Time-Course Changes of Steroidogenic Gene Expression and Steroidogenesis of Rat Leydig Cells after Acute Immobilization Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
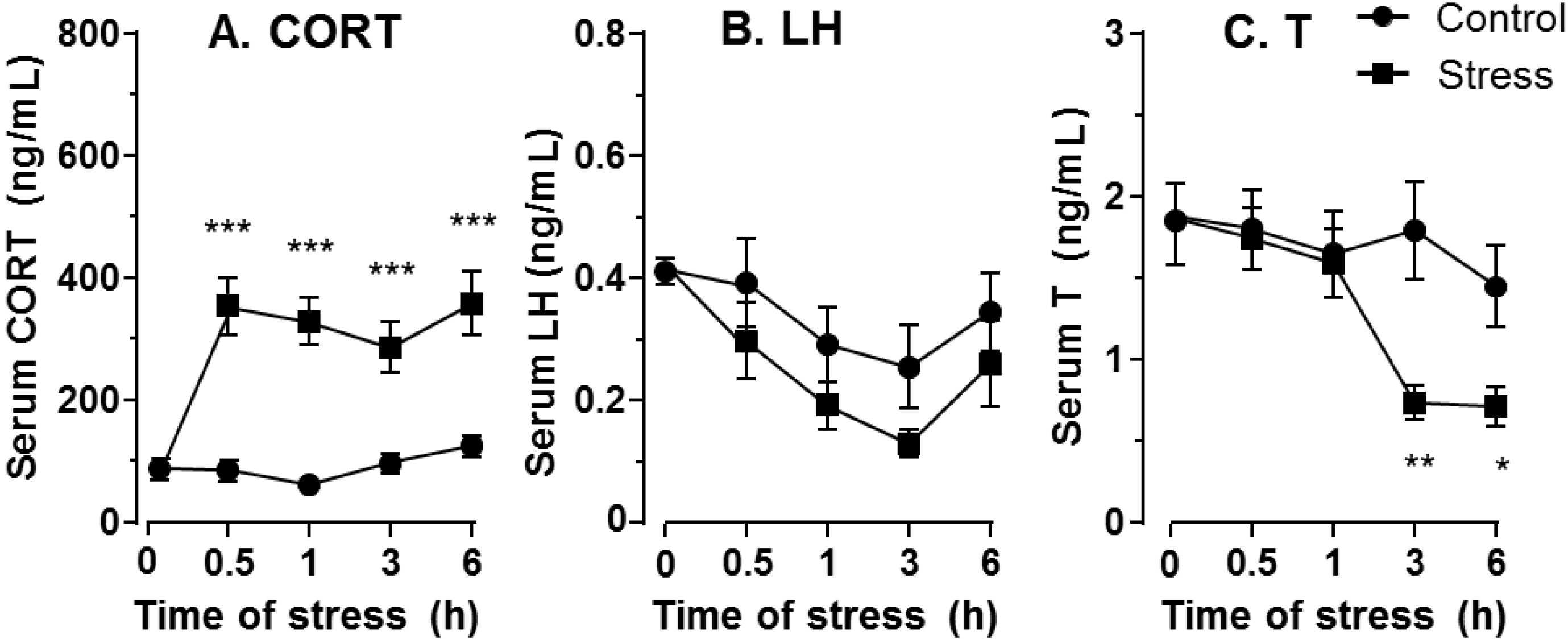
2.1. Plasma Hormone Levels


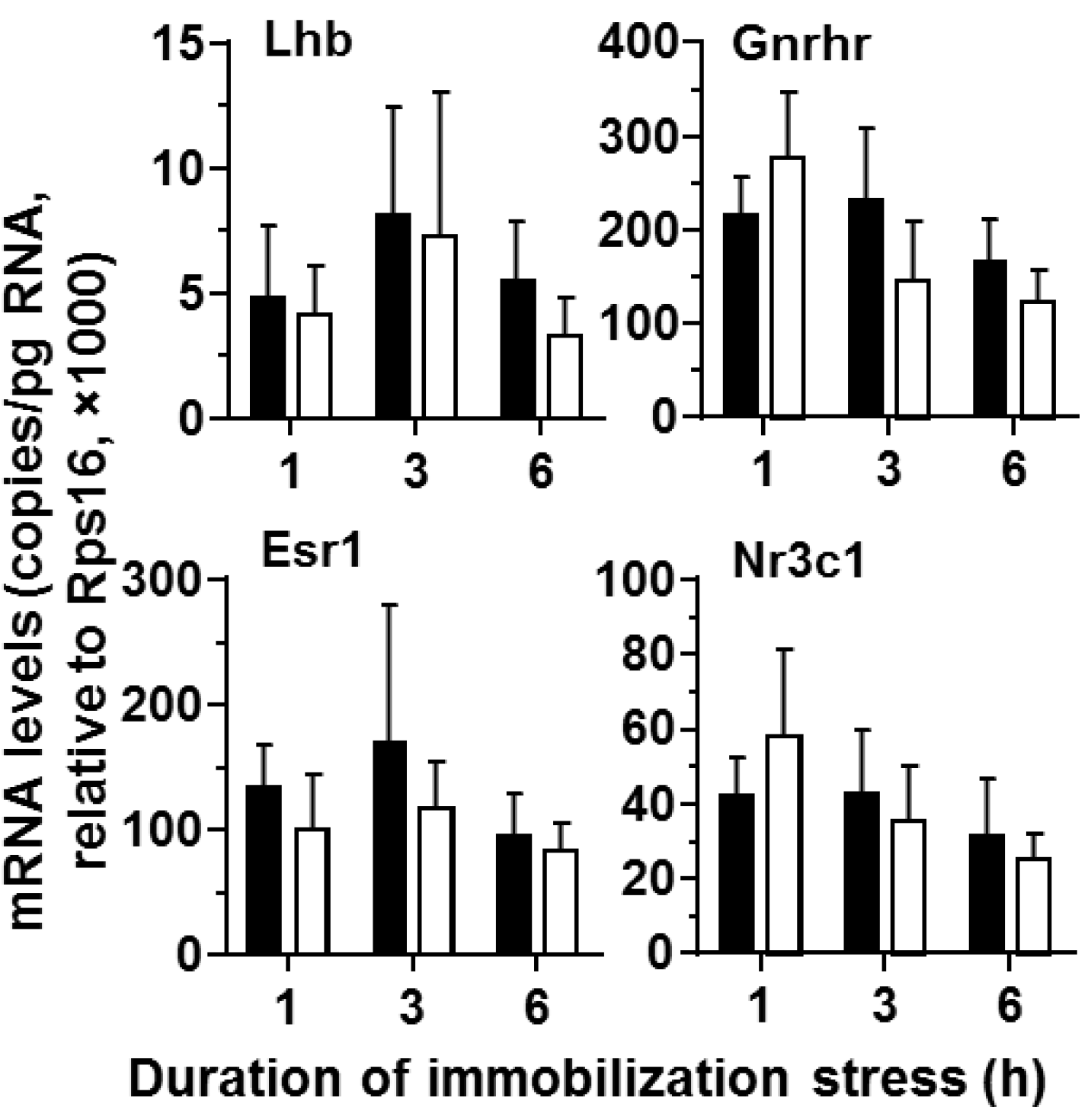
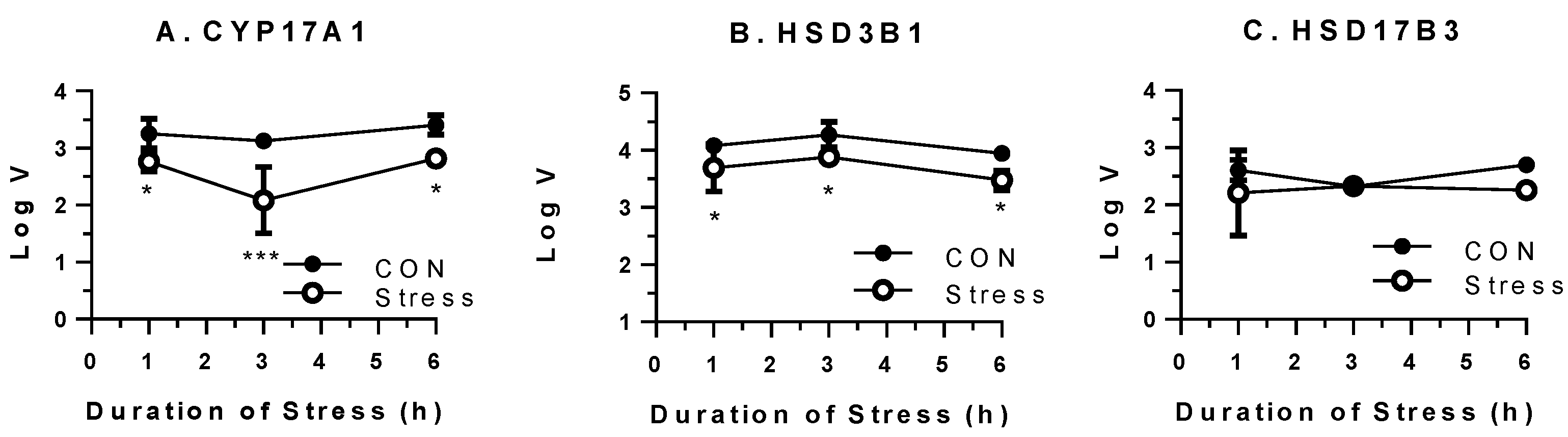
2.2. Gene Expression Levels of Pituitary and Testes


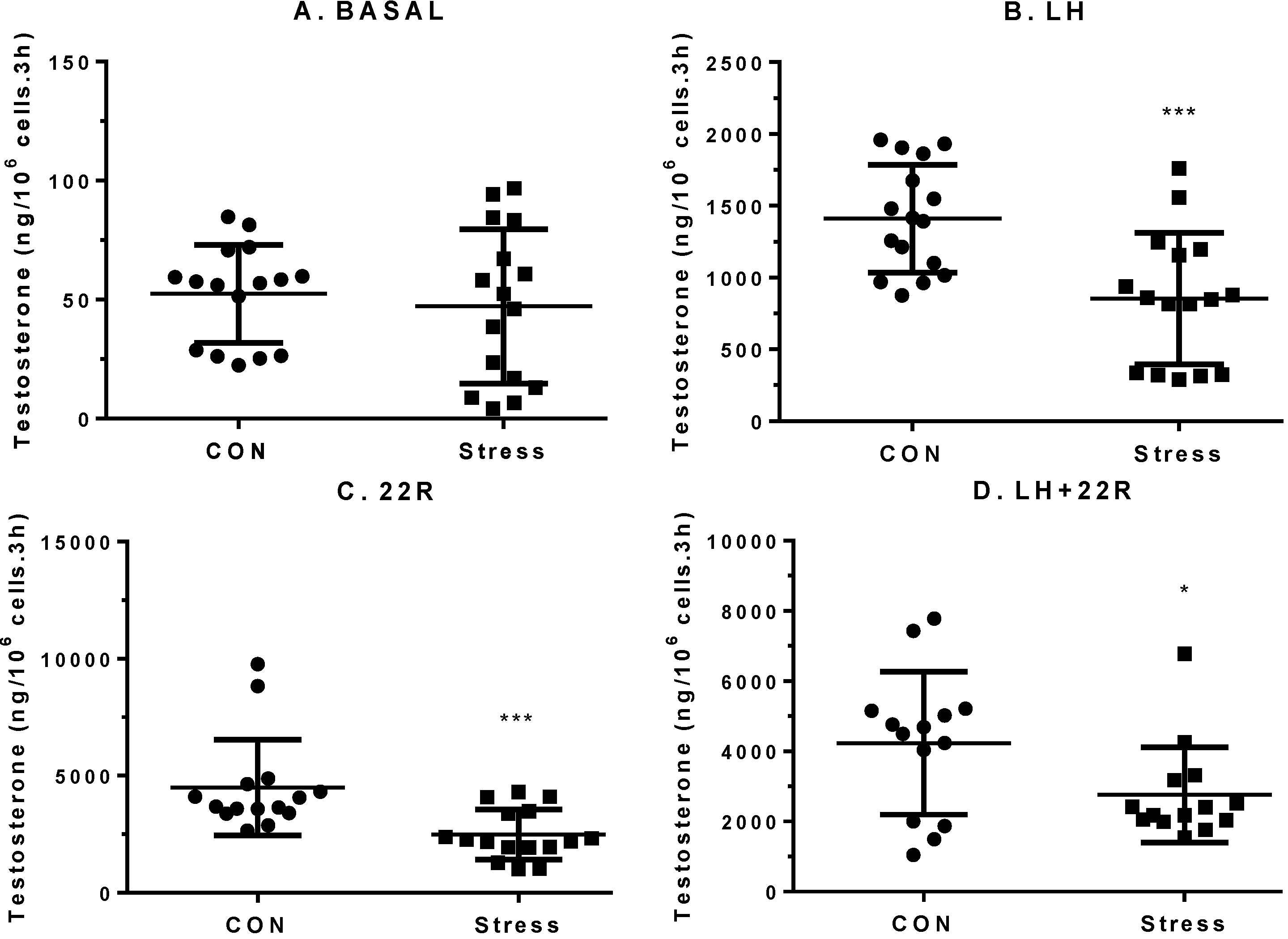
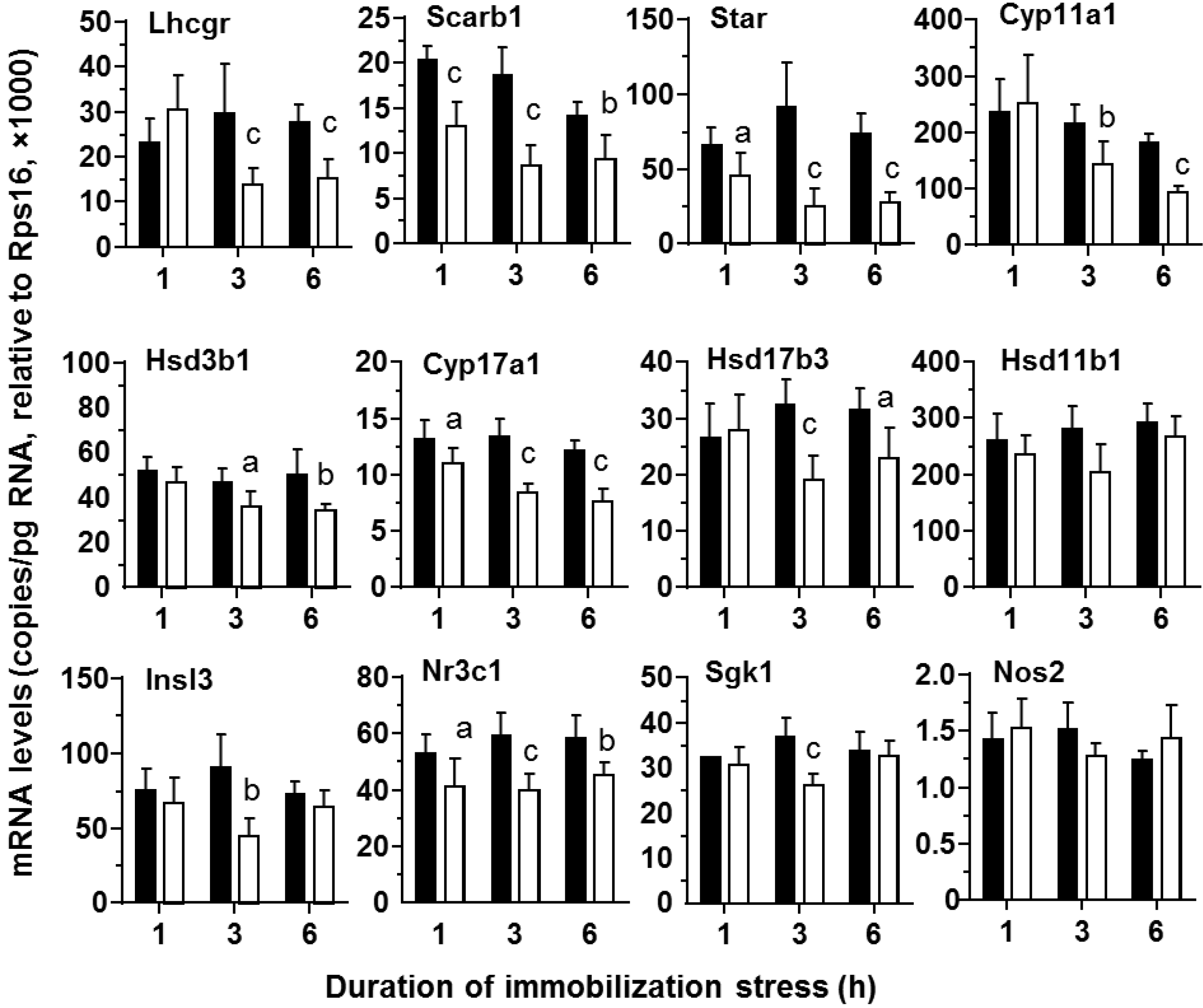
2.3. Levels of StAR Protein and Steroidogenic Enzymes


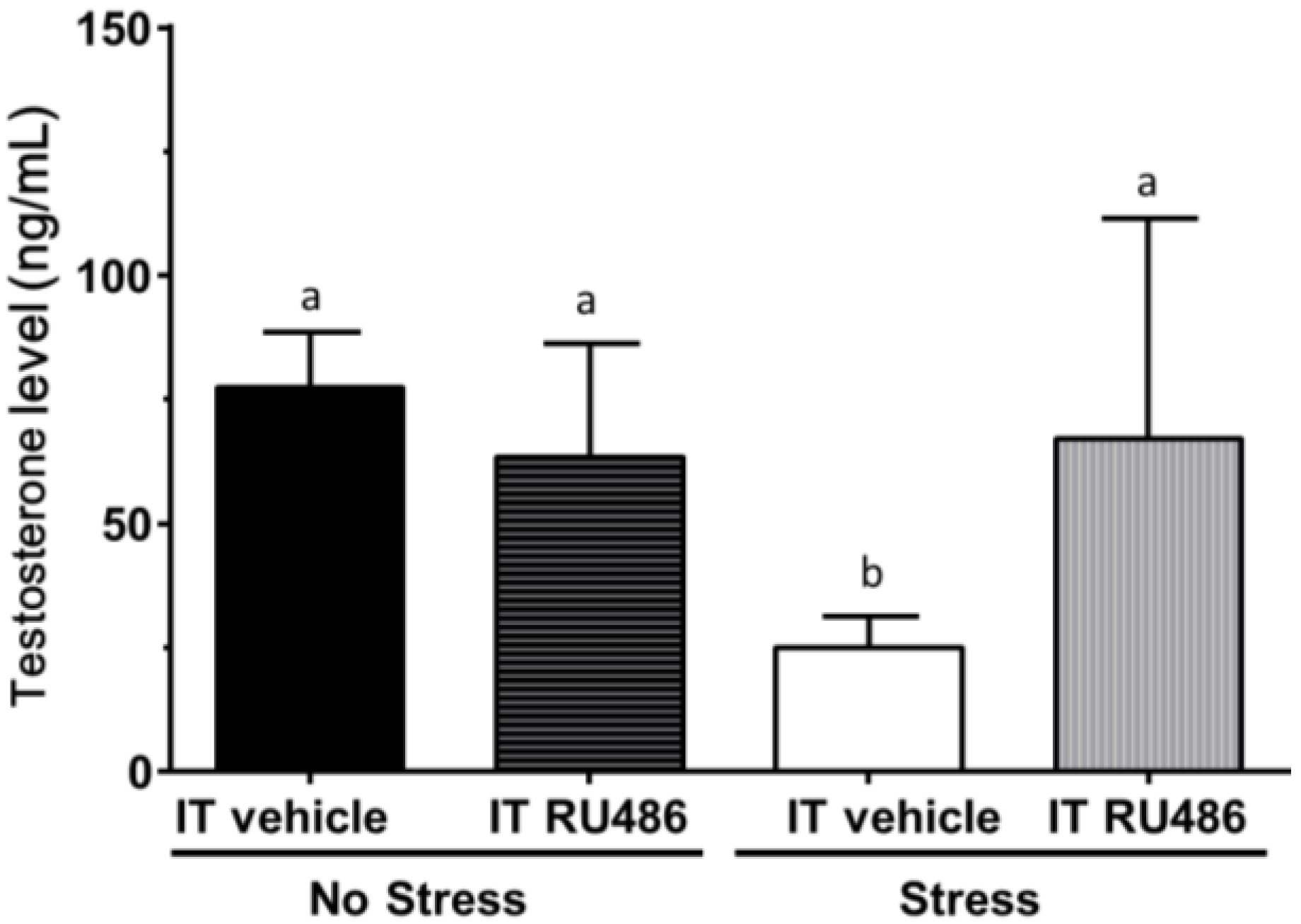
2.4. Antagonism by Glucocorticoid Receptor Antagonist RU486

3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Rat Stress Model: Time Course Experiment
4.3.1. Experiment 1
4.3.2. Experiment 2
4.4. Leydig Cell Isolation and Leydig Cell Steroidogenesis Ex Vivo
4.5. RIA of CORT, LH and Testosterone
4.6. Real-Time PCR (qPCR)
4.7. Analysis of Steroidogenic Enzyme Activities
4.8. Western Blot Analysis of StAR
4.9. Statistics
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stocco, D.M. Tracking the role of a StAR in the sky of the new millenium. Mol. Endocrinol. 2001, 15, 1245–1254. [Google Scholar] [PubMed]
- Ge, R.S.; Hardy, M.P. Regulation of Leydig cells during pubertal development. In The Leydig Cell in Health and Disease; Payne, A.H., Hardy, M.P., Eds.; Humana Press: Totowa, NJ, USA, 2007; pp. 55–70. [Google Scholar]
- Veldhuis, J.D.; Dufau, M.L. Steroidal regulation of biologically active luteinizing hormone secretion in men and women. Hum. Reprod. 1993, 8, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Ge, R.S.; Hardy, D.O.; Catterall, J.F.; Hardy, M.P. Developmental changes in glucocorticoid receptor and 11β-hydroxysteroid dehydrogenase oxidative and reductive activities in rat Leydig cells. Endocrinology 1997, 138, 5089–5095. [Google Scholar] [PubMed]
