
Functional Analysis of the Maize C-Repeat/DRE Motif-Binding Transcription Factor CBF3 Promoter in Response to Abiotic Stress


Abstract
:
1. Introduction
2. Results
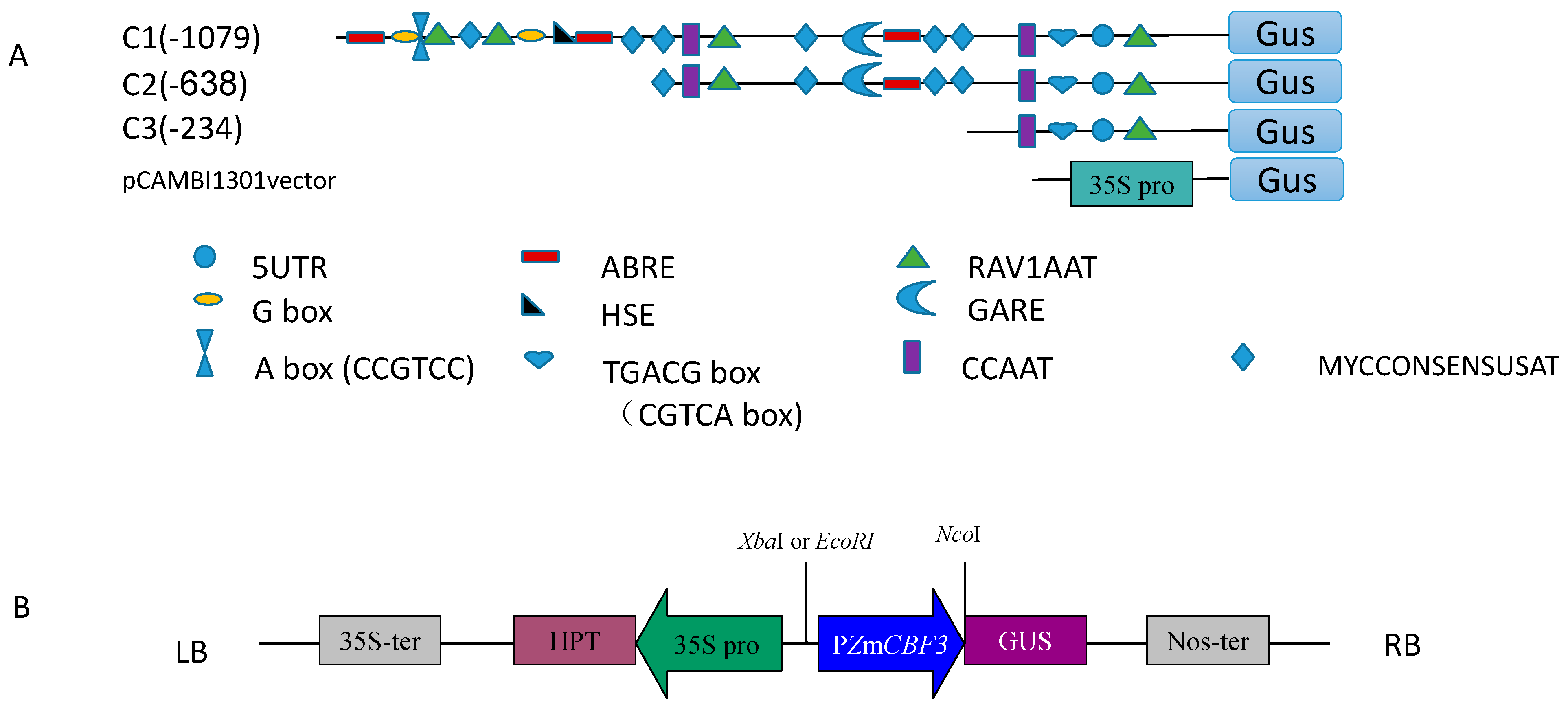
2.1. Promoter Sequence Analysis

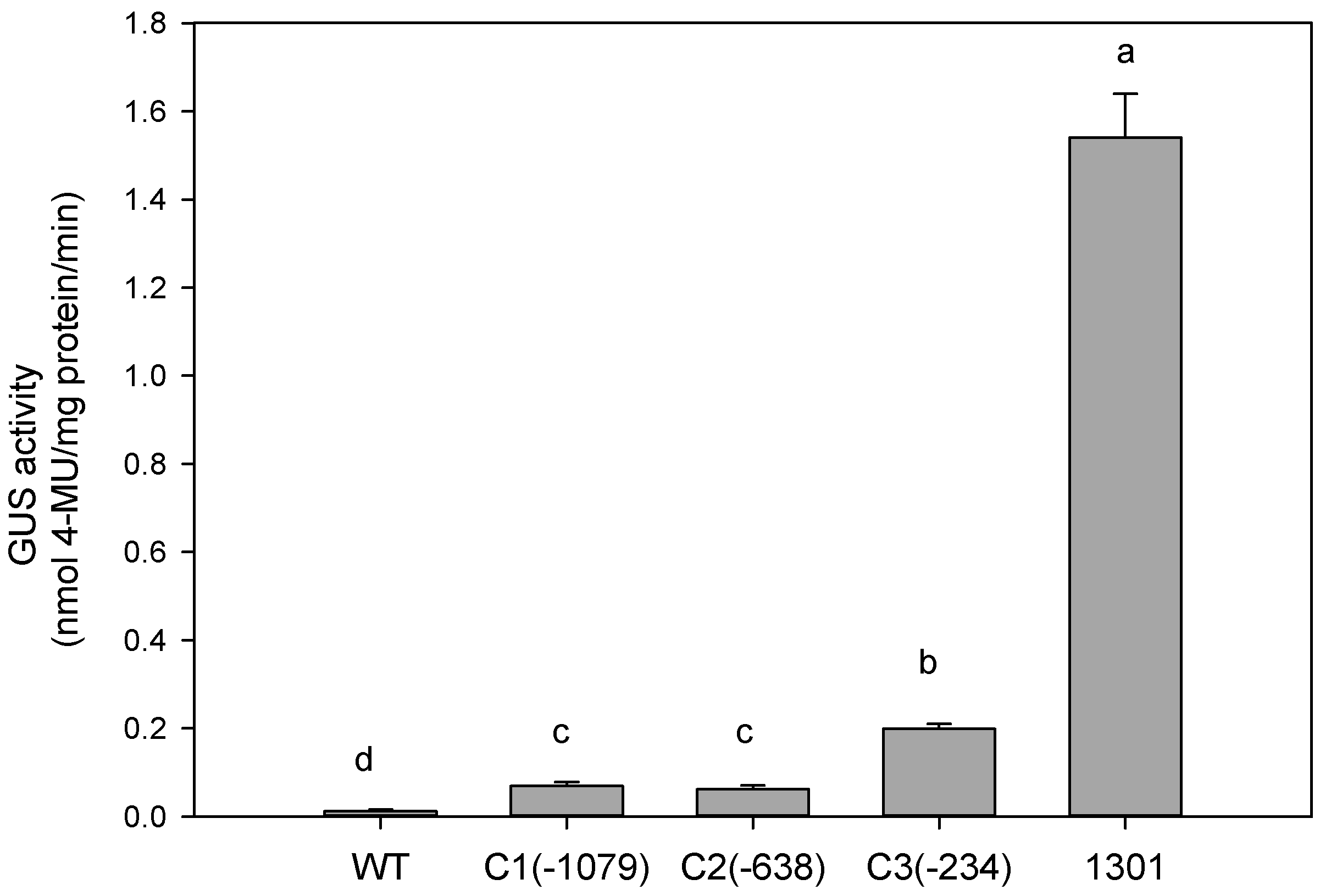
2.2. Basal Expression Analysis



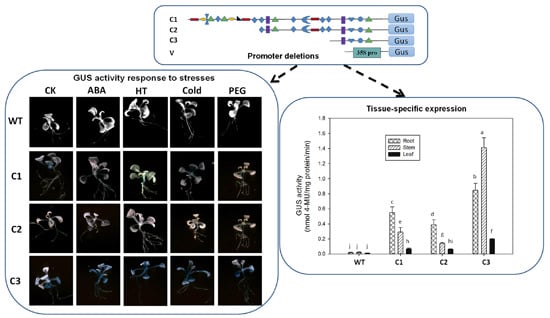
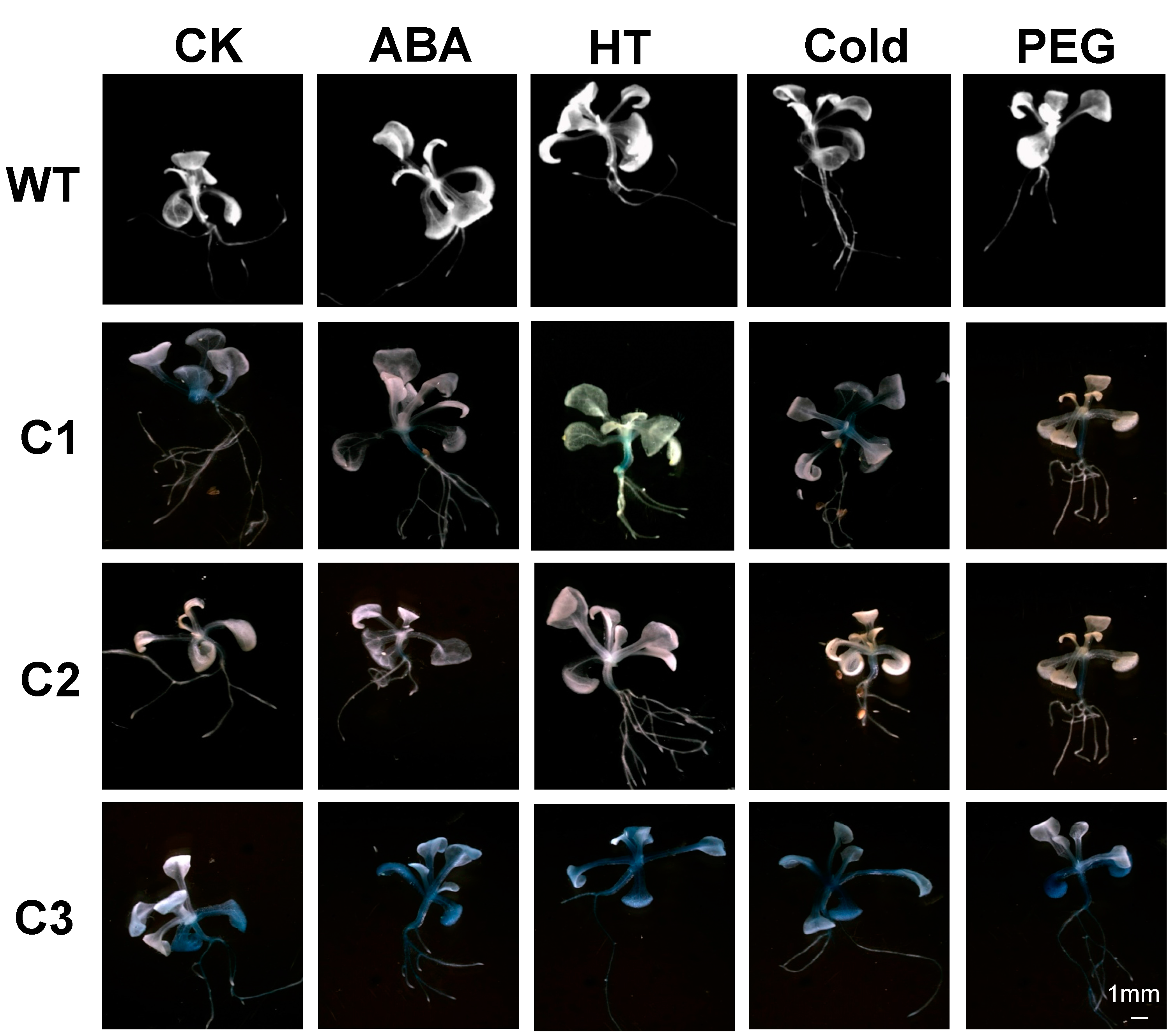
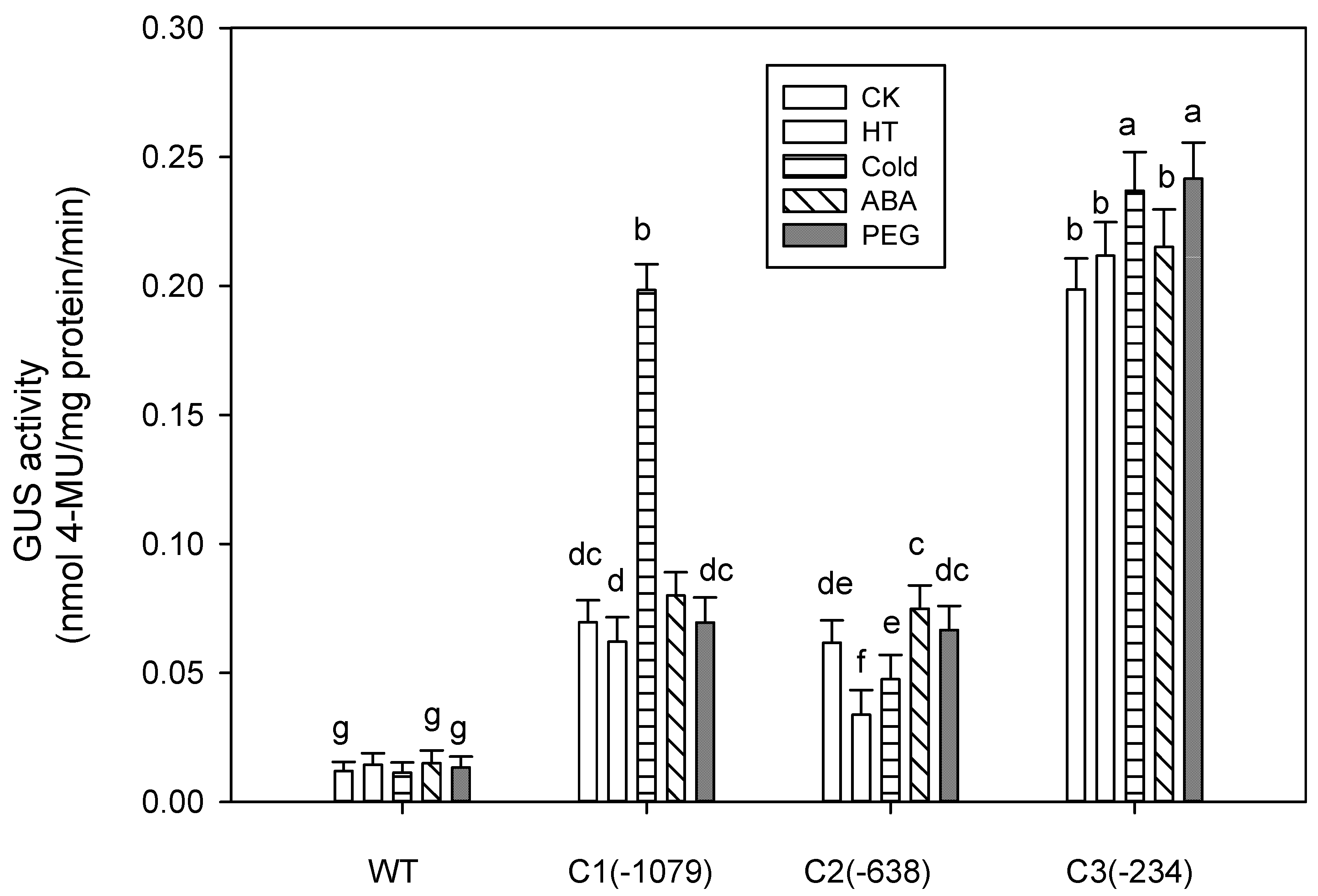
2.3. Abiotic Stress-Induced Expression Analysis

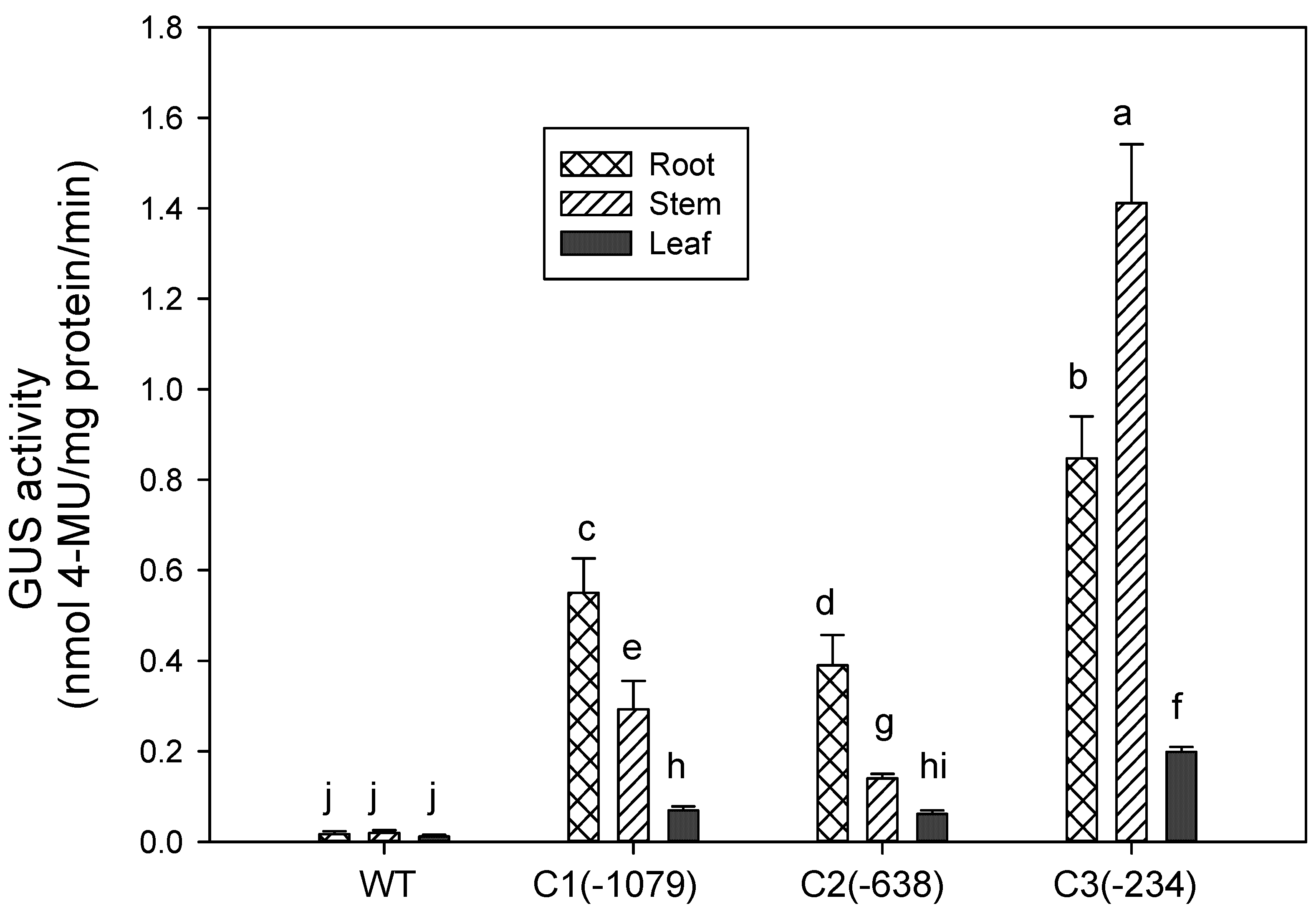
2.4. Tissue-Specific Expression Analysis

3. Discussion
3.1. Basal Expression Analysis of the Deletion Promoters
3.2. Abiotic Stress-Induced Expression Analysis
3.3. Expression of PZmCBF3 Is Tissue-Specific
4. Experimental Section
4.1. Plant Materials, Growth Condition, and Bacterial Strains
4.2. Genomic DNA Extraction
4.3. Isolation of the ZmCBF3 Gene Promoter

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligo Name | Sequence (5′→3′) |
|---|---|
| Pcbf-F | TGAGCCGGACGGTCTGCTAT |
| C2-638 | GGAATTCAGAGATAACGCCAGAACGAG |
| C3-234 | GGAATTCGCCTTGTTCCGTTGTTCG |
| Pcbf-R | TGCCATGGCTGCTGCTGTGACTGTGA |
| Hyg-F | GATGTTGGCGACCTCGTATT |
| Hyg-R | TCGTTATGTTTATCGGCACTTT |
4.4. Sequence Analysis
4.5. Construction of the PZmCBF3 Deletion Constructs
4.6. Arabidopsis Plant Transformation with Different PZmCBF3:GUS Constructs
