Protein Profiles Reveal Diverse Responsive Signaling Pathways in Kernels of Two Maize Inbred Lines with Contrasting Drought Sensitivity

Abstract
:1. Introduction
2. Results and Discussion
2.1. Maize Kernel Proteome Patterns in Response to Drought Stress Treatments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GI a | Protein Name b | Coverage (%) c /Peptide Fragments d | Fold Change (B73) e | Fold Change (Lo964) e | ||
|---|---|---|---|---|---|---|
| Cell rescue/defense | ||||||
| 219363419 | Dehydrin | 23.0/6 | 1.64 ± 0.09 | / | ||
| 295856 | RAB-17 | 37.5/10 | 1.36 ± 0.09 | / | ||
| 226491145 | Late embryogenesis abundant protein Lea14-A | 74.3/17 | 1.46 ± 0.15 | / | ||
| 195659191 | Embryonic abundant protein 1 | 84.6/25 | 1.47 ± 0.15 | −1.48 ± 0.08 | ||
| 226499304 | Pathogenesis-related protein 10 | 11.2/2 | 1.27 ± 0.08 | −1.30 ± 0.03 | ||
| 226531123 | Glycine-rich protein 2b | 44.2/3 | 1.61 ± 0.20 | 1.42 ± 0.09 | ||
| 226492587 | Stress-inducible membrane pore protein | 63.2/15 | 1.32 ± 0.07 | / | ||
| 226496775 | Xylanase inhibitor protein 1 | 24.5/3 | 1.63 ± 0.04 | −1.31 ± 0.02 | ||
| 293332305 | Serpin-ZXA-like | 13.6/5 | 1.30 ± 0.05 | / | ||
| 293335211 | Heavy-metal-associated Domain-containing protein | 11.9/2 | 1.54 ± 0.52 | / | ||
| 473187 | Protein kinase C inhibitor | 24.2/2 | −1.46 ± 0.04 | / | ||
| 75994217 | Hageman factor inhibitor | 18.7/2 | −1.45 ± 0.03 | / | ||
| Redox homeostasis | ||||||
| 257333334 | Glutathione transferase30 | 21.2/8 | 1.40 ± 0.14 | −1.42 ± 0.02 | ||
| 48374955 | Glutathione peroxidase | 44.0/8 | 1.33 ± 0.08 | / | ||
| 226494622 | Glutathione S-transferase,-like protein | 19.8/3 | 1.44 ± 0.06 | / | ||
| 257728955 | Grx_C2.2—glutaredoxin subgroup I | 69.9/8 | 1.26 ± 0.10 | 1.34 ± 0.09 | ||
| 87133468 | 1-Cys peroxiredoxin antioxidant | 76.4/57 | 1.32 ± 0.08 | / | ||
| 195652835 | Thioredoxin H-type | 61.5/9 | 1.33 ± 0.09 | / | ||
| 7548002 | Mn-superoxide dismutase | 47.6/18 | 1.59 ± 0.18 | / | ||
| 6018746 | Superoxide dismutase-4A | 46.7/6 | 1.28 ± 0.06 | / | ||
| 238008410 | NADH-ubiquinone oxidoreductase subunit B17.2 | 18.2/2 | 1.53 ± 0.01 | / | ||
| 195655511 | NAD dependent epimerase/dehydratase | 24.3/2 | 1.64 ± 0.05 | 1.35 ± 0.07 | ||
| Hormone response | ||||||
| 226497210 | ABA-responsive protein | 23.5/5 | 1.46 ± 0.04 | / | ||
| 226498678 | Ethylene-responsive protein | 28.7/6 | 1.50 ± 0.19 | / | ||
| 226508662 | Ethylene-responsive protein | 27.2/7 | 1.49 ± 0.15 | / | ||
| 301069326 | Auxin response factor 23 | 9.56/1 | 1.52 ± 0.19 | 1.35 ± 0.09 | ||
| 224028639 | Jasmonate-induced protein | 26.8/2 | −1.54 ± 0.008 | −1.31 ± 0.17 | ||
| Signal transduction | ||||||
| 229611800 | GTP-binding nuclear protein Ran-A1 | 40.1/11 | −1.35 ± 0.04 | −1.20 ± 0.01 | ||
| 226528736 | ATP/GTP binding protein | 16.1/4 | 1.34 ± 0.08 | / | ||
| 255037841 | Histidine kinase | 7.52/2 | −1.51 ± 0.02 | 1.28 ± 0.11 | ||
| 226493048 | Nucleoside diphosphate kinase 4 | 23.5/4 | 1.35 ± 0.09 | / | ||
| 254256262 | Pyruvate orthophosphate dikinase 1 | 23.0/12 | −2.11 ± 0.04 | −1.43 ± 0.05 | ||
| Storage proteins | ||||||
| 228310 | Globulin 2 | 77.6/156 | / | −1.51 ± 0.12 | ||
| 224030527 | Globulin 1 | 70.5/205 | 1.41 ± 0.10 | / | ||
| 195658011 | Globulin-1 S allele precursor | 77.6/152 | 1.63 ± 0.22 | / | ||
| 226500532 | Seed maturation protein PM41 | 18.0/9 | 1.42 ± 0.08 | / | ||