- Schultz, R.; Isola, J.; Parvinen, M.; Honkaniemi, J.; Wikstrom, A.C.; Gustafsson, J.A.; Pelto-Huikko, M. Localization of the glucocorticoid receptor in testis and accessory sexual organs of male rat. Mol. Cell Endocrinol. 1993, 95, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Macho, L.; Kvetnansky, R.; Fickova, M.; Popova, I.A.; Grigoriev, A. Effects of exposure to space flight on endocrine regulations in experimental animals. Endocr. Regul. 2001, 35, 101–114. [Google Scholar] [PubMed]
- Chrousos, G.P.; Gold, P.W. The concepts of stress and stress system disorders. Overview of physical and behavioral homeostasis. JAMA 1992, 267, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Munck, A.; Guyre, P.M.; Holbrook, N.J. Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocrinol. Rev. 1984, 5, 25–43. [Google Scholar] [CrossRef]
- Hardy, M.P.; Gao, H.B.; Dong, Q.; Ge, R.; Wang, Q.; Chai, W.R.; Feng, X.; Sottas, C. Stress hormone and male reproductive function. Cell. Tissue Res. 2005, 322, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Stojkov, N.J.; Janjic, M.M.; Baburski, A.Z.; Mihajlovic, A.I.; Drljaca, D.M.; Sokanovic, S.J.; Bjelic, M.M.; Kostic, T.S.; Andric, S.A. Sustained in vivo blockade of α1-adrenergic receptors prevented some of stress-triggered effects on steroidogenic machinery in Leydig cells. Am. J. Physiol. Endocrinol. Metab 2013, 305, E194–E204. [Google Scholar] [CrossRef] [PubMed]
- Stojkov, N.J.; Janjic, M.M.; Bjelic, M.M.; Mihajlovic, A.I.; Kostic, T.S.; Andric, S.A. Repeated immobilization stress disturbed steroidogenic machinery and stimulated the expression of cAMP signaling elements and adrenergic receptors in Leydig cells. Am. J. Physiol. Endocrinol. Metab 2012, 302, E1239–E1251. [Google Scholar] [CrossRef] [PubMed]
