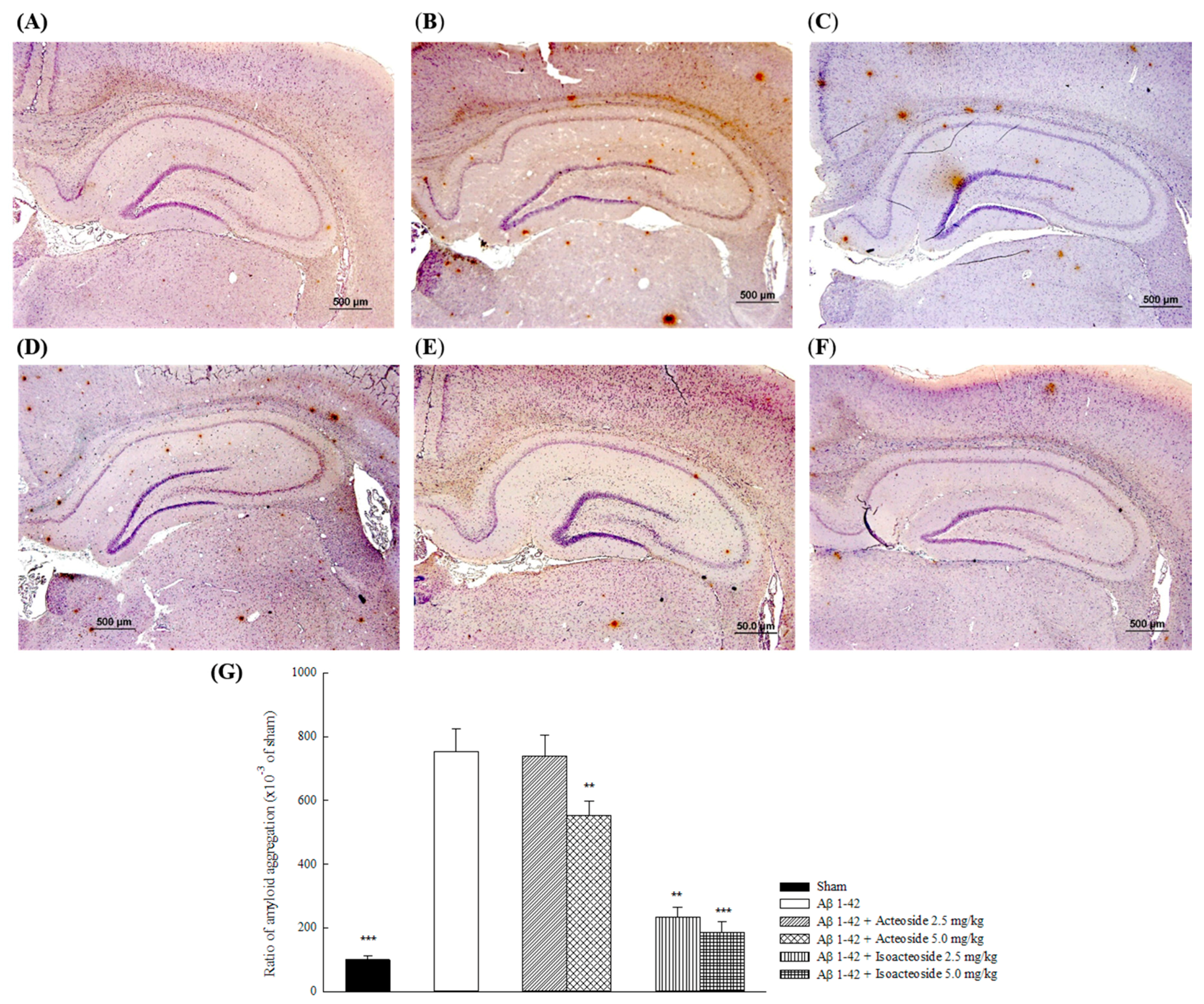
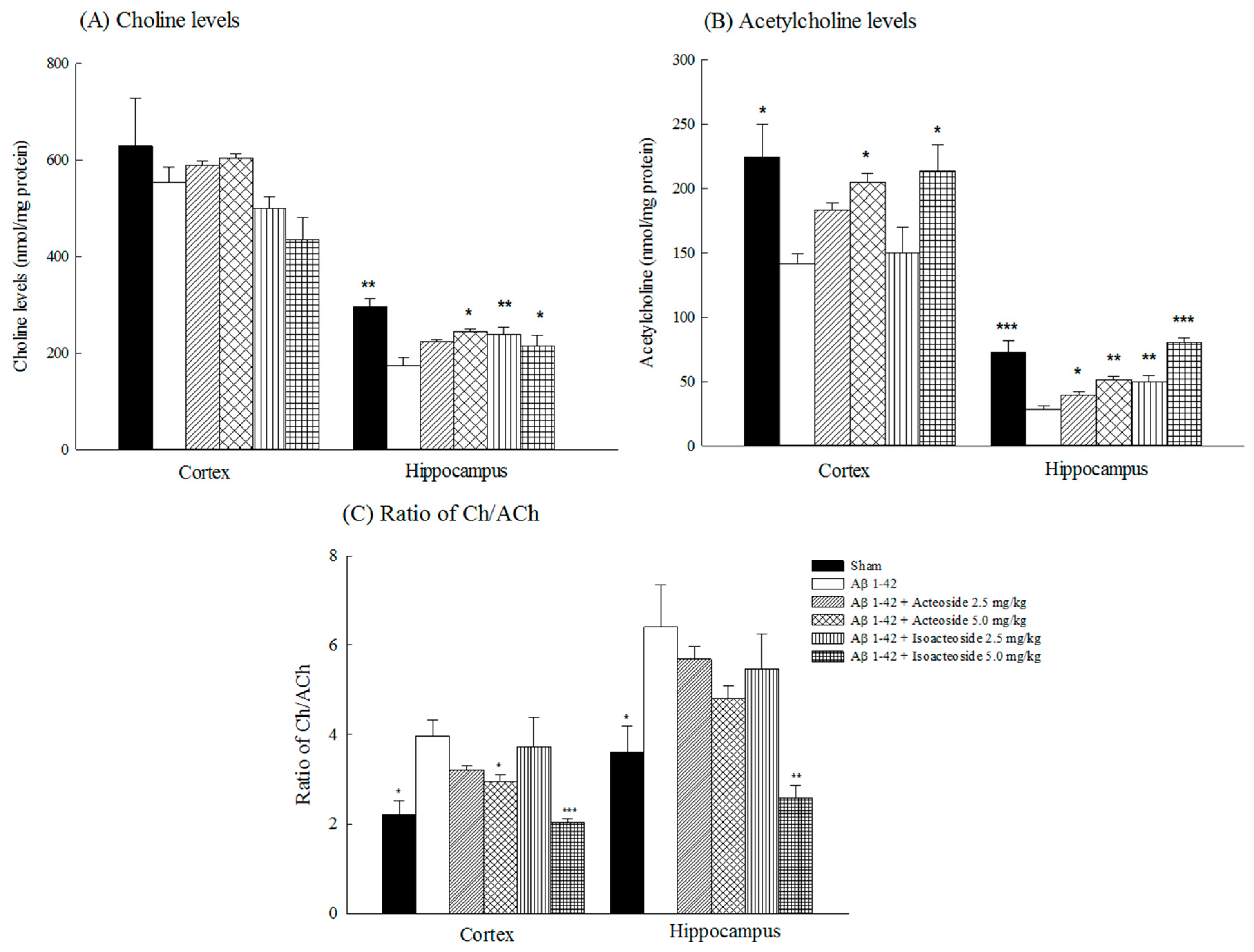
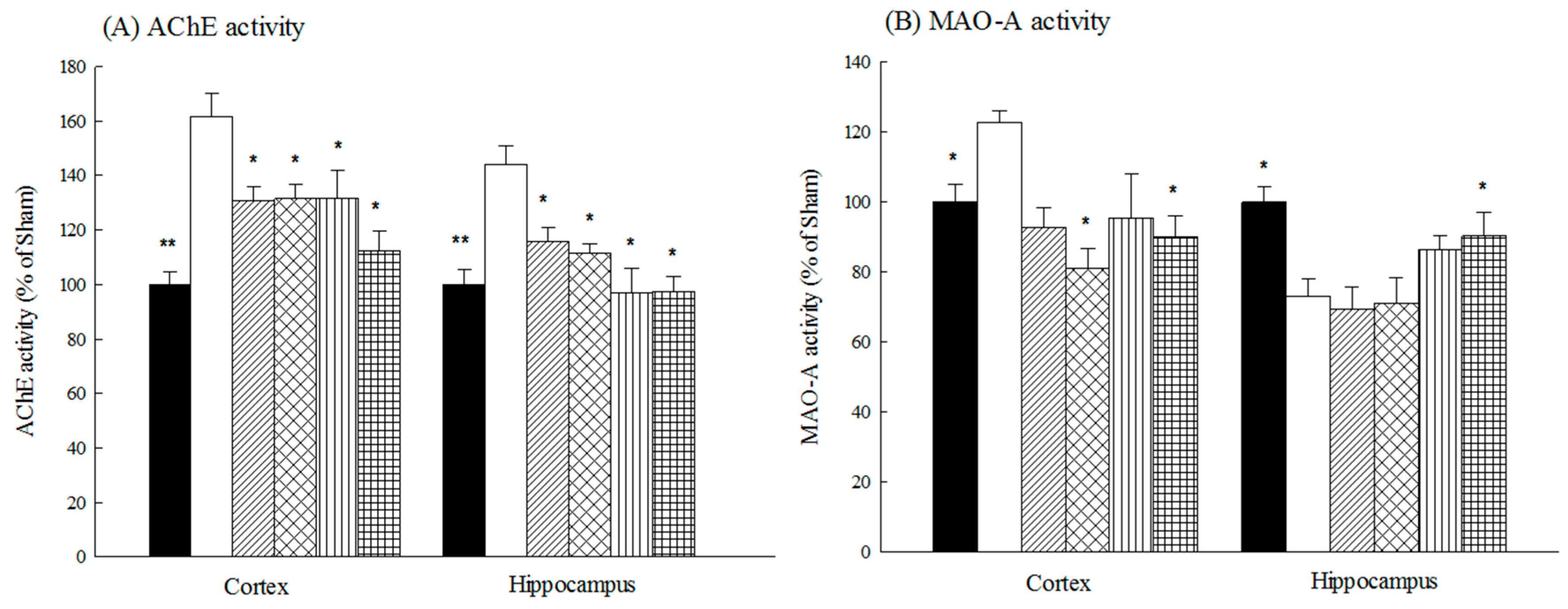
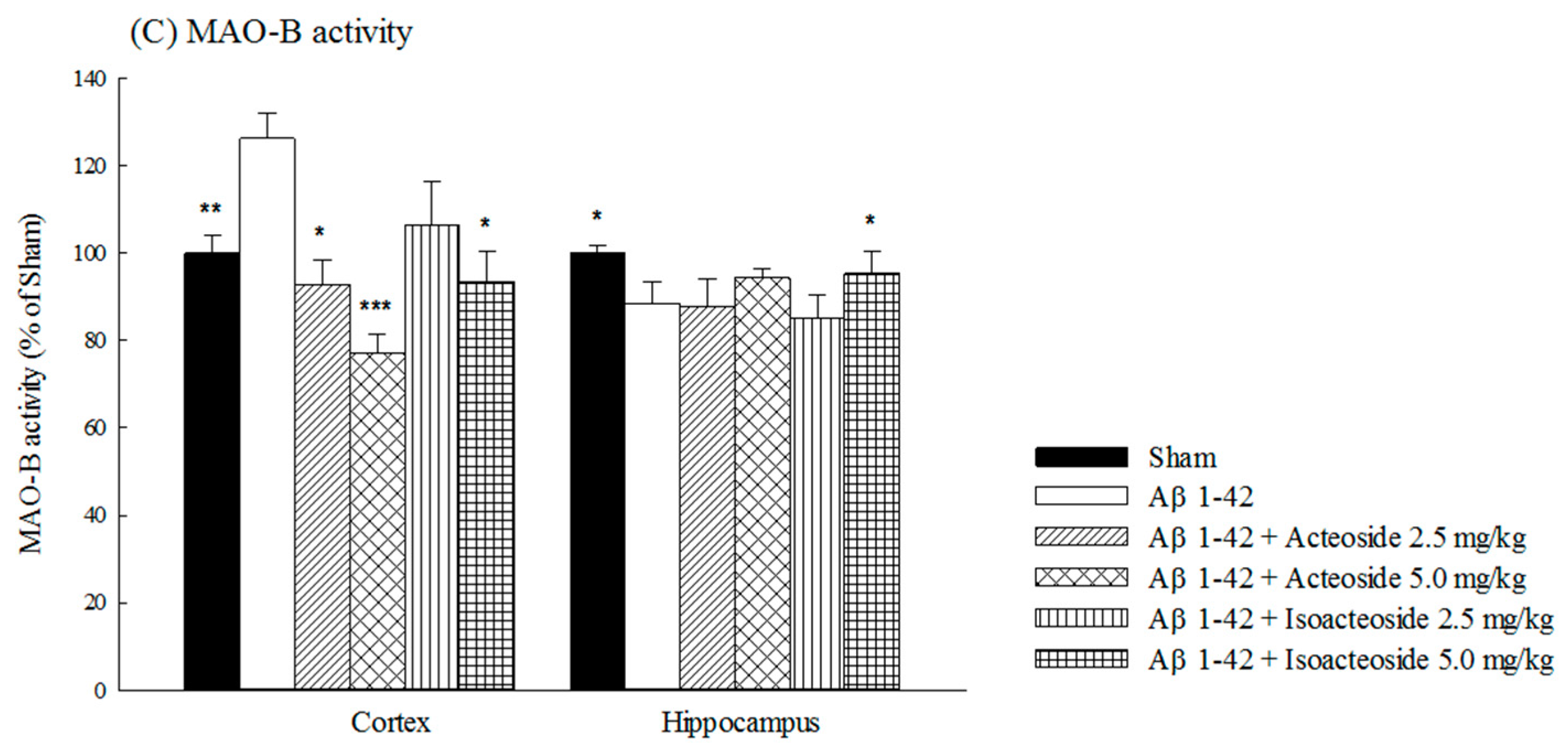
2.1.2. Effects of Acteoside and Isoacteoside on Amyloid Deposition and Neurochemical Disturbances Induced by Aβ 1-42 in Rats
2.2. In Vitro Test on Amyloid Cacade
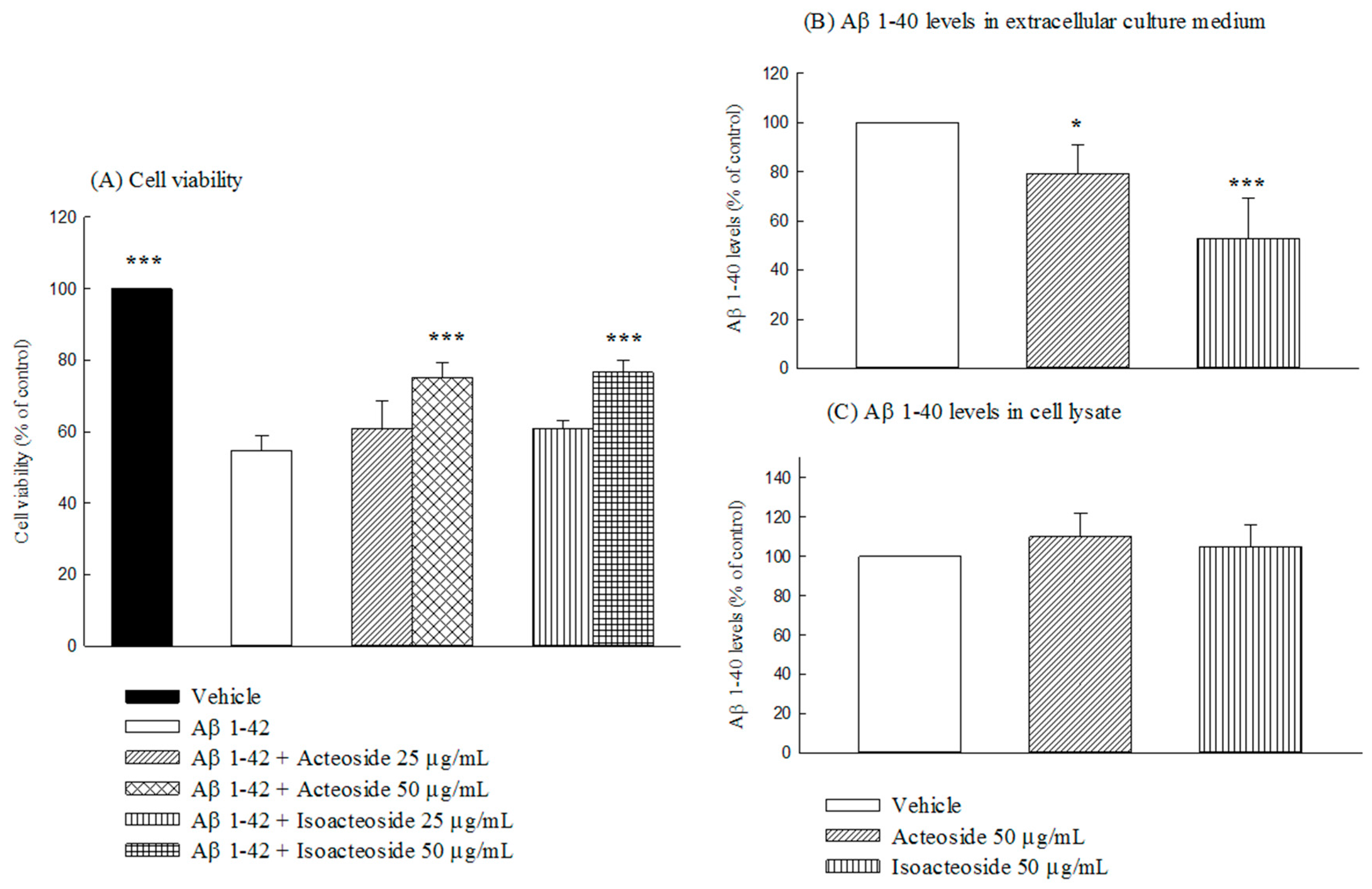
2.2.1. Effects of Acteoside and Isoacteoside on Neuronal Damage Induced by Aβ 1-42, and Intracellular and Extracellular Aβ 1-40 Levels in SH-SY5Y Cells
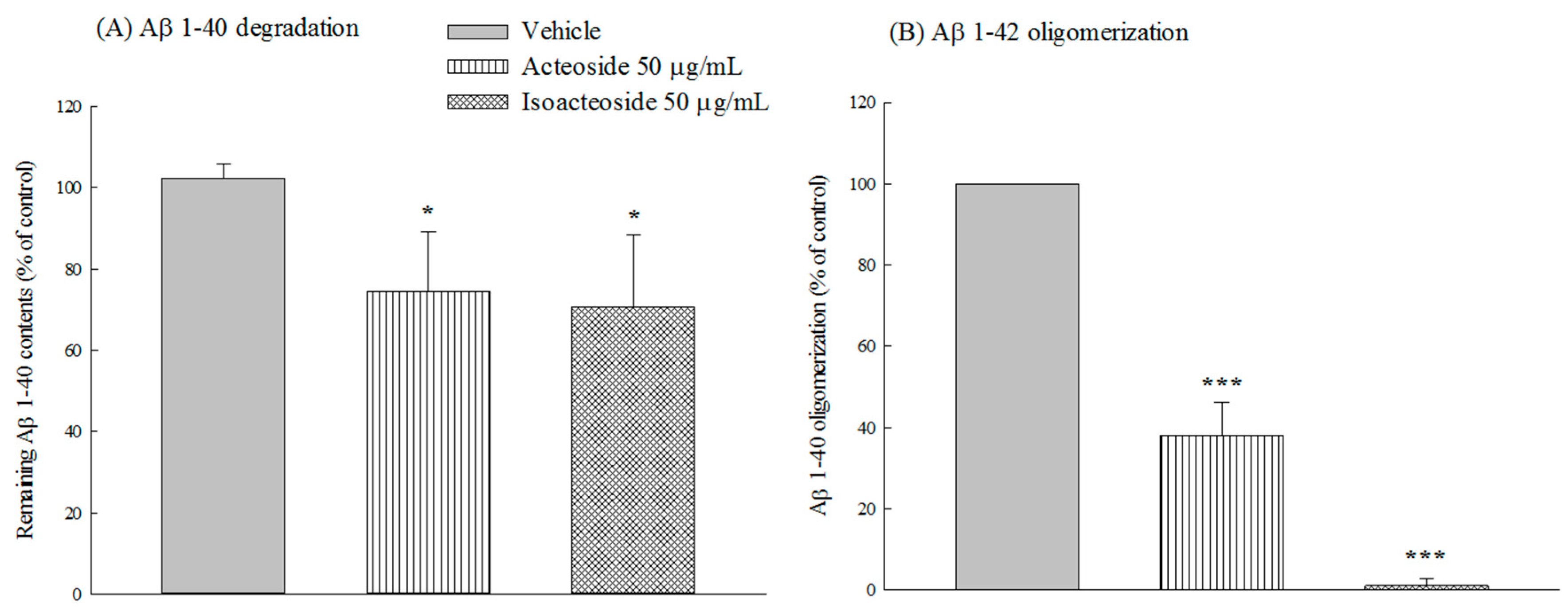
2.2.2. Effects of Acteoside and Isoacteoside on Aβ 1-40 Degradation and Aβ 1-42 Oligomerization In Vitro
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. In Vivo Aβ 1-42-Infused Model
4.3.1. Schedule of Aβ 1-42-Infused Model
4.3.2. Behavioral Tests
4.3.3. Assessment of Aβ 1-42 Deposition, Neurotransmitter Levels, and AChE and MAO Activity in Brain
4.4. In Vitro Test on Amyloid Cacade
4.4.1. Assessment of Cytoprotective Effect in SH-SY5Y Cells
4.4.2. Assessment of Intracellular and Extracellular Aβ 1-40 Levels in SH-SY5Y Cells
4.4.3. Assessment of Cell-Free Aβ 1-40 Degradation In Vitro
4.4.4. Assessment of Aβ 1-42 Oligomerization In Vitro
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Aβ | Amyloid β peptide |
| Ach | Acetylcholine |
| AChE | Acetylcholinesterase |
| AD | Alzheimer’s disease |
| APP | Amyloid precursor protein |
| BF | Basal forebrain |
| Ch | Choline |
| DA | Dopamine |
| MAO | Monoamine oxidase |
| MTT | 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide |
| MWM | Morris water maze |
| NE | Norepinephrine |
| PBS | Phosphate buffer saline |
| ThT | Thioflavin T |
References