| 22284 | Vicilin-like embryo storage protein | 69.8/180 | / | 1.46 ± 0.001 | ||
| 330732090 | γ-zein | 2.27/1 | −1.67 ± 0.01 | −1.13 ± 0.006 | ||
| Protein biosynthesis | ||||||
| 9931636 | Ribosomal protein s6 RPS6-2 | 23.1/5 | 1.65 ± 0.26 | / | ||
| 257667240 | 40S ribosomal protein S18 | 54.6/7 | 1.52 ± 0.09 | / | ||
| 6226702 | 40S ribosomal protein S8 | 33.5/7 | 1.81 ± 0.10 | / | ||
| 226502084 | 60S ribosomal protein L7a | 19.8/5 | 1.50 ± 0.14 | / | ||
| 258598734 | QM-like protein | 23.7/4 | 1.36 ± 0.08 | / | ||
| Protein folding and assembly | ||||||
| 7546186 | Heat shock protein HSP82 | 23.1/9 | −1.65 ± 0.03 | / | ||
| 293331695 | HSP protein | 23.3/10 | / | −1.51 ± 0.04 | ||
| 54299342 | Mitochondrial small heat shock protein 22 | 44.0/7 | / | −1.39 ± 0.02 | ||
| 293335765 | TCP-1/cpn60 chaperonin family protein | 8.94/2 | / | −1.34 ± 0.09 | ||
| 453670 | Heat shock protein 26 | 57.5/9 | / | −1.44 ± 0.07 | ||
| 257745378 | Peptidyl-prolyl cis-trans isomerase Family protein | 19.5/3 | / | 1.29 ± 0.02 | ||
| 226495869 | Histone deacetylase 6 | 6.49/2 | 1.45 ± 0.13 | −1.91 ± 0.09 | ||
| 59861271 | Protein disulfide isomerase | 6.56/2 | 1.90 ± 0.29 | / | ||
| Protein degradation | ||||||
| 224029787 | Proteasome subunit α type | 49.2/8 | 1.40 ± 0.12 | −1.26 ± 0.04 | ||
| 226531007 | Proteasome subunit α type 1 | 22.3/7 | 1.32 ± 0.08 | −1.17 ± 0.02 | ||
| Carbohydrate metabolism | ||||||
| 22488 | Sucrose synthase | 22.8/10 | −1.73 ± 0.03 | −1.42 ± 0.06 | ||
| 3342802 | Cytosolic 6-phosphogluconate dehydrogenase | 23.6/7 | 1.21 ± 0.04 | −1.39 ± 0.003 | ||
| 226530488 | Glucose-6-phosphate 1-epimerase-like | 11.9/3 | 1.28 ± 0.05 | / | ||
| 194688844 | Sucrose synthase2 | 29.5/11 | 1.61 ± 0.15 | / | ||
| 293333951 | Isocitrate dehydrogenase [NAD] Regulatory subunit 1 | 20.9/4 | / | −1.41 ± 0.14 | ||
| 226504732 | Sorbitol dehydrogenase homolog1 | 56.2/27 | 2.08 ± 0.10 | 1.39 ± 0.05 | ||
| 226499336 | Succinate dehydrogenase flavoprotein subunit | 12.9/2 | 1.45 ± 0.14 | / | ||
| 291047790 | Succinate semialdehyde dehydrogenase | 23.1/8 | 1.41 ± 0.12 | / | ||
| 257726331 | Cytokinin-O-glucosyltransferase 2 | 7.54/2 | 1.40 ± 0.04 | / | ||
| Transcription factor | ||||||
| 226496988 | Fibrillarin-2 | 20.5/2 | 1.51 ± 0.13 | −1.29 ± 0.15 | ||
| 308081068 | AP2-EREBP-type transcription factor | 12.9/2 | 1.33 ± 0.06 | / | ||
| Amino acid and lipid metabolism | ||||||
| 227478191 | Argininosuccinate synthase | 12.3/3 | 1.78 ± 0.23 | 1.27 ± 0.08 | ||
| 293332891 | Isovaleryl-CoA dehydrogenase | 13.0/2 | 1.77 ± 0.14 | / | ||
| 195605626 | Oleosin 16 kDa | 41.0/14 | 1.32 ± 0.07 | −1.30 ± 0.003 | ||
| Secondary metabolism | ||||||
| 226500722 | Anthocyanidin 5,3- O-glucosyltransferase | 5.86/2 | 1.23 ± 0.02 | −1.33 ± 0.11 | ||
| 293335591 | Dihydrolipoyl dehydrogenase | 15.0/6 | 1.32 ± 0.11 | / | ||
| 52699545 | Isopentenyl-diphosphate delta isomerase 2 | 8.72/2 | 1.47 ± 0.15 | / | ||
| 596080 | Thiamine biosynthetic enzyme | 16.4/4 | 1.39 ± 0.12 | 1.57 ± 0.23 | ||
| Membrane and transport | ||||||
| 75278333 | Aquaporin TIP1-1 | 6.4/3 | 1.34 ± 0.09 | / | ||
| 75308033 | Aquaporin TIP3-2 | 7.86/2 | 1.41 ± 0.05 | 1.84 ± 0.05 | ||
| 293332063 | Cytochrome b5 | 11.9/2 | −1.41 ± 0.11 | / | ||
| Others and unknown | ||||||
| 226495887 | Adaptin ear-binding coat-associated protein 1 | 12.8/1 | 1.25 ± 0.04 | −1.33 ± 0.17 | ||