- Hales, D.B.; Payne, A.H. Glucocorticoid-mediated repression of P450scc mRNA and de novo synthesis in cultured Leydig cells. Endocrinology 1989, 124, 2099–2104. [Google Scholar] [CrossRef] [PubMed]
- Orr, T.E.; Taylor, M.F.; Bhattacharyya, A.K.; Collins, D.C.; Mann, D.R. Acute immobilization stress disrupts testicular steroidogenesis in adult male rats by inhibiting the activities of 17α-hydroxylase and 17,20-lyase without affecting the binding of LH/hCG receptors. J. Androl. 1994, 15, 302–308. [Google Scholar] [PubMed]
- Ge, R.S.; Dong, Q.; Sottas, C.M.; Chen, H.; Zirkin, B.R.; Hardy, M.P. Gene expression in rat Leydig cells during development from the progenitor to adult stage: A cluster analysis. Biol. Reprod. 2005, 72, 1405–1415. [Google Scholar] [CrossRef] [PubMed]
- Ge, R.S.; Gao, H.B.; Nacharaju, V.L.; Gunsalus, G.L.; Hardy, M.P. Identification of a kinetically distinct activity of 11β-hydroxysteroid dehydrogenase in rat Leydig cells. Endocrinology 1997, 138, 2435–2442. [Google Scholar] [PubMed]
- Ge, R.S.; Dong, Q.; Niu, E.M.; Sottas, C.M.; Hardy, D.O.; Catterall, J.F.; Latif, S.A.; Morris, D.J.; Hardy, M.P. 11β-Hydroxysteroid dehydrogenase 2 in rat Leydig cells: Its role in blunting glucocorticoid action at physiological levels of substrate. Endocrinology 2005, 146, 2657–2664. [Google Scholar] [CrossRef] [PubMed]
- Fenster, L.; Katz, D.F.; Wyrobek, A.J.; Pieper, C.; Rempel, D.M.; Oman, D.; Swan, S.H. Effects of psychological stress on human semen quality. J. Androl. 1997, 18, 194–202. [Google Scholar] [PubMed]
- Hjollund, N.H.; Bonde, J.P.; Henriksen, T.B.; Giwercman, A.; Olsen, J. Job strain and male fertility. Epidemiology 2004, 15, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Hjollund, N.H.; Bonde, J.P.; Henriksen, T.B.; Giwercman, A.; Olsen, J. Reproductive effects of male psychologic stress. Epidemiology 2004, 15, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Eskiocak, S.; Gozen, A.S.; Yapar, S.B.; Tavas, F.; Kilic, A.S.; Eskiocak, M. Glutathione and free sulphydryl content of seminal plasma in healthy medical students during and after exam stress. Hum. Reprod. 2005, 20, 2595–2600. [Google Scholar] [CrossRef] [PubMed]