4.7. Abiotic Stress Treatments
4.8. Histochemical and Fluorometric Analyses of GUS Activity
4.9. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stockinger, E.J.; Gilmour, S.J.; Thomashow, M.F. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proc. Natl. Acad. Sci. USA 1997, 94, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, S.J.; Zarka, D.G.; Stockinger, E.J.; Salazar, M.P.; Houghton, J.M.; Thomashow, M.F. Low temperature regulation of the Arabidopsis CBF family of AP2 transcriptional activators as an early step in cold-induced COR gene expression. Plant J. 1998, 16, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, M.F. Molecular basis of plant cold acclimation: Insights gained from studying the CBF cold response pathway. Plant Physiol. 2010, 154, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell Online 1998, 10, 1391–1406. [Google Scholar] [CrossRef]
- Haake, V.; Cook, D.; Riechmann, J.; Pineda, O.; Thomashow, M.F.; Zhang, J.Z. Transcription factor CBF4 is a regulator of drought adaptation in Arabidopsis. Plant Physiol. 2002, 130, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Jaglo, K.R.; Kleff, S.; Amundsen, K.L.; Zhang, X.; Haake, V.; Zhang, J.Z.; Deits, T.; Thomashow, M.F. Components of the Arabidopsis C-repeat/dehydration-responsive element binding factor cold-response pathway are conserved in Brassica napus and other plant species. Plant Physiol. 