
Novel Hybrid Peptide Cecropin A (1–8)-LL37 (17–30) with Potential Antibacterial Activity

Abstract
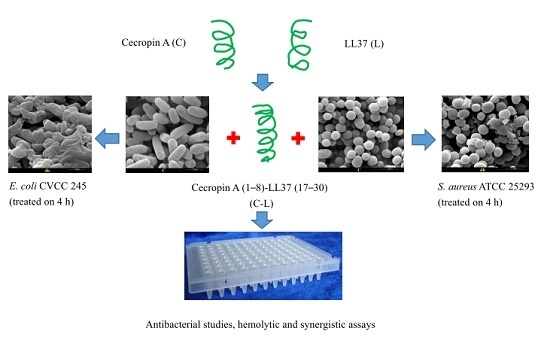
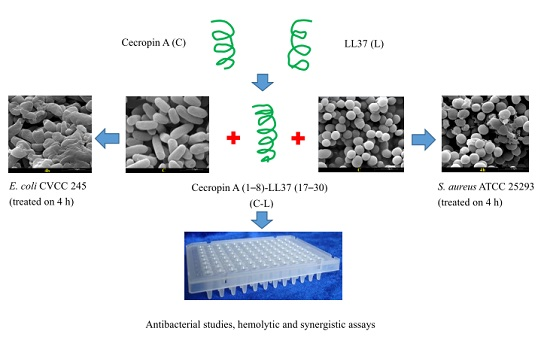
:
1. Introduction
2. Results
2.1. Primary Structural Parameters of Peptides
2.2. Antibacterial and Hemolytic Activity
2.3. Sterilization Speed and Efficacy
2.4. Disruption of Bacterial Cell Surface Structure Integrity
2.5. Synergistic Assay
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Antibiotics
4.2. Design, Biological Information Analysis, and Synthesis of Hybrid Peptides
4.3. Determination of Minimal Inhibitory Concentrations (MICs)
4.4. Hemolytic Assay
4.5. Antibacterial Studies
4.6. SEM Analysis of Bacterial Cells
4.7. Synergy Assays
4.8. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kim, L. Platforms for antibiotic discovery. Nat. Rev. Drug Discov. 2013, 12, 371–387. [Google Scholar]
- Hwang, I.S.; Hwang, J.S.; Hwang, J.H.; Choi, H.; Lee, E.; Kim, Y.; Lee, D.G. Synergistic effect and antibiofilm activity between the antimicrobial peptide coprisin and conventional antibiotics against opportunistic bacteria. Curr. Microbiol. 2013, 66, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.M.; Jin, X.B.; Zhu, J.Y.; Mei, H.F.; Ma, Y.; Chu, F.J.; Wang, Y.; Li, X.B. Expression of the antimicrobial peptide cecropin fused with human lysozyme in Escherichia coli. Appl. Microbiol. Biotechnol. 2010, 87, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Bowles, D.J. Defense-related proteins in higher plants. Annu. Rev. Biochem. 1990, 59, 873–907. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.L.; Hancock, R.E. Cationic host defense (antimicrobial) peptides. Curr. Opin. Immunol. 2006, 18, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, D.; Landuyt, B.; Luyten, W.; Schoofs, L. A comprehensive summary of LL-37, the factoctum human cathelicidin peptide. Cell. Immunol. 2012, 280, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Kyungik, L.; Song, Y.S.; Kyoungho, K.S.; Saeng, L.; Kyung, H.; Kim, Y. Antibiotic activity and structural analysis of the scorpion-derived antimicrobial peptide IsCT and its analogs. Biochem. Biophys. Res. Commun. 2004, 33, 712–719. [Google Scholar]
- Wu, R.J.; Wang, Q.; Zheng, Z.J.; Zhao, L.M.; Shang, Y.J.; Wei, X.B.; Liao, X.D.; Zhang, R.J. Design, characterization and expression of a novel hybrid peptides melittin (1–13)-LL37 (17–30). Mol. Biol. Rep. 2014, 41, 4163–4169. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.Y.; Rang, J.H.; Hahm, K.S. Structure-antibacterial, antitumor and hemolytic activity relationships of cecropin A-magainin 2 and cecropin A-melittin hybrid peptides. J. Pept. Res. 1999, 53, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Naghmouchi, K.; Baah, J.; Hober, D.; Jouy, E. Synergistic effect between colistin and bacteriocins in controlling Gram-negative pathogens and their potential to reduce antibiotic toxicity in mammalian epithelial cells. Antimicrob. Agents Chemother. 2013, 57, 2719–2725. [Google Scholar] [CrossRef] [PubMed]
