
Epigenomic and Other Evidence for Cannabis-Induced Aging Contextualized in a Synthetic Epidemiologic Overview of Cannabinoid-Related Teratogenesis and Cannabinoid-Related Carcinogenesis

Abstract
:1. Introduction
1.1. Key Definitions
1.2. Outline
2. Methods
3. Results and Discussion
3.1. Streams of Evidence for Cannabinoid Acceleration of Aging
3.1.1. Clinical Syndromes
3.1.2. Mitochondrial Inhibition
3.1.3. DNA Methylation
3.1.4. Mental Illnesses
3.1.5. Cardiovascular Age
3.1.6. Endocrine Suppression
3.1.7. Liver Inflammation
3.1.8. Cancer
3.1.9. Inheritable Cancer
3.1.10. Congenital Anomalies
3.1.11. Telomerase Inhibition
3.1.12. Elevated Mortality Rate
3.2. Pathogenetic Field of Interest
3.2.1. Epigenomic Overview
3.2.2. Stem-Cell Factors
3.2.3. Chromosomal Mechanics
3.2.4. Centromeres and Kinetochores
3.2.5. Prefrontal Cortex and Brain
3.2.6. Cardiovascular System
3.2.7. Cannabinoid-Related Teratogenesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
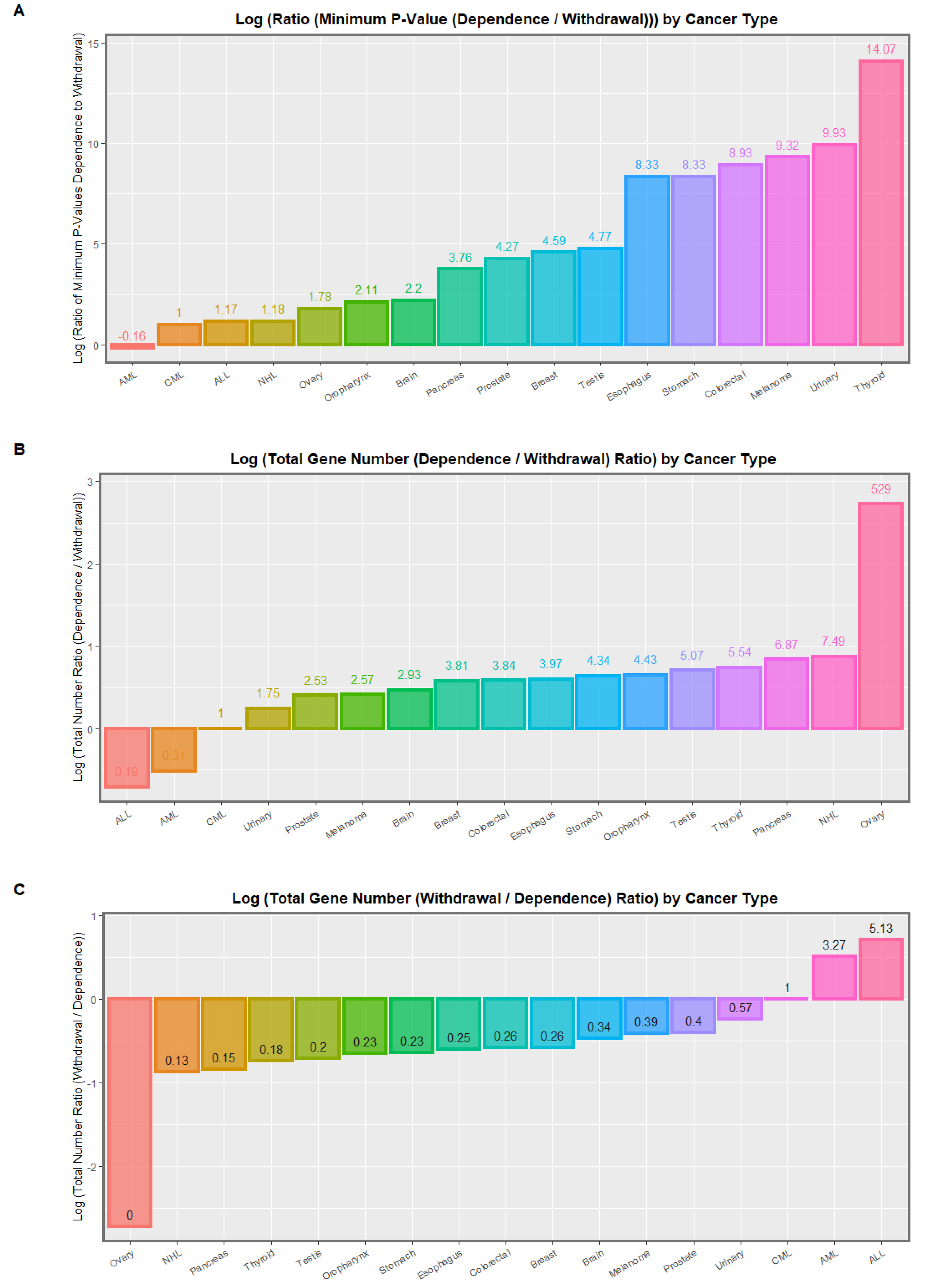{kind=link}
| (A) | |||||||||
| Nearest Gene Name | Nearest Gene Number | Page No. | Annotation | Chromosome Number | Dependency Status | Relative Position | Distance to Nearest Gene | p-Value | Bonferroni Adjusted p-Value |
| PTCH1 | ENSG00000185920 | 58 | Shh Receptor | 9 | Dependence | Intron | 0 | 3.46 × 10−6 | 0.012789 |
| PTCHD1-AS | ENSG00000233067 | 91 | lnc Promoter/enhancer | X | Dependence | Intron | 0 | 8.61 × 10−6 | 0.019678 |
| PTCHD1-AS | ENSG00000233067 | 129 | lnc Promoter/enhancer | X | Withdrawal | Intron | 0 | 8.21 × 10−8 | 0.002096 |
| PTCHD4 | ENSG00000244694 | 138 | Shh Receptor; Otopalatodigital syndrome | 6 | Withdrawal | Intron | 0 | 4.21 × 10−7 | 0.005104 |
| PTCH1 | ENSG00000185920 | 185 | Shh Receptor | 9 | Withdrawal | Intron | 0 | 5.80 × 10−6 | 0.017679 |
| SUFU | ENSG00000161996 | 207 | Hedgehog Inhibitor | 16 | Withdrawal | Exon | 0 | 1.01 × 10−5 | 0.022942 |
| Gli3 | ENSG00000106571 | 78 | Shh mediator | 7 | Dependence | Downstream | 81232 | 6.35 × 10−6 | 0.017090 |
| Gli3 | ENSG00000106571 | 99 | Shh mediator | 7 | Dependence | Intron | 0 | 1.00 × 10−5 | 0.021181 |
| Gli3 | ENSG00000106571 | 124 | Shh mediator | 7 | Withdrawal | Downstream | 20318 | 8.23 × 10−9 | 0.000646 |
| Gli3 | ENSG00000106571 | 182 | Shh mediator | 7 | Withdrawal | Intron | 0 | 5.28 × 10−6 | 0.001687 |
| Gli3 | ENSG00000106571 | 231 | Shh mediator | 7 | Withdrawal | Intron | 0 | 1.62 × 10−5 | 0.028539 |
| (B) | |||||||||
| Nearest Gene Name | Nearest Gene Number | Page No. | Annotation | Chromosome Number | Dependency Status | Number Genes Identified | Function | p-Value | |
| PTCH1 | ENSG00000185920 | 237 | Notch Processing | 9 | KEGG Pathway | 31 | Notch Processing | 0.044117 | |
| PTCH1 | ENSG00000185920 | 238 | Skin cancer | 9 | KEGG Pathway | 54 | Notch Processing | 0.067770 | |
| PSENEN | ENSG00000185920 | 326 | Cutaneous melanoma | 19 | Withdrawal | 110 | Notch Processing | 0.000008 | |
| Gli3 | ENSG00000106571 | 325 | Skin lesion | 7 | Withdrawal | 115 | Notch transcription factor | 1.65 × 10−6 | |
| Gli3 | ENSG00000106571 | 325 | Head and Neck SCC | 7 | Withdrawal | 53 | Notch transcription factor | 3.59 × 10−6 | |
| Gli3 | ENSG00000106571 | 325 | Skin cancer | 7 | Withdrawal | 113 | Notch transcription factor | 4.79 × 10−6 | |
| Gli3 | ENSG00000106571 | 325 | Lung adenocarcinoma | 7 | Withdrawal | 42 | Notch transcription factor | 5.84 × 10−6 | |
| Gli3 | ENSG00000106571 | 325 | Cancer | 7 | Withdrawal | 149 | Notch transcription factor | 7.17 × 10−6 | |
| Gli3 | ENSG00000106571 | 326 | Large bowel cancer | 7 | Withdrawal | 120 | Notch transcription factor | 7.45 × 10−6 | |
| Gli3 | ENSG00000106571 | 326 | Cutaneous melanoma | 7 | Withdrawal | 110 | Notch transcription factor | 7.71 × 10−6 | |
| Gli3 | ENSG00000106571 | 326 | High-grade astrocytoma | 7 | Withdrawal | 82 | Notch transcription factor | 8.42 × 10−6 | |
| Gli3 | ENSG00000106571 | 326 | Abdominal adenocarcinoma | 7 | Withdrawal | 135 | Notch transcription factor | 8.46 × 10−6 | |
| Gli3 | ENSG00000106571 | 327 | Solid cancer | 7 | Withdrawal | 150 | Notch transcription factor | 9.16 × 10−6 | |
| Gli3 | ENSG00000106571 | 327 | Head and Neck cancer | 7 | Withdrawal | 137 | Notch transcription factor | 9.54 × 10−6 | |
| Gli3 | ENSG00000106571 | 327 | Sensory development | 7 | Withdrawal | 18 | Notch transcription factor | 1.30 × 10−5 | |
| Gli3 | ENSG00000106571 | 327 | Carcinoma | 7 | Withdrawal | 148 | Notch transcription factor | 1.38 × 10−5 | |
3.2.8. Cannabinoid-Related Carcinogenesis
3.3. Implications of Findings
3.4. Spermatocytes
3.5. Oocytes
3.6. Zygotes
3.7. Cannabidiol and Δ8THC
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, D.; Furst, A.; Meaburn, K.; Lezaja, A.; Wen, Y.; Altmeyer, M.; Reina-San-Martin, B.; Soutoglou, E. Activation of homologous recombination in G1 preserves centromeric integrity. Nature 2021, 600, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafe, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging: An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Salvioli, S.; Monti, D.; Lanzarini, C.; Conte, M.; Pirazzini, C.; Bacalini, M.G.; Garagnani, P.; Giuliani, C.; Fontanesi, E.; Ostan, R.; et al. Immune system, cell senescence, aging and longevity--inflamm-aging reappraised. Curr. Pharm. Des. 2013, 19, 1675–1679. [Google Scholar] [PubMed]
- Beausejour, C.M.; Campisi, J. Ageing: Balancing regeneration and cancer. Nature 2006, 443, 404–405. [Google Scholar] [CrossRef] [PubMed]
- Busuttil, R.A.; Dollé, M.; Campisi, J.; Vijga, J. Genomic instability, aging, and cellular senescence. Ann. N. Y. Acad. Sci. 2004, 1019, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J. The biology of replicative senescence. Eur. J. Cancer 1997, 33, 703–709. [Google Scholar] [CrossRef]
- Rayess, H.; Wang, M.B.; Srivatsan, E.S. Cellular senescence and tumor suppressor gene p16. Int. J. Cancer 2012, 130, 1715–1725. [Google Scholar] [CrossRef] [Green Version]
- Waddington, C.H. Organizers and Genes; Cambridge University Press: Cambridge, UK, 1940; Volume 1. [Google Scholar]
- Gonzales, K.A.U.; Polak, L.; Matos, I.; Tierney, M.T.; Gola, A.; Wong, E.; Infarinato, N.R.; Nikolova, M.; Luo, S.; Liu, S.; et al. Stem cells expand potency and alter tissue fitness by accumulating diverse epigenetic memories. Science 2021, 374, eabh2444. [Google Scholar] [CrossRef]
- Hota, S.K.; Rao, K.S.; Blair, A.P.; Khalilimeybodi, A.; Hu, K.M.; Thomas, R.; So, K.; Kameswaran, V.; Xu, J.; Polacco, B.J.; et al. Brahma safeguards canalization of cardiac mesoderm differentiation. Nature 2022, 602, 129–134. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocampo, A.; Reddy, P.; Martinez-Redondo, P.; Platero-Luengo, A.; Hatanaka, F.; Hishida, T.; Li, M.; Lam, D.; Kurita, M.; Beyret, E.; et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell 2016, 167, 1719–1733.e1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Lüttmann, F.F.; Schoger, E.; Schöler, H.R.; Zelarayán, L.C.; Kim, K.P.; Haigh, J.J.; Kim, J.; Braun, T. Reversible reprogramming of cardiomyocytes to a fetal state drives heart regeneration in mice. Science 2021, 373, 1537–1540. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Brommer, B.; Tian, X.; Krishnan, A.; Meer, M.; Wang, C.; Vera, D.L.; Zeng, Q.; Yu, D.; Bonkowski, M.S.; et al. Reprogramming to recover youthful epigenetic information and restore vision. Nature 2020, 588, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M.B.; Sinclair, D.A. When stem cells grow old: Phenotypes and mechanisms of stem cell aging. Development 2016, 143, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkow, N.D.; Baler, R.D.; Compton, W.M.; Weiss, S.R. Adverse Health Effects of Marijuana Use. N. Engl. J. Med. 2014, 370, 2219–2227. [Google Scholar] [CrossRef] [Green Version]
- Substance Abuse and Mental Health Services Administration. Key Substance Use and Mental Health Indicators in the United States: Results from the 2020 National Survey on Drug Use and Health (NSDUH). Available online: https://www.samhsa.gov/data/sites/default/files/reports/rpt35325/NSDUHFFRPDFWHTMLFiles2020/2020NSDUHFFR1PDFW102121.pdf (accessed on 1 October 2021).
- Morishima, A. Effects of cannabis and natural cannabinoids on chromosomes and ova. NIDA Res. Monogr. 1984, 44, 25–45. [Google Scholar] [PubMed]
- Morishima, A.; Henrich, R.T.; Jayaraman, J.; Nahas, G.G. Hypoploid metaphases in cultured lymphocytes of marihuana smokers. Adv. Biosci. 1978, 22–23, 371–376. [Google Scholar]
- Leuchtenberger, C.; Leuchtenberger, R. Morphological and cytochemical effects of marijuana cigarette smoke on epithelioid cells of lung explants from mice. Nature 1971, 234, 227–229. [Google Scholar] [CrossRef]
- Leuchtenberger, C.; Leuchtenberger, R.; Schneider, A. Effects of marijuana and tobacco smoke on human lung physiology. Nature 1973, 241, 137–139. [Google Scholar] [CrossRef]
- Stenchever, M.A.; Kunysz, T.J.; Allen, M.A. Chromosome breakage in users of marihuana. Am. J. Obstet. Gynecol. 1974, 118, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.F.S.; Nahas, G.G.; Hembree, W.C. Effects of Marijuana Inhalation on Spermatogenesis of the Rat. In Marijuana in Medicine; Nahas, G.G., Sutin, K.M., Harvey, D.J., Agurell, S., Eds.; Human Press: Totowa, NY, USA, 1999; Volume 1, pp. 359–366. [Google Scholar]
- Reece, A.S.; Hulse, G.K. Causal inference multiple imputation investigation of the impact of cannabinoids and other substances on ethnic differentials in US testicular cancer incidence. BMC Pharmacol. Toxicol. 2021, 22, 40–71. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.K.; Itchon-Ramos, N.; Visco, Z.; Huang, Z.; Grenier, C.; Schrott, R.; Acharya, K.; Boudreau, M.H.; Price, T.M.; Raburn, D.J.; et al. Cannabinoid exposure and altered DNA methylation in rat and human sperm. Epigenetics 2018, 13, 1208–1221. [Google Scholar] [CrossRef] [Green Version]
- Schrott, R.; Murphy, S.K.; Modliszewski, J.L.; King, D.E.; Hill, B.; Itchon-Ramos, N.; Raburn, D.; Price, T.; Levin, E.D.; Vandrey, R.; et al. Refraining from use diminishes cannabis-associated epigenetic changes in human sperm. Environ. Epigenetics 2021, 7, dvab009. [Google Scholar] [CrossRef] [PubMed]
- DiNieri, J.A.; Wang, X.; Szutorisz, H.; Spano, S.M.; Kaur, J.; Casaccia, P.; Dow-Edwards, D.; Hurd, Y.L. Maternal cannabis use alters ventral striatal dopamine D2 gene regulation in the offspring. Biol. Psychiatry 2011, 70, 763–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szutorisz, H.; DiNieri, J.A.; Sweet, E.; Egervari, G.; Michaelides, M.; Carter, J.M.; Ren, Y.; Miller, M.L.; Blitzer, R.D.; Hurd, Y.L. Parental THC exposure leads to compulsive heroin-seeking and altered striatal synaptic plasticity in the subsequent generation. Neuropsychopharmacology 2014, 39, 1315–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szutorisz, H.; Hurd, Y.L. Epigenetic Effects of Cannabis Exposure. Biol. Psychiatry 2016, 79, 586–594. [Google Scholar] [CrossRef] [Green Version]
- Szutorisz, H.; Hurd, Y.L. High times for cannabis: Epigenetic imprint and its legacy on brain and behavior. Neurosci. Biobehav. Rev. 2018, 85, 93–101. [Google Scholar] [CrossRef]
- Watson, C.T.; Szutorisz, H.; Garg, P.; Martin, Q.; Landry, J.A.; Sharp, A.J.; Hurd, Y.L. Genome-Wide DNA Methylation Profiling Reveals Epigenetic Changes in the Rat Nucleus Accumbens Associated with Cross-Generational Effects of Adolescent THC Exposure. Neuropsychopharmacology 2015, 40, 2993–3005. [Google Scholar] [CrossRef] [Green Version]
- Rossato, M.; Pagano, C.; Vettor, R. The cannabinoid system and male reproductive functions. J. Neuroendocrinol. 2008, 20 (Suppl. 1), 90–93. [Google Scholar] [CrossRef]
- Chioccarelli, T.; Cacciola, G.; Altucci, L.; Lewis, S.E.; Simon, L.; Ricci, G.; Ledent, C.; Meccariello, R.; Fasano, S.; Pierantoni, R.; et al. Cannabinoid receptor 1 influences chromatin remodeling in mouse spermatids by affecting content of transition protein 2 mRNA and histone displacement. Endocrinology 2010, 151, 5017–5029. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, B.L.; Springs, A.E.; Kaminski, N.E. The profile of immune modulation by cannabidiol (CBD) involves deregulation of nuclear factor of activated T cells (NFAT). Biochem. Pharmacol. 2008, 76, 726–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenstein, T.K.; Meissler, J.J. Effects of Cannabinoids on T-cell Function and Resistance to Infection. J. Neuroimmune Pharmacol. 2015, 10, 204–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiurchiu, V. Endocannabinoids and Immunity. Cannabis Cannabinoid Res. 2016, 1, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, B.L.F. Evaluation of Marijuana Compounds on Neuroimmune Endpoints in Experimental Autoimmune Encephalomyelitis. Curr. Protoc. Toxicol. 2018, 75, 11.25.1–11.25.22. [Google Scholar] [CrossRef] [PubMed]
- Bindukumar, B.; Mahajan, S.D.; Reynolds, J.L.; Hu, Z.; Sykes, D.E.; Aalinkeel, R.; Schwartz, S.A. Genomic and proteomic analysis of the effects of cannabinoids on normal human astrocytes. Brain Res. 2008, 1191, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Hegde, V.L.; Rao, R.; Zhang, J.; Nagarkatti, P.S.; Nagarkatti, M. Histone modifications are associated with Delta9-tetrahydrocannabinol-mediated alterations in antigen-specific T cell responses. J. Biol. Chem. 2014, 289, 18707–18718. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.Y.; Huang, C.H.; Lin, Y.H.; Wang, C.C.; Jan, T.R. Cannabidiol induced apoptosis in human monocytes through mitochondrial permeability transition pore-mediated ROS production. Free Radic. Biol. Med. 2018, 124, 311–318. [Google Scholar] [CrossRef]
- Chiu, P.; Karler, R.; Craven, C.; Olsen, D.M.; Turkanis, S.A. The influence of delta9-tetrahydrocannabinol, cannabinol and cannabidiol on tissue oxygen consumption. Res. Commun. Chem. Pathol. Pharmacol. 1975, 12, 267–286. [Google Scholar]
- Harkany, T.; Horvath, T.L. (S)Pot on Mitochondria: Cannabinoids Disrupt Cellular Respiration to Limit Neuronal Activity. Cell Metab. 2017, 25, 8–10. [Google Scholar] [CrossRef] [Green Version]
- Hebert-Chatelain, E.; Desprez, T.; Serrat, R.; Bellocchio, L.; Soria-Gomez, E.; Busquets-Garcia, A.; Pagano Zottola, A.C.; Delamarre, A.; Cannich, A.; Vincent, P.; et al. A cannabinoid link between mitochondria and memory. Nature 2016, 539, 555–559. [Google Scholar] [CrossRef] [PubMed]
- McClean, D.K.; Zimmerman, A.M. Action of delta 9-tetrahydrocannabinol on cell division and macromolecular synthesis in division-synchronized protozoa. Pharmacology 1976, 14, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Tilak, S.; Zimmerman, S.; Zimmerman, A.M. Action of delta 9-tetrahydrocannabinol on the pool of acid soluble nucleotides. Cytobios 1984, 40, 71–85. [Google Scholar] [PubMed]
- Tahir SK, Zimmerman AM: Influence of marihuana on cellular structures and biochemical activities. Pharmacol. Biochem. Behav. 1991, 40, 617–623. [CrossRef]
- Parker, S.J.; Zuckerman, B.S.; Zimmermann, A.M. The Effects of Maternal Marijuana Use During Pregnancy on Fetal Growth. In Marijuana in Medicine; Nahas, G.G., Sutin, K.M., Harvey, D.J., Agurell, S., Eds.; Humana Press: Totowa, NY, USA, 1999; Volume 1, pp. 461–468. [Google Scholar]
- Mon, M.J.; Haas, A.E.; Stein, J.L.; Stein, G.S. Influence of psychoactive and nonpsychoactive cannabinoids on cell proliferation and macromolecular biosynthesis in human cells. Biochem. Pharmacol. 1981, 30, 31–43. [Google Scholar] [CrossRef]
- Zimmerman, A.M.; Raj, A.Y. Influence of cannabinoids on somatic cells in vivo. Pharmacology 1980, 21, 277–287. [Google Scholar] [CrossRef]