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Karran, E.; Mercken, M.; de Strooper, B. The amyloid cascade hypothesis for Alzheimer’s disease: An appraisal for the development of therapeutics. Nat. Rev. Drug Discov. 2011, 10, 698–712. [Google Scholar] [CrossRef] [PubMed]
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef] [PubMed]
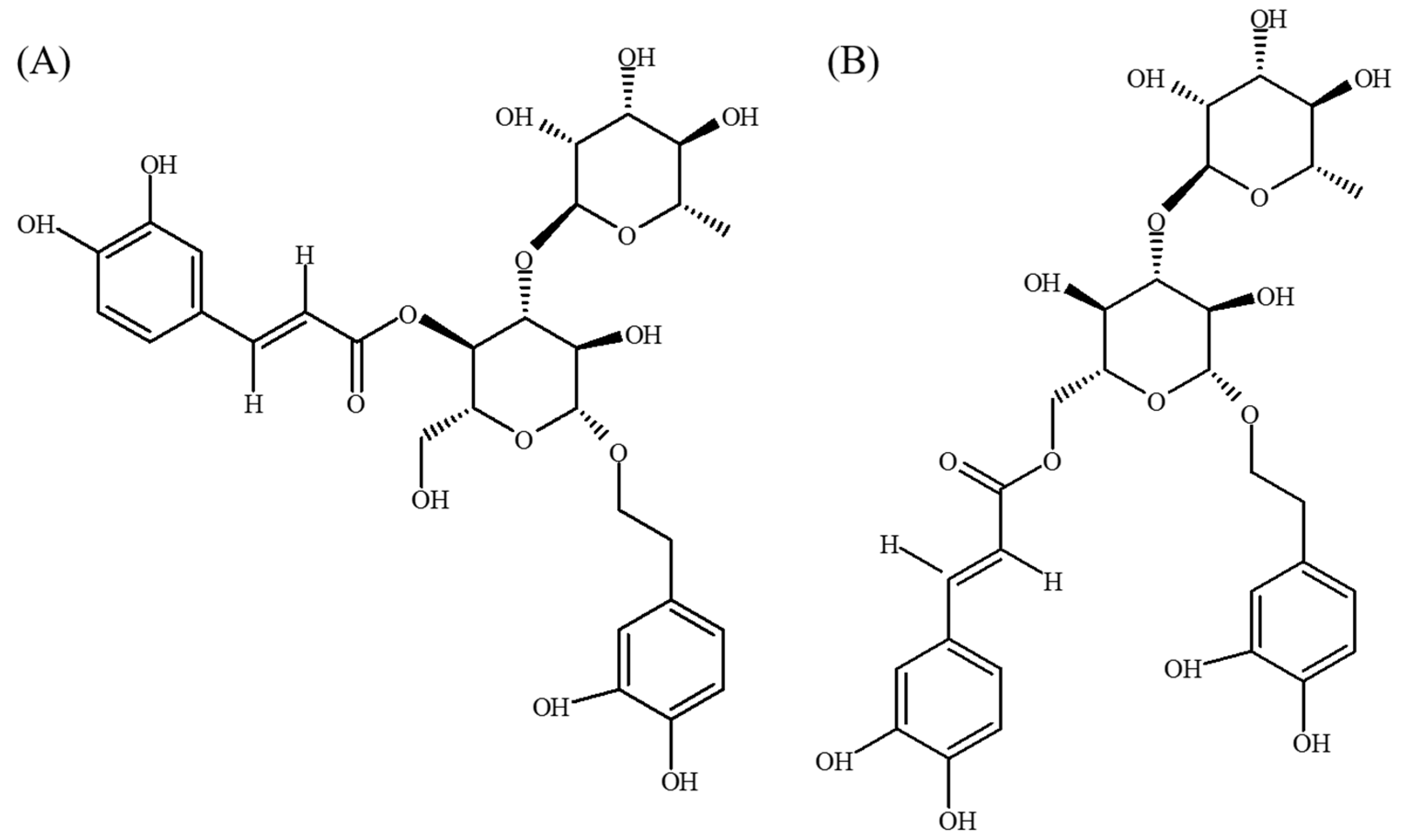
- Li, L.; Tsao, R.; Liu, Z.; Liu, S.; Yang, R.; Young, J.C.; Zhu, H.; Deng, Z.; Xie, M.; Fu, Z. Isolation and purification of acteoside and isoacteoside from Plantago psyllium L. by high-speed counter-current chromatography. J. Chromatogr. A 2005, 1063, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Numata, A.; Takemura, T.; Ode, R.H.; Narula, A.S.; Schmidt, J.M.; Cragg, G.M.; Pase, C.P. Antineoplastic agents, 107. Isolation of acteoside and isoacteoside from Castilleja linariaefolia. J. Nat. Prod. 1990, 53, 456–458. [Google Scholar] [CrossRef] [PubMed]
- Ohno, T.; Inoue, M.; Ogihara, Y.; Saracoglu, I. Antimetastatic activity of acteoside, a phenylethanoid glycoside. Biol. Pharm Bull. 2002, 25, 666–668. [Google Scholar] [CrossRef] [PubMed]
- Chiou, W.F.; Lin, L.C.; Chen, C.F. Acteoside protects endothelial cells against free radical-induced oxidative stress. J. Pharm Pharmacol. 2004, 56, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Woo, E.R.; Choi, C.Y.; Shin, D.W.; Lee, D.G.; You, H.J.; Jeong, H.G. Protective effect of acteoside on carbon tetrachloride-induced hepatotoxicity. Life Sci. 2004, 74, 1051–1064. [Google Scholar] [CrossRef] [PubMed]
