
DDX3X Biomarker Correlates with Poor Survival in Human Gliomas

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result
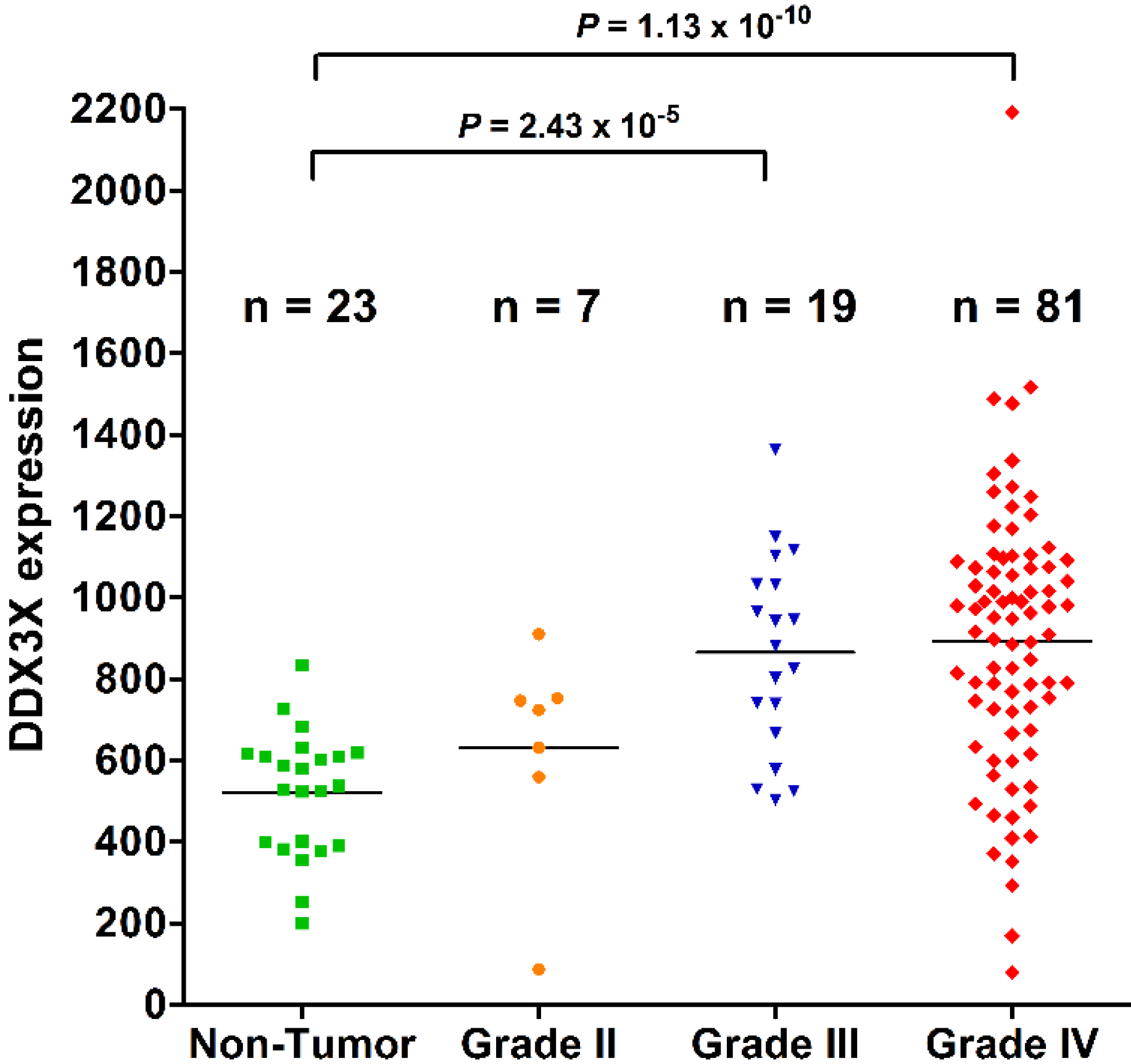
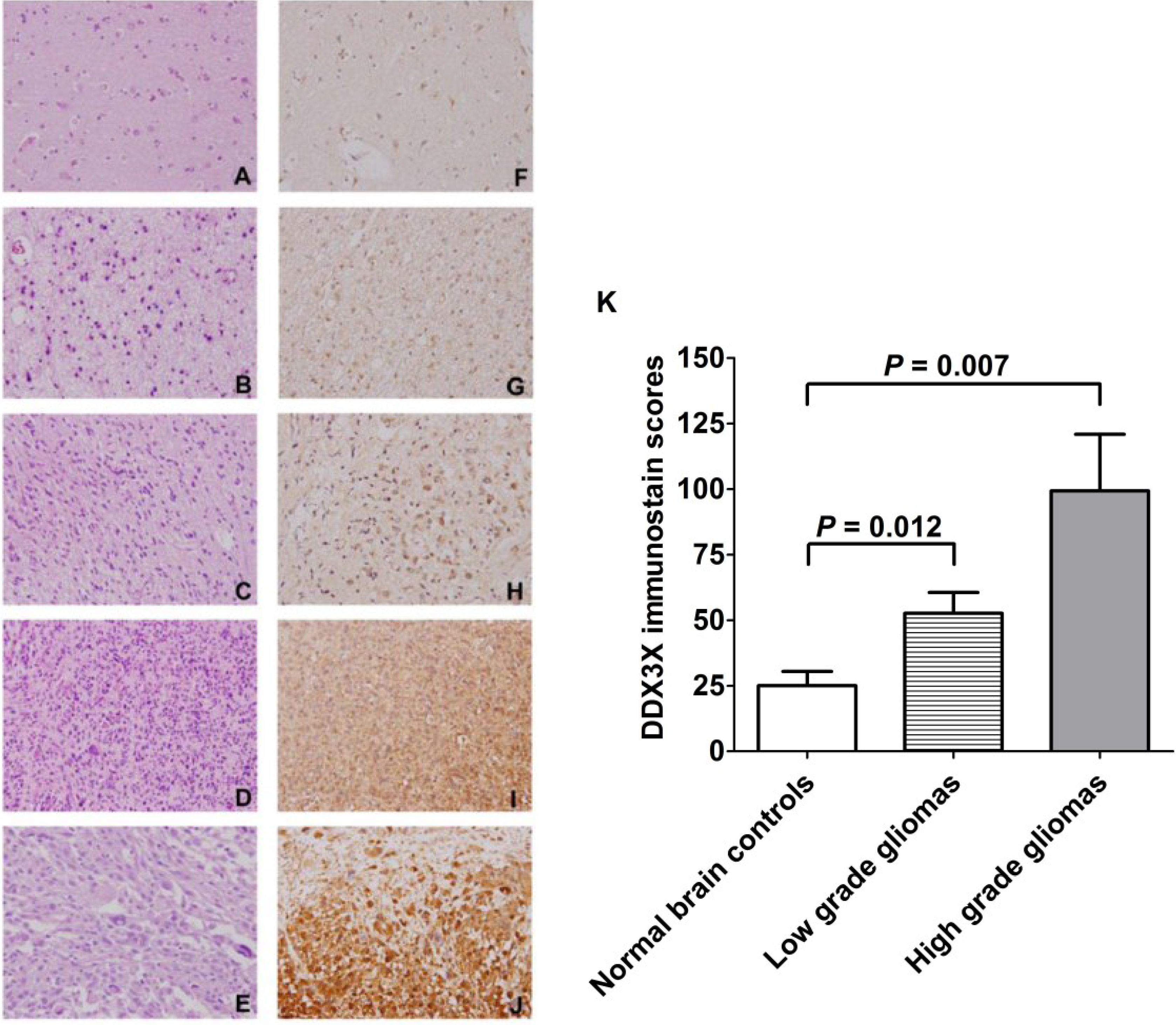
2.1. DDX3X Expression Positively Correlates with WHO Grading of Human Gliomas

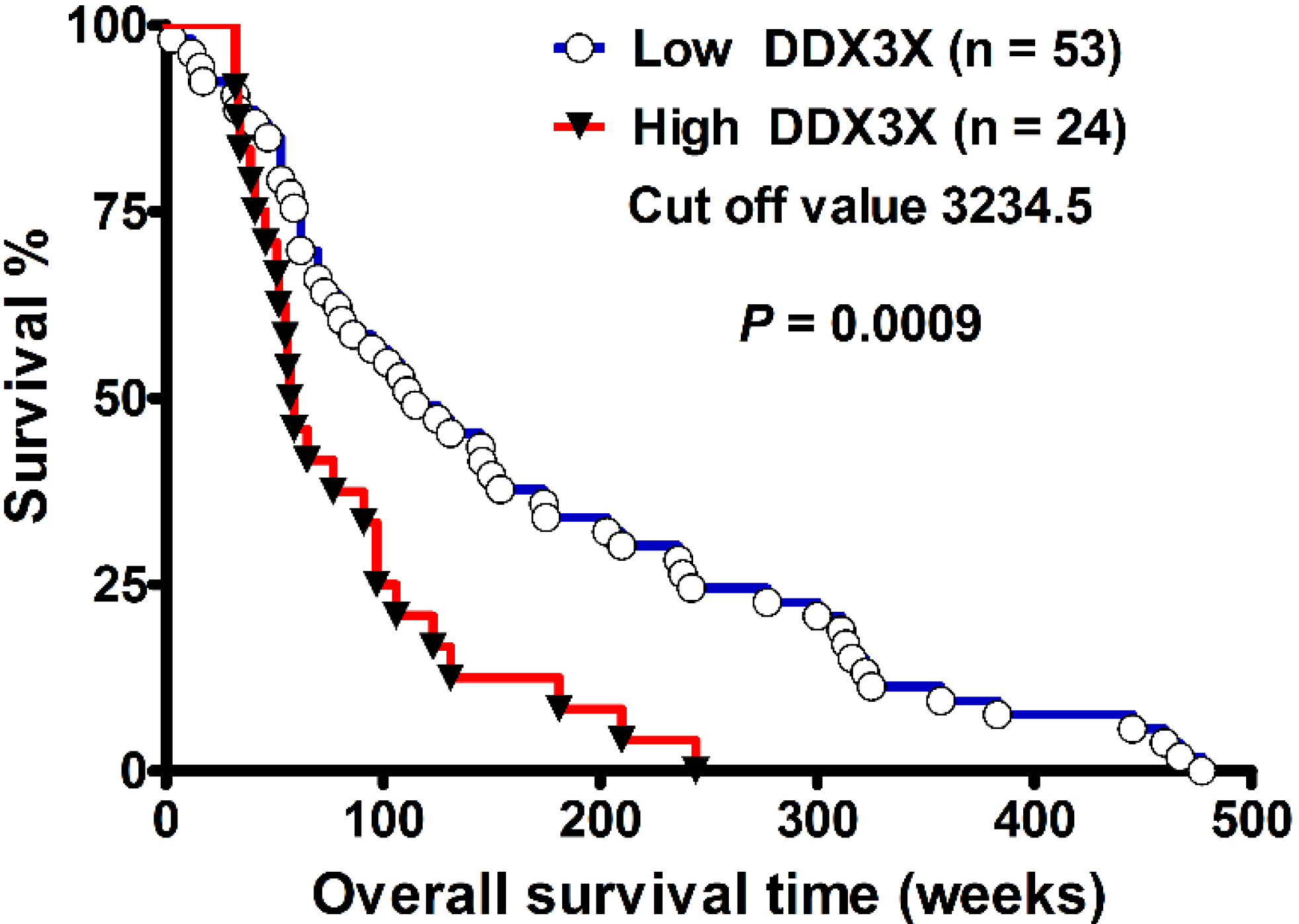
2.2. DDX3X Expression Correlates with Poor Survival in High-Grade Gliomas

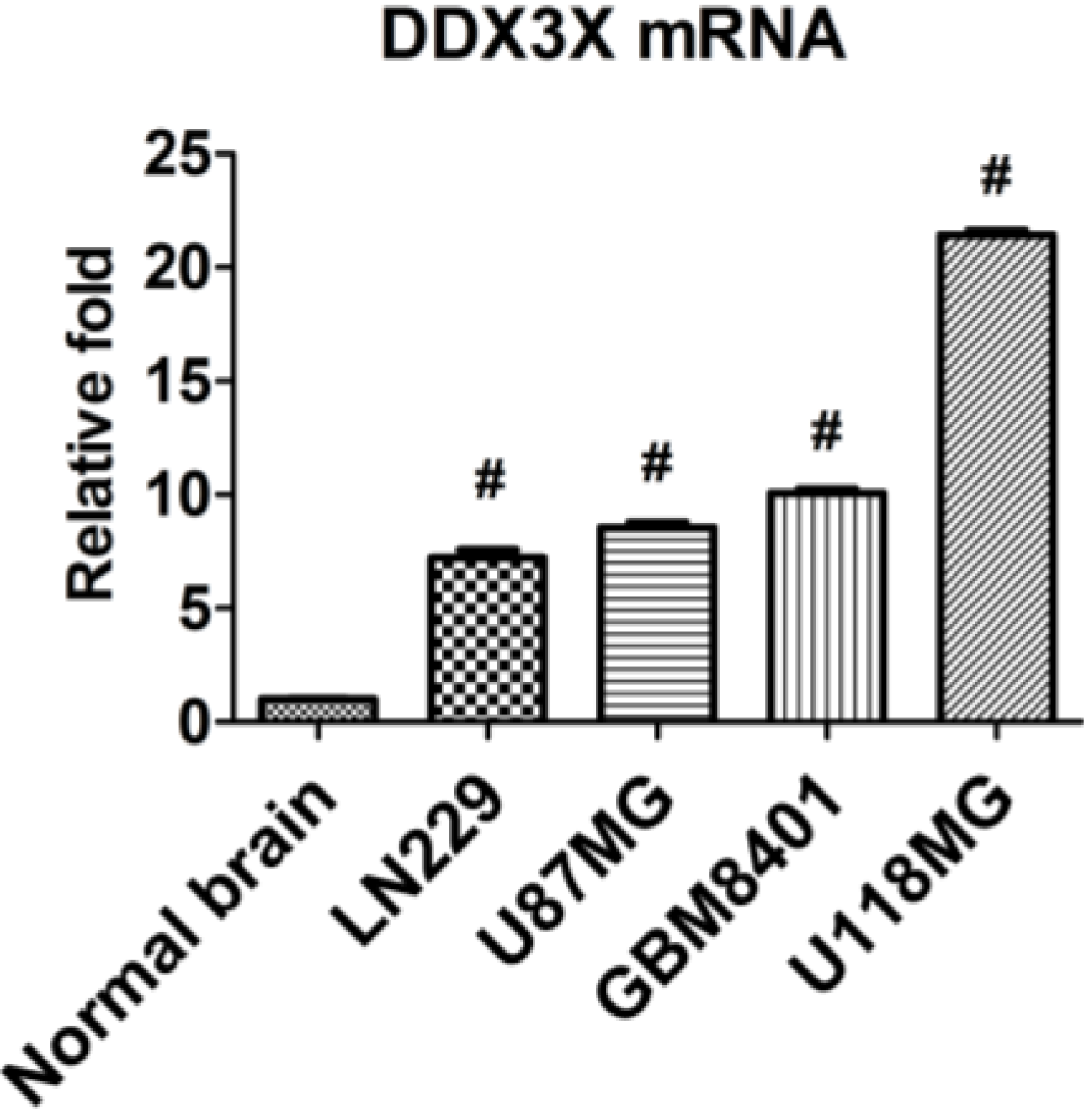
2.3. DDX3X mRNA Expression Is Increased in Human Glioma Cells
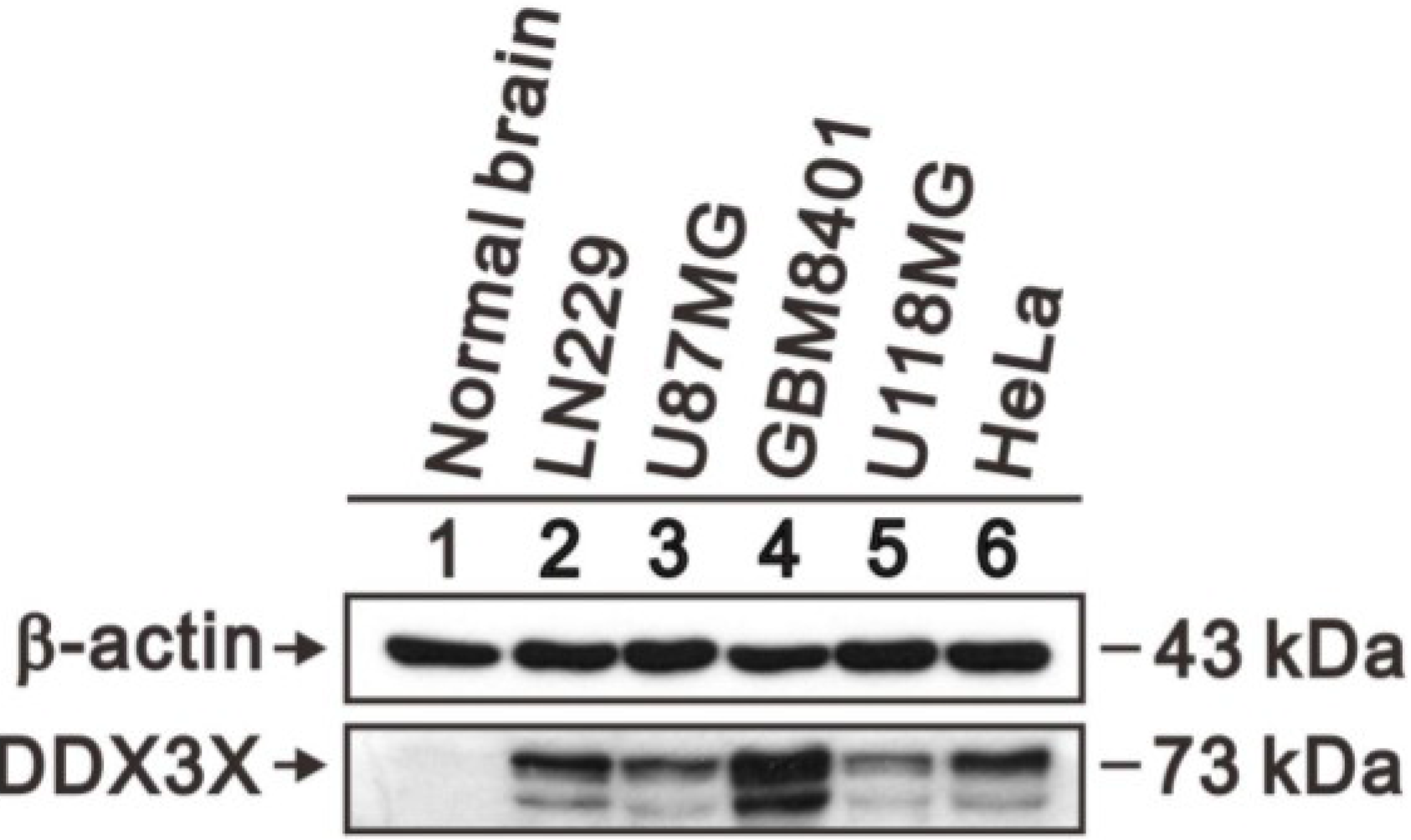
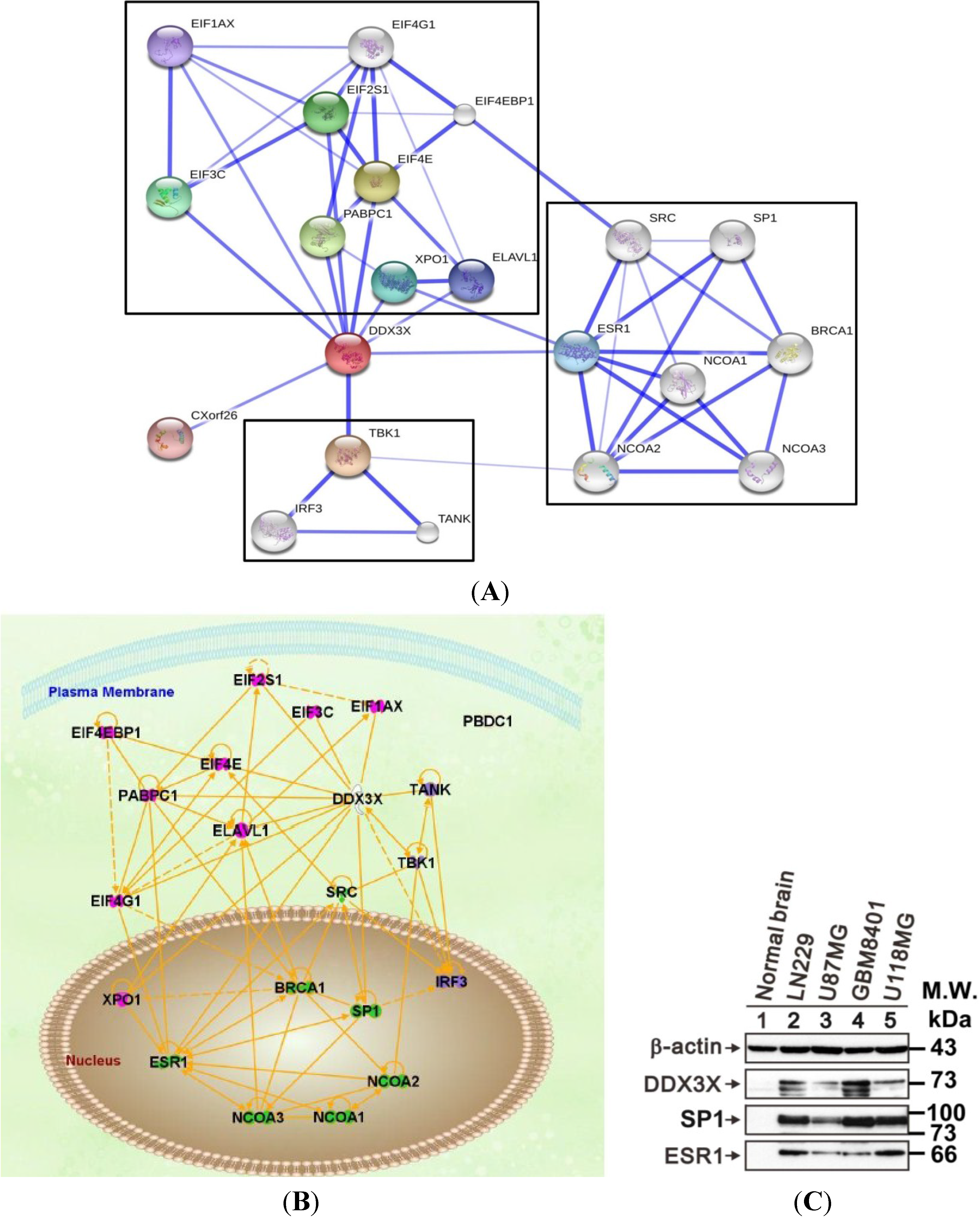
2.4. DDX3X Protein Overexpresses in Human Glioma Cells


2.5. DDX3X Protein Production Overexpressed in Human High-Grade Gliomas

2.6. DDX3X Hub Protein and the Protein–Protein Interactions

3. Discussion
4. Materials and Methods
4.1. Study Ethics and DDX3X Gene Expression in Human Brain Tumor
4.2. RNA Isolation and Quantitative RT-PCR (qRT-PCR)
4.3. Cell Lysate Preparation and Western Blots
4.4. Immunohistochemical (IHC) Staining
4.5. Evaluation of DDX3X Immunostain Scores
4.6. PPI Network and Signaling Pathways Analysis