- Tahir, S.K.; Trogadis, J.E.; Stevens, J.K.; Zimmerman, A.M. Cytoskeletal organization following cannabinoid treatment in undifferentiated and differentiated PC12 cells. Biochem. Cell Biol. 1992, 70, 1159–1173. [Google Scholar] [CrossRef]
- Aldington, S.; Harwood, M.; Cox, B.; Weatherall, M.; Beckert, L.; Hansell, A.; Pritchard, A.; Robinson, G.; Beasley, R. Cannabis use and risk of lung cancer: A case-control study. Eur. Respir. J. 2008, 31, 280–286. [Google Scholar] [CrossRef] [Green Version]
- Voirin, N.; Berthiller, J.; Benhaïm-Luzon, V.; Boniol, M.; Straif, K.; Ayoub, W.B.; Ayed, F.B.; Sasco, A.J. Risk of lung cancer and past use of cannabis in Tunisia. J. Thorac. Oncol. 2006, 1, 577–579. [Google Scholar] [CrossRef]
- Berthiller, J.; Straif, K.; Boniol, M.; Voirin, N.; Benhaïm-Luzon, V.; Ayoub, W.B.; Dari, I.; Laouamri, S.; Hamdi-Cherif, M.; Bartal, M.; et al. Cannabis smoking and risk of lung cancer in men: A pooled analysis of three studies in Maghreb. J. Thorac. Oncol. 2008, 3, 1398–1403. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.F.; Morgenstern, H.; Spitz, M.R.; Tashkin, D.P.; Yu, G.P.; Marshall, J.R.; Hsu, T.C.; Schantz, S.P. Marijuana use and increased risk of squamous cell carcinoma of the head and neck. Cancer Epidemiol. Biomark. Prev. 1999, 8, 1071–1078. [Google Scholar]
- Hashibe, M.; Ford, D.E.; Zhang, Z.F. Marijuana smoking and head and neck cancer. J. Clin. Pharmacol. 2002, 42 (Suppl. 11), 103S–107S. [Google Scholar] [CrossRef] [PubMed]
- Sidney, S.; Quesenberry, C.P., Jr.; Friedman, G.D.; Tekawa, I.S. Marijuana use and cancer incidence (California, United States). Cancer Causes Control 1997, 8, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Daling, J.R.; Doody, D.R.; Sun, X.; Trabert, B.L.; Weiss, N.S.; Chen, C.; Biggs, M.L.; Starr, J.R.; Dey, S.K.; Schwartz, S.M. Association of marijuana use and the incidence of testicular germ cell tumors. Cancer 2009, 115, 1215–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efird, J.T.; Friedman, G.D.; Sidney, S.; Klatsky, A.; Habel, L.A.; Udaltsova, N.V.; Van Den Eeden, S.; Nelson, L.M. The risk for malignant primary adult-onset glioma in a large, multiethnic, managed-care cohort: Cigarette smoking and other lifestyle behaviors. J. Neurooncol. 2004, 68, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Moiche Bokobo, P.; de la Presa, M.A.; Cuesta Angulo, J. Transitional cell carcinoma in a young heavy marihuana smoker. Arch. Esp. Urol. 2001, 54, 165–167. [Google Scholar]
- Chacko, J.A.; Heiner, J.G.; Siu, W.; Macy, M.; Terris, M.K. Association between marijuana use and transitional cell carcinoma. Urology 2006, 67, 100–104. [Google Scholar] [CrossRef]
- Nieder, A.M.; Lipke, M.C.; Madjar, S. Transitional cell carcinoma associated with marijuana: Case report and review of the literature. Urology 2006, 67, 200. [Google Scholar] [CrossRef]
- Bluhm, E.C.; Daniels, J.; Pollock, B.H.; Olshan, A.F. Maternal use of recreational drugs and neuroblastoma in offspring: A report from the Children’s Oncology Group (United States). Cancer Causes Control 2006, 17, 663–669. [Google Scholar] [CrossRef]
- Hashibe, M.; Straif, K.; Tashkin, D.P.; Morgenstern, H.; Greenlandm, S.; Zhang, Z.F. Epidemiologic review of marijuana use and cancer risk. Alcohol 2005, 35, 265–275. [Google Scholar] [CrossRef]
- Robison, L.L.; Buckley, J.D.; Daigle, A.E.; Wells, R.; Benjamin, D.; Arthur, D.C.; Hammond, G.D. Maternal drug use and risk of childhood nonlymphoblastic leukemia among offspring. An epidemiologic investigation implicating marijuana (a report from the Childrens Cancer Study Group). Cancer 1989, 63, 1904–1911. [Google Scholar] [CrossRef] [PubMed]
- Reece, A.S.; Hulse, G.K. Epidemiological Overview of Multidimensional Chromosomal and Genome Toxicity of Cannabis Exposure in Congenital Anomalies and Cancer Development. Sci. Rep. 2021, 11, 13892–13912. [Google Scholar] [CrossRef] [PubMed]
- Borowska, M.; Czarnywojtek, A.; Sawicka-Gutaj, N.; Woliński, K.; Płazińska, M.T.; Mikołajczak, P.; Ruchała, M. The effects of cannabinoids on the endocrine system. Endokrynol. Pol. 2018, 69, 705–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meah, F.; Lundholm, M.; Emanuele, N.; Amjed, H.; Poku, C.; Agrawal, L.; Emanuele, M.A. The effects of cannabis and cannabinoids on the endocrine system. Rev. Endocr. Metab. Disord. 2021, 23, 401–420. [Google Scholar] [CrossRef] [PubMed]
- Battista, N.; Pasquariello, N.; Di Tommaso, M.; Maccarrone, M. Interplay between endocannabinoids, steroids and cytokines in the control of human reproduction. J. Neuroendocrinol. 2008, 20 (Suppl. 1), 82–89. [Google Scholar] [CrossRef] [PubMed]
- Battista, N.; Rapino, C.; Di Tommaso, M.; Bari, M.; Pasquariello, N.; Maccarrone, M. Regulation of male fertility by the endocannabinoid system. Mol. Cell Endocrinol. 2008, 286 (Suppl. 1), S17–S23. [Google Scholar] [CrossRef] [Green Version]
- Battista, N.; Bari, M.; Maccarrone, M. Endocannabinoids and Reproductive Events in Health and Disease. Handb. Exp. Pharmacol. 2015, 231, 341–365. [Google Scholar]
- Smith, C.G.; Asch, R.H. Acute, short-term, and chronic effects of marijuana on the female primate reproductive function. NIDA Res. Monogr. 1984, 44, 82–96. [Google Scholar]
- Mendelson, J.H.; Mello, N.K. Effects of marijuana on neuroendocrine hormones in human males and females. NIDA Res. Monogr. 1984, 44, 97–114. [Google Scholar]
- Hillard, C.J. Endocannabinoids and the Endocrine System in Health and Disease. In Endocannabinoids; Pertwee, R.G., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 317–339. [Google Scholar]
- Teschendorff, A.E.; Menon, U.; Gentry-Maharaj, A.; Ramus, S.J.; Weisenberger, D.J.; Shen, H.; Campan, M.; Noushmehr, H.; Bell, C.G.; Maxwell, A.P.; et al. Age-dependent DNA methylation of genes that are suppressed in stem cells is a hallmark of cancer. Genome Res. 2010, 20, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Horvath, S.; Zhang, Y.; Langfelder, P.; Kahn, R.S.; Boks, M.P.; van Eijk, K.; van den Berg, L.H.; Ophoff, R.A. Aging effects on DNA methylation modules in human brain and blood tissue. Genome Biol. 2012, 13, R97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.I.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.B.; Gao, Y.; et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reece, A.S.; Norman, A.; Hulse, G.K. Acceleration of cardiovascular-biological age by amphetamine exposure is a power function of chronological age. Heart Asia 2017, 9, 30–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reece, A.S.; Hulse, G.K. Reduction in arterial stiffness and vascular age by naltrexone-induced interruption of opiate agonism: A cohort study. BMJ Open 2013, 3, e002610. [Google Scholar] [CrossRef] [Green Version]
- Reece, A.S.; Norman, A.; Hulse, G.K. Cannabis Exposure as an Interactive Cardiovascular Risk Factor and Accelerant of Organismal Ageing—A Longitudinal Study. BMJ Open 2016, 6, e011891–e011900. [Google Scholar] [CrossRef] [Green Version]
- Reece, A.S.; Hulse, G.K. Impact of Lifetime Opioid Exposure on Arterial Stiffness and Vascular Age: Cross-sectional and Longitudinal Studies in Men and Women. BMJ Open 2014, 4, e004521. [Google Scholar] [CrossRef]
- Sayed, N.; Huang, Y.; Nguyen, K.; Krejciova-Rajaniemi, Z.; Grawe, A.P.; Gao, T.; Tibshirani, R.; Hastie, T.; Alpert, A.; Cui, L.; et al. An inflammatory aging clock (iAge) based on deep learning tracks multimorbidity, immunosenescence, frailty and cardiovascular aging. Nat. Aging 2021, 1, 598–615. [Google Scholar] [CrossRef]
- Yu, Z.; Zhai, G.; Singmann, P.; He, Y.; Xu, T.; Prehn, C.; Römisch-Margl, W.; Lattka, E.; Gieger, C.; Soranzo, N.; et al. Human serum metabolic profiles are age dependent. Aging Cell 2012, 11, 960–967. [Google Scholar] [CrossRef]
- Menni, C.; Kastenmüller, G.; Petersen, A.K.; Bell, J.T.; Psatha, M.; Tsai, P.C.; Gieger, C.; Schulz, H.; Erte, I.; John, S.; et al. Metabolomic markers reveal novel pathways of ageing and early development in human populations. Int. J. Epidemiol. 2013, 42, 1111–1119. [Google Scholar] [CrossRef] [Green Version]
- Lawton, K.A.; Berger, A.; Mitchell, M.; Milgram, K.E.; Evans, A.M.; Guo, L.; Hanson, R.W.; Kalhan, S.C.; Ryals, J.A.; Milburn, M.V. Analysis of the adult human plasma metabolome. Pharmacogenomics 2008, 9, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Maekawa, K.; Saito, K.; Senoo, Y.; Urata, M.; Murayama, M.; Tajima, Y.; Kumagai, Y.; Saito, Y. Plasma and serum lipidomics of healthy white adults shows characteristic profiles by subjects’ gender and age. PLoS ONE 2014, 9, e91806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holly, A.C.; Melzer, D.; Pilling, L.C.; Henley, W.; Hernandez, D.G.; Singleton, A.B.; Bandinelli, S.; Guralnik, J.M.; Ferrucci, L.; Harries, L.W. Towards a gene expression biomarker set for human biological age. Aging Cell 2013, 12, 324–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertel, J.; Friedrich, N.; Wittfeld, K.; Pietzner, M.; Budde, K.; Van der Auwera, S.; Lohmann, T.; Teumer, A.; Völzke, H.; Nauck, M.; et al. Measuring Biological Age via Metabonomics: The Metabolic Age Score. J. Proteome Res. 2016, 15, 400–410. [Google Scholar] [CrossRef]
- Collino, S.; Montoliu, I.; Martin, F.P.J.; Scherer, M.; Mari, D.; Salvioli, S.; Bucci, L.; Ostan, R.; Monti, D.; Biagi, E.; et al. Metabolic signatures of extreme longevity in northern Italian centenarians reveal a complex remodeling of lipids, amino acids, and gut microbiota metabolism. PLoS ONE 2013, 8, e56564. [Google Scholar] [CrossRef]
- Tang, M.; Bolderson, E.; O’Byrne, K.J.; Richard, D.J. Tumor Hypoxia Drives Genomic Instability. Front. Cell Dev. Biol. 2021, 9, 626229. [Google Scholar] [CrossRef]
- Salmaninejad, A.; Ilkhani, K.; Marzban, H.; Navashenaq, J.G.; Rahimirad, S.; Radnia, F.; Yousefi, M.; Bahmanpour, Z.; Azhdari, S.; Sahebkar, A. Genomic Instability in Cancer: Molecular Mechanisms and Therapeutic Potentials. Curr. Pharm. Des. 2021, 27, 3161–3169. [Google Scholar] [CrossRef]
- Li, H.; Zimmerman, S.E.; Weyemi, U. Genomic instability and metabolism in cancer. Int. Rev. Cell Mol. Biol. 2021, 364, 241–265. [Google Scholar]
- Cardoso, A.P.F.; Banerjee, M.; Nail, A.N.; Lykoudi, A.; States, J.C. miRNA dysregulation is an emerging modulator of genomic instability. Semin. Cancer Biol. 2021, 76, 120–131. [Google Scholar] [CrossRef]
- De Majo, F.; Martens, L.; Hegenbarth, J.C.; Rühle, F.; Hamczyk, M.R.; Nevado, R.M.; Andrés, V.; Hilbold, E.; Bär, C.; Thum, T.; et al. Genomic instability in the naturally and prematurely aged myocardium. Proc. Natl. Acad. Sci. USA 2021, 118, e2022974118. [Google Scholar] [CrossRef]
- Bonora, M.; Missiroli, S.; Perrone, M.; Fiorica, F.; Pinton, P.; Giorgi, C. Mitochondrial Control of Genomic Instability in Cancer. Cancers 2021, 13, 1914. [Google Scholar] [CrossRef]
- Keefe, D.L. Telomeres and genomic instability during early development. Eur. J. Med. Genet. 2020, 63, 103638. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.O.; Gartner, J.; Rangel-Pozzo, A.; Mai, S. Genomic Instability in Circulating Tumor Cells. Cancers 2020, 12, 3001. [Google Scholar] [CrossRef]
- Duijf, P.H.; Nanayakkara, D.; Nones, K.; Srihari, S.; Kalimutho, M.; Khanna, K.K. Mechanisms of Genomic Instability in Breast Cancer. Trends Mol. Med. 2019, 25, 595–611. [Google Scholar] [CrossRef] [PubMed]
- Tubbs, A.; Nussenzweig, A. Endogenous DNA Damage as a Source of Genomic Instability in Cancer. Cell 2017, 168, 644–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andor, N.; Maley, C.C.; Ji, H.P. Genomic Instability in Cancer: Teetering on the Limit of Tolerance. Cancer Res. 2017, 77, 2179–2185. [Google Scholar] [CrossRef] [Green Version]
- Negrini, S.; Gorgoulis, V.G.; Halazonetis, T.D. Genomic instability—An evolving hallmark of cancer. Nat. Rev. Mol. Cell Biol. 2010, 11, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Reece, A.S.; Hulse, G.K. Geotemporospatial and causal inference epidemiological analysis of US survey and overview of cannabis, cannabidiol and cannabinoid genotoxicity in relation to congenital anomalies 2001–2015. BMC Pediatr. 2022, 22, 47–124. [Google Scholar] [CrossRef]
- Reece, A.S.; Hulse, G.K. A geospatiotemporal and causal inference epidemiological exploration of substance and cannabinoid exposure as drivers of rising US pediatric cancer rates. BMC Cancer 2021, 21, 197–230. [Google Scholar] [CrossRef]
- Reece AS, Hulse GK: Cannabinoid exposure as a major driver of pediatric acute lymphoid Leukaemia rates across the USA: Combined geospatial, multiple imputation and causal inference study. BMC Cancer 2021, 21, 984–1017. [CrossRef]
- Reece, A.S.; Hulse, G.K. Co-occurrence across time and space of drug- and cannabinoid- exposure and adverse mental health outcomes in the National Survey of Drug Use and Health: Combined geotemporospatial and causal inference analysis. BMC Public Health 2020, 20, 1655–1669. [Google Scholar] [CrossRef] [PubMed]
- Reece, A.S.; Hulse, G.K. Contemporary epidemiology of rising atrial septal defect trends across USA 1991–2016: A combined ecological geospatiotemporal and causal inferential study. BMC Pediatr. 2020, 20, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Reece, A.S.; Hulse, G.K. Canadian Cannabis Consumption and Patterns of Congenital Anomalies: An Ecological Geospatial Analysis. J. Addict. Med. 2020, 14, e195–e210. [Google Scholar] [CrossRef] [PubMed]
- Reece, A.S.; Hulse, G.K. Broad Spectrum epidemiological contribution of cannabis and other substances to the teratological profile of northern New South Wales: Geospatial and causal inference analysis. BMC Pharmacol. Toxicol. 2020, 21, 75–103. [Google Scholar] [CrossRef]
- Reece, A.S.; Hulse, G.K. Cannabis Consumption Patterns Explain the East-West Gradient in Canadian Neural Tube Defect Incidence: An Ecological Study. Glob. Pediatr. Health 2019, 6, 2333794X19894798. [Google Scholar] [CrossRef] [Green Version]
- Reece, A.S.; Hulse, G.K. Cannabis Teratology Explains Current Patterns of Coloradan Congenital Defects: The Contribution of Increased Cannabinoid Exposure to Rising Teratological Trends. Clin. Pediatr. 2019, 58, 1085–1123. [Google Scholar] [CrossRef]
- Reece, A.S.; Hulse, G.K. Geotemporospatial and Causal Inferential Epidemiological Overview and Survey of USA Cannabis, Cannabidiol and Cannabinoid Genotoxicity Expressed in Cancer Incidence 2003–2017: Part 3—Spatiotemporal, Multivariable and Causal Inferential Pathfinding and Exploratory Analyses of Prostate and Ovarian Cancers. Arch. Public Health 2022, 80, 100–136. [Google Scholar]
- Reece, A.S.; Hulse, G.K. Geotemporospatial and Causal Inferential Epidemiological Overview and Survey of USA Cannabis, Cannabidiol and Cannabinoid Genotoxicity Expressed in Cancer Incidence 2003–2017: Part 2—Categorical Bivariate Analysis and Attributable Fractions. Arch. Public Health 2022, 80, 100–135. [Google Scholar] [CrossRef]
- Reece, A.S.; Hulse, G.K. Geotemporospatial and Causal Inferential Epidemiological Overview and Survey of USA Cannabis, Cannabidiol and Cannabinoid Genotoxicity Expressed in Cancer Incidence 2003–2017: Part 1—Continuous Bivariate Analysis. Arch. Public Health 2022, 80, 99–133. [Google Scholar] [CrossRef]
- Reece, A.S.; Hulse, G.K. Cannabinoid- and Substance- Relationships of European Congenital Anomaly Patterns: A Space-Time Panel Regression and Causal Inferential Study. Environ. Epigenetics 2022, 8, dvab015. [Google Scholar] [CrossRef]
- Reece, A.S.; Hulse, G.K. Cannabis in Pregnancy—Rejoinder, Exposition and Cautionary Tales. Psychiatric Times, 9 October 2020. Available online: https://www.psychiatrictimes.com/view/cannabis-pregnancy-rejoinder-exposition-cautionary-tales (accessed on 1 November 2022).