| 219363567 | Hypothetical protein LOC100217226 | 9.47/1 | −1.71 ± 0.10 | −1.35 ± 0.06 | ||
| 219363597 | Hypothetical protein LOC100216972 | 12.0/1 | 1.32 ± 0.13 | −1.77 ± 0.17 | ||
| 226528357 | Hypothetical protein LOC100278634 | 34.8/3 | 1.59 ± 0.18 | / | ||
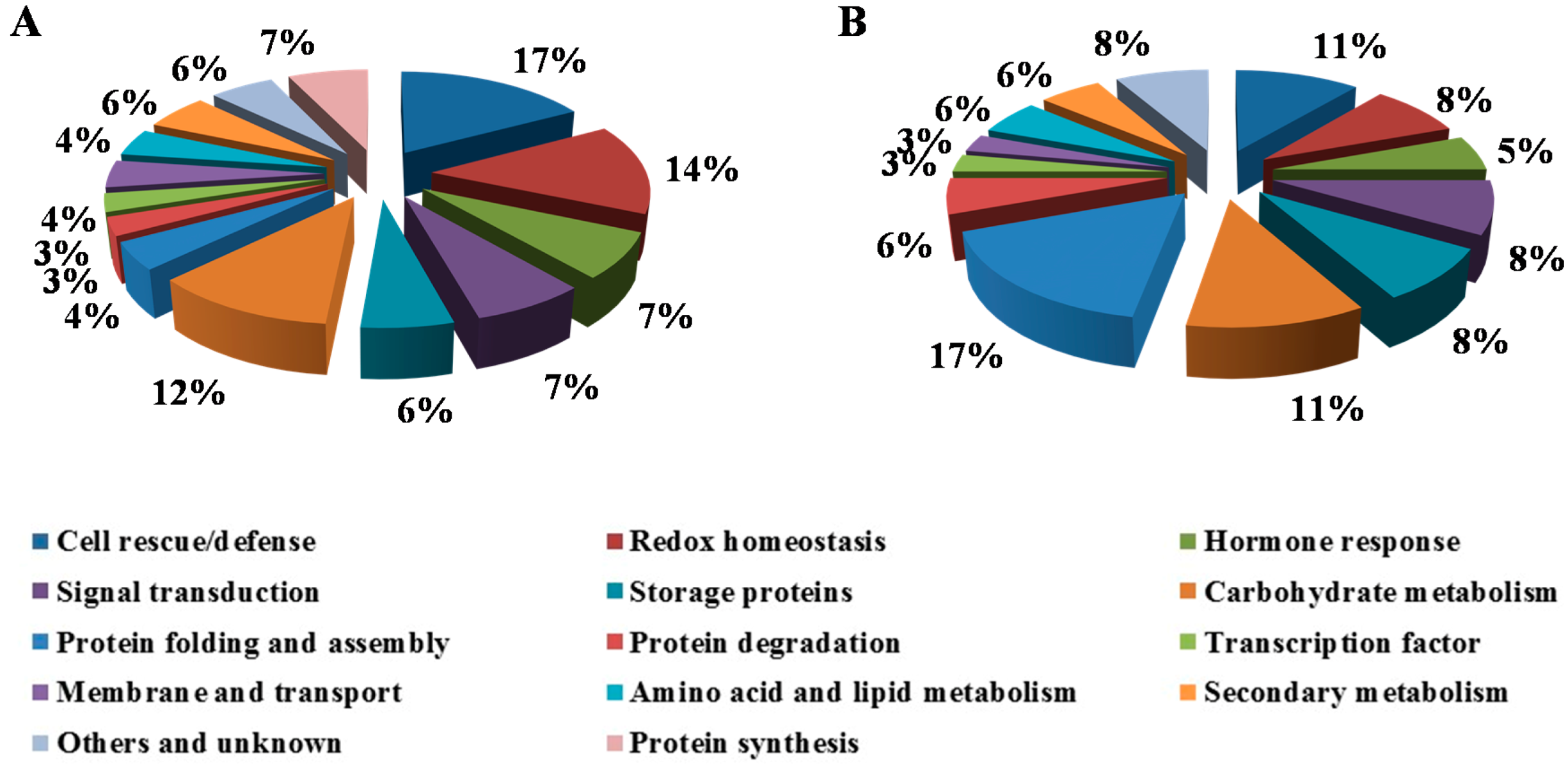
2.2. Functional Annotation and Categories of Differential Expressed Proteins

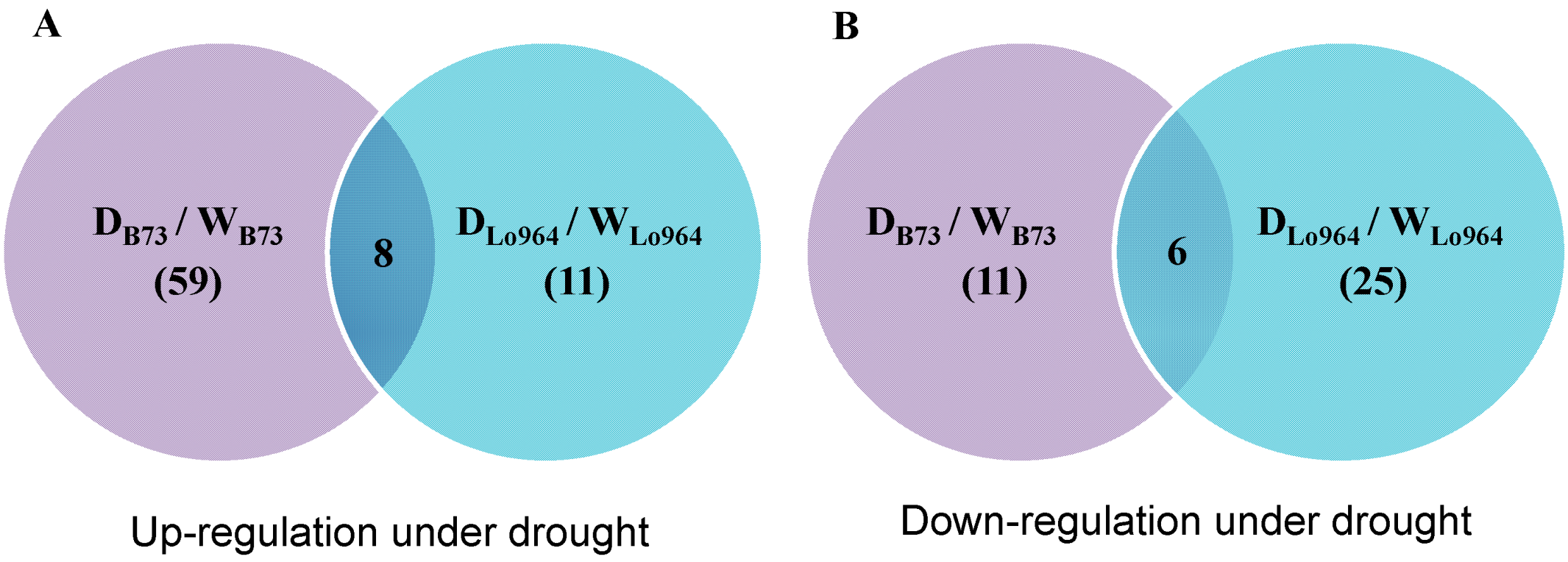
2.3. Overlapping and Specific Proteomic Response to Drought between B73 and Lo964 Lines

2.4. Gene Ontology Analysis of Drought-Responsive Proteins
2.5. Sensitivity to Drought Stress Is Characterized by the Up-Regulation of Antioxidative Proteins

2.6. The Response to Drought Stress May Be Connected to the Differential Regulation of Protein Metabolism
2.7. Differentially Expressed Proteins Implicated in Hormone Response and Signal Transduction Pathways
2.8. Differentially Expressed Proteins Are Involved in Carbohydrate Metabolism and Storage
2.9. Drought Response on Other Functional Groups
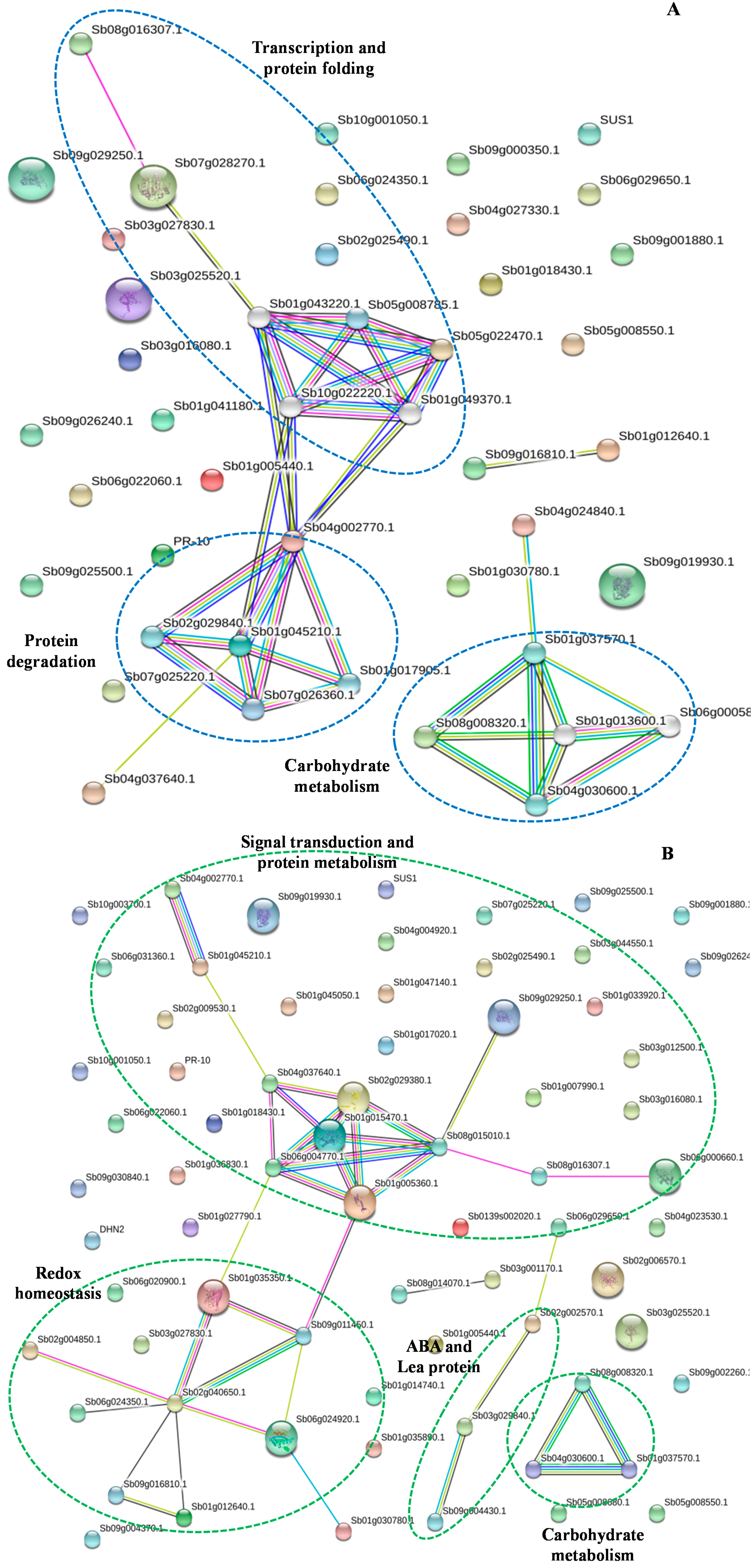
2.10. Protein–Protein Interaction Analysis
2.11. Models for Maize Kernels Tolerance and Sensitivity to Drought Stress


3. Materials and Methods
3.1. Plant Materials and Growth Conditions
3.2. Protein Extraction, Digestion, iTRAQ (Isobaric Tags for Relative and Absolute Quantitation) Labeling
3.3. Analysis of Strong Cation Exchange Fractionation, Reverse Phase Nanoflow HPLC and Tandem Mass Spectrometry
3.4. Protein Identification and Relative Quantification
3.5. Bioinformatics Analysis
4. Conclusions
Supplementary Materials
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Harrison, M.T.; Tardieu, F.; Dong, Z.; Messina, C.D.; Hammer, G.L. Characterizing drought stress and trait influence on maize yield under current and future conditions. Glob. Chang. Biol. 2014, 20, 867–878. [Google Scholar] [CrossRef]
- Edmeades, G.O. Progress in Achieving and Delivering Drought Tolerance in Maize—An Update; ISAAA: Ithaca, NY, USA, 2013. [Google Scholar]
- Payne, G.A. Process of contamination by aflatoxin-producing fungi and their impact on crops. In Mycotoxins in Agriculture and Food Safety; Sinha, K.K., Bhatnagar, D., Eds.; Marcel Dekker: New York, NY, USA, 1998; pp. 279–306. [Google Scholar]
- Guo, B.Z.; Chen, Z.Y.; Lee, R.D.; Scully, B.T. Drought stress and preharvest aflatoxin contamination in agricultural commodity: Genetics, genomics and proteomics. J. Integr. Plant Biol. 2008, 50, 1281–1291. [Google Scholar] [CrossRef]
- Scully, B.T.; Krakowsky, M.D.; Ni, X.Z.; Wilson, J.P.; Lee, R.D.; Guo, B.Z. Preharvest aflatoxin contamination of corn and other grain crops grown on the U.S. Southeastern Coastal Plain. Toxin Rev. 2009, 28, 169–179. [Google Scholar] [CrossRef]
- Fountain, J.C.; Scully, B.T.; Ni, X.; Kemerait, R.C.; Lee, R.D.; Chen, Z.Y.; Guo, B. Environmental influences on maize-Aspergillus flavus interactions and aflatoxin production. Front. Microbiol. 2014, 5, 40. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Brown, R.L.; Cleveland, T.E. Evidence for an association in corn between stress tolerance and resistance to Aspergillus flavus infection and aflatoxin contamination. Afr. J. Biotechnol. 2004, 3, 693–699. [Google Scholar]