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jan, H.J.; Lee, C.C.; Shih, Y.L.; Hueng, D.Y.; Ma, H.I.; Lai, J.H.; Wei, H.W.; Lee, H.M. Osteopontin regulates human glioma cell invasiveness and tumor growth in mice. Neuro Oncol. 2010, 12, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.I.; Hueng, D.Y.; Shui, H.A.; Han, J.M.; Wang, C.H.; Lai, Y.H.; Cheng, S.Y.; Xiao, X.; Chen, M.T.; Yang, Y.P. Intratumoral decorin gene delivery by AAV vector inhibits brain glioblastomas and prolongs survival of animals by inducing cell differentiation. Int. J. Mol. Sci. 2014, 15, 4393–4414. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.S.; Lin, C.; Cheng, Y.P.; Yu, Y.L.; Tang, C.T.; Hueng, D.Y. Epithelial cell transformation sequence 2 is a potential biomarker of unfavorable survival in human gliomas. Neurol. India 2014, 62, 406–409. [Google Scholar] [PubMed]
- Roversi, G.; Pfundt, R.; Moroni, R.F.; Magnani, I.; van Reijmersdal, S.; Pollo, B.; Straatman, H.; Larizza, L.; Schoenmakers, E.F. Identification of novel genomic markers related to progression to glioblastoma through genomic profiling of 25 primary glioma cell lines. Oncogene 2006, 25, 1571–1583. [Google Scholar] [CrossRef] [PubMed]