- Reece, A.S.; Hulse, G.K. Epidemiological Associations of Various Substances and Multiple Cannabinoids with Autism in USA. Clin. Pediatr. Open Access 2019, 4, 155. [Google Scholar] [CrossRef] [Green Version]
- Forrester, M.B.; Merz, R.D. Risk of selected birth defects with prenatal illicit drug use, Hawaii, 1986–2002. J. Toxicol. Environ. Health A 2007, 70, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Wagner, B.; Gerletti, P.; Fürst, P.; Keuth, O.; Bernsmann, T.; Martin, A.; Schäfer, B.; Numata, J.; Lorenzen, M.C.; Pieper, R. Transfer of cannabinoids into the milk of dairy cows fed with industrial hemp could lead to Δ9-THC exposure that exceeds acute reference dose. Nature Food 2022, 3, 921–932. [Google Scholar] [CrossRef]
- Reece, A.S.; Hulse, G.K. Cannabinoid Genotoxicity and Congenital Anomalies: A Convergent Synthesis of European and USA Datasets. In Cannabis, Cannabinoids and Endocannabinoids; Preedy, V., Patel, V., Eds.; Elsevier: London, UK, 2022; Volume 1, in press. [Google Scholar]
- Reece, A.S.; Hulse, G.K. Cannabis Genotoxicity and Cancer Incidence: A Highly Concordant Synthesis of European and USA Datasets. In Cannabis, Cannabinoids and Endocannabinoids; Preedy, V., Patel, V., Eds.; Elsevier: London, UK, 2022; Volume 1, in press. [Google Scholar]
- Reece, A.S.; Hulse, G.K. Epidemiological Overview of Cannabis- and Substance- Carcinogenesis in Europe: A Lagged Causal Inferential Panel Regression Modelling and Marginal Effects Study. 2022; Manuscript Submitted. [Google Scholar]
- Fine, J.D.; Moreau, A.L.; Karcher, N.R.; Agrawal, A.; Rogers, C.E.; Barch, D.M.; Bogdan, R. Association of Prenatal Cannabis Exposure with Psychosis Proneness Among Children in the Adolescent Brain Cognitive Development (ABCD) Study. JAMA Psychiatry 2019, 76, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Grech, A.; Van Os, J.; Jones, P.B.; Lewis, S.W.; Murray, R.M. Cannabis use and outcome of recent onset psychosis. Eur. Psychiatry 2005, 20, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Henquet, C.; Murray, R.; Linszen, D.; van Os, J. The environment and schizophrenia: The role of cannabis use. Schizophr. Bull. 2005, 31, 608–612. [Google Scholar] [CrossRef] [Green Version]
- Bartoli, F.; Crocamo, C.; Carra, G. Cannabis use disorder and suicide attempts in bipolar disorder: A meta-analysis. Neurosci. Biobehav. Rev. 2019, 103, 14–20. [Google Scholar] [CrossRef]
- Hanna, R.C.; Perez, J.M.; Ghose, S. Cannabis and development of dual diagnoses: A literature review. Am. J. Drug Alcohol Abus 2017, 43, 442–455. [Google Scholar] [CrossRef]
- Marwaha, S.; Winsper, C.; Bebbington, P.; Smith, D. Cannabis Use and Hypomania in Young People: A Prospective Analysis. Schizophr. Bull. 2018, 44, 1267–1274. [Google Scholar] [CrossRef]
- Tyler, E.; Jones, S.; Black, N.; Carter, L.A.; Barrowclough, C. The relationship between bipolar disorder and cannabis use in daily life: An experience sampling study. PLoS ONE 2015, 10, e0118916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dierker, L.; Selya, A.; Lanza, S.; Li, R.; Rose, J. Depression and marijuana use disorder symptoms among current marijuana users. Addict. Behav. 2018, 76, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Filbey, F.M.; Aslan, S.; Lu, H.; Peng, S.L. Residual Effects of THC via Novel Measures of Brain Perfusion and Metabolism in a Large Group of Chronic Cannabis Users. Neuropsychopharmacology 2018, 43, 700–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobbi, G.; Atkin, T.; Zytynski, T.; Wang, S.; Askari, S.; Boruff, J.; Ware, M.; Marmorstein, N.; Cipriani, A.; Dendukuri, N.; et al. Association of Cannabis Use in Adolescence and Risk of Depression, Anxiety, and Suicidality in Young Adulthood: A Systematic Review and Meta-analysis Cannabis Use in Adolescence and Risk of Depression, Anxiety, and Suicidality in Young AdulthoodCannabis Use in Adolescence and Risk of Depression, Anxiety, and Suicidality in Young Adulthood. JAMA Psychiatry 2019, 76, 426–434. [Google Scholar] [PubMed]
- Hézode, C.; Roudot-Thoraval, F.; Nguyen, S.; Grenard, P.; Julien, B.; Zafrani, E.S.; Pawlostky, J.M.; Dhumeaux, D.; Lotersztajn, S.; Mallat, A. Daily cannabis smoking as a risk factor for progression of fibrosis in chronic hepatitis C. Hepatology 2005, 42, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Patsenker, E.; Stickel, F. Cannabinoids in liver diseases. Clin. Liver Dis. 2016, 7, 21–25. [Google Scholar] [CrossRef]
- Mai, P.; Yang, L.; Tian, L.; Wang, L.; Jia, S.; Zhang, Y.; Liu, X.; Yang, L.; Li, L. Endocannabinoid System Contributes to Liver Injury and Inflammation by Activation of Bone Marrow-Derived Monocytes/Macrophages in a CB1-Dependent Manner. J. Immunol. 2015, 195, 3390–3401. [Google Scholar] [CrossRef] [Green Version]
- Patsenker, E.; Stoll, M.; Millonig, G.; Agaimy, A.; Wissniowski, T.; Schneider, V.; Mueller, S.; Brenneisen, R.; Seitz, H.K.; Ocker, M.; et al. Cannabinoid receptor type I modulates alcohol-induced liver fibrosis. Mol. Med. 2011, 17, 1285–1294. [Google Scholar] [CrossRef]
- Barber, P.A. Cannabis and Stroke. In Handbook of Cannabis and Related Pathologies: Biology, Pharmacology and Treatment; Preedy, V.R., Ed.; Academic Press: New York, NY, USA, 2017; Volume 1, pp. 486–493. [Google Scholar]
- Menahem, S. Cardiovascular Effects of Cannabis Usage. In Handbook of Cannabis and Related Pathologies: Biology, Pharmacology and Treatment; Preedy, V.R. Academic Press: New York, NY, USA, 2017; Volume 1, pp. 481–485. [Google Scholar]
- Volkow, N.D.; Compton, W.M.; Weiss, S.R. Adverse health effects of marijuana use. N. Engl. J. Med. 2014, 371, 878–879. [Google Scholar] [CrossRef] [Green Version]
- Rossato, M.; Ion Popa, F.; Ferigo, M.; Clari, G.; Foresta, C. Human sperm express cannabinoid receptor Cb1, the activation of which inhibits motility, acrosome reaction, and mitochondrial function. J. Clin. Endocrinol. Metab. 2005, 90, 984–991. [Google Scholar] [CrossRef]
- Mon, M.J.; Haas, A.E.; Stein, J.L.; Stein, G.S. Influence of psychoactive and nonpsychoactive cannabinoids on chromatin structure and function in human cells. Biochem. Pharmacol. 1981, 30, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yuan, W.; Li, M.D. Genes and pathways co-associated with the exposure to multiple drugs of abuse, including alcohol, amphetamine/methamphetamine, cocaine, marijuana, morphine, and/or nicotine: A review of proteomics analyses. Mol. Neurobiol. 2011, 44, 269–286. [Google Scholar] [CrossRef] [PubMed]
- Sarafian, T.A.; Habib, N.; Oldham, M.; Seeram, N.; Lee, R.P.; Lin, L.; Tashkin, D.P.; Roth, M.D. Inhaled marijuana smoke disrupts mitochondrial energetics in pulmonary epithelial cells in vivo. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 290, L1202–L1209. [Google Scholar] [CrossRef] [PubMed]
- Sarafian, T.A.; Kouyoumjian, S.; Khoshaghideh, F.; Tashkin, D.P.; Roth, M.D. Delta 9-tetrahydrocannabinol disrupts mitochondrial function and cell energetics. Am. J. Physiol. Lung Cell Mol. Physiol. 2003, 284, L298–L306. [Google Scholar] [CrossRef] [Green Version]
- Bénard, G.; Massa, F.; Puente, N.; Lourenço, J.; Bellocchio, L.; Soria-Gómez, E.; Matias, I.; Delamarre, A.; Metna-Laurent, M.; Cannich, A.; et al. Mitochondrial CB(1) receptors regulate neuronal energy metabolism. Nat. Neurosci. 2012, 15, 558–564. [Google Scholar] [CrossRef]
- Koch, M.; Varela, L.; Kim, J.G.; Kim, J.D.; Hernández-Nuño, F.; Simonds, S.E.; Castorena, C.M.; Vianna, C.R.; Elmquist, J.K.; Morozov, Y.M.; et al. Hypothalamic POMC neurons promote cannabinoid-induced feeding. Nature 2015, 519, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Wolff, V.; Schlagowski, A.I.; Rouyer, O.; Charles, A.L.; Singh, F.; Auger, C.; Schini-Kerth, V.; Marescaux, C.; Raul, J.S.; Zoll, J.; et al. Tetrahydrocannabinol induces brain mitochondrial respiratory chain dysfunction and increases oxidative stress: A potential mechanism involved in cannabis-related stroke. Biomed. Res. Int. 2015, 2015, 323706. [Google Scholar] [CrossRef] [Green Version]
- Canto, C.; Menzies, K.J.; Auwerx, J. NAD(+) Metabolism and the Control of Energy Homeostasis: A Balancing Act between Mitochondria and the Nucleus. Cell Metab. 2015, 22, 31–53. [Google Scholar] [CrossRef] [Green Version]
- Bárcena, C.; Mayoral, P.; Quirós, P.M. Mitohormesis, an Antiaging Paradigm. Int. Rev. Cell Mol. Biol. 2018, 340, 35–77. [Google Scholar]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, oxidants, and aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Kwong, J.M.; Caprioli, J.; Piri, N. DNA and RNA oxidative damage in the retina is associated with ganglion cell mitochondria. Sci. Rep. 2022, 12, 8705. [Google Scholar] [CrossRef] [PubMed]
- Krammer, U.D.B.; Sommer, A.; Tschida, S.; Mayer, A.; Lilja, S.V.; Switzeny, O.J.; Hippe, B.; Rust, P.; Haslberger, A.G. PGC-1α Methylation, miR-23a, and miR-30e Expression as Biomarkers for Exercise- and Diet-Induced Mitochondrial Biogenesis in Capillary Blood from Healthy Individuals: A Single-Arm Intervention. Sports 2022, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.K.S.; Wee, H.N.; Ching, J.; Kovalik, J.P.; Chan, A.W.; Matchar, D.B. Plasma acylcarnitines as metabolic signatures of declining health-related quality of life measure in community-dwelling older adults: A combined cross-sectional and longitudinal pilot study. J. Gerontol. A Biol. Sci. Med. Sci. 2022, 131, glac114. [Google Scholar] [CrossRef] [PubMed]
- Teng, H.; Hong, Y.; Cao, J.; Li, H.; Tian, F.; Sun, J.; Wen, K.; Han, G.; Whelchel, A.; Zhang, X.; et al. Senescence marker protein30 protects lens epithelial cells against oxidative damage by restoring mitochondrial function. Bioengineered 2022, 13, 12955–12971. [Google Scholar] [CrossRef] [PubMed]
- Wan, W.; Hua, F.; Fang, P.; Li, C.; Deng, F.; Chen, S.; Ying, J.; Wang, X. Regulation of Mitophagy by Sirtuin Family Proteins: A Vital Role in Aging and Age-Related Diseases. Front. Aging Neurosci. 2022, 14, 845330. [Google Scholar] [CrossRef]
- Zhong, W.; Rao, Z.; Xu, J.; Sun, Y.; Hu, H.; Wang, P.; Xia, Y.; Pan, X.; Tang, W.; Chen, Z.; et al. Defective mitophagy in aged macrophages promotes mitochondrial DNA cytosolic leakage to activate STING signaling during liver sterile inflammation. Aging Cell 2022, 21, e13622. [Google Scholar] [CrossRef]
- Hadley, E.C.; Lakatta, E.G.; Morrison-Bogorad, M.; Warner, H.R.; Hodes, R.J. The future of aging therapies. Cell 2005, 120, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Kirkwood, T.B. Understanding the odd science of aging. Cell 2005, 120, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Schrott, R.; Acharya, K.; Itchon-Ramos, N.; Hawkey, A.B.; Pippen, E.; Mitchell, J.T.; Kollins, S.H.; Levin, E.D.; Murphy, S.K. Cannabis use is associated with potentially heritable widespread changes in autism candidate gene DLGAP2 DNA methylation in sperm. Epigenetics 2019, 15, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Schrott, R.; Murphy, S.K. Cannabis use and the sperm epigenome: A budding concern? Environ. Epigenetics 2020, 6, dvaa002. [Google Scholar] [CrossRef] [Green Version]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C.; et al. Neuroinflammation and depression: A review. Eur. J. Neurosci. 2021, 53, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Buckley, P.F. Neuroinflammation and Schizophrenia. Curr. Psychiatry Rep. 2019, 21, 72. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, F.; Aggio, V.; Pratesi, M.L.; Greco, G.; Furlan, R. Neuroinflammation in Bipolar Depression. Front. Psychiatry 2020, 11, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najjar, S.; Pearlman, D.M.; Alper, K.; Najjar, A.; Devinsky, O. Neuroinflammation and psychiatric illness. J. Neuroinflammation 2013, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Imbeault, S.; Goiny, M.; Liu, X.; Erhardt, S. Effects of IDO1 and TDO2 inhibition on cognitive deficits and anxiety following LPS-induced neuroinflammation. Acta Neuropsychiatr. 2020, 32, 43–53. [Google Scholar] [CrossRef]
- Paiva, I.H.R.; Duarte-Silva, E.; Peixoto, C.A. The role of prebiotics in cognition, anxiety, and depression. Eur. Neuropsychopharmacol. 2020, 34, 1–18. [Google Scholar] [CrossRef]
- Xu, Y.; Sheng, H.; Bao, Q.; Wang, Y.; Lu, J.; Ni, X. NLRP3 inflammasome activation mediates estrogen deficiency-induced depression- and anxiety-like behavior and hippocampal inflammation in mice. Brain Behav. Immun. 2016, 56, 175–186. [Google Scholar] [CrossRef]
- Zheng, Z.H.; Tu, J.L.; Li, X.H.; Hua, Q.; Liu, W.Z.; Liu, Y.; Pan, B.X.; Hu, P.; Zhang, W.H. Neuroinflammation induces anxiety- and depressive-like behavior by modulating neuronal plasticity in the basolateral amygdala. Brain Behav. Immun. 2021, 91, 505–518. [Google Scholar] [CrossRef]
- Cai, Z.; Hussain, M.D.; Yan, L.J. Microglia, neuroinflammation, and beta-amyloid protein in Alzheimer’s disease. Int. J. Neurosci. 2014, 124, 307–321. [Google Scholar] [CrossRef]
- Calsolaro, V.; Edison, P. Neuroinflammation in Alzheimer’s disease: Current evidence and future directions. Alzheimers Dement. 2016, 12, 719–732. [Google Scholar] [CrossRef]
- Leng, F.; Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: Where do we go from here? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Zheng, L.J.; Zhang, L.J. Neuroinflammation, Gut Microbiome, and Alzheimer’s Disease. Mol. Neurobiol. 2018, 55, 8243–8250. [Google Scholar] [CrossRef] [PubMed]
- Cannizzo, E.S.; Clement, C.C.; Sahu, R.; Follo, C.; Santambrogio, L. Oxidative stress, inflamm-aging and immunosenescence. J. Proteomics 2011, 74, 2313–2323. [Google Scholar] [CrossRef] [PubMed]
- Reece, A.S.; Hulse, G.K. Effect of Cannabis Legalization on US Autism Incidence and Medium Term Projections. Clin. Pediatr. Open Access 2019, 4, 154. [Google Scholar] [CrossRef] [Green Version]
- Reece, A.S.; Hulse, G.K. Impact of Converging Sociocultural and Substance-Related Trends on US Autism Rates: Combined Geospatiotemporal and Causal Inferential Analysis. Eur. Arch. Psychiatry Clinial Neurosci. 2022, 19, 7726–7752. [Google Scholar] [CrossRef] [PubMed]
- Schrott, R.; Modliszewski, J.L.; Hawkey, A.B.; Grenier, C.; Holloway, Z.; Evans, J.; Pippen, E.; Corcoran, D.L.; Levin, E.D.; Murphy, S.K. Sperm DNA methylation alterations from cannabis extract exposure are evident in offspring. Epigenetics Chromatin 2022, 15, 33. [Google Scholar] [CrossRef] [PubMed]
- Schrott, R.; Greeson, K.W.; King, D.; Symosko Crow, K.M.; Easley IV, C.A.; Murphy, S.K. Cannabis alters DNA methylation at maternally imprinted and autism candidate genes in spermatogenic cells. Syst. Biol. Reprod. Med. 2022, 68, 357–369. [Google Scholar] [CrossRef]
- Mona, K.; Ntlantsana, V.; Tomita, A.M.; Paruk, S. Prevalence of cannabis use in people with psychosis in KwaZulu-Natal, South Africa. S. Afr. J. Psychiatry 2022, 28, 1927. [Google Scholar] [CrossRef]
- Mallet, J.; Godin, O.; Dansou, Y.; Mazer, N.; Scognamiglio, C.; Berna, F.; Boyer, L.; Capdevielle, D.; Chéreau, I.; D’Amato, T.; et al. Current (but not ex) cigarette smoking is associated with worse cognitive performances in schizophrenia: Results from the FACE-SZ cohort. Psychol. Med. 2022, 1–12. [Google Scholar] [CrossRef]
- Little, R.; D’Mello, D. A Cannabinoid Hypothesis of Schizophrenia: Pathways to Psychosis. Innov. Clin. Neurosci. 2022, 19, 38–43. [Google Scholar]
- Kayir, H.; Ruffolo, J.; McCunn, P.; Khokhar, J.Y. The Relationship Between Cannabis, Cognition, and Schizophrenia: It’s Complicated. In Current Topics in Behavioral Neurosciences; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Ibarra-Lecue, I.; Unzueta-Larrinaga, P.; Barrena-Barbadillo, R.; Villate, A.; Horrillo, I.; Mendivil, B.; Landabaso, M.A.; Meana, J.J.; Etxebarria, N.; Callado, L.F.; et al. Cannabis use selectively modulates circulating biomarkers in the blood of schizophrenia patients. Addict. Biol. 2022, 27, e13233. [Google Scholar] [CrossRef] [PubMed]
- Greco, L.A.; Reay, W.R.; Dayas, C.V.; Cairns, M.J. Pairwise genetic meta-analyses between schizophrenia and substance dependence phenotypes reveals novel association signals with pharmacological significance. Transl. Psychiatry 2022, 12, 403. [Google Scholar] [CrossRef] [PubMed]
- Fusar-Poli, L.; Pries, L.K.; van Os, J.; Radhakrishnan, R.; Pençe, A.Y.; Erzin, G.; Delespaul, P.; Kenis, G.; Luykx, J.J.; Lin, B.D.; et al. The association between cannabis use and facial emotion recognition in schizophrenia, siblings, and healthy controls: Results from the EUGEI study. Eur. Neuropsychopharmacol. 2022, 63, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Dennen, C.A.; Blum, K.; Bowirrat, A.; Khalsa, J.; Thanos, P.K.; Baron, D.; Badgaiyan, R.D.; Gupta, A.; Braverman, E.R.; Gold, M.S. Neurogenetic and Epigenetic Aspects of Cannabinoids. Epigenomes 2022, 6, 27. [Google Scholar] [CrossRef]
- Del Re, E.C. Emerging themes in schizophrenia research at SIRS 2022: Inclusivity, equality and Cannabis impact on mental health. Schizophr. Res. 2022, 250, 39–40. [Google Scholar] [CrossRef]
- Crawford, P.; Go, K.V. Schizophrenia. Am. Fam. Physician 2022, 106, 388–396. [Google Scholar]
- Argote, M.; Sescousse, G.; Brunelin, J.; Fakra, E.; Nourredine, M.; Rolland, B. Association between formal thought disorder and cannabis use: A systematic review and meta-analysis. Schizophrenia 2022, 8, 78. [Google Scholar] [CrossRef]
- Pearson, N.T.; Berry, J.H. Cannabis and Psychosis Through the Lens of DSM-5. Int. J. Environ. Res. Public Health 2019, 16, 4149. [Google Scholar] [CrossRef] [Green Version]
- Vaucher, J.; Keating, B.J.; Lasserre, A.M.; Gan, W.; Lyall, D.M.; Ward, J.; Smith, D.J.; Pell, J.P.; Sattar, N.; Paré, G.; et al. Cannabis use and risk of schizophrenia: A Mendelian randomization study. Mol. Psychiatry 2018, 23, 1287–1292. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, A.A.; Salama, M.; Peak, R.; Tindle, R.; Salem, A.; Keri, S.; Misiak, B.; Frydecka, D.; Mohamed, W. Interactions between cannabis and schizophrenia in humans and rodents. Rev. Neurosci. 2017, 28, 811–823. [Google Scholar] [CrossRef]
- Marconi, A.; Di Forti, M.; Lewis, C.M.; Murray, R.M.; Vassos, E. Meta-analysis of the Association Between the Level of Cannabis Use and Risk of Psychosis. Schizophr. Bull. 2016, 42, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Gage, S.H.; Hickman, M.; Zammit, S. Association Between Cannabis and Psychosis: Epidemiologic Evidence. Biol. Psychiatry 2016, 79, 549–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pushpa-Rajah, J.A.; McLoughlin, B.C.; Gillies, D.; Rathbone, J.; Variend, H.; Kalakouti, E.; Kyprianou, K. Cannabis and schizophrenia. Schizophr. Bull. 2015, 41, 336–337. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, D.C.; Sewell, R.A.; Ranganathan, M. Cannabis and psychosis/schizophrenia: Human studies. Eur. Arch. Psychiatry Clin. Neurosci. 2009, 259, 413–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andréasson, S.; Engström, A.; Allebeck, P.; Rydberg, U. Cannabis and schizophrenia. A longitudinal study of Swedish conscripts. Lancet 1987, 2, 1483–1486. [Google Scholar] [CrossRef]
- Reece, A.S.; Thomas, M.R.; Norman, A.; Hulse, G.K. Dramatic acceleration of reproductive aging, contraction of biochemical fecundity and healthspan-lifespan implications of opioid-induced endocrinopathy-FSH/LH ratio and other interrelationships. Reprod. Toxicol. 2016, 66, 20–30. [Google Scholar] [CrossRef]
- Ruth, K.S.; Day, F.R.; Hussain, J.; Martínez-Marchal, A.; Aiken, C.E.; Azad, A.; Thompson, D.J.; Knoblochova, L.; Abe, H.; Tarry-Adkins, J.L.; et al. Genetic insights into biological mechanisms governing human ovarian ageing. Nature 2021, 596, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Berta, D.G.; Kuisma, H.; Välimäki, N.; Räisänen, M.; Jäntti, M.; Pasanen, A.; Karhu, A.; Kaukomaa, J.; Taira, A.; Cajuso, T.; et al. Deficient H2A.Z deposition is associated with genesis of uterine leiomyoma. Nature 2021, 596, 398–403. [Google Scholar] [CrossRef]
- Partridge, L.; Gems, D.; Withers, D.J. Sex and death: What is the connection? Cell 2005, 120, 461–472. [Google Scholar] [CrossRef]
- Kaymak, I.; Williams, K.S.; Cantor, J.R.; Jones, R.G. Immunometabolic Interplay in the Tumor Microenvironment. Cancer Cell 2021, 39, 28–37. [Google Scholar] [CrossRef]
- Spadaro, O.; Youm, Y.; Shchukina, I.; Ryu, S.; Sidorov, S.; Ravussin, A.; Nguyen, K.; Aladyeva, E.; Predeus, A.N.; Smith, S.R.; et al. Caloric restriction in humans reveals immunometabolic regulators of health span. Science 2022, 375, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Lombard, D.B.; Chua, K.F.; Mostoslavsky, R.; Franco, S.; Gostissa, M.; Alt, F.W. DNA repair, genome stability, and aging. Cell 2005, 120, 497–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gröbner, S.N.; Worst, B.C.; Weischenfeldt, J.; Buchhalter, I.; Kleinheinz, K.; Rudneva, V.A.; Johann, P.D.; Balasubramanian, G.P.; Segura-Wang, M.; Brabetz, S.; et al. The landscape of genomic alterations across childhood cancers. Nature 2018, 555, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Liu, Y.U.; Liu, Y.; Alexandrov, L.B.; Edmonson, M.N.; Gawad, C.; Zhou, X.; Li, Y.; Rusch, M.C.; Easton, J.; et al. Pan-cancer genome and transcriptome analyses of 1,699 paediatric leukaemias and solid tumours. Nature 2018, 555, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Trivers, K.F.; Mertens, A.C.; Ross, J.A.; Steinbuch, M.; Olshan, A.F.; Robison, L.L. Children’s Cancer G: Parental marijuana use and risk of childhood acute myeloid leukaemia: A report from the Children’s Cancer Group (United States and Canada). Paediatr. Perinat. Epidemiol. 2006, 20, 110–118. [Google Scholar] [CrossRef]
- Grufferman, S.; Schwartz, A.G.; Ruymann, F.B.; Maurer, H.M. Parents’ use of cocaine and marijuana and increased risk of rhabdomyosarcoma in their children. Cancer Causes Control 1993, 4, 217–224. [Google Scholar] [CrossRef]
- Kuijten, R.R.; Bunin, G.R.; Nass, C.C.; Meadows, A.T. Gestational and familial risk factors for childhood astrocytoma: Results of a case-control study. Cancer Res. 1990, 50, 2608–2612. [Google Scholar]
- Carlson, B.M. Human Embryology and Developmental Biology, 6 th ed.; Elsevier: Pennsylvania, PA, USA, 2019; Volume 1. [Google Scholar]
- Gill, S.K.; Broussard, C.; Devine, O.; Green, R.F.; Rasmussen, S.A.; Reefhuis, J.; National Birth Defects Prevention Study. Association between maternal age and birth defects of unknown etiology: United States, 1997–2007. Birth Defects Res. A Clin. Mol. Teratol. 2012, 94, 1010–1018. [Google Scholar] [CrossRef] [Green Version]
- Hussein, N.A.E.M.; El-Toukhy, M.A.E.F.; Kazem, A.H.; Ali, M.E.S.; Ahmad, M.A.E.R.; Ghazy, H.M.R.; El-Din, A.M.G. Protective and therapeutic effects of cannabis plant extract on liver cancer induced by dimethylnitrosamine in mice. Alex. J. Med. 2014, 50, 241–251. [Google Scholar] [CrossRef]
- Arendt, M.; Munk-Jørgensen, P.; Sher, L.; Jensen, S.O. Mortality among individuals with cannabis, cocaine, amphetamine, MDMA, and opioid use disorders: A nationwide follow-up study of Danish substance users in treatment. Drug Alcohol Depend. 2011, 114, 134–139. [Google Scholar] [CrossRef]
- Calabria, B.; Degenhardt, L.; Hall, W.; Lynskey, M. Does cannabis use increase the risk of death? Systematic review of epidemiological evidence on adverse effects of cannabis use. Drug Alcohol Rev. 2010, 29, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, R.C.; Cunningham, J.K.; Verdichevski, M.; Sykes, J.; Jaffer, S.R.; Kish, S.J. All-cause mortality among individuals with disorders related to the use of methamphetamine: A comparative cohort study. Drug Alcohol Depend. 2012, 125, 290–294. [Google Scholar] [CrossRef]
- Davstad, I.; Allebeck, P.; Leifman, A.; Stenbacka, M.; Romelsjö, A. Self-reported drug use and mortality among a nationwide sample of Swedish conscripts—A 35-year follow-up. Drug Alcohol Depend. 2011, 118, 383–390. [Google Scholar] [CrossRef] [PubMed]
- DeFilippis, E.M.; Singh, A.; Divakaran, S.; Gupta, A.; Collins, B.L.; Biery, D.; Qamar, A.; Fatima, A.; Ramsis, M.; Pipilas, D.; et al. Cocaine and Marijuana Use among Young Adults Presenting with Myocardial Infarction: The Partners YOUNG-MI Registry. J. Am. Coll. Cardiol. 2018, in press.