- Campagne, D.M. Should fertilization treatment start with reducing stress? Hum. Reprod. 2006, 21, 1651–1658. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.B.; Tong, M.H.; Hu, Y.Q.; Guo, Q.S.; Ge, R.; Hardy, M.P. Glucocorticoid induces apoptosis in rat Leydig cells. Endocrinology 2002, 143, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, Q.; Wang, F.F.; Gao, H.B.; Zhang, P. Stress induces glucocorticoid-mediated apoptosis of rat Leydig cells in vivo. Stress 2012, 15, 74–84. [Google Scholar] [PubMed]
- Spencer, R.L.; Miller, A.H.; Moday, H.; McEwen, B.S.; Blanchard, R.J.; Blanchard, D.C.; Sakai, R.R. Chronic social stress produces reductions in available splenic type II corticosteroid receptor binding and plasma corticosteroid binding globulin levels. Psychoneuroendocrinology 1996, 21, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, D.C.; Spencer, R.L.; Weiss, S.M.; Blanchard, R.J.; McEwen, B.; Sakai, R.R. Visible burrow system as a model of chronic social stress: Behavioral and neuroendocrine correlates. Psychoneuroendocrinology 1995, 20, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Ge, R.S.; Hardy, M.P. Initial predominance of the oxidative activity of type I 11β-hydroxysteroid dehydrogenase in primary rat Leydig cells and transfected cell lines. J. Androl. 2000, 21, 303–310. [Google Scholar] [PubMed]
- Monder, C.; Miroff, Y.; Marandici, A.; Hardy, M.P. 11β-Hydroxysteroid dehydrogenase alleviates glucocorticoid-mediated inhibition of steroidogenesis in rat Leydig cells. Endocrinology 1994, 134, 1199–1204. [Google Scholar] [PubMed]