- Yedavalli, V.S.; Neuveut, C.; Chi, Y.H.; Kleiman, L.; Jeang, K.T. Requirement of DDX3 DEAD box RNA helicase for HIV-1 Rev-RRE export function. Cell 2004, 119, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Garbelli, A.; Beermann, S.; di Cicco, G.; Dietrich, U.; Maga, G. A motif unique to the human DEAD-box protein DDX3 is important for nucleic acid binding, ATP hydrolysis, RNA/DNA unwinding and HIV-1 replication. PLoS ONE 2011, 6, e19810. [Google Scholar] [CrossRef] [PubMed]
- Dolan, P.T.; Roth, A.P.; Xue, B.; Sun, R.; Dunker, A.K.; Uversky, V.N.; LaCount, D.J. Intrinsic disorder mediates hepatitis C virus core-host cell protein interactions. Protein Sci. 2015, 24, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Cheng, X.R.; Zhou, W.X.; Zhang, Y.X. Gene expression patterns of hippocampus and cerebral cortex of senescence-accelerated mouse treated with Huang-Lian-Jie-Du decoction. Neurosci. Lett. 2008, 439, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Botlagunta, M.; Vesuna, F.; Mironchik, Y.; Raman, A.; Lisok, A.; Winnard, P., Jr.; Winnard, P.; Mukadam, S.; van