- Koo, K.A.; Kim, S.H.; Oh, T.H.; Kim, Y.C. Acteoside and its aglycones protect primary cultures of rat cortical cells from glutamate-induced excitotoxicity. Life Sci. 2006, 79, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, Y.; Yan, J.; Zhao, X.; Sun, X.; Zhang, Y.; Guo, J.; Zhu, C. Acteoside protects human neuroblastoma SH-SY5Y cells against β-amyloid-induced cell injury. Brain Res. 2009, 1283, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Yamada, P.; Iijima, R.; Han, J.; Shigemori, H.; Yokota, S.; Isoda, H. Inhibitory effect of acteoside isolated from Cistanche tubulosa on chemical mediator release and inflammatory cytokine production by RBL-2H3 and KU812 cells. Planta Med. 2010, 76, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Chunhua, M.; Shumin, W. Effects of acteoside on lipopolysaccharide-induced inflammation in acute lung injury via regulation of NF-κB pathway in vivo and in vitro. Toxicol. Appl. Pharmacol. 2015, 285, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Gao, L.; Huo, S.X.; Peng, X.M.; Wu, P.P.; Cai, L.M.; Yan, M. Effect of acteoside on learning and memory impairment induced by scopolamine in mice. China J. Chin. Mater. Med. 2012, 37, 2956–2959. [Google Scholar]
- Xiong, L.; Mao, S.; Lu, B.; Yang, J.; Zhou, F.; Hu, Y.; Jiang, Y.; Shen, C.; Zhao, Y. Osmanthus fragrans flower extract and acteoside protect against d-galactose-induced aging in an ICR mouse model. J. Med. Food 2016, 19, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Peng, X.; Huo, S.; He, Y.; Yan, M. Acteoside enhances expression of neurotrophin-3 in brain tissues of subacute aging mice induced by d-galactose combined with aluminum trichloride. Chin. J. Cell. Mol. Immunol. 2014, 30, 1022–1025. [Google Scholar]