- Desai, R.; Patel, U.; Sharma, S.; Amin, P.; Bhuva, R.; Patel, M.S.; Sharma, N.; Shah, M.; Patel, S.; Savani, S.; et al. Recreational Marijuana Use and Acute Myocardial Infarction: Insights from Nationwide Inpatient Sample in the United States. Cureus 2017, 9, e1816. [Google Scholar] [CrossRef]
- Fridell, M.; Bäckström, M.; Hesse, M.; Krantz, P.; Perrin, S.; Nyhlén, A. Prediction of psychiatric comorbidity on premature death in a cohort of patients with substance use disorders: A 42-year follow-up. BMC Psychiatry 2019, 19, 150. [Google Scholar] [CrossRef] [Green Version]
- Frost, L.; Mostofsky, E.; Rosenbloom, J.I.; Mukamal, K.J.; Mittleman, M.A. Marijuana use and long-term mortality among survivors of acute myocardial infarction. Am. Heart J. 2013, 165, 170–175. [Google Scholar] [CrossRef] [Green Version]
- Hser, Y.I.; Kagihara, J.; Huang, D.; Evans, E.; Messina, N. Mortality among substance-using mothers in California: A 10-year prospective study. Addiction 2012, 107, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Muhuri, P.K.; Gfroerer, J.C. Mortality associated with illegal drug use among adults in the United States. Am. J. Drug Alcohol Abus 2011, 37, 155–164. [Google Scholar] [CrossRef]
- Pavarin, R.M.; Berardi, D. Mortality risk in a cohort of subjects reported by authorities for cannabis possession for personal use. Results of a longitudinal study. Epidemiol. Prev. 2011, 35, 89–93. [Google Scholar]
- Von Greiff, N.; Skogens, L.; Berlin, M.; Bergmark, A. Mortality and Cause of Death-A 30-Year Follow-Up of Substance Misusers in Sweden. Subst. Use Misuse 2018, 53, 2043–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fergusson, D.M.; Boden, J.M.; Horwood, L.J. Cannabis use and other illicit drug use: Testing the cannabis gateway hypothesis. Addiction 2006, 101, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Secades-Villa, R.; Garcia-Rodríguez, O.; Jin, C.J.; Wang, S.; Blanco, C. Probability and predictors of the cannabis gateway effect: A national study. Int. J. Drug Policy 2015, 26, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Nkansah-Amankra, S.; Minelli, M. Gateway hypothesis” and early drug use: Additional findings from tracking a population-based sample of adolescents to adulthood. Prev. Med. Rep. 2016, 4, 134–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Association, A.P. Diagnostic and Statistical Manual of Mental Disorders (DSM-5); American Psychiatric Association: Washington, DC, USA, 2013; Volume 1. [Google Scholar]
- GeneCards: The Human Gene Database: UHRF1. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=Uhrf1 (accessed on 1 June 2022).
- Mudbhary, R.; Hoshida, Y.; Chernyavskaya, Y.; Jacob, V.; Villanueva, A.; Fiel, M.I.; Chen, X.; Kojima, K.; Thung, S.; Bronson, R.T.; et al. UHRF1 overexpression drives DNA hypomethylation and hepatocellular carcinoma. Cancer Cell 2014, 25, 196–209. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, Z.; Chen, J.; Liu, W.; Lai, W.; Liu, B.; Li, X.; Liu, L.; Xu, S.; Dong, Q.; et al. Stella safeguards the oocyte methylome by preventing de novo methylation mediated by DNMT1. Nature 2018, 564, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Parolia, A.; Qiao, Y.; Bawa, P.; Eyunni, S.; Mannan, R.; Carson, S.E.; Chang, Y.; Wang, X.; Zhang, Y.; et al. Targeting SWI/SNF ATPases in enhancer-addicted prostate cancer. Nature 2022, 601, 434–439. [Google Scholar] [CrossRef]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Chi, Y.; Sauve, A.A. Nicotinamide riboside, a trace nutrient in foods, is a vitamin B3 with effects on energy metabolism and neuroprotection. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 657–661. [Google Scholar] [CrossRef]
- Hollis, F.; van der Kooij, M.A.; Zanoletti, O.; Lozano, L.; Cantó, C.; Sandi, C. Mitochondrial function in the brain links anxiety with social subordination. Proc. Natl. Acad. Sci. USA 2015, 112, 15486–15491. [Google Scholar] [CrossRef] [Green Version]
- Gómez, C.A.; Sutin, J.; Wu, W.; Fu, B.; Uhlirova, H.; Devor, A.; Boas, D.A.; Sakadžić, S.; Yaseen, M.A. Phasor analysis of NADH FLIM identifies pharmacological disruptions to mitochondrial metabolic processes in the rodent cerebral cortex. PLoS ONE 2018, 13, e0194578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.; Kim, H.R.; Shin, M.G. Rejuvenating Aged Hematopoietic Stem Cells Through Improvement of Mitochondrial Function. Ann. Lab. Med. 2018, 38, 395–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidd, P.M. Neurodegeneration from mitochondrial insufficiency: Nutrients, stem cells, growth factors, and prospects for brain rebuilding using integrative management. Altern. Med. Rev. 2005, 10, 268–293. [Google Scholar]
- Cimadamore, F.; Curchoe, C.L.; Alderson, N.; Scott, F.; Salvesen, G.; Terskikh, A.V. Nicotinamide rescues human embryonic stem cell-derived neuroectoderm from parthanatic cell death. Stem Cells 2009, 27, 1772–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.Y.; Wang, F.; Zhang, X.Y.; Huang, P.; Lu, Y.B.; Wei, E.Q.; Zhang, W.P. Nicotinamide phosphoribosyltransferase may be involved in age-related brain diseases. PLoS ONE 2012, 7, e44933. [Google Scholar] [CrossRef] [Green Version]
- Griffin, S.M.; Pickard, M.R.; Orme, R.P.; Hawkins, C.P.; Fricker, R.A. Nicotinamide promotes neuronal differentiation of mouse embryonic stem cells in vitro. Neuroreport 2013, 24, 1041–1046. [Google Scholar] [CrossRef]
- Maruotti, J.; Sripathi, S.R.; Bharti, K.; Fuller, J.; Wahlin, K.J.; Ranganathan, V.; Sluch, V.M.; Berlinicke, C.A.; Davis, J.; Kim, C.; et al. Small-molecule-directed, efficient generation of retinal pigment epithelium from human pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2015, 112, 10950–10955. [Google Scholar] [CrossRef]
- Black, B.E. Preface to: Centromeres and Kinetochores. In Centromeres and Kinetochores; Black, B.E., Ed.; Springer: Cham, Switzerland, 2017; Volume 1, pp. v–viii. [Google Scholar]
- Sathananthan, A.H.; Kola, I.; Osborne, J.; Trounson, A.; Ng, S.C.; Bongso, A.; Ratnam, S. Centrioles in the beginning of human development. Proc. Natl. Acad. Sci. USA 1991, 88, 4806–4810. [Google Scholar] [CrossRef] [Green Version]
- Sathananthan, A.H. Mitosis in the human embryo: The vital role of the sperm centrosome (centriole). Histol. Histopathol. 1997, 12, 827–856. [Google Scholar]
- Blengini, C.S.; Schindler, K. Acentriolar spindle assembly in mammalian female meiosis and the consequences of its perturbations on human reproduction. Biol. Reprod. 2021, 106, 253–263. [Google Scholar] [CrossRef]
- Cavazza, T.; Takeda, Y.; Politi, A.Z.; Aushev, M.; Aldag, P.; Baker, C.; Choudhary, M.; Bucevičius, J.; Lukinavičius, G.; Elder, K.; et al. Parental genome unification is highly error-prone in mammalian embryos. Cell 2021, 184, 2860–2877.e2822. [Google Scholar] [CrossRef] [PubMed]
- Cavin-Meza, G.; Kwan, M.M.; Wignall, S.M. Multiple motors cooperate to establish and maintain acentrosomal spindle bipolarity in C. elegans oocyte meiosis. eLife 2022, 11, e72872. [Google Scholar] [CrossRef]
- So, C.; Menelaou, K.; Uraji, J.; Harasimov, K.; Steyer, A.M.; Seres, K.B.; Bucevičius, J.; Lukinavičius, G.; Möbius, W.; Sibold, C.; et al. Mechanism of spindle pole organization and instability in human oocytes. Science 2022, 375, eabj3944. [Google Scholar] [CrossRef] [PubMed]
- Tischer, T.; Yang, J.; Barford, D. The APC/C targets the Cep152-Cep63 complex at the centrosome to regulate mitotic spindle assembly. J. Cell Sci. 2022, 135, jcs259273. [Google Scholar] [CrossRef] [PubMed]
- Segbert, C.; Barkus, R.; Powers, J.; Strome, S.; Saxton, W.M.; Bossinger, O. KLP-18, a Klp2 kinesin, is required for assembly of acentrosomal meiotic spindles in Caenorhabditis elegans. Mol. Biol. Cell 2003, 14, 4458–4469. [Google Scholar] [CrossRef] [Green Version]
- Alberts, B.; Johnson, A.; Lewis, J.; Morgan, D.; Raff, M.; Roberts, K.; Walter, P. (Eds.) Molecular Biology of the Cell, 6th ed.; Garland Science: New York, NY, USA, 2014. [Google Scholar]
- Moutin, M.J.; Bosc, C.; Peris, L.; Andrieux, A. Tubulin post-translational modifications control neuronal development and functions. Dev. Neurobiol. 2021, 81, 253–272. [Google Scholar] [CrossRef]
- Janke, C.; Montagnac, G. Causes and Consequences of Microtubule Acetylation. Curr. Biol. 2017, 27, R1287–R1292. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Fukagawa, T. Critical Foundation of the Kinetochore: The Constitutive Centromere—Associated Network (CCAN). In Centromeres and Kinetochores; Black, B.E., Ed.; Springer: Philadelphia, PA, USA, 2017; Volume 1, pp. 1–554. [Google Scholar]
- French, B.T.; Straight, A.F. The Power of Xenopus Egg Extract for Reconstitution of Centromere and Kinetochore Function. In Centromeres and Kinetochores; Black, B.E., Ed.; Springer: Philadelphia, PA, USA, 2017; Volume 1, pp. 1–554. [Google Scholar]
- Grishchuk, E.L. Biophysics of Microtubule End Coupling at the Kinetochore. In Centromeres and Kinetochores; Black, B.E., Ed.; Springer: Philadelphia, PA, USA, 2017; Volume 1, pp. 1–554. [Google Scholar]
- Hsu, J.M.; Huang, J.; Meluh, P.B.; Laurent, B.C. The yeast RSC chromatin-remodeling complex is required for kinetochore function in chromosome segregation. Mol. Cell Biol. 2003, 23, 3202–3215. [Google Scholar] [CrossRef]
- Ryu, H.Y.; Hochstrasser, M. Histone sumoylation and chromatin dynamics. Nucleic Acids Res. 2021, 49, 6043–6052. [Google Scholar] [CrossRef]
- Gowran, A.; Murphy, C.E.; Campbell, V.A. Delta(9)-tetrahydrocannabinol regulates the p53 post-translational modifiers Murine double minute 2 and the Small Ubiquitin MOdifier protein in the rat brain. FEBS Lett. 2009, 583, 3412–3418. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, A.M.; Zimmerman, S.; Raj, A.Y. Effects of Cannabinoids on Spermatogensis in Mice. In Marijuana and Medicine, 1st ed.; Nahas, G.G., Sutin, K.M., Harvey, D.J., Agurell, S., Eds.; Humana Press: Totowa, NY, USA, 1999; Volume 1, pp. 347–358. [Google Scholar]
- Zimmerman, S.; Zimmerman, A.M. Genetic effects of marijuana. Int. J. Addict. 1990, 25, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Henrich, R.T.; Nogawa, T.; Morishima, A. In vitro induction of segregational errors of chromosomes by natural cannabinoids in normal human lymphocytes. Environ. Mutagen. 1980, 2, 139–147. [Google Scholar] [CrossRef] [PubMed]
- GeneCards: Down Syndrome Cell Adhesion Molecule. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=DSCAM (accessed on 1 April 2022).
- Grossman, T.R.; Gamliel, A.; Wessells, R.J.; Taghli-Lamallem, O.; Jepsen, K.; Ocorr, K.; Korenberg, J.R.; Peterson, K.L.; Rosenfeld, M.G.; Bodmer, R.; et al. Over-expression of DSCAM and COL6A2 cooperatively generates congenital heart defects. PLoS Genet. 2011, 7, e1002344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrell, V.; Cárdenas, A.; Ciceri, G.; Galcerán, J.; Flames, N.; Pla, R.; Nóbrega-Pereira, S.; García-Frigola, C.; Peregrín, S.; Zhao, Z.; et al. Slit/Robo signaling modulates the proliferation of central nervous system progenitors. Neuron 2012, 76, 338–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cárdenas, A.; Villalba, A.; de Juan Romero, C.; Picó, E.; Kyrousi, C.; Tzika, A.C.; Tessier-Lavigne, M.; Ma, L.; Drukker, M.; Cappello, S.; et al. Evolution of Cortical Neurogenesis in Amniotes Controlled by Robo Signaling Levels. Cell 2018, 174, 590–606.e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, M.L.; Gonda, Y.; Mommersteeg, M.T.; Barber, M.; Ypsilanti, A.R.; Hanashima, C.; Parnavelas, J.G.; Andrews, W.D. Robo1 modulates proliferation and neurogenesis in the developing neocortex. J. Neurosci. 2014, 34, 5717–5731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alpár, A.; Tortoriello, G.; Calvigioni, D.; Niphakis, M.J.; Milenkovic, I.; Bakker, J.; Cameron, G.A.; Hanics, J.; Morris, C.V.; Fuzik, J.; et al. Endocannabinoids modulate cortical development by configuring Slit2/Robo1 signalling. Nat. Commun. 2014, 5, 4421. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.R.; Zhao, H.T.; Park, J.M.; Dipoppa, M.; Monsalve-Mercado, M.M.; Dahan, J.B.; Rodgers, C.C.; Lejeune, A.; Hillman, E.; Miller, K.D.; et al. A human-specific modifier of cortical connectivity and circuit function. Nature 2021, 599, 640–644. [Google Scholar] [CrossRef]
- Shibata, M.; Pattabiraman, K.; Lorente-Galdos, B.; Andrijevic, D.; Kim, S.K.; Kaur, N.; Muchnik, S.K.; Xing, X.; Santpere, G.; Sousa, A.M.; et al. Regulation of prefrontal patterning and connectivity by retinoic acid. Nature 2021, 598, 483–488. [Google Scholar] [CrossRef]
- Scuteri, A.; Orru, M.; Morrell, C.; Piras, M.G.; Taub, D.; Schlessinger, D.; Uda, M.; Lakatta, E.G. Independent and additive effects of cytokine patterns and the metabolic syndrome on arterial aging in the SardiNIA Study. Atherosclerosis 2011, 215, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Strait, J.B.; Lakatta, E.G. Aging-associated cardiovascular changes and their relationship to heart failure. Heart Fail. Clin. 2012, 8, 143–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutin, A.R.; Scuteri, A.; Lakatta, E.G.; Tarasov, K.V.; Ferrucci, L.; Costa, P.T., Jr.; Schlessinger, D.; Uda, M.; Terracciano, A. Trait antagonism and the progression of arterial thickening: Women with antagonistic traits have similar carotid arterial thickness as men. Hypertension 2010, 56, 617–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Khazan, B.; Lakatta, E. Central Arterial Aging and Angiotensin II Signaling. Curr. Hypertens. Rev. 2010, 6, 266–281. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Monticone, R.E.; Lakatta, E.G. Arterial aging: A journey into subclinical arterial disease. Curr. Opin. Nephrol. Hypertens. 2010, 19, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhang, J.; Spinetti, G.; Jiang, L.Q.; Monticone, R.; Zhao, D.; Cheng, L.; Krawczyk, M.; Talan, M.; Pintus, G.; et al. Angiotensin II activates matrix metalloproteinase type II and mimics age-associated carotid arterial remodeling in young rats. Am. J. Pathol. 2005, 167, 1429–1442. [Google Scholar] [CrossRef] [Green Version]
- Chien, K.R.; Karsenty, G. Longevity and lineages: Toward the integrative biology of degenerative diseases in heart, muscle, and bone. Cell 2005, 120, 533–544. [Google Scholar] [CrossRef]
- Lakatta, E.G. Arterial aging is risky. J. Appl. Physiol. 2008, 105, 1321–1322. [Google Scholar] [CrossRef]
- Le Couteur, D.G.; Lakatta, E.G. A vascular theory of aging. J. Gerontol. A Biol. Sci. Med. Sci. 2010, 65, 1025–1027. [Google Scholar] [CrossRef]
- Itkin, T.; Rafii, S. Cardiovascular diseases disrupt the bone-marrow niche. Nature 2022, 601, 515–517. [Google Scholar] [CrossRef]
- Rohde, D.; Vandoorne, K.; Lee, I.; Grune, J.; Zhang, S.; McAlpine, C.S.; Schloss, M.J.; Nayar, R.; Courties, G.; Frodermann, V.; et al. Bone marrow endothelial dysfunction promotes myeloid cell expansion in cardiovascular disease. Nat. Cardiovasc. Res. 2022, 1, 28–44. [Google Scholar] [CrossRef]
- Matissek, S.J.; Elsawa, S.F. GLI3: A mediator of genetic diseases, development and cancer. Cell Commun. Signal. 2020, 18, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungricht, R.; Guibbal, L.; Lasbennes, M.C.; Orsini, V.; Beibel, M.; Waldt, A.; Cuttat, R.; Carbone, W.; Basler, A.; Roma, G.; et al. Genome-wide screening in human kidney organoids identifies developmental and disease-related aspects of nephrogenesis. Cell Stem Cell 2022, 29, 160–175.e167. [Google Scholar] [CrossRef] [PubMed]
- GeneCards: PSENEN. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=PSENEN&keywords=psenen (accessed on 1 April 2022).