- Martin, L.J.; Tremblay, J.J. Glucocorticoids antagonize cAMP-induced StAR transcription in Leydig cells through the orphan nuclear receptor NR4A1. J. Mol. Endocrinol. 2008, 41, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.C.; Huang, Y.D.; Hardy, D.O.; Li, X.K.; Ge, R.S. Glucocorticoid suppresses steroidogenesis in rat progenitor Leydig cells. J. Androl. 2010, 31, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Stocco, D.M.; Clark, B.J. Regulation of the acute production of steroids in steroidogenic cells. Endocr. Rev. 1996, 17, 221–244. [Google Scholar] [PubMed]
- Dong, Q.; Salva, A.; Sottas, C. M.; Niu, E.; Holmes, M.; Hardy, M.P. Rapid glucocorticoid mediation of suppressed testosterone biosynthesis in male mice subjected to immobilization stress. J. Androl. 2004, 25, 973–981. [Google Scholar] [PubMed]
- Gao, H.B.; Shan, L.X.; Monder, C.; Hardy, M.P. Suppression of endogenous corticosterone levels in vivo increases the steroidogenic capacity of purified rat Leydig cells in vitro. Endocrinology 1996, 137, 1714–1718. [Google Scholar] [PubMed]
- Monder, C.; Hardy, M.P.; Blanchard, R.J.; Blanchard, D.C. Comparative aspects of 11β-hydroxysteroid dehydrogenase. Testicular 11β-hydroxysteroid dehydrogenase: Development of a model for the mediation of Leydig cell function by corticosteroids. Steroids 1994, 59, 69–73. [Google Scholar] [CrossRef]
- Orr, T.E.; Mann, D.R. Effects of restraint stress on plasma LH and testosterone concentrations, Leydig cell LH/hCG receptors, and in vitro testicular steroidogenesis in adult rats. Horm. Behav. 1990, 24, 324–341. [Google Scholar] [CrossRef] [PubMed]
- Akibami, M.A.; Mann, D.R. Mechanism of stress-induced attenuation of the testicular response to gonadotropin: Possible involvement of testicular opioids, a pertussis toxin-sensitive G-protein, and phosphodiesterase. J. Androl. 1996, 17, 10–16. [Google Scholar] [PubMed]