2001, 127, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.-W.; Rodriguez, E.M.; Close, T.J. Barley CBF3 gene identification, expression pattern, and map location. Plant Physiol. 2002, 129, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Skinner, J.S.; von Zitzewitz, J.; Szűcs, P.; Marquez-Cedillo, L.; Filichkin, T.; Amundsen, K.; Stockinger, E.J.; Thomashow, M.F.; Chen, T.H.; Hayes, P.M. Structural, functional, and phylogenetic characterization of a large CBF gene family in barley. Plant Mol. Biol. 2005, 59, 533–551. [Google Scholar] [CrossRef] [PubMed]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt-and cold-responsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Fei, S.Z. Functional and phylogenetic analysis of a DREB/CBF-like gene in perennial ryegrass (Lolium perenne L.). Planta 2006, 224, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Chen, M.; Li, L.; Xu, Z.; Chen, X.; Guo, J.; Ma, Y. Over-expression of the soybean GmERF3 gene, an AP2/ERF type transcription factor for increased tolerances to salt, drought, and diseases in transgenic tobacco. J. Exp. Bot. 2009, 60, 3781–3796. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Sakuma, Y.; Li, J.; Liu, Q.; Li, Y.-Q.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Cloning and functional analysis of a novel DREB1/CBF transcription factor involved in cold-responsive gene expression in Zea mays L. Plant Cell Physiol. 