- Peng, X.M.; Gao, L.; Huo, S.X.; Liu, X.M.; Yan, M. The mechanism of memory enhancement of acteoside (Verbascoside) in the senescent mouse model induced by a combination of d-gal and AlCl3. Phytother. Res. 2015, 29, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Peng, X.M.; Huo, S.X.; Liu, X.M.; Yan, M. Memory enhancement of acteoside (Verbascoside) in a senescent mice model induced by a combination of d-gal and AlCl3. Phytother. Res. 2015, 29, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Q.; Xu, Y.X.; Zhu, C.Q. Upregulation of heme oxygenase-1 by acteoside through ERK and PI3 K/Akt pathway confer neuroprotection against β-amyloid-induced neurotoxicity. Neurotox. Res. 2012, 21, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Kurisu, M.; Miyamae, Y.; Murakami, K.; Han, J.; Isoda, H.; Irie, K.; Shigemori, H. Inhibition of amyloid β aggregation by acteoside, a phenylethanoid glycoside. Biosci. Biotechnol. Biochem. 2013, 77, 1329–1332. [Google Scholar] [CrossRef] [PubMed]
- Myhrer, T. Neurotransmitter systems involved in learning and memory in the rat: A meta-analysis based on studies of four behavioral tasks. Brain Res. Rev. 2003, 41, 268–287. [Google Scholar] [CrossRef]
- Itoh, A.; Nitta, A.; Nadai, M.; Nishimura, K.; Hirose, M.; Hasegawa, T.; Nabeshima, T. Dysfunction of cholinergic and dopaminergic neuronal systems in β-amyloid protein—Infused rats. J. Neurochem. 1996, 66, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Tran, M.H.; Yamada, K.; Nabeshima, T. Amyloid β-peptide induces cholinergic dysfunction and cognitive deficits: A minireview. Peptides 2002, 23, 1271–1283. [Google Scholar] [CrossRef]