- Kostic, T.; Andric, S.; Kovacevic, R.; Maric, D. The involvement of nitric oxide in stress-impaired testicular steroidogenesis. Eur. J. Pharmacol. 1998, 346, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Andric, S.A.; Janjic, M.M.; Stojkov, N.J.; Kostic, T.S. Testosterone-induced modulation of nitric oxide-cGMP signaling pathway and androgenesis in the rat Leydig cells. Biol. Reprod. 2010, 83, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Weissman, B.A.; Sottas, C.M.; Zhou, P.; Iadecola, C.; Hardy, M.P. Testosterone production in mice lacking inducible nitric oxide synthase expression is sensitive to restraint stress. Am. J. Physiol. Endocrinol. Metab 2007, 292, E615–E620. [Google Scholar] [CrossRef] [PubMed]
- Weissman, B.A.; Sottas, C.M.; Holmes, M.; Zhou, P.; Iadecola, C.; Hardy, D.; Ge, R.S.; Hardy, M.P. Normal responses to restraint stress in mice lacking the gene for neuronal nitric oxide synthase. J. Androl. 2009, 30, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.C.; Chen, Y.; Lang, F. Glucocorticoid activation of Na+/H+ exchanger isoform 3 revisited. the roles of SGK1 and NHERF2. J. Biol. Chem. 2002, 277, 7676–7683. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, S.; Sharpe, R.M. Interstitial fluid volume in the rat testis: Androgen-dependent regulation by the seminiferous tubules? J. Endocrinol. 1989, 120, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Klinefelter, G.R.; Hall, P.F.; Ewing, L.L. Effect of luteinizing hormone deprivation in situ on steroidogenesis of rat Leydig cells purified by a multi-step procedure. Biol. Reprod. 1987, 36, 769–783. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.H.; Downing, J.R.; Wong, K.L. Luteinizing hormone receptor and testosterone synthesis in two distinct populations of Leydig cells. Endocrinology 1980, 106, 1424–1429. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Didolkar, A.K.; Monder, C.; Bardin, C.W.; Sundaram, K. The biological activity of 7α-methyl-19-nortestosterone is not amplified in male reproductive tract as is that of testosterone. Endocrinology 1992, 130, 3677–3683. [Google Scholar] [PubMed]