2004, 45, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Meyerowitz, E.M. The AP2/EREBP family of plant transcription factors. Biol. Chem. 1998, 379, 633–646. [Google Scholar] [PubMed]
- Sakuma, Y.; Liu, Q.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration-and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 2002, 290, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Wang, F.; Jia, D.; Li, J.; Zhang, Y.; Jia, C.; Wang, D.; Pan, H. Cloning and functional analysis of the promoter of a stress-inducible gene (ZmRXO1) in maize. Plant Mol. Biol. Rep. 2014, 33, 1–9. [Google Scholar] [CrossRef]
- Yang, Q.; Yuan, D.; Shi, L.; Capell, T.; Bai, C.; Wen, N.; Lu, X.; Sandmann, G.; Christou, P.; Zhu, C. Functional characterization of the Gentiana lutea zeaxanthin epoxidase (GlZEP) promoter in transgenic tomato plants. Transgenic Res. 2012, 21, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, J.; Li, J.; Zhang, S.; Pan, H. Functional analyses of the maize CKS2 gene promoter in response to abiotic stresses and hormones. Acta Physiol. Plant. 2014, 36, 1867–1878. [Google Scholar] [CrossRef]
- Dalal, M.; Chinnusamy, V.; Bansal, K.C. Isolation and functional characterization of lycopene β-cyclase (CYC-B) promoter from Solanum habrochaites. BMC Plant Biol. 2010, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Hübel, A.; Schöffl, F. Arabidopsis heat shock factor: Isolation and characterization of the gene and the recombinant protein. Plant Mol. Biol. 1994, 26, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Schöffl, F.; Prändl, R.; Reindl, A. Regulation of the heat-shock response. Plant Physiol. 1998, 117, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.-H.; Hong, X.; Agarwal, M.; Zhu, J.-K. ICE1: A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev. 2003, 17, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Hobo, T.; Asada, M.; Kowyama, Y.; Hattori, T. ACGT-containing abscisic acid response element (ABRE) and coupling element 3 (CE3) are functionally equivalent. Plant J. 1999, 19, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Zhang, P.; Ho, T. Modular nature of abscisic acid (ABA) response complexes: Composite promoter units that are necessary and sufficient for ABA induction of gene expression in barley. Plant Cell Online 1996, 8, 1107–1119. [Google Scholar] [CrossRef]
- Hobo, T.; Kowyama, Y.; Hattori, T. A bZIP factor, TRAB1, interacts with VP1 and mediates abscisic acid-induced transcription. Proc. Natl. Acad. Sci. USA 1999, 96, 15348–15353. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.I.; Hong, J.H.; Ha, J.O.; Kang, J.Y.; Kim, S.Y. ABFs, a family of ABA-responsive element binding factors. J. Biol. Chem. 2000, 275, 1723–1730. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, L.G.G.; Riaño-Pachón, D.M.; Schrago, C.G.; dos Santos, R.V.; Mueller-Roeber, B.; Vincentz, M. The role of bZIP transcription factors in green plant evolution: Adaptive features emerging from four founder genes. PLoS ONE 2008, 3, e2944. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Lata, C.; Mishra, A.K.; Muthamilarasan, M.; Bonthala, V.S.; Khan, Y.; Prasad, M. Genome-wide investigation and expression profiling of AP2/ERF transcription factor superfamily in foxtail millet (Setaria italica L.). PLoS ONE 2014, 9, e113092. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.T.; Taylor, C.G.; Acedo, G.N.; Cheng, C.L.; Conkling, M.A. Characterization of cis-acting sequences regulating root-specific gene expression in tobacco. Plant Cell Online 1991, 3, 371–382. [Google Scholar]
- Kim, D.W.; Lee, S.H.; Choi, S.B.; Won, S.K.; Heo, Y.K.; Cho, M.; Park, Y.I.; Cho, H.T. Functional conservation of a root hair cell-specific cis-element in angiosperms with different root hair distribution patterns. Plant Cell Online 2006, 18, 2958–2970. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.T.; Cosgrove, D.J. Regulation of root hair initiation and expansin gene expression in Arabidopsis. Plant Cell Online 2002, 14, 3237–3253. [Google Scholar] [CrossRef]