- Yamada, K.; Tanaka, T.; Mamiya, T.; Shiotani, T.; Kameyama, T.; Nabeshima, T. Improvement by nefiracetam of β-amyloid-(1-42)-induced learning and memory impairments in rats. Br. J. Pharmacol. 1999, 126, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.R.; Lin, H.C.; Su, M.H. Reversal by aqueous extracts of Cistanche tubulosa from behavioral deficits in Alzheimer’s disease-like rat model: Relevance for amyloid deposition and central neurotransmitter function. BMC Complement. Altern. Med. 2014, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Jeong, E.J.; Lee, H.S.; Kim, Y.C. Acteoside of Callicarpa dichotoma attenuates scopolamine-induced memory impairments. Biol. Pharm Bull. 2006, 29, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Arai, H.; Ichimiya, Y.; Kosaka, K.; Moroji, T.; Iizuka, R. Neurotransmitter changes in early- and late-onset Alzheimer-type dementia. Prog. Neuropsychopharmacol. Biol. Psychiatry 1992, 16, 883–890. [Google Scholar] [PubMed]
- Gulyas, B.; Pavlova, E.; Kasa, P.; Gulya, K.; Bakota, L.; Varszegi, S.; Keller, E.; Horvath, M.C.; Nag, S.; Hermecz, I.; et al. Activated MAO-B in the brain of Alzheimer patients, demonstrated by 11C-l-deprenyl using whole hemisphere autoradiography. Neurochem. Int. 2011, 58, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Parihar, M.S.; Hemnani, T. Alzheimer’s disease pathogenesis and therapeutic interventions. J. Clin. Neurosci. 2004, 11, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Baik, S.H.; Kang, S.; Cho, S.W.; Bae, J.; Cha, M.Y.; Sailor, M.J.; Mook-Jung, I.; Ahn, K.H. Close correlation of monoamine oxidase activity with progress of Alzheimer’s disease in mice, observed by in vivo two-photon imaging. ACS Cent. Sci. 2016, 2, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Kawamata, T.; Akiguchi, I.; Kameyama, M.; Nakamura, N.; Kimura, H. Expression of monoamine oxidase B activity in astrocytes of senile plaques. Acta Neuropathol. 