- Robinson, G.I.; Ye, F.; Lu, X.; Laviolette, S.R.; Feng, Q. Maternal Delta-9-Tetrahydrocannabinol Exposure Induces Abnormalities of the Developing Heart in Mice. Cannabis Cannabinoid Res. 2022; ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Laviolette, S.R.; Hardy, D.B. Exposure to Δ9-tetrahydrocannabinol during rat pregnancy leads to impaired cardiac dysfunction in postnatal life. Pediatr. Res. 2021, 90, 532–539. [Google Scholar] [CrossRef]
- Jameson, J.L.; Fauci, A.S.; Hauser, S.L.; Longo, D.L.; Jameson, J.L.; Loscalzo, J. (Eds.) Harrison’s Principles of Internal Medicine, 20th ed.; McGraw Hill: New York, NY, USA, 2018. [Google Scholar]
- Reece, A.S.; Hulse, G.K. Gastroschisis and Autism-Dual Canaries in the Californian Coalmine. JAMA Surg. 2019, 154, 366–367. [Google Scholar] [CrossRef]
- Reece, A.S.; Hulse, G.K. Cannabis and Pregnancy Don’t Mix. Mo. Med. 2020, 117, 530–531. [Google Scholar]
- Corsi, D.J.; Donelle, J.; Sucha, E.; Hawken, S.; Hsu, H.; El-Chaâr, D.; Bisnaire, L.; Fell, D.; Wen, S.W.; Walker, M. Maternal cannabis use in pregnancy and child neurodevelopmental outcomes. Nat. Med. 2020, 26, 1536–1540. [Google Scholar] [CrossRef]
- Corsi, D.J.; Walsh, L.; Weiss, D.; Hsu, H.; El-Chaar, D.; Hawken, S.; Fell, D.B.; Walker, M. Association Between Self-reported Prenatal Cannabis Use and Maternal, Perinatal, and Neonatal Outcomes. JAMA 2019, 322, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Brents, L. Correlates and consequences of Prenatal Cannabis Exposure (PCE): Identifying and Characterizing Vulnerable Maternal Populations and Determining Outcomes in Exposed Offspring. In Handbook of Cannabis and Related Pathologies: Biology, Pharmacology, Diagnosis and Treatment; Preedy, V.R., Ed.; Academic Press: London, UK, 2017; Volume 1, pp. 160–170. [Google Scholar]
- Fried, P.; Watkinson, B.; James, D.; Gray, R. Current and former marijuana use: Preliminary findings of a longitudinal study of effects on IQ in young adults. CMAJ 2002, 166, 887–891. [Google Scholar]
- Fried, P.A.; Smith, A.M. A literature review of the consequences of prenatal marihuana exposure. An emerging theme of a deficiency in aspects of executive function. Neurotoxicol. Teratol. 2001, 23, 1–11. [Google Scholar] [CrossRef]
- Fried, P.A.; Watkinson, B.; Gray, R. Neurocognitive consequences of marihuana—A comparison with pre-drug performance. Neurotoxicol. Teratol. 2005, 27, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Mioduszewski, O.; Hatchard, T.; Byron-Alhassan, A.; Fall, C.; Fried, P.A. Prenatal marijuana exposure impacts executive functioning into young adulthood: An fMRI study. Neurotoxicol. Teratol. 2016, 58, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Longo, C.A.; Fried, P.A.; Hogan, M.J.; Cameron, I. Effects of marijuana on visuospatial working memory: An fMRI study in young adults. Psychopharmacology 2010, 210, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Fried, P.A.; Hogan, M.J.; Cameron, I. Effects of prenatal marijuana on visuospatial working memory: An fMRI study in young adults. Neurotoxicol. Teratol. 2006, 28, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Fried, P.A.; Hogan, M.J.; Cameron, I. Effects of prenatal marijuana on response inhibition: An fMRI study of young adults. Neurotoxicol. Teratol. 2004, 26, 533–542. [Google Scholar] [CrossRef]
- Smith, A.; Fried, P.; Hogan, M.; Cameron, I. The effects of prenatal and current marijuana exposure on response inhibition: A functional magnetic resonance imaging study. Brain Cogn. 2004, 54, 147–149. [Google Scholar]
- Hockings, N. Cuvier’s Objection, Morphogenesis and the Evolution Of Evolvability. 2020. Available online: https://www.researchgate.net/publication/342438770_Cuvier%27s_objection_morphogenesis_and_the_evolution_of_evolvability (accessed on 1 April 2022).
- Reece, A.S.; Hulse, G.K. Chapter 1: Close Parallels between Cannabis Use and Deteriorating US Mental Health at Four Levels Supports and Extends the Epidemiological Salience of Demonstrated Causal Mental Health Relationships: A Geospatiotemporal Study. In Epidemiology of Cannabis: Genotoxicity and Neurotoxicity, Epigenomics and Aging; Elsevier: New York, NY, USA, 2023; Volume 1, in press. [Google Scholar]
- Reece, A.S.; Hulse, G.K. Chapter 2: Linked Rise of Cannabis Use and Autism Incidence Demonstrated by Close Three Level Geospatiotemporal Relationships, USA, 1990–2011. In Epidemiology of Cannabis: Genotoxicity and Neurotoxicity, Epigenomics and Aging; Elsevier: New York, NY, USA, 2023; Volume 1, in press. [Google Scholar]
- Reece, A.S.; Hulse, G.K. Chapter 3: Geospatiotemporal and Causal Inferential Analysis of United States Congenital Anomalies as a Function of Multiple Cannabinoid- and Substance- Exposures: Phenocopying Thalidomide and Hundred Megabase-Scale Genotoxicity. In Epidemiology of Cannabis: Genotoxicity and Neurotoxicity, Epigenomics and Aging; Elsevier: New York, NY, USA, 2023; Volume 1, in press. [Google Scholar]
- Reece, A.S.; Hulse, G.K. Chapter 4: Geospatiotemporal and Causal Inferential Epidemiological Survey and Exploration of Cannabinoid- and Substance- Related Carcinogenesis in USA 2003–2017. In Epidemiology of Cannabis: Genotoxicity and Neurotoxicity, Epigenomics and Aging; Elsevier: New York, NY, USA, 2023; Volume 1, in press. [Google Scholar]
- Reece, A.S.; Hulse, G.K. Chapter 5: Multivalent Cannabinoid Epigenotoxicities and Multigenerational Aging. In Epidemiology of Cannabis: Genotoxicity and Neurotoxicity, Epigenomics and Aging; Elsevier: New York, NY, USA, 2023; Volume 1, in press. [Google Scholar]
- Reece, A.S.; Hulse, G.K. Epidemiology of Cannabis: Genotoxicity and Neurotoxicity, Epigenomics and Aging; Elsevier: New York, NY, USA, 2023. [Google Scholar]
- Jenkins, K.J.; Correa, A.; Feinstein, J.A.; Botto, L.; Britt, A.E.; Daniels, S.R.; Elixson, M.; Warnes, C.A.; Webb, C.L. Noninherited risk factors and congenital cardiovascular defects: Current knowledge: A scientific statement from the American Heart Association Council on Cardiovascular Disease in the Young: Endorsed by the American Academy of Pediatrics. Circulation 2007, 115, 2995–3014. [Google Scholar] [CrossRef] [Green Version]
- Alberry, B.; Laufer, B.I.; Chater-Diehl, E.; Singh, S.M. Epigenetic Impacts of Early Life Stress in Fetal Alcohol Spectrum Disorders Shape the Neurodevelopmental Continuum. Front. Mol. Neurosci. 2021, 14, 671891. [Google Scholar] [CrossRef]
- Chang, R.C.; Thomas, K.N.; Mehta, N.A.; Veazey, K.J.; Parnell, S.E.; Golding, M.C. Programmed suppression of oxidative phosphorylation and mitochondrial function by gestational alcohol exposure correlate with widespread increases in H3K9me2 that do not suppress transcription. Epigenetics Chromatin 2021, 14, 27. [Google Scholar] [CrossRef]
- Darbinian, N.; Selzer, M.E. Oligodendrocyte pathology in fetal alcohol spectrum disorders. Neural Regen. Res. 2022, 17, 497–502. [Google Scholar]
- Gutherz, O.R.; Deyssenroth, M.; Li, Q.; Hao, K.; Jacobson, J.L.; Chen, J.; Jacobson, S.W.; Carter, R.C. Potential roles of imprinted genes in the teratogenic effects of alcohol on the placenta, somatic growth, and the developing brain. Exp. Neurol. 2022, 347, 113919. [Google Scholar] [CrossRef] [PubMed]
- Kruithof, P.; Ban, S. A brief overview of fetal alcohol syndrome for health professionals. Br. J. Nurs. 2021, 30, 890–893. [Google Scholar] [CrossRef] [PubMed]
- Legault, L.M.; Doiron, K.; Breton-Larrivée, M.; Langford-Avelar, A.; Lemieux, A.; Caron, M.; Jerome-Majewska, L.A.; Sinnett, D.; McGraw, S. Pre-implantation alcohol exposure induces lasting sex-specific DNA methylation programming errors in the developing forebrain. Clin. Epigenetics 2021, 13, 164. [Google Scholar] [CrossRef] [PubMed]
- Lussier, A.A.; Bodnar, T.S.; Moksa, M.; Hirst, M.; Kobor, M.S.; Weinberg, J. Prenatal Adversity Alters the Epigenetic Profile of the Prefrontal Cortex: Sexually Dimorphic Effects of Prenatal Alcohol Exposure and Food-Related Stress. Genes 2021, 12, 1773. [Google Scholar] [CrossRef] [PubMed]
- Lussier, A.A.; Bodnar, T.S.; Weinberg, J. Intersection of Epigenetic and Immune Alterations: Implications for Fetal Alcohol Spectrum Disorder and Mental Health. Front. Neurosci. 2021, 15, 788630. [Google Scholar] [CrossRef]
- Smith, S.M.; Virdee, M.S.; Eckerle, J.K.; Sandness, K.E.; Georgieff, M.K.; Boys, C.J.; Zeisel, S.H.; Wozniak, J.R. Polymorphisms in SLC44A1 are associated with cognitive improvement in children diagnosed with fetal alcohol spectrum disorder: An exploratory study of oral choline supplementation. Am. J. Clin. Nutr. 2021, 114, 617–627. [Google Scholar] [CrossRef]
- Terracina, S.; Ferraguti, G.; Tarani, L.; Messina, M.P.; Lucarelli, M.; Vitali, M.; De Persis, S.; Greco, A.; Minni, A.; Polimeni, A.; et al. Transgenerational Abnormalities Induced by Paternal Preconceptual Alcohol Drinking. Findings from Humans and Animal Models. Curr. Neuropharmacol. 2021, 20, 1158–1173. [Google Scholar] [CrossRef]
- Thomas, K.N.; Zimmel, K.N.; Roach, A.N.; Basel, A.; Mehta, N.A.; Bedi, Y.S.; Golding, M.C. Maternal background alters the penetrance of growth phenotypes and sex-specific placental adaptation of offspring sired by alcohol-exposed males. FASEB J. 2021, 35, e22035. [Google Scholar] [CrossRef]
- Wallén, E.; Auvinen, P.; Kaminen-Ahola, N. The Effects of Early Prenatal Alcohol Exposure on Epigenome and Embryonic Development. Genes 2021, 12, 1095. [Google Scholar] [CrossRef]
- Subbanna, S.; Shivakumar, M.; Psychoyos, D.; Xie, S.; Basavarajappa, B.S. Anandamide-CB1 receptor signaling contributes to postnatal ethanol-induced neonatal neurodegeneration, adult synaptic, and memory deficits. J. Neurosci. 2013, 33, 6350–6366. [Google Scholar] [CrossRef] [Green Version]
- Subbanna, S.; Nagre, N.N.; Umapathy, N.S.; Pace, B.S.; Basavarajappa, B.S. Ethanol exposure induces neonatal neurodegeneration by enhancing CB1R Exon1 histone H4K8 acetylation and up-regulating CB1R function causing neurobehavioral abnormalities in adult mice. Int. J. Neuropsychopharmacol. 2014, 18, pyu028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subbanna, S.; Nagre, N.N.; Shivakumar, M.; Joshi, V.; Psychoyos, D.; Kutlar, A.; Umapathy, N.S.; Basavarajappa, B.S. CB1R-Mediated Activation of Caspase-3 Causes Epigenetic and Neurobehavioral Abnormalities in Postnatal Ethanol-Exposed Mice. Front. Mol. Neurosci. 2018, 11, 45. [Google Scholar] [CrossRef] [PubMed]
- Joshi, V.; Subbanna, S.; Shivakumar, M.; Basavarajappa, B.S. CB1R regulates CDK5 signaling and epigenetically controls Rac1 expression contributing to neurobehavioral abnormalities in mice postnatally exposed to ethanol. Neuropsychopharmacology 2019, 44, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivakumar, M.; Subbanna, S.; Joshi, V.; Basavarajappa, B.S. Postnatal Ethanol Exposure Activates HDAC-Mediated Histone Deacetylation, Impairs Synaptic Plasticity Gene Expression and Behavior in Mice. Int. J. Neuropsychopharmacol. 2020, 23, 324–338. [Google Scholar] [CrossRef]
- Subbanna, S.; Basavarajappa, B.S. Postnatal Ethanol-Induced Neurodegeneration Involves CB1R-Mediated β-Catenin Degradation in Neonatal Mice. Brain Sci. 2020, 10, 271. [Google Scholar] [CrossRef]
- Kulaga, V.; Shor, S.; Koren, G. Correlation between drugs of abuse and alcohol by hair analysis: Parents at risk for having children with fetal alcohol spectrum disorder. Alcohol 2010, 44, 615–621. [Google Scholar] [CrossRef]
- Shor, S.; Nulman, I.; Kulaga, V.; Koren, G. Heavy in utero ethanol exposure is associated with the use of other drugs of abuse in a high-risk population. Alcohol 2010, 44, 623–627. [Google Scholar] [CrossRef]
- Buchi, K.F.; Suarez, C.; Varner, M.W. The prevalence of prenatal opioid and other drug use in Utah. Am. J. Perinatol. 2013, 30, 241–244. [Google Scholar]
- Subbanna, S.; Psychoyos, D.; Xie, S.; Basavarajappa, B.S. Postnatal ethanol exposure alters levels of 2-arachidonylglycerol-metabolizing enzymes and pharmacological inhibition of monoacylglycerol lipase does not cause neurodegeneration in neonatal mice. J. Neurochem. 2015, 134, 276–287. [Google Scholar] [CrossRef] [Green Version]
- Seleverstov, O.; Tobiasz, A.; Jackson, J.S.; Sullivan, R.; Ma, D.; Sullivan, J.P.; Davison, S.; Akkhawattanangkul, Y.; Tate, D.L.; Costello, T.; et al. Maternal alcohol exposure during mid-pregnancy dilates fetal cerebral arteries via endocannabinoid receptors. Alcohol 2017, 61, 51–61. [Google Scholar] [CrossRef]
- Gal, P.; Sharpless, M.K. Fetal drug exposure-behavioral teratogenesis. Drug Intell. Clin. Pharm. 1984, 18, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Faden, V.B.; Graubard, B.I. Maternal substance use during pregnancy and developmental outcome at age three. J. Subst. Abuse 2000, 12, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Psychoyos, D.; Hungund, B.; Cooper, T.; Finnell, R.H. A cannabinoid analogue of Delta9-tetrahydrocannabinol disrupts neural development in chick. Birth Defects Res. B Dev. Reprod. Toxicol. 2008, 83, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.; Lee, J.; Mitchell, E.; Moore, L.; Baxter, E.J.; Hewinson, J.; Dawson, K.J.; Menzies, A.; Godfrey, A.L.; Green, A.R.; et al. Life histories of myeloproliferative neoplasms inferred from phylogenies. Nature 2022, 602, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Malouf, C.; Ottersbach, K. Molecular processes involved in B cell acute lymphoblastic leukaemia. Cell Mol. Life Sci. 2018, 75, 417–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Shih, J.; Hollern, D.P.; Wang, L.; Bowlby, R.; Tickoo, S.K.; Thorsson, V.; Mungall, A.J.; Newton, Y.; Hegde, A.M.; et al. Integrated Molecular Characterization of Testicular Germ Cell Tumors. Cell Rep. 2018, 23, 3392–3406. [Google Scholar] [CrossRef] [PubMed]
- Kagawa, H.; Javali, A.; Khoei, H.H.; Sommer, T.M.; Sestini, G.; Novatchkova, M.; Scholte op Reimer, Y.; Castel, G.; Bruneau, A.; Maenhoudt, N.; et al. Human blastoids model blastocyst development and implantation. Nature 2022, 601, 600–605. [Google Scholar] [CrossRef]
- Chioccarelli, T.; Falco, G.; Cappetta, D.; De Angelis, A.; Roberto, L.; Addeo, M.; Ragusa, M.; Barbagallo, D.; Berrino, L.; Purrello, M.; et al. FUS driven circCNOT6L biogenesis in mouse and human spermatozoa supports zygote development. Cell Mol. Life Sci. 2021, 79, 50. [Google Scholar] [CrossRef]
- Nahas, G.G. Keep Off the Grass; Elsevier: Middlebury, VT, USA, 1990; Volume 1. [Google Scholar]
- Nahas, G.G. Cannabis Physiopathology Epidemiology Detection; CRC Press Revivals: Boca Raton, FL, USA, 1990; Volume 1. [Google Scholar]
- Russo, C.; Ferk, F.; Mišík, M.; Ropek, N.; Nersesyan, A.; Mejri, D.; Holzmann, K.; Lavorgna, M.; Isidori, M.; Knasmüller, S. Low doses of widely consumed cannabinoids (cannabidiol and cannabidivarin) cause DNA damage and chromosomal aberrations in human-derived cells. Arch. Toxicol. 2019, 93, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Pucci, M.; Rapino, C.; Di Francesco, A.; Dainese, E.; D’Addario, C.; Maccarrone, M. Epigenetic control of skin differentiation genes by phytocannabinoids. Br. J. Pharmacol. 2013, 170, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Maor, Y.; Yu, J.; Kuzontkoski, P.M.; Dezube, B.J.; Zhang, X.; Groopman, J.E. Cannabidiol inhibits growth and induces programmed cell death in kaposi sarcoma-associated herpesvirus-infected endothelium. Genes Cancer 2012, 3, 512–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karmaus, P.W.; Wagner, J.G.; Harkema, J.R.; Kaminski, N.E.; Kaplan, B.L. Cannabidiol (CBD) enhances lipopolysaccharide (LPS)-induced pulmonary inflammation in C57BL/6 mice. J. Immunotoxicol. 2013, 10, 321–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hind, W.H.; England, T.J.; O’Sullivan, S.E. Cannabidiol protects an in vitro model of the blood-brain barrier from oxygen-glucose deprivation via PPARgamma and 5-HT1A receptors. Br. J. Pharmacol. 2016, 173, 815–825. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.E.; Sun, Y.; Bennett, A.J.; Randall, M.D.; Kendall, D.A. Time-dependent vascular actions of cannabidiol in the rat aorta. Eur. J. Pharmacol. 2009, 612, 61–68. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.E.; Kendall, D.A. Cannabinoid activation of peroxisome proliferator-activated receptors: Potential for modulation of inflammatory disease. Immunobiology 2010, 215, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Hegde, V.L.; Singh, U.P.; Nagarkatti, P.S.; Nagarkatti, M. Critical Role of Mast Cells and Peroxisome Proliferator-Activated Receptor gamma in the Induction of Myeloid-Derived Suppressor Cells by Marijuana Cannabidiol In Vivo. J. Immunol. 2015, 194, 5211–5222. [Google Scholar] [CrossRef] [Green Version]
- Ramer, R.; Heinemann, K.; Merkord, J.; Rohde, H.; Salamon, A.; Linnebacher, M.; Hinz, B. COX-2 and PPAR-gamma confer cannabidiol-induced apoptosis of human lung cancer cells. Mol. Cancer Ther. 2013, 12, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Scuderi, C.; Steardo, L.; Esposito, G. Cannabidiol promotes amyloid precursor protein ubiquitination and reduction of beta amyloid expression in SHSY5YAPP+ cells through PPARgamma involvement. Phytother. Res. 2014, 28, 1007–1013. [Google Scholar] [CrossRef]
- De Filippis, D.; Esposito, G.; Cirillo, C.; Cipriano, M.; De Winter, B.Y.; Scuderi, C.; Sarnelli, G.; Cuomo, R.; Steardo, L.; De Man, J.G.; et al. Cannabidiol reduces intestinal inflammation through the control of neuroimmune axis. PLoS ONE 2011, 6, e28159. [Google Scholar] [CrossRef] [Green Version]
- Esposito, G.; Scuderi, C.; Valenza, M.; Togna, G.I.; Latina, V.; De Filippis, D.; Cipriano, M.; Carratù, M.R.; Iuvone, T.; Steardo, L. Cannabidiol reduces Abeta-induced neuroinflammation and promotes hippocampal neurogenesis through PPARgamma involvement. PLoS ONE 2011, 6, e28668. [Google Scholar] [CrossRef]
- Chan, J.Z.; Duncan, R.E. Regulatory Effects of Cannabidiol on Mitochondrial Functions: A Review. Cells 2021, 10, 1251. [Google Scholar] [CrossRef] [PubMed]
- Olivas-Aguirre, M.; Torres-López, L.; Pottosin, I.; Dobrovinskaya, O. Phenolic Compounds Cannabidiol, Curcumin and Quercetin Cause Mitochondrial Dysfunction and Suppress Acute Lymphoblastic Leukemia Cells. Int. J. Mol. Sci. 2020, 22, 204. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.; Drysdale, A.J.; Lafourcade, C.; Pertwee, R.G.; Platt, B. Cannabidiol targets mitochondria to regulate intracellular Ca2+ levels. J. Neurosci. 2009, 29, 2053–2063. [Google Scholar] [CrossRef] [Green Version]
- Winklmayr, M.; Gaisberger, M.; Kittl, M.; Fuchs, J.; Ritter, M.; Jakab, M. Dose-Dependent Cannabidiol-Induced Elevation of Intracellular Calcium and Apoptosis in Human Articular Chondrocytes. J. Orthop. Res. 2019, 37, 2540–2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayakawa, K.; Mishima, K.; Hazekawa, M.; Sano, K.; Irie, K.; Orito, K.; Egawa, T.; Kitamura, Y.; Uchida, N.; Nishimura, R.; et al. Cannabidiol potentiates pharmacological effects of Delta(9)-tetrahydrocannabinol via CB(1) receptor-dependent mechanism. Brain Res. 2008, 1188, 157–164. [Google Scholar] [CrossRef]
- Silvestri, C.; Di Marzo, V. The endocannabinoid system in energy homeostasis and the etiopathology of metabolic disorders. Cell Metab. 2013, 17, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Mato, S.; Victoria Sánchez-Gómez, M.; Matute, C. Cannabidiol induces intracellular calcium elevation and cytotoxicity in oligodendrocytes. Glia 2010, 58, 1739–1747. [Google Scholar] [CrossRef]
- Fišar, Z.; Singh, N.; Hroudová, J. Cannabinoid-induced changes in respiration of brain mitochondria. Toxicol. Lett. 2014, 231, 62–71. [Google Scholar] [CrossRef]
- Alhamoruni, A.; Lee, A.C.; Wright, K.L.; Larvin, M.; O’Sullivan, S.E. Pharmacological effects of cannabinoids on the Caco-2 cell culture model of intestinal permeability. J. Pharmacol. Exp. Ther. 2010, 335, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, J.A.; Biagioni, A.F.; Almada, R.C.; de Souza Crippa, J.A.; Cecilio Hallak, J.E.; Zuardi, A.W.; Coimbra, N.C. Dissociation between the panicolytic effect of cannabidiol microinjected into the substantia nigra, pars reticulata, and fear-induced antinociception elicited by bicuculline administration in deep layers of the superior colliculus: The role of CB1-cannabinoid receptor in the ventral mesencephalon. Eur. J. Pharmacol. 2015, 758, 153–163. [Google Scholar]
- Laprairie, R.B.; Bagher, A.M.; Kelly, M.E.M.; Denovan-Wright, E. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol. 2015, 172, 4790–4805. [Google Scholar] [CrossRef] [Green Version]
- Stanley, C.P.; Hind, W.H.; Tufarelli, C.; O’Sullivan, S.E. Cannabidiol causes endothelium-dependent vasorelaxation of human mesenteric arteries via CB1 activation. Cardiovasc. Res. 2015, 107, 568–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartim, A.G.; Guimaraes, F.S.; Joca, S.R. Antidepressant-like effect of cannabidiol injection into the ventral medial prefrontal cortex-Possible involvement of 5-HT1A and CB1 receptors. Behav. Brain Res. 2016, 303, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.S.; Kim, Y.J.; Kim, M.O.; Kang, M.; Oh, S.W.; Nho, Y.H.; Park, S.H.; Lee, J. Cannabidiol upregulates melanogenesis through CB1 dependent pathway by activating p38 MAPK and p42/44 MAPK. Chem. Biol. Interact. 2017, 273, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.R.; Gomes, F.V.; Fonseca, M.D.; Mechoulam, R.; Breuer, A.; Cunha, T.M.; Guimaraes, F.S. Antinociceptive effects of HUF-101, a fluorinated cannabidiol derivative. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79, 369–377. [Google Scholar] [CrossRef]
- Stern, C.A.; da Silva, T.R.; Raymundi, A.M.; de Souza, C.P.; Hiroaki-Sato, V.A.; Kato, L.; Guimarães, F.S.; Andreatini, R.; Takahashi, R.N.; Bertoglio, L.J. Cannabidiol disrupts the consolidation of specific and generalized fear memories via dorsal hippocampus CB1 and CB2 receptors. Neuropharmacology 2017, 125, 220–230. [Google Scholar] [CrossRef]
- Fogaça, M.V.; Campos, A.C.; Coelho, L.D.; Duman, R.S.; Guimarães, F.S. The anxiolytic effects of cannabidiol in chronically stressed mice are mediated by the endocannabinoid system: Role of neurogenesis and dendritic remodeling. Neuropharmacology 2018, 135, 22–33. [Google Scholar] [CrossRef]
- Mahoney, J.M.; Harris, R.A. Effect of 9 -tetrahydrocannabinol on mitochondrial processes. Biochem. Pharmacol. 1972, 21, 1217–1226. [Google Scholar] [CrossRef]
- Bartova, A.; Birmingham, M.K. Effect of delta9-tetrahydrocannabinol on mitochondrial NADH-oxidase activity. J. Biol. Chem. 1976, 251, 5002–5006. [Google Scholar] [CrossRef]
- Hebert-Chatelain, E.; Reguero, L.; Puente, N.; Lutz, B.; Chaouloff, F.; Rossignol, R.; Piazza, P.V.; Benard, G.; Grandes, P.; Marsicano, G. Cannabinoid control of brain bioenergetics: Exploring the subcellular localization of the CB1 receptor. Mol. Metab. 2014, 3, 495–504. [Google Scholar] [CrossRef]
- Jimenez-Blasco, D.; Busquets-Garcia, A.; Hebert-Chatelain, E.; Serrat, R.; Vicente-Gutierrez, C.; Ioannidou, C.; Gómez-Sotres, P.; Lopez-Fabuel, I.; Resch-Beusher, M.; Resel, E.; et al. Glucose metabolism links astroglial mitochondria to cannabinoid effects. Nature 2020, 583, 603–608. [Google Scholar] [CrossRef] [PubMed]
- PPARG Peroxisome Proliferator Activated Receptor Gamma [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=ShowDetailView&TermToSearch=5468 (accessed on 1 April 2022).