- Guo, B.Z.; Holbrook, C.C.; Yu, J.; Lee, R.D.; Lynch, R.E. Application of technology of gene expression in response to drought stress and elimination of preharvest aflatoxin contamination. In Aflatoxin and Food Safety; Abbas, H.K., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 313–331. [Google Scholar]
- Chenu, K.; Chapman, S.C.; Tardieu, F.; McLean, G.; Welcker, C.; Hammer, G.L. Simulating the yield impacts of organ-level quantitative trait loci associated with drought response in maize: A “Gene-to-Phenotype” modeling approach. Genetics 2009, 183, 1507–1523. [Google Scholar] [CrossRef]
- Tsonev, S.; Todorovska, E.; Avramova, V.; Kolev, S.; Abu-Mhadi, N.; Christov, N.K. Genomics assisted improvement of drought tolerance in maize: QTL approaches. Biotechnol. Biotechnol. Equip. 2009, 23, 1410–1413. [Google Scholar] [CrossRef]
- Bolaños, J.; Edmeades, G.O.; Martinez, L. Eight cycles of selection for drought tolerance in lowland tropical maize. III. Responses in drought-adaptive physiological and morphological traits. Field Crops Res. 1993, 31, 269–286. [Google Scholar] [CrossRef]
- Ribaut, J.M.; Ragot, M. Marker-assisted selection to improve drought adaptation in maize: The backcross approach, perspectives, limitations, and alternatives. J. Exp. Bot. 2007, 58, 351–360. [Google Scholar] [CrossRef]
- Benešová, M.; Holá, D.; Fischer, L.; Jedelský, P.L.; Hnilička, F.; Wilhelmová, N.; Rothová, O.; Kočová, M.; Procházková, D.; Honnerová, J.; et al. The physiology and proteomics of drought tolerance in maize: Early stomatal closure as a cause of lower tolerance to short-term dehydration? PLoS One 2012, 7, e38017. [Google Scholar] [CrossRef]
- Luo, M.; Liu, J.; Lee, R.D.; Scully, B.T.; Guo, B.Z. Monitoring the expression of maize genes in developing kernels under drought stress using oligo-microarray. J. Integr. Plant Biol. 2010, 52, 1059–1074. [Google Scholar] [CrossRef]
- Jiang, T.B.; Fountain, J.; Davis, G.; Kemerait, R.; Scully, B.T.; Lee, R.D.; Guo, B.Z. Root morphology and gene expression analysis in response to drought stress in maize (Zea mays). Plant Mol. Biol. Rep. 2012, 30, 360–369. [Google Scholar] [CrossRef]
- Sanguineti, M.C.; Giuliani, M.M.; Govi, G.; Tuberosa, R.; Landi, P. Root and shoot traits of maize inbred lines grown in the field and in hydroponic culture and their relationship with root lodging. Maydica 1998, 43, 211–216. [Google Scholar]
- Sen, S.; Smith, M.E.; Setter, T.L. Analysis of maize root traits in response to low nitrogen. Asian J. Plant Sci. Res. 2013, 3, 121–125. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, pp. 403–410. Available online: http://blast.ncbi.nlm.nih.gov/ (accessed on 6 December 2013). [CrossRef]
- Bray, E.A. Molecular and physiological responses to water-deficit stress. In Advances in Molecular Breeding toward Drought and Salt Tolerant Crops; Jenks, M.A., Hasegawa, P.M., Jain, S.M., Eds.; Springer: New York, NY, USA, 2007; pp. 121–140. [Google Scholar]
- Ashraf, M. Inducing drought tolerance in plants: Recent advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef]
- Fountain, J.C.; Chen, Z.Y.; Scully, B.T.; Kemerait, R.C.; Lee, R.D.; Guo, B.Z. Pathogenesis-related gene expressions in different maize genotypes under drought stressed conditions. Afr. J. Plant Sci. 2010, 4, 433–440. [Google Scholar]
- Jiang, T.B.; Zhou, B.R.; Luo, M.; Abbas, H.K.; Kemerait, R.; Lee, R.D.; Scully, B.T.; Guo, B.Z. Expression analysis of stress-related genes in kernels of different maize (Zea mays L.) inbred lines with different resistance to aflatoxin contamination. Toxins 2011, 3, 538–550. [Google Scholar] [CrossRef]