Diest, P.; Chen, J.H.; et al. Oncogenic role of DDX3 in breast cancer biogenesis. Oncogene 2008, 27, 3912–3922. [Google Scholar] [CrossRef] [PubMed]
- Bol, G.M.; Raman, V.; van der Groep, P.; Vermeulen, J.F.; Patel, A.H.; van der Wall, E.; van Diest, P.J. Expression of the RNA helicase DDX3 and the hypoxia response in breast cancer. PLoS ONE 2013, 8, e63548. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.W.; Lee, M.C.; Wang, J.; Chen, C.Y.; Cheng, Y.W.; Lee, H. DDX3 loss by p53 inactivation promotes tumor malignancy via the MDM2/Slug/E-cadherin pathway and poor patient outcome in non-small-cell lung cancer. Oncogene 2014, 33, 1515–1526. [Google Scholar] [CrossRef] [PubMed]
- Koshio, J.; Kagamu, H.; Nozaki, K.; Saida, Y.; Tanaka, T.; Shoji, S.; Igarashi, N.; Miura, S.; Okajima, M.; Watanabe, S.; et al. DEAD/H (Asp-Glu-Ala-Asp/His) box polypeptide 3, X-linked is an immunogenic target of cancer stem cells. Cancer Immunol. Immunother. 