- Reece, A.S.; Hulse, G.K. Epidemiology of Δ8THC–Related Carcinogenesis in USA: A Panel Regression and Causal Inferential Study. Int. J. Environ. Res. Public Health 2022, 19, 7726–7752. [Google Scholar] [CrossRef] [PubMed]
- Reece, A.S.; Hulse, G.K. Congenital Anomaly Epidemiological Correlates of Δ8THC Across USA 2003–2016: Panel Regression and Causal Inferential Study. Environ. Epigenetics 2022, in press.
- Gant, J. Scientists are baffled by spatter of babies born without hands or arms in France, as investigation fails to discover a cause. The Daily Mail, 13 July 2019. [Google Scholar]
- Willsher, K. Baby arm defects prompt nationwide investigation in France. The Guardian, 31 October 2018. [Google Scholar]
- Agence France-Presse in Paris. France to investigate cause of upper limb defects in babies. The Guardian, 21 October 2018.
- Babies Born with Deformed Hands Spark Investigation in Germany. Available online: https://edition.cnn.com/2019/09/16/health/hand-deformities-babies-gelsenkirchen-germany-intl-scli-grm/index.html (accessed on 1 April 2022).
- Wang, Y.X.; Blau, H.M. Reversing aging for heart repair. Science 2021, 373, 1439–1440. [Google Scholar] [CrossRef] [PubMed]
- Bejaoui, Y.; Razzaq, A.; Yousri, N.A.; Oshima, J.; Megarbane, A.; Qannan, A.; Potabattula, R.; Alam, T.; Martin, G.M.; Horn, H.F.; et al. DNA methylation signatures in Blood DNA of Hutchinson-Gilford Progeria syndrome. Aging Cell 2022, 21, e13555. [Google Scholar] [CrossRef] [PubMed]
- Do, W.L.; Nguyen, S.; Yao, J.; Guo, X.; Whitsel, E.A.; Demerath, E.; Rotter, J.I.; Rich, S.S.; Lange, L.; Ding, J.; et al. Associations between DNA methylation and BMI vary by metabolic health status: A potential link to disparate cardiovascular outcomes. Clin. Epigenetics 2021, 13, 230. [Google Scholar] [CrossRef]
- Fu, K.A.; Paul, K.C.; Lu, A.T.; Horvath, S.; Keener, A.M.; Bordelon, Y.; Bronstein, J.M.; Ritz, B. DNA methylation-based surrogates of plasma proteins are associated with Parkinson’s disease risk. J. Neurol. Sci. 2021, 431, 120046. [Google Scholar] [CrossRef]
- Gao, T.; Wilkins, J.T.; Zheng, Y.; Joyce, B.T.; Jacobs, D.R., Jr.; Schreiner, P.J.; Horvath, S.; Greenland, P.; Lloyd-Jones, D.; Hou, L. Plasma lipid profiles in early adulthood are associated with epigenetic aging in the Coronary Artery Risk Development in Young Adults (CARDIA) Study. Clin. Epigenetics 2022, 14, 16. [Google Scholar] [CrossRef]
- Roberts, J.D.; Vittinghoff, E.; Lu, A.T.; Alonso, A.; Wang, B.; Sitlani, C.M.; Mohammadi-Shemirani, P.; Fornage, M.; Kornej, J.; Brody, J.A.; et al. Epigenetic Age and the Risk of Incident Atrial Fibrillation. Circulation 2021, 144, 1899–1911. [Google Scholar] [CrossRef]
- Schlosser, P.; Tin, A.; Matias-Garcia, P.R.; Thio, C.H.; Joehanes, R.; Liu, H.; Weihs, A.; Yu, Z.; Hoppmann, A.; Grundner-Culemann, F.; et al. Meta-analyses identify DNA methylation associated with kidney function and damage. Nat. Commun. 2021, 12, 7174. [Google Scholar] [CrossRef] [PubMed]
- Tin, A.; Schlosser, P.; Matias-Garcia, P.R.; Thio, C.H.; Joehanes, R.; Liu, H.; Yu, Z.; Weihs, A.; Hoppmann, A.; Grundner-Culemann, F.; et al. Epigenome-wide association study of serum urate reveals insights into urate co-regulation and the SLC2A9 locus. Nat. Commun. 2021, 12, 7173. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Bara, A.; Vargas, C.A.; Frick, A.L.; Loh, E.; Landry, J.; Uzamere, T.O.; Callens, J.E.; Martin, Q.; Rajarajan, P.; et al. Prenatal Δ(9)-Tetrahydrocannabinol Exposure in Males Leads to Motivational Disturbances Related to Striatal Epigenetic Dysregulation. Biol. Psychiatry 2021, 92, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Porath, A.J.; Fried, P.A. Effects of prenatal cigarette and marijuana exposure on drug use among offspring. Neurotoxicol. Teratol. 2005, 27, 267–277. [Google Scholar] [CrossRef] [PubMed]





| No. | Streams of Evidence | Focus of the Discussion |
|---|---|---|
| Section 3.1.1 | Clinical syndromes | Clinical phenomenology |
| Section 3.1.2 | Mitochondrial inhibition | Cellular systems and mechanisms |
| Section 3.1.3 | DNA Methylation | Cellular systems and mechanisms |
| Section 3.1.4 | Mental illnesses | Organ systems |
| Section 3.1.5 | Cardiovascular age | Organ systems |
| Section 3.1.6 | Endocrine suppression | Organ systems |
| Section 3.1.7 | Liver inflammation | Organ systems |
| Section 3.1.8 | Cancer | Heath disorders and Population impacts |
| Section 3.1.9 | Inheritable cancer | Heath disorders and Population impacts |
| Section 3.1.10 | Congenital Anomalies | Heath disorders and Population impacts |
| Section 3.1.11 | Telomerase inhibition | Cellular systems and mechanisms |
| Section 3.1.12 | Elevated Mortality rate | Epidemiological Studies |
| Pathogenetic Field of Interest | ||
| Section 3.2.1 | Epigenomic Overview | Cellular systems and mechanisms |
| Section 3.2.2 | Stem-Cell Factors | Cellular systems and mechanisms |
| Section 3.2.3 | Chromosomal Mechanics | Cellular systems and mechanisms |
| Section 3.2.4 | Centromeres and Kinetochores | Cellular systems and mechanisms |
| Section 3.2.5 | Prefrontal cortex and Brain | Organ systems |
| Section 3.2.6 | Cardiovascular System | Organ systems |
| Section 3.2.7 | Teratogenesis | Analysis DNA Methylation data and epidemiological impacts |
| Section 3.2.8 | Carcinogenesis | Analysis DNA Methylation data and epidemiological impacts |
| Nearest Gene Name | Chromosome Number | Nearest Gene Number | Dependency Status | Functional Annotation | Page | Distance from Nearest Gene | Relative Position | p-Value | Bonferroni Adjusted p-Value |
|---|---|---|---|---|---|---|---|---|---|
| DNA Methyltransferases | |||||||||
| DNMT1 | 19 | ENSG00000130816 | Withdrawal | Maintenance DNA methyltransferase | 156 | 0 | Intron | 1.89 × 10−6 | 0.010563 |
| DNMT1 | 19 | ENSG00000130816 | Withdrawal | Maintenance DNA methyltransferase | 179 | 0 | Intron | 4.81 × 10−6 | 0.016176 |
| DNMT3B | 20 | ENSG00000088305 | Dependence | de novo DNA methyltransferase | 109 | 0 | Intron | 1.22 × 10−5 | 0.023205 |
| DNMT3B | 20 | ENSG00000088305 | Withdrawal | de novo DNA methyltransferase | 125 | 1067 | Upstream | 2.08 × 10−8 | 0.001062 |
| DNMT3A | 2 | ENSG00000119772 | Withdrawal | de novo DNA methyltransferase | 194 | 0 | Intron | 7.57 × 10−6 | 0.020149 |
| DNA Demethylases | |||||||||
| TET1 | 10 | ENSG00000138336 | Dependence | Ten-Eleven translocase | 107 | 0 | Intron | 1.18 × 10−5 | 0.022782 |
| TET1P1 | 13 | ENSG00000232204 | Dependence | Pseudogene for TET | 63 | 36,150 | Downstream | 4.14 × 10−6 | 0.013905 |
| TET1P1 | 13 | ENSG00000232204 | Dependence | Pseudogene for TET | 85 | 47,940 | Upstream | 7.47 × 10−6 | 0.018443 |
| TET1P1 | 13 | ENSG00000232204 | Dependence | Pseudogene for TET | 98 | 9930 | Downstream | 9.97 × 10−6 | 0.021086 |
| TET1P1 | 13 | ENSG00000232204 | Dependence | Pseudogene for TET | 98 | 55,192 | Upstream | 6.32 × 10−6 | 0.018533 |
| Others | |||||||||
| UHRF1 | 19 | ENSG00000276043 | Withdrawal | Integrator of epigenetic information | 128 | 0 | Intron | 5.74 × 10−8 | 0.001782 |
| UHRF1BP1L | 12 | ENSG00000111647 | Withdrawal | Regulator of UHRF1 | 155 | 0 | Intron | 1.79 × 10−6 | 0.010239 |
| UHRF1BP1L | 12 | ENSG00000111647 | Withdrawal | Regulator of UHRF1 | 233 | 0 | Intron | 1.67 × 10−5 | 0.028881 |
| DPPA2 | 3 | ENSG00000163530 | Dependence | Developmental Pluripotency Associated 2 | 40 | 15,599 | Downstream | 1.66 × 10−6 | 0.009001 |
| DPPA2 | 3 | ENSG00000163530 | Dependence | Developmental Pluripotency Associated 2 | 133 | 6894 | Downstream | 1.90 × 10−7 | 0.003298 |
| DPPA2P1 | Y | ENSG00000223915 | Withdrawal | Pseudogene for DPPA2A | 135 | 26,055 | Upstream | 2.78 × 10−7 | 0.004034 |
| Telomerase | |||||||||
| TERT | 5 | ENSG00000223915 | Dependence | Telomerase | 44 | 4227 | Upstream | 2.82 × 10−6 | 0.012582 |
| Polycomb Repressors | |||||||||
| PCGF6 in PRC1 | 10 | ENSG00000156374 | Dependence | Polycomb Repressive Complex 1 | 65 | 0 | Intron | 4.37 × 10−6 | 0.014300 |
| PCGF6 in PRC1 | 10 | ENSG00000156374 | Withdrawal | Polycomb Repressive Complex 1 | 137 | 0 | Intron | 4.03 × 10−7 | 0.004978 |
| EZH2 in PRC2 | 7 | ENSG00000180628 | Dependence | Polycomb Repressive Complex 2 | 94 | 0 | Intron | 9.22 × 10−6 | 0.020342 |
| Chromatin Remodellers | |||||||||
| SMARCA2 | 9 | ENSG00000080503 | Dependence | SWI/SNF Matrix, Actin Chromatin Regulator 2 | 6 | 0 | Intron | 5.27 × 10−9 | 0.000438 |
| SMARCA2 | 9 | ENSG00000080503 | Dependence | SWI/SNF Matrix, Actin Chromatin Regulator 2 | 62 | 3071 | Downstream | 4.00 × 10−6 | 0.013641 |
| SMARCA2 | 9 | ENSG00000080503 | Dependence | SWI/SNF Matrix, Actin Chromatin Regulator 2 | 114 | 0 | Intron | 1.34 × 10−5 | 0.024371 |
| SMARCA4 | 19 | ENSG00000127616 | Withdrawal | SWI/SNF Matrix, Actin Chromatin Regulator 4 | 145 | 9567 | Upstream | 8.86 × 10−7 | 0.007300 |
| SMARCA4 | 19 | ENSG00000127616 | Withdrawal | SWI/SNF Matrix, Actin Chromatin Regulator 4 | 199 | 9258 | Upstream | 8.54 × 10−6 | 0.021311 |
| Nearest Gene Name | Chromosome Number | Nearest Gene Number | Dependency Status | Functional Annotation | Page | Distance from Nearest Gene | Relative Position | p-Value | Bonferroni Adjusted p-Value |
|---|---|---|---|---|---|---|---|---|---|
| POU5F1P2 | 8 | ENSG00000253382 | Dependence | Oct3/4 Pseudogene | 5 | 2871 | Downstream | 1.49 × 10−9 | 0.000216 |
| SOX2-OT | 3 | ENSG00000242808 | Dependence | Sox2 Overlapping Transcript | 6 | 0 | Intron | 5.25 × 10−9 | 0.000438 |
| SOX2-OT | 3 | ENSG00000242808 | Dependence | Sox2 Overlapping Transcript | 48 | 0 | Intron | 2.38 × 10−6 | 0.017245 |
| SOX2-OT | 3 | ENSG00000242808 | Dependence | Sox2 Overlapping Transcript | 88 | 0 | Intron | 8.12 × 10−6 | 0.019185 |
| SOX2-OT | 3 | ENSG00000242808 | Withdrawal | Sox2 Overlapping Transcript | 116 | 0 | Intron | 1.40 × 10−5 | 0.024849 |
| SOX2-OT | 3 | ENSG00000242808 | Withdrawal | Sox2 Overlapping Transcript | 146 | 0 | Intron | 9.74 × 10−7 | 0.007679 |
| SOX2-OT | 3 | ENSG00000242808 | Withdrawal | Sox2 Overlapping Transcript | 211 | 0 | Intron | 1.11 × 10−5 | 0.023974 |
| Klf4 | 9 | ENSG00000136826 | Dependence | Kruppel-like factor 4 | 117 | 12,186 | Upstream | 1.41 × 10−5 | 0.024968 |
| MycBP2 | 13 | ENSG00000005810 | Dependence | Myc Binding Protein 2 | 49 | 0 | Intron | 2.50 × 10−6 | 0.010960 |
| MycBP2 | 13 | ENSG00000005810 | Withdrawal | Myc Binding Protein 2 | 153 | 0 | Intron | 1.58 × 10−6 | 0.009647 |
| Myc | 8 | ENSG00000136826 | Withdrawal | Myc proto-oncogene | 227 | 23,489 | Downstream | 1.49 × 10−5 | 0.027466 |
| Nearest Gene Name | Nearest Gene Number | Chromosome Number | Relative Location | Distance to Nearest Gene (Bases) | Number of Annotations | p-Value | Bonferroni-Adjusted p-Value |
|---|---|---|---|---|---|---|---|
| Centrosomal Proteins | |||||||
| CENPIP1 | ENSG00000224778 | 13 | Upstream | 1100 | 1 | 2.38 × 10−9 | 0.000279 |
| CENPF | ENSG00000117724 | 1 | Downstream | 72,569 | 3 | 2.98 × 10−8 | 0.001109 |
| CNEPVL3 | ENSG00000224109 | X | Downstream | 2146 | 1 | 2.80 × 10−6 | 0.001153 |
| CENPK | ENSG00000123219 | 5 | Intron | 0 | 1 | 8.01 × 10−6 | 0.019098 |
| CNEPP | ENSG00000188312 | 9 | Intron | 0 | 2 | 8.26 × 10−6 | 0.019330 |
| CNEPJ | ENSG00000151849 | 13 | Exon | 0 | 1 | 4.66 × 10−7 | 0.005279 |
| CNEPUP1 | ENSG00000255075 | 11 | Upstream | 8401 | 1 | 2.81 × 10−6 | 0.012567 |
| INCENP | ENSG00000149503 | 11 | Intron | 0 | 1 | 3.07 × 10−6 | 0.013077 |
| CNEPO | ENSG00000138092 | 2 | Exon | 0 | 1 | 6.25 × 10−6 | 0.018393 |
| CNEPI | ENSG00000102384 | X | Intron | 0 | 2 | 7.54 × 10−6 | 0.020123 |
| CNEPL | ENSG00000120334 | 1 | Intron | 0 | 1 | 8.22 × 10−6 | 0.020943 |
| CNEPX | ENSG00000169689 | 17 | Exon | 0 | 1 | 9.35 × 10−6 | 0.022176 |
| CNEPC | ENSG00000145241 | 4 | Intron | 0 | 1 | 9.60 × 10−6 | 0.002248 |
| CENPV | ENSG00000166582 | 17 | Upstream | 13,237 | 2 | 1.63 × 10−5 | 0.002861 |
| CENPN | ENSG00000166451 | 16 | 86 | 7.73 × 10−20 | |||
| Others | |||||||
| KNL1 | ENSG00000137812 | 15 | 3UTR | 0 | 1 | 7.71 × 10−7 | 0.006173 |
| ZWINT | ENSG00000122952 | 10 | Downstream | 58,081 | 1 | 6.00 × 10−6 | 0.016644 |
| NUF2 | ENSG00000143228 | 1 | Intron | 0 | 1 | 1.12 × 10−6 | 0.007421 |
| SPC24 | ENSG00000161888 | 19 | 3UTR | 0 | 1 | 1.61 × 10−6 | 0.009713 |
| Sumoylation | |||||||
| SUMO1 | ENSG00000112701 | 2 | Intron | 0 | 1 | 1.25 × 10−5 | 0.023445 |
| ZNF451 | ENSG00000226803 | 6 | Intron | 0 | 1 | 2.22 × 10−6 | 0.011398 |
| SENP6 | ENSG00000112701 | 6 | Intron | 0 | 1 | 3.12 × 10−6 | 0.013217 |
| SENP7 | ENSG00000138468 | 3 | Intron | 0 | 1 | 4.73 × 10−6 | 0.014903 |
| SENP7 | ENSG00000138468 | 3 | Intron | 0 | 1 | 1.16 × 10−5 | 0.024458 |
| No. | Europe | USA | ||||||
|---|---|---|---|---|---|---|---|---|
| Congenital Anomaly | Term | Model | p-Value | Congenital Anomaly | Term | Model | p-Value | |
| 1 | Abdominal Wall Defects | pm.Resin.Daily | Categorical | 3.01 × 10−120 | ||||
| 2 | All Anomalies | Daily_Use | Categorical | <2.2 × 10−320 | ||||
| 3 | Amniotic band | pm.Resin.Daily | Categorical | 1.09 × 10−47 | ||||
| 4 | Anencephalus and similar | Resin_THC | Categorical | 1.53 × 10−212 | ||||
| 5 | Annular Pancreas | Daily_Use | Categorical | 1.52 × 10−13 | ||||
| 6 | Anophthalmos | Daily_Use | Categorical | 1.06 × 10−6 | ||||
| 7 | Ano-rectal atresia and stenosis | pm.Resin.Daily | Categorical | 4.03 × 10−39 | Large intestinal and Rectal atresia/stenosis | Cannabidiol_Estimates | Continuous | 0.0040 |
| 8 | Anotia | Herb_THC | Categorical | 4.63 × 10−13 | Anotia/microtia | LM_Cannabis | Continuous | 7.57 × 10−4 |
| 9 | Aortic atresia/interrupted aortic arch | LM.Cann_Resin_THC | Categorical | 5.71 × 10−25 | Interrupted aortic arch | LM_Cannabis | Continuous | 3.40 × 10−6 |
| 10 | Aortic Valve stenosis/atresia | Herb_THC | Categorical | 7.14 × 10−13 | Aortic valve stenosis | LM_Cannabis | Continuous | 0.0019 |
| 11 | Arhinencephaly/holoprosencephaly | LM_Herb.Daily | Continuous | 0.0052 | ||||
| 12 | Arterial Truncus | pm.Herb.Daily | Categorical | 9.92 × 10−7 | ||||
| 13 | Atrial septal defect (ASD) | Herb_THC | Categorical | <2.2 × 10−320 | Atrial septal defect (ASD) | LM_Cannabis | Continuous | 0.0378 |
| 14 | Atrioventricular septal defect (AVSD) | pm.Resin.Daily | Categorical | 1.65 × 10−101 | Atrioventricular septal defect (AVSD) | LM_Cannabis | Categorical | 0.0470 |
| 15 | Bilateral renal agenesis including Potter syndrome | Herb_THC | Categorical | 1.08 × 10−47 | Renal agenesis/hypoplasia | LM_Cannabis | Continuous | 7.34 × 10−4 |
| 16 | Bile duct atresia | Daily_Use | Categorical | 1.00 × 10−40 | Biliary atresia | Cannabidiol_Estimates | Continuous | 2.43 × 10−4 |
| 17 | Bladder Extrophy/Epispadias | pm.Resin.Daily | Categorical | 1.56 × 10−18 | Bladder extrophy | LM_Cannabis | Continuous | 0.0170 |
| 18 | Choanal Atresia | Herb_THC | Categorical | 7.34 × 10−94 | Choanal atresia | Δ9THC_Estimates | Continuous | 0.0033 |
| 19 | Chromosomal | Daily_Use | Categorical | <2.2 × 10−320 | Chromosomal | LM_Cannabis | Mixed Effects | 9.38 × 10−30 |
| 20 | Cleft lip with or without palate | Herb_THC | Categorical | 1.80 × 10−101 | Cleft lip with and without cleft palate | Cannabidiol_Estimates | Categorical | 0.0159 |