1990, 80, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Smale, G.; Nichols, N.R.; Brady, D.R.; Finch, C.E.; Horton, W.E., Jr. Evidence for apoptotic cell death in Alzheimer’s disease. Exp. Neurol. 1995, 133, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Blanco, N.J.; Love, B.C.; Cooper, J.A.; McGeary, J.E.; Knopik, V.S.; Maddox, W.T. A frontal dopamine system for reflective exploratory behavior. Neurobiol. Learn. Mem. 2015, 123, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Young, J.W.; Kooistra, K.; Geyer, M.A. Dopamine receptor mediation of the exploratory/hyperactivity effects of modafinil. Neuropsychopharmacology 2011, 36, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Alttoa, A.; Seeman, P.; Koiv, K.; Eller, M.; Harro, J. Rats with persistently high exploratory activity have both higher extracellular dopamine levels and higher proportion of D2 (High) receptors in the striatum. Synapse 2009, 63, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Bai, P.; Peng, X.M.; Gao, L.; Huo, S.X.; Zhao, P.P.; Yan, M. Study on protective effect of acteoside on cellular model of Alzheimer’s disease induced by okadaic acid. China J. Chin. Mater. Med. 2013, 38, 1323–1326. [Google Scholar]
- Pu, X.; Song, Z.; Li, Y.; Tu, P.; Li, H. Acteoside from Cistanche salsa inhibits apoptosis by 1-methyl-4-phenylpyridinium ion in cerebellar granule neurons. Planta Med. 2003, 69, 65–66. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D. The neurotrophin family of neurotrophic factors: An overview. Methods Mol. Biol. 2012, 846, 1–12. [Google Scholar] [PubMed]