- Zadražnik, T.; Hollung, K.; Egge-Jacobsen, W.; Meglič, V.; Šuštar-Vozlič, J. Differential proteomic analysis of drought stress response in leaves of common bean (Phaseolus vulgaris L.). J. Proteomics 2013, 78, 254–272. [Google Scholar] [CrossRef]
- Alvarez, S.; Roy Choudhury, S.; Pandey, S. Comparative quantitative proteomics analysis of the ABA response of roots of drought-sensitive and drought-tolerant wheat varieties identifies proteomic signatures of drought adaptability. J. Proteome Res. 2014, 13, 1688–1701. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Brown, R.L.; Damann, K.E.; Cleveland, T.E. Identification of unique or elevated levels of kernel proteins in aflatoxin-resistant maize genotypes through proteome analysis. Phytopathology 2002, 92, 1084–1094. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Brown, R.L.; Damann, K.E.; Cleveland, T.E. Identification of maize kernel endosperm proteins associated with resistance to aflatoxin contamination by Aspergillus flavus. Phytopathology 2007, 97, 1094–1103. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Brown, R.L.; Menkir, A.; Cleveland, T.E. Identification of resistance-associated proteins in closely-related maize lines varying in aflatoxin accumulation. Mol. Breed. 2012, 30, 53–68. [Google Scholar] [CrossRef]
- Hanin, M.; Brini, F.; Ebel, C.; Toda, Y.; Takeda, S.; Masmoudi, K. Plant dehydrins and stress tolerance: Versatile proteins for complex mechanisms. Plant Signal. Behav. 2011, 6, 1503–1509. [Google Scholar] [CrossRef]
- Zhao, Y.; Du, H.M.; Wang, Z.L.; Huang, B.R. Identification of proteins associated with water-deficit tolerance in C4 perennial grass species, Cynodon dactylon × Cynodon transvaalensis and Cynodon dactylon. Physiol. Plant 2011, 141, 40–55. [Google Scholar] [CrossRef]
- Borovskii, G.B.; Stupnikova, I.V.; Antipina, A.I.; Vladimirova, S.V.; Voinikov, V.K. Accumulation of dehydrin-like proteins in the mitochondria of cereals in response to cold, freezing, drought and ABA treatment. BMC Plant Biol. 2002, 2, 5. [Google Scholar] [CrossRef]
- Koag, M.C.; Fenton, R.D.; Wilkens, S.; Close, T.J. The binding of maize DHN1 to lipid vesicles. Gain of structure and lipid specificity. Plant Physiol. 2003, 131, 309–316. [Google Scholar] [CrossRef]
- Mohammadkhani, N.; Heidari, R. Effects of drought stress on soluble proteins in two maize varieties. Turk. J. Biol. 2008, 32, 23–30. [Google Scholar]
- Chen, Z.Y.; Brown, R.L.; Lax, A.R.; Cleveland, T.E.; Russin, J.S. Inhibition of plant-pathogenic fungi by a corn trypsin inhibitor overexpressed in Escherichia coli. Appl. Environ. Microbiol. 1999, 65, 1320–1324. [Google Scholar]
- Xie, Y.R.; Chen, Z.Y.; Brown, R.L.; Bhatnagar, D. Expression and functional characterization of two pathogenesis-related protein 10 genes from Zea mays. J. Plant Physiol. 2010, 167, 121–130. [Google Scholar] [CrossRef]
- Fan, S.G.; Wu, G.J. Characteristics of plant proteinase inhibitors and their applications in combating phytophagous insects. Bot. Bull. Acad. Sin. 2005, 46, 273–292. [Google Scholar]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
- Wan, X.Y.; Liu, J.Y. Comparative proteomics analysis reveals an intimate protein network provoked by hydrogen peroxide stress in rice seedling leaves. Mol. Cell. Proteomics 2008, 7, 1469–1488. [Google Scholar] [CrossRef]