- Cochran, R.C.; Ewing, L.L.; Niswender, G.D. Serum levels of follicle-stimulating hormone, prolactin, testosterone, 5α-dihydrotestosterone, 5α-androstane-3α, 17β-diol, 5α-androstane-3β, 17b-diol and 17β-estradiol from male beagles with spontaneous or induced benign prostatic hyperplasia. Invest. Urol. 1981, 19, 142–147. [Google Scholar] [PubMed]
- Lin, H.; Ge, R.S.; Chen, G.R.; Hu, G.X.; Dong, L.; Lian, Q.Q.; Hardy, D.O.; Sottas, C.M.; Li, X.K.; Hardy, M.P. Involvement of testicular growth factors in fetal Leydig cell aggregation after exposure to phthalate in utero. Proc. Nat. Acad. Sci. USA 2008, 105, 7218–7222. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhou, H.; Su, Z.; Chen, B.; Wang, G.; Wang, C. Q.; Xu, Y.; Ge, R.S. Comparison of cell types in the rat Leydig cell lineage after ethane dimethanesulfonate treatment. Reproduction 2013, 145, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Yuan, K.M.; Liang, Y.; Chu, Y.H.; Lian, Q.Q.; Ge, Y.F.; Zhen, W.; Sottas, C.M.; Su, Z.J.; Ge, R.S. Alterations of gene profiles in Leydig-cell-regenerating adult rat testis after ethane dimethane sulfonate-treatment. Asian J. Androl. 2014. [Google Scholar] [CrossRef]
- Huyghe, S.; Schmalbruch, H.; de Gendt, K.; Verhoeven, G.; Guillou, F.; van Veldhoven, P.P.; Baes, M. Peroxisomal multifunctional protein 2 is essential for lipid homeostasis in Sertoli cells and male fertility in mice. Endocrinology 2006, 147, 2228–2236. [Google Scholar] [CrossRef] [PubMed]