- Korsching, S.; Auburger, G.; Heumann, R.; Scott, J.; Thoenen, H. Levels of nerve growth factor and its mRNA in the central nervous system of the rat correlate with cholinergic innervation. EMBO J. 1985, 4, 1389–1393. [Google Scholar] [PubMed]
- Auld, D.S.; Mennicken, F.; Quirion, R. Nerve growth factor rapidly induces prolonged acetylcholine release from cultured basal forebrain neurons: Differentiation between neuromodulatory and neurotrophic influences. J. Neurosci. 2001, 21, 3375–3382. [Google Scholar] [PubMed]
- Mufson, E.J.; Counts, S.E.; Perez, S.E.; Ginsberg, S.D. Cholinergic system during the progression of Alzheimer’s disease: Therapeutic implications. Expert Rev. Neurother. 2008, 8, 1703–1718. [Google Scholar] [CrossRef] [PubMed]
- Glowinski, J.; Iversen, L.L. Regional studies of catecholamines in the rat brain. I. The disposition of [3H]norepinephrine, [3H]dopamine and [3H]dopa in various regions of the brain. J. Neurochem. 1966, 13, 655–669. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Levels of Cortical Neurotransmitters and Their Metabolites (ng/g Protein) | |||||
| - | MHPG | NE | DOPAC | HVA | DA |
| Vehicle | 26.05 ± 1.11 * | 15.63 ± 0.41 | 17.13 ± 0.85 * | 3.76 ± 0.78 ** | 3.83 ± 0.16 ** |
| Aβ 1-42 | 22.87 ± 0.81 | 13.61 ± 1.20 | 11.97 ± 0.66 | 2.90 ± 0.23 | 2.18 ± 0.11 |
| Acteoside | |||||
| 2.5 mg/kg | 22.40 ± 1.24 | 13.08 ± 1.06 | 12.91 ± 0.66 | 2.87 ± 0.11 | 2.26 ± 0.06 |
| 5.0 mg/kg | 23.84 ± 1.94 | 12.91 ± 0.67 | 13.25 ± 1.07 | 2.90 ± 0.15 | 2.23 ± 0.15 |
| Isoacteoside | |||||
| 2.5 mg/kg | 31.71 ± 3.21 * | 12.60 ± 1.56 | 12.53 ± 3.33 | 2.91 ± 0.58 | 2.67 ± 0.43 |