2013, 62, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Schroder, P.C.; Fernandez-Irigoyen, J.; Bigaud, E.; Serna, A.; Renandez-Alcoceba, R.; Lu, S.C.; Mato, J.M.; Prieto, J.; Corrales, F.J. Proteomic analysis of human hepatoma cells expressing methionine adenosyltransferase I/III: Characterization of DDX3X as a target of S-adenosylmethionine. J. Proteomics 2012, 75, 2855–2868. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Yang, Z.L.; Xiong, L.; Zou, Q.; Yuan, Y.; Li, J.; Liang, L.; Chen, M.; Chen, S. Nectin-2 and DDX3 are biomarkers for metastasis and poor prognosis of squamous cell/adenosquamous carcinomas and adenocarcinoma of gallbladder. Int. J. Clin. Exp. Pathol. 2013, 6, 179–190. [Google Scholar] [PubMed]
- Sun, M.; Song, L.; Zhou, T.; Gillespie, G.Y.; Jope, R.S. The role of DDX3 in regulating Snail. Biochim. Biophys. Acta 2011, 1813, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.P.; Lin, C.; Lin, P.Y.; Cheng, C.Y.; Ma, H.I.; Chen, C.M.; Hueng, D.Y. Midkine expression in high grade gliomas: Correlation of this novel marker with proliferation and survival in human gliomas. Surg. Neurol. Int. 2014, 5. [Google Scholar] [CrossRef]