- Bacalum, M.; Radu, M. Cationic antimicrobial peptides cytotoxicity on mammalian cells: An analysis using therapeutic index integrative concept. Int. J. Pept. Res. Ther. 2015, 21, 47–55. [Google Scholar] [CrossRef]
- Marassi, F.M.; Opella, S.J.; Juvvadi, P.; Merrifield, R.B. Orientation of Cecropin A helices in phospholipid bilayers determined by solid-state NMR spectroscopy. Biophys. J. 1999, 77, 3152–3155. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Kamysz, W.; Amato, G.; Silvestri, C.; del Prete, M.S.; Lukasiak, J.; Scalise, G. Comparative activities of cecropin A, melittin, and cecropin A–melittin peptide CA(1–7)M(2–9)NH2 against multidrug-resistant nosocomial isolates of Acinetobacter baumannii. Peptides 2003, 24, 1315–1318. [Google Scholar] [CrossRef] [PubMed]
- Pavon, N.; Buelna-Chontal, M.; Hernandez-Esquivel, L.; Hernandez, S.; Chavez, E.; Conde, R.; Lanz-Mendoza, H. Mitochondrial inactivation by Anopheles albimanus cecropin 3: Molecular mechanisms. Peptides 2014, 53, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Shin, S.Y.; Maeng, C.Y.; Hahm, K.S. CecropinA-melittin hybrid peptide exerts its antifungal effects by damaging on the plasma membranes of Trichosporon beigelii. Biotechnol. Lett. 1998, 20, 211–214. [Google Scholar] [CrossRef]
- Shin, S.Y.; Lee, M.K.; Kim, K.L.; Hahm, K.S. Structure-antitumor and hemolytic activity relationships of synthetic peptides derived from cecropin A-magainin 2 and cecropin A-melittin hybrid peptides. J. Pept. Res. 1997, 50, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Saugar, J.M.; Rodriguez-Hernandez, M.J.; de la Torre, B.G.; Pachon-Ibanez, M.E.; Fernandez-Reyes, M.; Andreu, D.; Pachon, J.; Rivas, L. Activity of cecropin A-melittin hybrid peptides against colistin-resistant clinical strains of Acinetobacter baumannii: Molecular basis for the differential mechanisms of action. Antimicrob. Agents Chemother. 2006, 50, 1251–1256. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G. Peptide antibiotics and their roles in innate immune. Annu. Rev. Immunol. 1995, 13, 61–92. [Google Scholar] [CrossRef] [PubMed]
- Andreu, D.; Ubach, J.; Boman, A.; Wahlin, B.; Wade, D.; Merrifield, R.B.; Boman, H.G. Shortened cecropin A-melittin hybrids: Significant size reduction retains potent antibiotic activity. FEBS Lett. 1992, 296, 190–194. [Google Scholar] [CrossRef]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta-Biomembr. 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Splith, K.; Neundorf, I. Antimicrobial peptides with cell-penetrating peptide properties and vice versa. Eur. Biophys. J. 2011, 40, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.P.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2008, 30, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Ramos, R.; Silva, J.P.; Rodrigues, A.C.; Costa, R.; Guardao, L.; Schmitt, F.; Soares, R.; Vilanova, M.; Domingues, L.; Gama, M. Wound healing activity of the human antimicrobial peptide LL37. Peptides 2011, 32, 146914–146976. [Google Scholar] [CrossRef] [PubMed]