| 5.0 mg/kg | 34.41 ± 4.81 * | 14.18 ± 1.01 | 13.80 ± 3.03 | 2.74 ± 0.69 | 3.84 ± 0.47 * |
| The Levels of Hippocampal Neurotransmitters and Their Metabolites (ng/g Protein) | |||||
| MHPG | NE | DOPAC | HVA | DA | |
| Vehicle | 622.30 ± 17.58 | 64.49 ± 1.65 ** | 5.90 ± 0.39 | 3.60 ± 0.20 | 5.41 ± 0.35 *** |
| Aβ 1-42 | 620.97 ± 25.79 | 51.96 ± 1.54 | 5.12 ± 0.36 | 3.24 ± 0.16 | 0.97 ± 0.07 |
| Acteoside | |||||
| 2.5 mg/kg | 641.15 ± 23.10 | 53.16 ± 2.29 | 5.22 ± 0.27 | 3.26 ± 0.12 | 1.07 ± 0.15 |
| 5.0 mg/kg | 631.13 ± 36.81 | 53.53 ± 1.83 | 5.32 ± 0.14 | 3.34 ± 0.10 | 1.17 ± 0.13 |
| Isoacteoside | |||||
| 2.5 mg/kg | 689.99 ± 79.95 | 54.01 ± 6.20 | 4.98 ± 1.15 | 3.68 ± 0.46 | 5.28 ± 0.83 *** |
| 5.0 mg/kg | 574.02 ± 54.16 | 45.13 ± 3.36 | 4.13 ± 0.45 | 3.29 ± 0.25 | 5.66 ± 0.36 *** |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiao, Y.-J.; Su, M.-H.; Lin, H.-C.; Wu, C.-R. Acteoside and Isoacteoside Protect Amyloid β Peptide Induced Cytotoxicity, Cognitive Deficit and Neurochemical Disturbances In Vitro and In Vivo. Int. J. Mol. Sci. 2017, 18, 895. https://doi.org/10.3390/ijms18040895
Shiao Y-J, Su M-H, Lin H-C, Wu C-R. Acteoside and Isoacteoside Protect Amyloid β Peptide Induced Cytotoxicity, Cognitive Deficit and Neurochemical Disturbances In Vitro and In Vivo. International Journal of Molecular Sciences. 2017; 18(4):895. https://doi.org/10.3390/ijms18040895
Chicago/Turabian StyleShiao, Young-Ji, Muh-Hwan Su, Hang-Ching Lin, and Chi-Rei Wu. 2017. "Acteoside and Isoacteoside Protect Amyloid β Peptide Induced Cytotoxicity, Cognitive Deficit and Neurochemical Disturbances In Vitro and In Vivo" International Journal of Molecular Sciences 18, no. 4: 895. https://doi.org/10.3390/ijms18040895