- Lin, C.; Choi, H.-S.; Cho, H.-T. Root hair-specific EXPANSIN A7 is required for root hair elongation in Arabidopsis. Mol. Cells 2011, 31, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wen, F.; Zhao, X.; Hawes, M. Isolation of the promoter of a root cap expressed pectinmethylesterase gene from Pisum sativum L. (RCPME1) and its use in the study of gene activity. Plant Soil 2004, 265, 47–59. [Google Scholar] [CrossRef]
- Sunitha, S.; Mahajan, N.; Veluthambi, K. The TrAP/REn monodirectional promoter of Mungbean yellow mosaic geminivirus (MYMV) displays root-specific expression in transgenic tobacco. Plant Cell Tissue Organ Cult. 2012, 109, 535–545. [Google Scholar] [CrossRef]
- Kagaya, Y.; Ohmiya, K.; Hattori, T. RAV1, a novel DNA-binding protein, binds to bipartite recognition sequence through two distinct DNA-binding domains uniquely found in higher plants. Nucleic Acids Res. 1999, 27, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Simon, L.; Lalonde, M. DNA amplification from vegetative and sexual tissues of trees using polymerase chain reaction. Can. J. For. Res. 1990, 20, 254–257. [Google Scholar] [CrossRef]
- TIANquick Midi Purification Kit. Available online: http://www.tiangen.com/?productShow/t1/1/id/48.html (accessed on 1 March 2015).
- PLACE. Available online: http://www.dna.affrc.go.jp/PLACE/ (accessed on 1 March 2015).