- Marrs, K.A. The functions and regulation of glutathione S-transferases in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 127–158. [Google Scholar] [CrossRef]
- Vieira dos Santos, C.; Rey, P. Plant thioredoxins are key actors in the oxidative stress response. Trends Plant Sci. 2006, 11, 329–334. [Google Scholar] [CrossRef]
- Yu, L.X.; Setter, T.L. Comparative transcriptional profiling of placenta and endosperm in developing maize kernels in response to water deficit. Plant Physiol. 2003, 131, 568–582. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Lyzenga, W.J.; Stone, S.L. Abiotic stress tolerance mediated by protein ubiquitination. J. Exp. Bot. 2011, 63, 599–616. [Google Scholar] [CrossRef]
- Harb, A.; Krishnan, A.; Ambavaram, M.M.; Pereira, A. Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol. 2010, 154, 1254–1271. [Google Scholar] [CrossRef]
- Aimar, D.; Calafat, M.; Andrade, A.M.; Carassay, L.; Abdala, G.; Molas, M.L. Drought tolerance and stress hormones: From model organisms to forage crops. In Plants and Environment; Vasanthaiah, H.K.N., Kambiranda, D., Eds.; InTech: Rijeka, Croatia, 2011; pp. 137–164. [Google Scholar]
- Kim, J.I.; Baek, D.; Park, H.C.; Chun, H.J.; Oh, D.H.; Lee, M.K.; Cha, J.Y.; Kim, W.Y.; Kim, M.C.; Chung, W.S.; et al. Overexpression of Arabidopsis YUCCA6 in potato results in high-auxin developmental phenotypes and enhanced resistance to water deficit. Mol. Plant 2013, 6, 337–349. [Google Scholar] [CrossRef]
- De Micco, V.; Aronne, G. Morpho-Anatomical traits for plant adaptation to drought. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Ricardo, A., Ed.; Springer: New York, NY, USA, 2012; pp. 37–61. [Google Scholar]
- Wilkinson, S.; Davies, W.J. Drought, ozone, ABA and ethylene: new insights from cell to plant to community. Plant Cell Environ. 2010, 33, 510–525. [Google Scholar] [CrossRef]
- Askari, H.; Edqvist, J.; Hajheidari, M.; Kafi, M.; Salekdeh, G.H. Effects of salinity levels on proteome of Suaeda aegyptiaca leaves. Proteomics 2006, 6, 2542–2554. [Google Scholar] [CrossRef]
- Gu, J.F.; Qiu, M.; Yang, J.C. Enhanced tolerance to drought in transgenic rice plants overexpressing C4 photosynthesis enzymes. Crop J. 2013, 1, 105–114. [Google Scholar] [CrossRef]
- Jiang, S.S.; Liang, X.N.; Li, X.; Wang, S.L.; Lv, D.W.; Ma, C.Y.; Li, X.H.; Ma, W.J.; Yan, Y.M. Wheat drought-responsive grain proteome analysis by linear and nonlinear 2-DE and MALDI-TOF mass spectrometry. Int. J. Mol. Sci. 2012, 13, 16065–16083. [Google Scholar] [CrossRef]
- Acevedo, R.M.; Maiale, S.J.; Pessino, S.C.; Bottini, R.; Ruiz, O.A.; Sansberro, P.A. A succinate dehydrogenase flavoprotein subunit-like transcript is up-regulated in Ilex paraguariensis leaves in response to water deficit and abscisic acid. Plant Physiol. Biochem. 2013, 65, 48–54. [Google Scholar] [CrossRef]
- Mohammadkhani, N.; Heidari, R. Drought-induced accumulation of soluble sugars and proline in two maize varieties. World Appl. Sci. J. 2008, 3, 448–453. [Google Scholar]
- Arabzadeh, N. The effect of drought stress on soluble carbohydrates (sugars) in two species of Haloxylon persicum and Haloxylon aphyllum. Plant Sci. 2012, 11, 44–51. [Google Scholar]
- Maurel, C.; Verdoucq, L.; Luu, D.T.; Santoni, V. Plant aquaporins: Membrane channels with multiple integrated functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef]