- Tsai, W.C.; Chen, Y.; Huang, L.C.; Lee, H.S.; Ma, H.I.; Huang, S.M.; Sytwu, H.K.; Hueng, D.Y. EMMPRIN expression positively correlates with WHO grades of astrocytomas and meningiomas. J. Neurooncol. 2013, 114, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.I.; Chiou, S.H.; Hueng, D.Y.; Tai, L.K.; Huang, P.I.; Kao, C.L.; Chen, Y.W.; Sytwu, H.K. Celecoxib and radioresistant glioblastoma-derived CD133+ cells: improvement in radiotherapeutic effects. Laboratory investigation. J. Neurosurg. 2011, 114, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.C.; Chi, C.W.; Chau, G.Y.; Li, F.Y.; Tsai, Y.H.; Wu, J.C.; Wu Lee, Y.H. DDX3, a DEAD box RNA helicase, is deregulated in hepatitis virus-associated hepatocellular carcinoma and is involved in cell growth control. Oncogene 2006, 25, 1991–2003. [Google Scholar] [CrossRef] [PubMed]
- Porter, W.; Wang, F.; Wang, W.; Duan, R.; Safe, S. Role of estrogen receptor/Sp1 complexes in estrogen-induced heat shock protein 27 gene expression. Mol. Endocrinol. 1996, 10, 1371–1378. [Google Scholar] [PubMed]