| 21 | Cleft palate | Herb_THC | Categorical | 1.79 × 10−34 | Cleft palate alone | LM_Cannabis | Continuous | 0.0014 |
| 22 | Cloacal exstrophy | LM_Cannabis | Categorical | 2.13 × 10−86 | ||||
| 23 | Club foot-talipes equinovarus | Daily_Use | Categorical | 4.23 × 10−292 | Clubfoot | LM_Cannabis | Continuous | 3.16 × 10−5 |
| 24 | Coarctation Aorta | Daily_Use | Categorical | 5.78 × 10−33 | Coarctation of the aorta | LM_Cannabis | Categorical | 9.74 × 10−45 |
| 25 | Congenital cataract | Daily_Use | Categorical | 4.88 × 10−66 | Congenital cataract | LM_Cannabis | Continuous | 0.0479 |
| 26 | Congenital glaucoma | Daily_Use | Categorical | 1.52 × 10−43 | ||||
| 27 | Congenital Heart | pm.Herb.Daily | Categorical | <2.2 × 10−320 | ||||
| 28 | Conjoined twins | Daily_Use | Categorical | 8.62 × 10−14 | ||||
| 29 | Craniosynostosis | Daily_Use | Categorical | 5.72 × 10−155 | ||||
| 30 | Cystic adenomatous malformation of lung | Daily_Use | Categorical | 4.05 × 10−80 | ||||
| 31 | Diaphragmatic Hernia | Daily_Use | Categorical | 8.77 × 10−57 | Diaphragmatic hernia | LM_Cannabis | Categorical | 2.11 × 10−8 |
| 32 | Digestive system | pm.Herb.Daily | Categorical | 1.61 × 10−264 | ||||
| 33 | Double outlet right ventricle | pm.Herb.Daily | Categorical | 1.28 × 10−46 | Double outlet right ventricle | LM_Cannabis | Categorical | 7.31 × 10−4 |
| 34 | Down Syndrome | Daily_Use | Categorical | <2.2 × 10−320 | Trisomy 21 (Down syndrome) | LM_Cannabis | Categorical | 4.02 × 10−26 |
| 35 | Duodenal stenosis/atresia | Herb_THC | Categorical | 1.50 × 10−10 | ||||
| 36 | Ear, face and neck | Daily_Use | Categorical | 3.38 × 10−44 | ||||
| 37 | Ebstein’s Anomaly | pm.Resin.Daily | Categorical | 3.23 × 10−17 | ||||
| 38 | Edward syndrome/Trisomy 18 | Daily_Use | Categorical | <2.2 × 10−320 | Edward syndrome/Trisomy 18 | LM_Cannabis | Categorical | 1.06 × 10−61 |
| 39 | Encephalocele | pm.Resin.Daily | Categorical | 4.76 × 10−21 | Encephalocele | LM_Cannabis | Continuous | 0.0013 |
| 40 | Epispadias | LM_Cannabis | Continuous | 0.0111 | ||||
| 41 | Eye | Daily_Use | Categorical | 2.27 × 10−175 | ||||
| 42 | Fetal alcohol syndrome | pm.Resin.Daily | Categorical | 5.88 × 10−57 | ||||
| 43 | Gastroschisis | Herb_THC | Categorical | 6.55 × 10−39 | ||||
| 44 | Genetic syndromes + microdeletions | pm.Herb.Daily | Categorical | 1.38 × 10−228 | Deletion 22q11.2 | LM_Cannabis | Continuous | 0.0024 |
| 45 | Genital | pm.Herb.Daily | Categorical | 2.55 × 10−243 | ||||
| 46 | Hip dislocation and/or dysplasia | Daily_Use | Categorical | <2.2 × 10−320 | Congenital hip dislocation | LM_Cannabis | Categorical | 7.27 × 10−70 |
| 47 | Hirschsprung’s disease | Daily_Use | Categorical | 2.54 × 10−88 | Hirschsprung disease (congenital megacolon) | LM_Cannabis | Categorical | 6.69 × 10−6 |
| 48 | Holoprosencephaly/Arhinencephaly | LM_Cannabis | Categorical | 1.22 × 10−72 | Holoprosencephaly | LM_Cannabis | Categorical | 2.90 × 10−12 |
| 49 | Hydrocephalus | pm.Herb.Daily | Categorical | 1.76 × 10−110 | ||||
| 50 | Hydronephrosis | Herb_THC | Categorical | <2.2 × 10−320 | ||||
| 51 | Hypoplastic Left Heart | Daily_Use | Categorical | 3.37 × 10−61 | Hypoplastic left heart syndrome | LM_Cannabis | Continuous | 0.0047 |
| 52 | Hypoplastic right heart | Resin_THC | Categorical | 2.85 × 10−59 | ||||
| 53 | Hypospadias | pm.Herb.Daily | Categorical | 2.92 × 10−177 | Hypospadias | LM_Cannabis | Continuous | 1.16 × 10−5 |
| 54 | Klinefelter syndrome | Daily_Use | Categorical | 1.75 × 10−41 | ||||
| 55 | Large intestinal and Rectal atresia/stenosis | Cannabidiol_Estimates | Continuous | 0.0040 | ||||
| 56 | Lateral anomalies | LM.Cann_Herb_THC | Categorical | 2.36 × 10−48 | ||||
| 57 | Limb anomalies | pm.Herb.Daily | Categorical | <2.2 × 10−320 | ||||
| 58 | Limb reductions | Daily_Use | Categorical | 8.20 × 10−65 | Limb deficiencies (reduction defects) | LM_Cannabis | Continuous | 0.0134 |
| 59 | Lower limb Reduction deformity | LM_Cannabis | Continuous | 0.0420 | ||||
| 60 | Maternal infections resulting in malformations | Daily_Use | Categorical | 4.15 × 10−87 | ||||
| 61 | Microphthalmos/Anophthalmos | Daily_Use | Categorical | 1.25 × 10−55 | Microphthalmos/Anophthalmos | Δ9THC_Estimates | Continuous | 0.0045 |
| 62 | Mitral valve anomalies | pm.Herb.Daily | Categorical | 8.99 × 10−58 | ||||
| 63 | Multicystic renal dysplasia | pm.Resin.Daily | Categorical | 6.70 × 10−251 | ||||
| 64 | Nervous system | pm.Herb.Daily | Categorical | <2.2 × 10−320 | ||||
| 65 | Neural Tube Defects | Resin_THC | Categorical | 9.97 × 10−269 | ||||
| 66 | Obstructive genitourinary defect | Cannabidiol_Estimates | Categorical | 2.22 × 10−15 | ||||
| 67 | Oesophageal stenosis/atresia | Daily_Use | Categorical | 3.49 × 10−44 | Oesophageal atresia/tracheoesophageal fistula | LM_Cannabis | Continuous | 4.83 × 10−6 |
| 68 | Omphalocele | pm.Resin.Daily | Categorical | 4.94 × 10−131 | Omphalocele | LM_Cannabis | Continuous | 0.0025 |
| 69 | Oro-facial clefts | Herb_THC | Categorical | 3.99 × 10−133 | ||||
| 70 | Patau syndrome/trisomy 13 | Daily_Use | Categorical | 1.08 × 10−144 | Patau syndrome/trisomy 13 | LM_Cannabis | Continuous | 2.08 × 10−7 |
| 71 | PDA as only CHD in term infants (>=37 weeks) | pm.Herb.Daily | Categorical | 2.14 × 10−20 | ||||
| 72 | Polydactyly | pm.Resin.Daily | Categorical | 1.46 × 10−292 | ||||
| 73 | Posterior urethral valve and/or prune belly | pm.Resin.Daily | Categorical | 1.28 × 10−42 | Congenital posterior urethral valves | LM_Cannabis | Continuous | 1.18 × 10−4 |
| 74 | Pulmonary valve atresia | Daily_Use | Categorical | 1.42 × 10−27 | Pulmonary valve atresia | Cannabidiol_Estimates | Categorical | 1.02 × 10−5 |
| 75 | Pulmonary valve stenosis | Daily_Use | Categorical | 2.09 × 10−95 | ||||
| 76 | Respiratory | pm.Herb.Daily | Categorical | 2.57 × 10−203 | ||||
| 77 | Severe CHD | Herb_THC | Categorical | 1.81 × 10-317 | ||||
| 78 | Severe microcephaly | pm.Herb.Daily | Categorical | 3.17 × 10−148 | ||||
| 79 | Single ventricle | Daily_Use | Categorical | 1.03 × 10−25 | Single ventricle | LM_Cannabis | Categorical | 0.0060 |
| 80 | Situs inversus | Daily_Use | Categorical | 1.42 × 10−44 | ||||
| 81 | Skeletal dysplasias | Daily_Use | Categorical | 5.12 × 10−74 | ||||
| 82 | Small Intestine stenosis/atresia | pm.Herb.Daily | Categorical | 8.23 × 10−31 | Small intestinal atresia/stenosis | Cannabidiol_Estimates | Continuous | 3.39 × 10−6 |
| 83 | Spina Bifida | Resin_THC | Categorical | 3.93 × 10−84 | Spina bifida without anencephalus | Δ9THC_Estimates | Continuous | 0.0008 |
| 84 | Syndactyly | pm.Resin.Daily | Categorical | 3.47 × 10−16 | ||||
| 85 | Teratogenic syndromes with malformations | Daily_Use | Categorical | 1.42 × 10−139 | ||||
| 86 | Tetralogy of Fallot | Daily_Use | Categorical | 3.12 × 10−47 | Tetralogy of Fallot | LM_Cannabis | Continuous | 0.0168 |
| 87 | Total Anomalous Pulmonary Venous Return | Herb_THC | Categorical | 4.07 × 10−09 | Total anomalous pulmonary venous connection | LM_Cannabis | Continuous | 0.0299 |
| 88 | Transposition of great vessels | Resin_THC | Categorical | 9.96 × 10−33 | Transposition of great arteries | Cannabidiol_Estimates | Continuous | 0.0479 |
| 89 | Turner syndrome | Daily_Use | Categorical | 1.10 × 10−146 | Turner syndrome | LM_Cannabis | Categorical | 7.69 × 10−49 |
| 90 | Tricuspid valve stenosis/atresia | Daily_Use | Categorical | 6.86 × 10−24 | ||||
| 91 | Urinary | pm.Resin.Daily | Categorical | <2.2 × 10−320 | ||||
| 92 | Valproate syndrome | Daily_Use | Categorical | 1.57 × 10−7 | ||||
| 93 | Vascular disruption anomalies | Herb_THC | Categorical | 3.46 × 10−101 | ||||
| 94 | VATER/VACTERL | pm.Herb.Daily | Categorical | 2.43 × 10−36 | ||||
| 95 | Ventricular septal defect (VSD) | pm.Resin.Daily | Categorical | <2.2 × 10−320 | Ventricular septal defect | LM_Cannabis | Continuous | 0.0021 |
| Gene Acronym | Gene Name | Gene Number | Functional Annotation | Status | Page Number | Number of Genes Annotated | p-Value |
|---|---|---|---|---|---|---|---|
| Meis1 | Meis Homeobox 1 | ENSG00000143995 | Withdrawal | 194 | 37 | 7.55 × 10−6 | |
| Meis1 | Meis Homeobox 1 | ENSG00000143995 | Cancer growth | Withdrawal | 325 | 149 | 7.17 × 10−6 |
| Meis1 | Meis Homeobox 1 | ENSG00000143995 | Sensory organ development | Withdrawal | 327 | 18 | 1.30 × 10−5 |
| Meis1 | Meis Homeobox 1 | ENSG00000143995 | Eye formation | Withdrawal | 328 | 15 | 2.81 × 10−5 |
| Meis1 | Meis Homeobox 1 | ENSG00000143995 | Cancer | Withdrawal | 329 | 151 | 4.32 × 10−5 |
| Meis1 | Meis Homeobox 1 | ENSG00000143995 | Lens formation | Withdrawal | 333 | 4 | 9.17 × 10−5 |
| Meis1 | Meis Homeobox 1 | ENSG00000143995 | Cancer | Withdrawal | 334 | 88 | 1.22 × 10−4 |
| Meis1 | Meis Homeobox 1 | ENSG00000143995 | Eye formation | Withdrawal | 334 | 11 | 1.23 × 10−4 |
| Meis2 | Meis Homeobox 2 | ENSG00000134138 | Withdrawal | 134 | 97 | 2.36 × 10−7 | |
| Meis2 | Meis Homeobox 2 | ENSG00000134138 | Withdrawal | 181 | 1 | 0.016676 | |
| Meis2 | Meis Homeobox 2 | ENSG00000134138 | Withdrawal | 209 | 1 | 0.023289 | |
| Meis2 | Meis Homeobox 2 | ENSG00000134138 | Upper Aerodigestive SCC | Withdrawal | 325 | 40 | 1.28 × 10−6 |
| Meis2 | Meis Homeobox 2 | ENSG00000134138 | Upper Aerodigestive SCC | Withdrawal | 325 | 53 | 3.59 × 10−6 |
| Meis2 | Meis Homeobox 2 | ENSG00000134138 | Cranial nerve abnormality | Withdrawal | 325 | 7 | 6.34 × 10−6 |
| Meis2 | Meis Homeobox 2 | ENSG00000134138 | Cancer | Withdrawal | 325 | 149 | 7.17 × 10−6 |
| FGFs | Fibroblast Growth Factor | Withdrawal | 175 | ||||
| FGFR1OP | FGF Receptor 1 Oncogene Partner | ENSG00000213066 | Withdrawal | 13 | 1 | 0.002226 | |
| FGF5 | Fibroblast Growth Factor 5 | ENSG00000138675 | Withdrawal | 21 | 1 | 0.004362 | |
| FGF14 | Fibroblast Growth Factor 14 | ENSG00000102466 | Withdrawal | 25 | 1 | 0.005329 | |
| FGFR2 | Fibroblast Growth Factor Receptor 2 | ENSG00000066468 | Withdrawal | 28 | 1 | 0.005981 | |
| FGF14 | Fibroblast Growth Factor 14 | ENSG00000102466 | Dependence | 30 | 1 | 8.68 × 10−7 | |
| FGF12 | Fibroblast Growth Factor 12 | ENSG00000114279 | Dependence | 41 | 1 | 0.009199 | |
| FGF12 | Fibroblast Growth Factor 12 | ENSG00000114279 | Dependence | 54 | 1 | 0.001187 | |
| FGF3 | Fibroblast Growth Factor 3 | ENSG00000186895 | Dependence | 81 | 1 | 0.017663 | |
| FGFRL1 | FGF Receptor Like 3 | ENSG00000127418 | Dependence | 86 | 1 | 0.018855 | |
| FGF14 | Fibroblast Growth Factor 14 | ENSG00000102466 | Dependence | 106 | 1 | 0.002259 | |
| FGF4 | Fibroblast Growth Factor 4 | ENSG00000122642 | Dependence | 17 | 7 | 2.34 × 10−7 | |
| FGF4 | Fibroblast Growth Factor 4 | ENSG00000122642 | KEGG: Rap1 signaling | 236 | 41 | 0.000353 | |
| FGF4 | Fibroblast Growth Factor 4 | ENSG00000122642 | KEGG: actin cytoskeleton | 237 | 37 | 0.004586 | |
| FGF4 | Fibroblast Growth Factor 4 | ENSG00000122642 | KEGG: melanoma | 237 | 15 | 0.021590 | |
| FGF4 | Fibroblast Growth Factor 4 | ENSG00000122642 | KEGG: MAP kinase pathway | 237 | 39 | 0.029222 | |
| FGF4 | Fibroblast Growth Factor 4 | ENSG00000122642 | KEGG: Cancer pathways | 238 | 54 | 0.067770 | |
| FGF4 | Fibroblast Growth Factor 4 | ENSG00000122642 | KEGG: Ras signaling | 328 | 38 | 0.008745 | |
| RXRA | Retinoid X Receptor Alpha | ENSG00000186350 | Withdrawal | 125 | 1 | 1.48 × 10−8 | |
| RXRG | Retinoid X Receptor Gamma | ENSG00000143171 | Withdrawal | 136 | 1 | 3.40 × 10−7 | |
| RXRA | Retinoid X Receptor Alpha | ENSG00000186350 | Withdrawal | 144 | 1 | 8.40 × 10−7 | |
| RARA | Retinoic Acid Receptor Alpha | ENSG00000131759 | Dependence | 44 | 1 | 1.95 × 10−6 | |
| RARB | Retinoic Acid Receptor Beta | ENSG00000077092 | Dependence | 73 | 1 | 5.54 × 10−6 | |
| RARB | Retinoic Acid Receptor Beta | ENSG00000077092 | Withdrawal | 124 | 1 | 7.94 × 10−9 | |
| RARB | Retinoic Acid Receptor Beta | ENSG00000077092 | Withdrawal | 168 | 1 | 3.25 × 10−6 | |
| RARB | Retinoic Acid Receptor Beta | ENSG00000077092 | Withdrawal | 190 | 1 | 6.89 × 10−6 | |
| RARB | Retinoic Acid Receptor Beta | ENSG00000077092 | Withdrawal | 215 | 1 | 1.20 × 10−5 | |
| RARA | Retinoic Acid Receptor Alpha | ENSG00000131759 | KEGG: Cancer pathways | 238 | 54 | 0.067777 | |
| WNT’s | Wnt’s | Withdrawal | 203 | ||||
| WNT7B | Wnt family member 7B | ENSG00000188064 | Dependence | 74 | 1 | 5.78 × 10−6 | |
| WNT7A | Wnt family member 7A | ENSG00000154764 | Dependence | 119 | 1 | 1.47 × 10−0 | |
| WNT7A | Wnt family member 7A | ENSG00000154764 | Dependence | 123 | 1 | 4.13 × 10−9 | |
| WNT3A | Wnt family member 3A | ENSG00000154342 | Head and neck cancer | Withdrawal | 239 | 356 | 7.73 × 10−20 |
| WNT8B | Wnt family member 8B | ENSG00000075290 | Head and neck cancer | Withdrawal | 239 | 342 | 7.74 × 10−20 |
| TBX4 | T-Box transcription factor 4 | ENSG00000121075 | Dependence | 52 | 1 | 2.72 × 10−6 | |
| TBX4 | T-Box transcription factor 4 | ENSG00000121075 | Withdrawal | 235 | 1 | 1.71 × 10−5 | |
| TBX5-AS1 | T-Box transcription factor 5 Antisense 1 | ENSG00000255399 | Withdrawal | 202 | 1 | 9.18 × 10−6 | |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Dependence | 37 | 124 | 1.37 × 10−6 | |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Upper aerodigestive SCC | Withdrawal | 325 | 40 | 1.28 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Upper aerodigestive SCC | Withdrawal | 325 | 115 | 1.65 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Skin lesion | Withdrawal | 325 | 53 | 3.59 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Skin cancer | Withdrawal | 325 | 113 | 4.79 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Cancer | Withdrawal | 325 | 149 | 7.17 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Large bowel adenocarcinoma | Withdrawal | 326 | 120 | 7.45 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Cutaneous melanoma | Withdrawal | 326 | 110 | 7.71 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | High grade astocytoma | Withdrawal | 326 | 82 | 8.42 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Abdominal adenocarcinoma | Withdrawal | 326 | 135 | 8.46 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Solid organ cancer | Withdrawal | 327 | 150 | 9.16 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Head and neck cancer | Withdrawal | 327 | 137 | 9.54 × 10−6 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Sensory organ development | Withdrawal | 327 | 18 | 1.30 × 10−5 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Carcinoma | Withdrawal | 327 | 148 | 1.38 × 10−5 |