- Tuberosa, R.; Sanguineti, M.C.; Landi, P.; Giuliani, M.M.; Salvi, S.; Conti, S. Identification of QTLs for root characteristics in maize grown in hydroponics and analysis of their overlap with QTLs for grain yield in the field at two water regimes. Plant Mol. Biol. 2002, 48, 697–712. [Google Scholar] [CrossRef]
- De Micco, V.; Aronne, G. Root structure of Rumex scutatus L. growing on slopes. IAWA J. 2010, 31, 13–28. [Google Scholar] [CrossRef]
- Tardieu, F. Any trait or trait-related allele can confer drought tolerance: Just design the right drought scenario. J. Exp. Bot. 2012, 63, 25–31. [Google Scholar] [CrossRef]
- Omori, F.; Mano, Y. QTL mapping of root angle in F2 populations from maize “B73” teosinte “Zea luxurians”. Plant Root 2007, 1, 57–65. [Google Scholar] [CrossRef]
- Zheng, J.; Fu, J.; Gou, M.; Huai, J.; Liu, Y.; Jian, M.; Huang, Q.; Guo, X.; Dong, Z.; Wang, H.; et al. Genome-wide transcriptome analysis of two maize inbred lines under drought stress. Plant Mol. Biol. 2010, 72, 407–421. [Google Scholar] [CrossRef]
- Kausar, R.; Arshad, M.; Shahzad, A.; Komatsu, S. Proteomics analysis of sensitive and tolerant barley genotypes under drought stress. Amino Acids 2013, 44, 345–359. [Google Scholar] [CrossRef]
- Pechanova, O.; Pechan, T.; Williams, W.P.; Luthe, D.S. Proteomic analysis of the maize rachis: Potential roles of constitutive and induced proteins in resistance to Aspergillus flavus infection and aflatoxin accumulation. Proteomics 2011, 11, 114–127. [Google Scholar] [CrossRef]
- Yang, L.M.; Tian, D.G.; Todd, C.D.; Luo, Y.M.; Hu, X.Y. Comparative proteome analyses reveal that nitric oxide is an important signal molecule in the response of rice to aluminum toxicity. J. Proteome Res. 2013, 12, 1316–1330. [Google Scholar] [CrossRef]
- Yang, L.M.; Luo, Y.M.; Wei, J.F.; Ren, C.M.; Zhou, X.; He, S.H. Methods for protein identification using expressed sequence tags and peptide mass fingerprinting for seed crops without complete genome sequences. Seed Sci. Res. 2010, 20, 257–262. [Google Scholar] [CrossRef]
- Schägger, H. Tricine-SDS-PAGE. Nat. Protoc. 2006, 1, 16–22. [Google Scholar] [CrossRef]
- Chen, S. High throughput protein identification using direct infusion nanoelectrospray ionization mass spectrometry. Proteomics 2006, 6, 16–25. [Google Scholar] [CrossRef]
- Zhu, M.; Dai, S.; McClung, S.; Yan, X.; Chen, S. Functional differentiation of Brassica napus guard cells and mesophyll cells revealed by comparative proteomics. Mol. Cell. Proteomics 2009, 8, 752–766. [Google Scholar] [CrossRef]
- Bai, X.G.; Yang, L.M.; Yang, Y.Q.; Ahmad, P.; Yang, Y.P.; Hu, X.Y. Deciphering the protective role of nitric oxide against salt stress at the physiological and proteomic levels in maize. J. Proteome Res. 2011, 10, 4349–4364. [Google Scholar] [CrossRef]
- Tang, W.H.; Shilov, I.V.; Seymour, S.L. Nonlinear fitting method for determining local false discovery rates from decoy database searches. J. Proteome Res. 2008, 7, 3661–3667. [Google Scholar] [CrossRef]
- Millioni, R.; Tolin, S.; Puricelli, L.; Sbrignadello, S.; Fadini, G.P.; Tessari, P.; Arrigoni, G. High abundance proteins depletion vs. low abundance proteins enrichment: Comparison of methods to reduce the plasma proteome complexity. PLoS One 2011, 6, e19603. [Google Scholar] [CrossRef]