- Wu, F.; Ivanov, I.; Xu, R.; Safe, S. Role of SP transcription factors in hormone-dependent modulation of genes in MCF-7 breast cancer cells: Microarray and RNA interference studies. J. Mol. Endocrinol. 2009, 42, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Soulat, D.; Burckstummer, T.; Westermayer, S.; Goncalves, A.; Bauch, A.; Stefanovic, A.; Hantschel, O.; Bennett, K.L.; Decker, T.; Superti-Furga, G. The DEAD-box helicase DDX3X is a critical component of the TANK-binding kinase 1-dependent innate immune response. EMBO J. 2008, 27, 2135–2146. [Google Scholar] [CrossRef] [PubMed]
- Hueng, D.Y.; Lin, G.J.; Huang, S.H.; Liu, L.W.; Ju, D.T.; Chen, Y.W.; Sytwu, H.K.; Chang, C.; Huang, S.M.; Yeh, Y.S.; et al. Inhibition of Nodal suppresses angiogenesis and growth of human gliomas. J. Neurooncol. 2011, 104, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Jan, H.J.; Lai, J.H.; Ma, H.I.; Hueng, D.Y.; Lee, Y.C.; Cheng, Y.Y.; Liu, L.W.; Wei, H.W.; Lee, H.M. Nodal promotes growth and invasion in human gliomas. Oncogene 2010, 29, 3110–3123. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, Q.; Liu, D.; Wang, D.; Chen, H.; Xiao, S.; Fang, L. Molecular cloning, functional characterization and antiviral activity of porcine DDX3X. Biochem. Biophys. Res. Commun. 