- Wang, G. Structures of human host defense cathelicidin LL-37 and its smallest antimicrobial peptide KR-12 in lipid micelles. J. Biol. Chem. 2008, 283, 32637–32643. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Y.F.; Han, H.Y.; Miller, D.W.; Wang, G.S. Solution structures of human LL-37 fragments and NMR-based identification of a minimal membrane-targeting antimicrobial and anticancer region. J. Am. Chem. Soc. 2006, 128, 5776–5785. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.; Shi, Y.; Han, J.; Li, Q.; Tang, Y.; Le, G. Design and analysis of structure-activity relationship of novel antimicrobial peptides derived from the conserved sequence of cecropin. J. Pept. Sci. 2008, 14, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.; Hancock, R. The relationship between peptide structure and antibacterial activity. Peptides 2003, 24, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Moraes, L.G.M.; Fazio, M.A.; Vieira, R.F.F.; Nakaie, C.R.; Miranda, M.T.M.; Schreier, S. Conformational and functional studies of gomesin analogues by CD, EPR and fluorescence spectroscopies. Biochim. Biophys. Acta-Biomembr. 2007, 1768, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, S.J.; Park, Y.A.; Kanneganti, N.P.; Mukkisa, H.R.; Crisman, L.L.; Davis, S.E. Modified cysteine-deleted tachyplesin (CDT) analogs as linear antimicrobial peptides: Influence of chain length, positive charge, and hydrophobicity on antimicrobial and hemolytic activity. Int. J. Pept. Res. Ther. 2014, 20, 519–530. [Google Scholar] [CrossRef]
- Christensen, B.; Fink, J.; Merrifiel, R.B.; Mauzerall, D. Channel-forming properties of cecropins and related model compounds incorporated into planar lipid-membranes. Proc. Natl. Acad. Sci. USA 1988, 85, 5072–5076. [Google Scholar] [CrossRef] [PubMed]
- Dennison, S.R.; Wallace, J.; Harris, F.; Phoenix, D.A. Amphiphilic α-helical antimicrobial peptides and their structure/function relationships. Protein Pept. Lett. 2005, 12, 31–39. [Google Scholar] [CrossRef] [PubMed]
- SSpro. Available online: http://scratch.proteomics.ics.uci.edu/ (accessed on 17 June 2016).
- Bonucci, A.; Balducci, E.; Pistolesi, S.; Pogni, R. The defensin-lipid interaction: Insights on the binding states of the human antimicrobial peptide HNP-1 to model bacterial membranes. Biochim. Biophys. Acta-Biomembr. 2013, 1828, 758–764. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Felix, J.B. Peptide-membrane interactions and mechanisms of membrane destruction by amphipathic α-helical antimicrobial peptides. Biochim. Biophys. Acta-Biomembr. 2006, 1758, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Lee, D.G.; Park, Y.; Kim, H.N.; Choi, B.H.; Choi, C.H.; Hahm, K.S. Antibacterial activities of peptides designed as hybrids of antimicrobial peptides. Biotechnol. Lett. 2002, 24, 347–353. [Google Scholar]
- Epand, R.M.; Vogel, H.J. Diversity of antimicrobial peptides and their mechanisms of action. Biochim. Biophys. Acta-Biomembr. 1999, 1462, 11–28. [Google Scholar] [CrossRef]