- Pang, Q.; Chen, S.; Dai, S.; Chen, Y.; Wang, Y.; Yan, X. Comparative proteomics of salt tolerance in Arabidopsis thaliana and Thellungiella halophila. J. Proteome Res. 2010, 9, 2584–2599. [Google Scholar] [CrossRef]
- Neilson, K.A.; Mariani, M.; Haynes, P.A. Quantitative proteomic analysis of cold-responsive proteins in rice. Proteomics 2011, 11, 1696–1706. [Google Scholar] [CrossRef]
- Lin, Z.; Zhang, X.; Yang, X.; Li, G.; Tang, S.; Wang, S.; Ding, Y.; Liu, Z. Proteomic analysis of proteins related to rice grain chalkiness using iTRAQ and a novel comparison system based on a notched-belly mutant with white-belly. BMC Plant Biol. 2014, 14, 163. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, pp. D222–D230. Available online: http://www.sanger.ac.uk/Software/Pfam (accessed on 12 March 2014). [CrossRef]
- Hunter, S.; Jones, P.; Mitchell, A.; Apweiler, R.; Attwood, T.K.; Bateman, A.; Bernard, T.; Binns, D.; Bork, P.; Burge, S.; et al. InterPro in 2011: New developments in the family and domain prediction database. Nucleic Acids Res. 2012, 40, pp. D306–D312. Available online: http://www.ebi.ac.uk/interpro (accessed on 12 March 2014). [CrossRef]
- Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Birney, E.; Biswas, M. The InterPro database, an integrated documentation resource for protein families, domains and functional sites. Nucleic Acids Res. 2001, 29, 37–40. [Google Scholar] [CrossRef]
- Bateman, A.; Birney, E.; Cerruti, L.; Durbin, R.; Etwiller, L.; Eddy, S.R. The Pfam protein families database. Nucleic Acids Res. 2002, 30, 276–280. [Google Scholar] [CrossRef]
- Bancroft, I.; Bent, E.; Love, K.; Goodman, H.; Dean, C.; Bergkamp, R. Analysis of 1.9 Mb of contiguous sequence from chromosome 4. Nature 1998, 391, 485–488. [Google Scholar] [CrossRef]
- Wu, C.H.; Yeh, L.S.; Huang, H.; Arminski, L.; Castro-Alvear, J.; Chen, Y.; Hu, Z.; Kourtesis, P.; Ledley, R.S.; Suzek, B.E.; et al. The Protein Information Resource. Nucleic Acids Res. 2003, 31, pp. 345–347. Available online: http://pir.georgetown.edu (accessed on 20 March 2014). [CrossRef]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; von Mering, C.; et al. STRING v9.1: Protein–protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, pp. D808–815. Available online: http://string-db.org (accessed on 20 March 2014). [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Jiang, T.; Fountain, J.C.; Scully, B.T.; Lee, R.D.; Kemerait, R.C.; Chen, S.; Guo, B. Protein Profiles Reveal Diverse Responsive Signaling Pathways in Kernels of Two Maize Inbred Lines with Contrasting Drought Sensitivity. Int. J. Mol. Sci. 2014, 15, 18892-18918. https://doi.org/10.3390/ijms151018892
Yang L, Jiang T, Fountain JC, Scully BT, Lee RD, Kemerait RC, Chen S, Guo B. Protein Profiles Reveal Diverse Responsive Signaling Pathways in Kernels of Two Maize Inbred Lines with Contrasting Drought Sensitivity. International Journal of Molecular Sciences. 2014; 15(10):18892-18918. https://doi.org/10.3390/ijms151018892
Chicago/Turabian StyleYang, Liming, Tingbo Jiang, Jake C. Fountain, Brian T. Scully, Robert D. Lee, Robert C. Kemerait, Sixue Chen, and Baozhu Guo. 2014. "Protein Profiles Reveal Diverse Responsive Signaling Pathways in Kernels of Two Maize Inbred Lines with Contrasting Drought Sensitivity" International Journal of Molecular Sciences 15, no. 10: 18892-18918. https://doi.org/10.3390/ijms151018892