- Corton, J.C.; Bocos, C.; Moreno, E.S.; Merritt, A.; Marsman, D.S.; Sausen, P.J.; Cattley, R.C.; Gustafsson, J.A. Rat 17β-hydroxysteroid dehydrogenase type IV is a novel peroxisome proliferator-inducible gene. Mol. Pharmacol. 1996, 50, 1157–1166. [Google Scholar] [PubMed]
- Hu, G.X.; Zhou, H.Y.; Li, X.W.; Chen, B.B.; Xiao, Y.C.; Lian, Q.Q.; Liang, G.; Kim, H.H.; Zheng, Z.Q.; Hardy, D.O.; Ge, R.S. The (+)- and (−)-gossypols potently inhibit both 3β-hydroxysteroid dehydrogenase and 17β-hydroxysteroid dehydrogenase 3 in human and rat testes. J. Steroid Biochem. Mol. Biol. 2009, 115, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, X.; Zhu, Q.; Chen, D.; Guo, J.; Yao, W.; Dong, Y.; Wei, J.; Lian, Q.; Ge, R.S.; Yuan, B. Disrupting androgen production of Leydig cells by resveratrol via direct inhibition of human and rat 3β-hydroxysteroid dehydrogenase. Toxicol. Lett. 2014, 226, 14–19. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, H.; Yuan, K.-m.; Zhou, H.-y.; Bu, T.; Su, H.; Liu, S.; Zhu, Q.; Wang, Y.; Hu, Y.; Shan, Y.; et al. Time-Course Changes of Steroidogenic Gene Expression and Steroidogenesis of Rat Leydig Cells after Acute Immobilization Stress. Int. J. Mol. Sci. 2014, 15, 21028-21044. https://doi.org/10.3390/ijms151121028
Lin H, Yuan K-m, Zhou H-y, Bu T, Su H, Liu S, Zhu Q, Wang Y, Hu Y, Shan Y, et al. Time-Course Changes of Steroidogenic Gene Expression and Steroidogenesis of Rat Leydig Cells after Acute Immobilization Stress. International Journal of Molecular Sciences. 2014; 15(11):21028-21044. https://doi.org/10.3390/ijms151121028
Chicago/Turabian StyleLin, Han, Kai-ming Yuan, Hong-yu Zhou, Tiao Bu, Huina Su, Shiwen Liu, Qiqi Zhu, Yiyan Wang, Yuanyuan Hu, Yuanyuan Shan, and et al. 2014. "Time-Course Changes of Steroidogenic Gene Expression and Steroidogenesis of Rat Leydig Cells after Acute Immobilization Stress" International Journal of Molecular Sciences 15, no. 11: 21028-21044. https://doi.org/10.3390/ijms151121028