- Prenner, E.J.; Lewis, R.; Mcelhaney, R.N. The interaction of the antimicrobial peptide gramicidins S with lipid bilater model and biological membranes. Biochim. Biophys. Acta-Biomembr. 1999, 1462, 201–221. [Google Scholar] [CrossRef]
- Durell, S.R.; Raghunathan, G.; Guy, H.R. Modeling the ion channel structure of cecropin. Biophys. J. 1992, 63, 1623–1631. [Google Scholar] [CrossRef]
- Wang, G.S.; Mishra, B.; Epand, R.F.; Epand, R.M. High-quality 3D structures shine light on antibacterial, anti-biofilm and antiviral activities of human cathelicidin LL-37 and its fragments. Biochim. Biophys. Acta-Biomembr. 2014, 1838, 2160–2172. [Google Scholar] [CrossRef] [PubMed]
- Thennarasu, S.; Tan, A.M.; Penumatchu, R.; Shelburne, C.E.; Heyl, D.L.; Ramamoorthy, A. Antimicrobial and membrane disrupting activities of a peptide derived from the human cathelicidin antimicrobial peptide LL37. Biophys. J. 2010, 98, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Gronberg, A.; Zettergren, L.; Agren, M.S. Stability of the cathelicidin peptide LL-37 in a non-healing wound environment. Acta Derm-Venereol. 2011, 91, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.C.; Hui, C.F.; Chen, J.Y.; Wu, J.L. Truncated antimicrobial peptides from marine organisms retain anticancer activity and antibacterial activity against multidrug-resistant Staphylococcus aureus. Peptides 2011, 44, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Dosler, S.; Gerceker, A.A. In vitro activities of nisin alone or in combination with vancomycin and ciprofloxacin against methicillin-resistant and methicillin-susceptible Staphylococcus aureus strains. Chemotherapy 2011, 57, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Dosler, S.; Mataraci, E. In vitro pharmacokinetics of antimicrobial cationic peptides alone and in combination with antibiotics against methicillin resistant Staphylococcus aureus biofilms. Peptides 2013, 49, 53–58. [Google Scholar] [CrossRef] [PubMed]
- APD. Available online: http://aps.unmc.edu/AP/main.php (accessed on 17 June 2016).
- Jpred. Available online: http://www.compbio.dundee.ac.uk/jpred4/index.html (accessed on 17 June 2016).
- Tian, Z.G.; Dong, T.T.; Teng, D.; Yang, Y.I.; Wang, J.H. Design and characterization of novel hybrid peptides from LFB15(W4,10), HP(2–20), and cecropin A based on structure parameters by computer-aided method. Appl. Microbiol. Biotechnol. 2009, 82, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Krogstad, D.J.; Moellering, R.C.J.R. Antimicrobial combinations. In Antibiotics in Laboratory Medicine, 2nd ed.; Lorian, V., Ed.; Williams & Wilkin: Easton, MD, USA, 1986; pp. 537–595. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Amino Acid Sequence | Theoretical MW | Measured MW |
|---|---|---|---|
| C-L | KWKLFKKIFKRIVQRIKDFLRN | 2905.70 | 2905.60 |
| L | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | 4490.57 | 4490.60 |
| C | KWKLFKKIEKVGQNIRDGIIKAGPAVAVVGQATQIAK | 4010.60 | 4010.82 |