| CHD7 | Chromodomain Helicase DNA Binding Protein 7 | ENSG00000171316 | Upper aerodigestive SCC | Withdrawal | 327 | 44 | 1.60 × 10−43 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Skin lesion | Withdrawal | 325 | 105 | 1.65 × 10−6 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Skin cancer | Withdrawal | 325 | 113 | 4.79 × 10−6 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Cranial nerve abnormality | Withdrawal | 325 | 7 | 6.34 × 10−6 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Cancer | Withdrawal | 325 | 149 | 7.17 × 10−6 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Large bowel adenocarcinoma | Withdrawal | 326 | 120 | 7.45 × 10−6 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Cutaneous melanoma | Withdrawal | 326 | 110 | 7.71 × 10−6 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | High grade astocytoma | Withdrawal | 326 | 82 | 8.42 × 10−6 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Abdominal adenocarcinoma | Withdrawal | 326 | 135 | 8.46 × 10−6 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Solid organ cancer | Withdrawal | 327 | 150 | 9.16 × 10−6 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Head and neck cancer | Withdrawal | 327 | 137 | 9.54 × 10−6 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Carcinoma | Withdrawal | 327 | 148 | 1.38 × 10−5 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Carcinoma | Withdrawal | 329 | 151 | 4.32 × 10−5 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Squamous cell tumor | Withdrawal | 332 | 65 | 7.59 × 10−5 |
| MEGF8 | Multiple EGF-like domains 8 | ENSG00000105429 | Preaxial polydactyly | Withdrawal | 333 | 3 | 9.19 × 10−5 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Upper aerodigestive SCC | Withdrawal | 325 | 22 | 1.28 × 10−6 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Cancer | Withdrawal | 325 | 149 | 7.17 × 10−6 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Solid organ cancer | Withdrawal | 327 | 150 | 9.16 × 10−6 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Head and neck cancer | Withdrawal | 327 | 137 | 9.54 × 10−6 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Carcinoma | Withdrawal | 327 | 148 | 1.38 × 10−5 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Carcinoma | Withdrawal | 329 | 151 | 4.32 × 10−5 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Squamous cell tumor | Withdrawal | 331 | 65 | 7.59 × 10−5 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Preaxial polydactyly | Withdrawal | 333 | 3 | 9.19 × 10−5 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Squamous cell tumor | Withdrawal | 334 | 64 | 1.45 × 10−4 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Head and neck cancer | Withdrawal | 335 | 127 | 1.75 × 10−4 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Cancer | Withdrawal | 337 | 79 | 2.83 × 10−4 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Head abnormalities | Withdrawal | 338 | 21 | 3.27 × 10−4 |
| TMEM107 | Transmembrane protein 107 | ENSG00000179029 | Haemopoietic stimulation | Withdrawal | 338 | 23 | 3.51 × 10−4 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Upper aerodigestive SCC | Withdrawal | 325 | 166 | 1.28 × 10−6 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Upper aerodigestive SCC | Withdrawal | 325 | 115 | 1.65 × 10−6 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Cranial nerve abnormality | Withdrawal | 325 | 7 | 6.34 × 10−6 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Cancer | Withdrawal | 325 | 149 | 7.17 × 10−6 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Large bowel adenocarcinoma | Withdrawal | 326 | 120 | 7.45 × 10−6 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Abdominal adenocarcinoma | Withdrawal | 326 | 135 | 8.46 × 10−6 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Solid organ cancer | Withdrawal | 327 | 150 | 9.16 × 10-=6 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Head and neck cancer | Withdrawal | 327 | 137 | 9.54 × 10−6 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Sensory organ development | Withdrawal | 327 | 18 | 1.30 × 10−5 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Carcinoma | Withdrawal | 327 | 148 | 1.38 × 10−5 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Upper aerodigestive SCC | Withdrawal | 327 | 44 | 1.60 × 10−5 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Carcinoma | Withdrawal | 328 | 119 | 2.47 × 10−5 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Eye formation | Withdrawal | 328 | 15 | 2.81 × 10−5 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | Upper GIT carcinoma | Withdrawal | 328 | 75 | 3.42 × 10−5 |
| BMP4 | Bone morphogenetic protein 4 | ENSG00000125378 | GIT adenocarcinoma | Withdrawal | 328 | 121 | 3.56 × 10−5 |
| GREM1 | GREM1, DAN family BMP antagonist | ENSG00000126873 | Withdrawal | 171 | 1 | 3.61 × 10−6 | |
| GREM2 | GREM2, DAN family BMP antagonist | ENSG00000180875 | Withdrawal | 85 | 1 | 9.90 × 10−6 | |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Skin lesion | Withdrawal | 325 | 183 | 1.28 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Head and neck squamous carcinoma | Withdrawal | 325 | 53 | 1.65 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Skin cancer | Withdrawal | 325 | 113 | 3.59 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Lung adenocarcinoma | Withdrawal | 325 | 42 | 4.79 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Cancer | Withdrawal | 325 | 149 | 7.17 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Large bowel adenocarcinoma | Withdrawal | 326 | 120 | 7.45 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Cutaneous melanoma | Withdrawal | 326 | 110 | 7.71 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | High grade astocytoma | Withdrawal | 326 | 82 | 8.42 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Abdominal adenocarcinoma | Withdrawal | 326 | 135 | 8.46 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Solid organ cancer | Withdrawal | 327 | 150 | 9.16 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Head and neck cancer | Withdrawal | 327 | 137 | 9.54 × 10−6 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Sensory organ development | Withdrawal | 327 | 18 | 1.30 × 10−5 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Carcinoma | Withdrawal | 327 | 148 | 1.38 × 10−5 |
| GLI3 | GLI zinc finger family 3 | ENSG00000106571 | Upper aerodigestive SCC | Withdrawal | 327 | 44 | 1.60 × 10−43 |
| System | Mean p-Value | Median p-Value |
|---|---|---|
| Gastrointestinal | 0.0011 | 7.45 × 10−6 |
| Chromosomes | 0.0018 | 1.31 × 10−4 |
| Neurological | 0.0035 | 6.15 × 10−4 |
| Cardiovascular | 0.0011 | 0.0011 |
| Face | 0.0021 | 0.0014 |
| Body Wall | 0.0018 | 0.0016 |
| General | 0.0026 | 0.0017 |
| Uronephrology | 0.0021 | 0.0022 |
| Limb | 0.0036 | 0.0037 |
| Target | Cannabis Dependence | Cannabis Withdrawal | ||||||
|---|---|---|---|---|---|---|---|---|
| Number of Annotations | Cumulative Genes | Minimum p-Value | Median p-Value | Number of Annotations | Cumulative Genes | Minimum p-Value | Median p-Value | |
| Gastrointestinal | 8 | 2561 | 4.60 × 10−16 | 1.13 × 10−15 | - | - | - | - |
| Large Intestine | 5 | 1240 | 7.65 × 10−15 | 6.40 × 10−14 | 3 | 363 | 7.45 × 10−6 | 6.80 × 10−5 |
| Esophagus | 4 | 393 | 3.15 × 10−13 | 9.40 × 10−4 | 3 | 69 | 0.0020 | 0.0028 |
| Neurological | 8 | 710 | 5.33 × 10−8 | 4.45 × 10−4 | 1 | 2 | 7.20 × 10−4 | 7.20 × 10−4 |
| Heart | 5 | 53 | 8.83 × 10−8 | 1.57 × 10−4 | - | - | - | - |
| Liver | 2 | 404 | 1.28 × 10−7 | 1.79 × 10−7 | - | - | - | - |
| Brain | 6 | 750 | 1.39 × 10−7 | 1.86 × 10−5 | 1 | 3 | 1.16 × 10−4 | 1.16 × 10−4 |
| Pancreas | 8 | 769 | 9.10 × 10−7 | 1.25 × 10−5 | 4 | 112 | 0.0052 | 0.0061 |
| Embryo | 9 | 285 | 8.20 × 10−6 | 3.61 × 10−4 | - | - | - | - |
| Atrioventricular valves | 3 | 13 | 9.04 × 10−6 | 4.00 × 10−5 | - | - | - | - |
| Neurons | 14 | 336 | 9.27 × 10−6 | 1.88 × 10−4 | 3 | 11 | 0.0020 | 0.0031 |
| DNA | 12 | 373 | 1.50 × 10−5 | 0.0011 | 5 | 33 | 3.58 × 10−4 | 0.0070 |
| Chromosomes | 3 | 16 | 1.60 × 10−5 | 7.90 × 10−5 | 1 | 1 | 0.0070 | 0.0070 |
| Cardiovascular | 4 | 85 | 2.10 × 10−5 | 0.0019 | - | - | - | - |
| Synapse | 15 | 308 | 3.12 × 10−5 | 0.0018 | 7 | 36 | 1.43 × 10−4 | 0.0013 |
| Microtubules | 1 | 58 | 3.30 × 10−5 | 3.30 × 10−5 | 1 | 24 | 0.0045 | 0.0045 |
| Embryo | 6 | 93 | 3.60 × 10−5 | 0.0018 | 2 | 8 | 0.0023 | 0.0046 |
| Ventricle | 4 | 23 | 5.10 × 10−5 | 6.09 × 10−4 | - | - | - | - |
| Body | 5 | 132 | 7.80 × 10−5 | 0.0016 | 2 | 51 | 1.93 × 10−4 | 3.74 × 10−4 |
| Eye | 6 | 65 | 7.90 × 10−5 | 0.0010 | 13 | 73 | 2.80 × 10−5 | 6.89 × 10−4 |
| Cerebrum | 2 | 153 | 1.20 × 10−4 | 7.35 × 10−4 | 4 | 22 | 7.41 × 10−4 | 0.0020 |
| Head | 1 | 47 | 1.20 × 10−4 | 1.20 × 10−4 | - | - | - | - |
| Bone | 7 | 50 | 1.40 × 10−4 | 0.0018 | 14 | 48 | 1.93 × 10−4 | 0.0070 |
| Sensory | 1 | 29 | 1.64 × 10−4 | 1.64 × 10−4 | - | - | - | - |
| Body Axis | 1 | 1 | 1.93 × 10−4 | 1.93 × 10−4 | - | - | - | - |
| Urinary system | 1 | 17 | 2.20 × 10−4 | 2.20 × 10−4 | 1 | 8 | 0.0044 | 0.0044 |
| Kidney | 5 | 84 | 4.29 × 10−4 | 0.0022 | 1 | 4 | 0.0042 | 0.0042 |
| Breast | 1 | 3 | 5.73 × 10−4 | 5.73 × 10−4 | 2 | 9 | 0.0021 | 0.0023 |
| Granulocytes | 1 | 3 | 5.73 × 10−4 | 5.73 × 10−4 | - | - | - | - |
| Ear | 6 | 36 | 7.20 × 10−4 | 0.0021 | 5 | 21 | 1.65 × 10−4 | 8.04 × 10−4 |
| Atria | 4 | 15 | 8.55 × 10−4 | 0.0017 | - | - | - | - |
| Body trunk | 1 | 50 | 0.0015 | 0.0015 | - | - | - | - |
| Myogenesis | 2 | 4 | 0.0018 | 0.0018 | - | - | - | - |
| Vertebra | 1 | 3 | 0.0049 | 0.0049 | - | - | - | - |
| Limb | - | - | - | - | 6 | 18 | 9.20 × 10−5 | 0.0037 |
| Nose | - | - | - | - | 1 | 3 | 0.0011 | 0.0011 |
| Ovarian reserve | - | - | - | - | 1 | 2 | 0.0031 | 0.0031 |
| Mitochondria | - | - | - | - | 1 | 1 | 0.0070 | 0.0070 |
| Palate | - | - | - | - | 1 | 1 | 0.0070 | 0.0070 |
| No. | Europe | USA | |||||
|---|---|---|---|---|---|---|---|
| Model | Cancer | Minimum p-Value | Model | Correlate | Cancer | Minimum p-Value | |
| 1 | Categorical | Acute Lymphoid Leukemia | 8.70 × 10−24 | Categorical | Δ9THC | Acute Lymphoid Leukemia | 7.65 × 10−25 |
| 2 | Continuous | Acute Myeloid Leukemia | 2.11 × 10−4 | Categorical | Δ9THC | Acute Myeloid Leukemia | 3.11 × 10−110 |
| 3 | Categorical | Cannabidiol | All_Cancer | <2.2 × 10−320 | |||
| 4 | Categorical | Anus | 6.71 × 10−35 | ||||
| 5 | Categorical | Bladder | <2.2 × 10−320 | Categorical | Cannabidiol | Bladder | <2.2 × 10−320 |
| 6 | Continuous | Brain.Medulloblastoma | 5.64 × 10−42 | Categorical | Cannabidiol | Brain | 5.67 × 10−33 |
| 7 | Categorical | Breast | 4.03 × 10−17 | Categorical | Δ9THC | Breast | 8.06 × 10−146 |
| 8 | Continuous | Chronic Lymphoid Leukemia | 1.20 × 10−34 | Categorical | Cannabidiol | Chronic Lymphoid Leukemia | 2.98 × 10−12 |
| 9 | Continuous | Chronic Myeloid Leukemia | 1.32 × 10−32 | Categorical | Δ9THC | Chronic Myeloid Leukemia | 1.52 × 10−12 |
| 10 | Categorical | Colorectum | 6.14 × 10−242 | Categorical | Cannabidiol | Colorectum | <2.2 × 10−320 |
| 11 | Categorical | Corpus uteri | 2.28 × 10−4 | ||||
| 12 | Categorical | Esophagus | 1.12 × 10−110 | Categorical | Cannabidiol | Esophagus | 2.31 × 10−43 |
| 13 | Categorical | Gallbladder | 2.24 × 10−4 | ||||
| 14 | Continuous | Hepatocellular Cancer | 2.29 × 10−42 | ||||
| 15 | Categorical | Hodgkin lymphoma | 1.80 × 10−8 | Categorical | Cannabidiol | Hodgkins | 1.22 × 10−30 |
| 16 | Categorical | Kaposi sarcoma | 1.16 × 10−7 | Categorical | Cannabidiol | Kaposi | 4.75 × 10−29 |
| 17 | Categorical | Kidney | 7.46 × 10−5 | Continuous | Cannabinol | Kidney | 0.0067 |
| 18 | Categorical | Larynx | <2.2 × 10−320 | ||||
| 19 | Categorical | Liver | <2.2 × 10−320 | Categorical | Δ9THC | Liver | <2.2 × 10−320 |
| 20 | Categorical | Lung | 1.45 × 10−8 | Categorical | Cannabidiol | Lung | 6.87 × 10−194 |
| 21 | Categorical | Melanoma of skin | <2.2 × 10−320 | Categorical | Cannabidiol | Melanoma | <2.2 × 10−320 |
| 22 | Categorical | Mesothelioma | 3.37 × 10−111 | ||||
| 23 | Categorical | Multiple myeloma | 6.92 × 10−8 | Categorical | Δ9THC | Multiple myeloma | 1.73 × 10−30 |
| 24 | Categorical | Non-Hodgkin lymphoma | 1.60 × 10−44 | Categorical | Cannabidiol | Non-Hodgkin lymphoma | 3.15 × 10−145 |
| 25 | Continuous | Oropharynx | 7.02 × 10−21 | Continuous | Δ9THC | Oropharynx | 3.21 × 10−6 |
| 26 | Categorical | Ovary.Germ Cell Tumor | 1.07 × 10−38 | Categorical | Cannabidiol | Ovary | 2.49 × 10−312 |
| 27 | Categorical | Pancreas | 4.09 × 10−9 | Categorical | Δ9THC | Pancreas | 4.57 × 10−166 |
| 28 | Categorical | Penis | 1.64 × 10−19 | ||||
| 29 | Categorical | Prostate | <2.2 × 10−320 | Categorical | Cannabidiol | Prostate | <2.2 × 10−320 |
| 30 | Categorical | Cannabidiol | Stomach | 2.30 × 10−192 | |||
| 31 | Categorical | Testis | 3.83 × 10−81 | Continuous | Cannabinol | Testis | 1.47 × 10−5 |
| 32 | Continuous | Testis.Non-Seminoma Germ | 1.25 × 10−75 | ||||
| 33 | Categorical | Testis.Seminoma | 5.14 × 10−58 | ||||
| 34 | Categorical | Thyroid | <2.2 × 10−320 | Categorical | Δ9THC | Thyroid | <2.2 × 10−320 |
| 35 | Continuous | Vulva | 8.88 × 10−44 | ||||
| Cancer | Minimum p-Value Dependence | Minimum p-Value Withdrawal | p-Value Ratio Dependence/Withdrawal | Total Gene Number Dependence | Total Gene Number Withdrawal | Gene Number Ratio Dependence/Withdrawal |
|---|---|---|---|---|---|---|
| Thyroid | 1.21 × 10−17 | 0.0014 | 1.17 × 1014 | 637 | 115 | 5.54 |
| Melanoma | 3.70 × 10−15 | 7.71 × 10−6 | 2.08 × 109 | 579 | 225 | 2.57 |
| Urinary | 2.54 × 10−14 | 2.16 × 10−4 | 8.50 × 109 | 1191 | 679 | 1.75 |
| Esophagus | 3.15 × 10−13 | 6.80 × 10−5 | 2.16 × 108 | 465 | 117 | 3.97 |
| Stomach | 3.15 × 10−13 | 6.80 × 10−5 | 2.16 × 108 | 443 | 102 | 4.34 |
| Colorectal | 7.27 × 10−13 | 6.17 × 10−4 | 8.49 × 108 | 1734 | 452 | 3.84 |
| Testis | 1.14 × 10−8 | 6.75 × 10−4 | 5.92 × 104 | 304 | 60 | 5.07 |
| Liver | 1.17 × 10−8 | NA | NA | 890 | NA | NA |
| Prostate | 2.88 × 10−8 | 5.33 × 10−4 | 1.85 × 104 | 399 | 158 | 2.53 |
| Breast | 3.25 × 10−8 | 0.0013 | 3.91 × 104 | 674 | 177 | 3.81 |
| Brain | 5.33 × 10−8 | 8.42 × 10−6 | 157.97 | 2779 | 947 | 2.93 |
| Oropharynx | 1.25 × 10−7 | 1.60 × 10−5 | 128.00 | 195 | 44 | 4.43 |
| Pancreas | 9.10 × 10−7 | 0.0052 | 5.73 × 103 | 769 | 112 | 6.87 |
| ALL | 4.08 × 10−5 | 6.01 × 10−4 | 14.73 | 23 | 118 | 0.19 |
| NHL | 4.08 × 10−5 | 6.11 × 10−4 | 14.98 | 322 | 43 | 7.49 |
| Ovary | 1.16 × 10−4 | 0.0070 | 60.43 | 529 | 1 | 529.00 |
| CML | 2.13 × 10−4 | 0.0021 | 9.95 | 11 | 11 | 1.00 |
| AML | 8.96 × 10−4 | 6.26 × 10−4 | 0.70 | 11 | 36 | 0.31 |
| Kidney | 0.00101 | NA | NA | 89 | NA | NA |
| Myeloma | NA | 0.0016 | NA | NA | 10 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reece, A.S.; Hulse, G.K. Epigenomic and Other Evidence for Cannabis-Induced Aging Contextualized in a Synthetic Epidemiologic Overview of Cannabinoid-Related Teratogenesis and Cannabinoid-Related Carcinogenesis. Int. J. Environ. Res. Public Health 2022, 19, 16721. https://doi.org/10.3390/ijerph192416721
Reece AS, Hulse GK. Epigenomic and Other Evidence for Cannabis-Induced Aging Contextualized in a Synthetic Epidemiologic Overview of Cannabinoid-Related Teratogenesis and Cannabinoid-Related Carcinogenesis. International Journal of Environmental Research and Public Health. 2022; 19(24):16721. https://doi.org/10.3390/ijerph192416721
Chicago/Turabian StyleReece, Albert Stuart, and Gary Kenneth Hulse. 2022. "Epigenomic and Other Evidence for Cannabis-Induced Aging Contextualized in a Synthetic Epidemiologic Overview of Cannabinoid-Related Teratogenesis and Cannabinoid-Related Carcinogenesis" International Journal of Environmental Research and Public Health 19, no. 24: 16721. https://doi.org/10.3390/ijerph192416721