- PlantCARE. Available online: http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ (accessed on 1 March 2015).
- Chen, H.; Nelson, R.; Sherwood, J. Enhanced recovery of transformants of Agrobacterium tumefaciens after freeze-thaw transformation and drug selection. Biotechniques 1994, 16, 664–668. [Google Scholar] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A. Plant reporter genes: The GUS gene fusion system. Genet. Eng. 1988, 10, 247–263. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Wang, F.; Yu, G.; Zhang, X.; Jia, C.; Qin, J.; Pan, H. Functional Analysis of the Maize C-Repeat/DRE Motif-Binding Transcription Factor CBF3 Promoter in Response to Abiotic Stress. Int. J. Mol. Sci. 2015, 16, 12131-12146. https://doi.org/10.3390/ijms160612131
Liu J, Wang F, Yu G, Zhang X, Jia C, Qin J, Pan H. Functional Analysis of the Maize C-Repeat/DRE Motif-Binding Transcription Factor CBF3 Promoter in Response to Abiotic Stress. International Journal of Molecular Sciences. 2015; 16(6):12131-12146. https://doi.org/10.3390/ijms160612131
Chicago/Turabian StyleLiu, Jinliang, Fengting Wang, Gang Yu, Xianghui Zhang, Chengguo Jia, Jianchun Qin, and Hongyu Pan. 2015. "Functional Analysis of the Maize C-Repeat/DRE Motif-Binding Transcription Factor CBF3 Promoter in Response to Abiotic Stress" International Journal of Molecular Sciences 16, no. 6: 12131-12146. https://doi.org/10.3390/ijms160612131