2014, 443, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.C.; Hueng, D.Y.; Lin, C.K. Nuclear overexpression of urocortin discriminates primary brain tumors from reactive gliosis. APMIS 2015, 123, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hueng, D.-Y.; Tsai, W.-C.; Chiou, H.-Y.C.; Feng, S.-W.; Lin, C.; Li, Y.-F.; Huang, L.-C.; Lin, M.-H. DDX3X Biomarker Correlates with Poor Survival in Human Gliomas. Int. J. Mol. Sci. 2015, 16, 15578-15591. https://doi.org/10.3390/ijms160715578
Hueng D-Y, Tsai W-C, Chiou H-YC, Feng S-W, Lin C, Li Y-F, Huang L-C, Lin M-H. DDX3X Biomarker Correlates with Poor Survival in Human Gliomas. International Journal of Molecular Sciences. 2015; 16(7):15578-15591. https://doi.org/10.3390/ijms160715578
Chicago/Turabian StyleHueng, Dueng-Yuan, Wen-Chiuan Tsai, Hsin-Ying Clair Chiou, Shao-Wei Feng, Chin Lin, Yao-Feng Li, Li-Chun Huang, and Ming-Hong Lin. 2015. "DDX3X Biomarker Correlates with Poor Survival in Human Gliomas" International Journal of Molecular Sciences 16, no. 7: 15578-15591. https://doi.org/10.3390/ijms160715578