| Peptide | Structure | Net Charge | Helical Residue (%) | Hydrophobic Residue (%) |
|---|---|---|---|---|
| C-L | α-helical | +8 | 90.9 | 45 |
| L | α-helical | +6 | 81.1 | 35 |
| C | α-helical | +7 | 81.1 | 45 |
| Bacteria Strains | Antimicrobial Peptide | p-Value | |||
|---|---|---|---|---|---|
| C | L | C-L | |||
| MIC (µg/mL) | E. coli K88 | 64 ± 0.72 a | 61 ± 1.78 b | 8 ± 0.33 c | <0.01 |
| E. coli CVCC 245 | 4 ± 0.28 c | 84 ± 1.54 a | 7 ± 0.29 b | <0.01 | |
| S. aureus CVCC 26003 | 128 ± 7.57 a | 36 ± 1.55 b | 7 ± 0.28 c | <0.01 | |
| S. aureus ATCC 25923 | 198 ± 10.1 a | 58 ± 1.68 b | 2 ± 0.03 c | <0.01 | |
| L. mono. CVCC 1599 | 64 ± 2.99 a | 13 ± 0.43 b | 2 ± 0.06 c | <0.01 | |
| M. luteus CVCC 28001 | 154 ± 5.49 a | 90 ± 1.43 b | 16 ± 0.18 c | <0.01 | |
| HC50 (µg/mL) | Sheep erythrocyte cell | 169 ± 15.1 b | 32 ± 0.68 c | 221 ± 3.27 a | <0.01 |
| Bacteria Strains | Time (h) | Antimicrobial Peptides | p-Value | ||
|---|---|---|---|---|---|
| C | L | C-L | |||
| Viable S. aureus concentration (×108 CFU/mL) | 1 | 0.54 ± 0.007 b | 0.90 ± 0.020 a | 0.43 ± 0.017 c | <0.01 |
| 2 | 0.40 ± 0.035 b | 0.53 ± 0.026 a | 0.30 ± 0.012 c | <0.01 | |
| 3 | 0.21 ± 0.014 a | 0.23 ± 0.011 a | 0.18 ± 0.020 b | 0.014 | |
| 4 | 0.09 ± 0.005 | 0.09 ± 0.002 | 0.08 ± 0.003 | 0.079 | |
| 5 | 0.01 ± 0.0005 | 0.01 ± 0.0003 | 0.01 ± 0.0001 | 0.298 | |
| Viable E. coli concentration (×108 CFU/mL) | 1 | 0.44 ± 0.028 a | 0.44 ± 0.027 a | 0.29 ± 0.015 b | <0.01 |
| 2 | 0.30 ± 0.021 b | 0.35 ± 0.014 a | 0.16 ± 0.026 c | <0.01 | |
| 3 | 0.15 ± 0.014 a | 0.14 ± 0.020 a | 0.05 ± 0.003 b | <0.01 | |
| 4 | 0.06 ± 0.002 | 0.05 ± 0.002 | 0.05 ± 0.003 | 0.061 | |
| 5 | 0.01 ± 0.0002 | 0.01 ± 0.003 | 0.01 ± 0.0001 | 0.659 | |
| Combination Group | Fractional Inhibitory Concentration (FIC) Index | p-Value | ||||
|---|---|---|---|---|---|---|
| C-L + Chloramphenicol | C-L + Thiamphenicol | C-L + Neomycin Sulfate | C-L + Penicillin G | C-L + Kanamycin | ||
| E. coli CVCC 245 | 1.00 ± 0.141 c | 1.50 ± 0.001 a | 0.313 ± 0.001 d | 1.50 ± 0.003 a | 1.25 ± 0.002 b | <0.01 |
| S. aureus ATCC 25923 | 0.375 ± 0.005 c | 0.188 ± 0.002 e | 0.250 ± 0.004 d | 1.13 ± 0.005 a | 0.750 ± 0.003 b | <0.01 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, X.-B.; Wu, R.-J.; Si, D.-Y.; Liao, X.-D.; Zhang, L.-L.; Zhang, R.-J. Novel Hybrid Peptide Cecropin A (1–8)-LL37 (17–30) with Potential Antibacterial Activity. Int. J. Mol. Sci. 2016, 17, 983. https://doi.org/10.3390/ijms17070983
Wei X-B, Wu R-J, Si D-Y, Liao X-D, Zhang L-L, Zhang R-J. Novel Hybrid Peptide Cecropin A (1–8)-LL37 (17–30) with Potential Antibacterial Activity. International Journal of Molecular Sciences. 2016; 17(7):983. https://doi.org/10.3390/ijms17070983
Chicago/Turabian StyleWei, Xu-Biao, Ru-Juan Wu, Da-Yong Si, Xiu-Dong Liao, Lu-Lu Zhang, and Ri-Jun Zhang. 2016. "Novel Hybrid Peptide Cecropin A (1–8)-LL37 (17–30) with Potential Antibacterial Activity" International Journal of Molecular Sciences 17, no. 7: 983. https://doi.org/10.3390/ijms17070983