
Essential Factors for a Healthy Microbiome: A Scoping Review


1
Health Sciences, College of Health Professions, Sacred Heart University, Fairfield, CT 06825, USA
2
Department of Nutrition, School of Mathematics, Science and Engineering, University of the Incarnate Word, San Antonio, TX 78209, USA
3
Public Health and Physical Therapy and Human Movement Sciences, College of Health Professions, Sacred Heart University, Fairfield, CT 06825, USA
*
Author to whom correspondence should be addressed.
Int. J. Environ. Res. Public Health 2022, 19(14), 8361; https://doi.org/10.3390/ijerph19148361
Submission received: 16 May 2022
/
Revised: 5 July 2022
/
Accepted: 6 July 2022
/
Published: 8 July 2022
(This article belongs to the Special Issue Healthy Nutrition and Physical Activity)
Abstract
:Recent discoveries of the purpose and potential of microbial interactions with humans have broad implications for our understanding of metabolism, immunity, the host–microbe genetic interactions. Bioavailability and bioaccessibility of phytonutrients in foods not only enrich microbial diversity in the lower human gastrointestinal tract (GIT) but also direct the functioning of the metagenome of the microbiota. Thus, healthy choices must include foods that contain nutrients that satisfy both the needs of humans and their microbes. Physical activity interventions at a moderate level of intensity have shown positive effects on metabolism and the microbiome, while intense training (>70% VO2max) reduces diversity in the short term. The microbiome of elite endurance athletes is a robust producer of short-chain fatty acids. A lifestyle lacking activity is associated with the development of chronic disease, and experimental conditions simulating weightlessness in humans demonstrate loss of muscle mass occurring in conjunction with a decline in gut short-chain fatty acid (SCFA) production and the microbes that produce them. This review summarizes evidence addressing the relationship between the intestinal microbiome, diet, and physical activity. Data from the studies reviewed suggest that food choices and physical fitness in developed countries promote a resource “curse” dilemma for the microbiome and our health.
1. Introduction
Microbial species in the gut, including fungi, viruses, and bacteria, are key influencers in human development, immune function, and health. The gut microbiome functions as both a nutritional competitor and supplier of nutraceuticals and facilitates human metabolism. In this way, the microbiome behaves as an economy that trades in nutrients and the end products of fermentation; thus, supply and demand forces may apply. Economists have observed that developing countries often suffer from the paradox of plenty, such that resource riches lead to marginalization and poverty in some groups [1]. The “resource curse” framework can be applied to the human microbiome, where richness in some nutritional components leads to bacterial overgrowth of certain species, thus crowding out the growth and sustainability of others [2]. The western pattern diet (WPD) due to its nutrient resources which are low in fiber and carbohydrate polymers of 10 or more monomeric units, disrupts microbial populations resulting in damage to the intestinal epithelial barrier (IEB)–the gatekeeper of nutrient permeability into the endothelium [3]. Once the intercellular junction has been breached, inflammation and tissue injury occur. Pathogens take advantage of this disruption, while commensal bacteria become displaced by dysregulation. Resource imbalance creates a lack of “leadership” among microbes and results in a lawless state of dysbiosis where some species go into decline.
The formation of the protective mucous barrier on the lining of the intestinal lumen creates a physical space for the host–microbe exchange of nutrients and metabolites [4]. In addition, the production of short-chain fatty acids (SCFAs) through the fermentation of polysaccharides supports the energy needs of endothelial cells [5]. Commensal bacteria also play a key role in the post-natal development of the gut-associated immune system by regulating immune homeostasis, thus providing another defense mechanism against invading pathogens [6]. The adoption of dietary patterns must be viewed within a broader context of a system of interactions between the host and its microbiome.
Physical activity (PA) has been shown to prevent and treat a number of chronic diseases, including heart disease, type 2 diabetes, high blood pressure, some cancers, depression, and dementia [7]. In an experiment using dry immersion to simulate extreme inactivity while controlling for diet, researchers observed that participants not only developed muscle loss, but experienced a reduction in microbial production of propionate and an increase in Firmicutes [8]. Studies on habitual athletes have shown that they also have high levels of Firmicutes, but individual species vary by sport and activity level [9]. The gut microbiome of sedentary adults in a short-term exercise intervention given supplemental protein developed an increase in microbial and viral diversity [10]. However, one would expect that when athletes are compared to sedentary individuals that bacterial species associated with health would be more abundant in the more active group, as well as a greater consumption of fruits and vegetables [9].
The literature on lifestyle patterns, including diet and physical activity, and their effect on the microbiome, identifies some differences by activity level and microbe–food matrix preferences and supports a resource environmental theoretical framework for future research. This scoping review seeks to review the role of diet and exercise-induced adaptations in shaping the gut microbiome to reveal the current state of our knowledge of their impact on health.
2. Methods
This scoping review of systematic is reported using guidelines from Joanna Briggs Institute Reviewer’s Manual and managed with JBI System for the Unified Management, Assessment, and Review of Information (SUMARI) [11,12].
2.1. Eligibility Criteria
As presented in the Table 1 PICOS chart, papers selected for inclusion were English language systematic reviews of cross-sectional, prospective cohort studies, randomized-controlled trials of either parallel or crossover design with healthy adult subjects aged 18 years or older. The exposure or intervention had to include diet pattern (Western-style, plant-based, vegan, omnivore, carbohydrate exclusion) and/or physical activity or exercise. Therapeutic interventions using pre- and pro-biotics as therapeutic agents were rejected, but studies using fermented whole foods were included. Studies should include comparisons between the type of diet and/or physical activity levels and changes in microbial composition of the gastrointestinal tract (GIT). Reviews examining probiotics outside of the context of dietary pattern and non-interventional reviews were excluded. Outcomes of the intervention or exposure should include gut microbiota composition through fecal samples and abundance composition or abundance of specific intestinal bacteria.
2.2. Search Strategy
A systematic search for systematic reviews was conducted on PubMed, CINAHL-EBSCO, Cochrane Database of Systematic Reviews, Prospective Register of Systematic Reviews (PROSPERO), PEDro, and PubMed was conducted in June 2022 from January 2012 to June 2022. The search aimed to identify systematic review papers investigating the influence of diet and/or physical activity on the microbiota composition of gut microbiota of healthy adults using a combination of MeSH and individual search terms. The search terms were microbiome OR microbiota OR gut microbiome in combination with diet AND/OR exercise AND/OR physical activity resulting in 282 articles after the elimination of duplicates. Study identification and selection are detailed in the PRISMA-ScR flow diagram (Figure 1) PRISM-ScR [13].
The initial searches produced several papers with questionable authority due to heterogeneity in reporting outcomes. Strengthening The Organization and Reporting of Microbiome Studies (STORMS) was developed from STROBE and STREGA (STrengthening the REporting of Genetic Association Studies) guidelines in 2021 to standardize the organization of essential information for papers reporting on the microbiome with the purpose of reducing reporting heterogeneity [14,15,16].
2.3. Study Selection and Data Extraction Process
Two reviewers (PGF and HF) independently screened the titles and abstracts, as well as assessed papers for eligibility. The Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA-SCR) guideline was used as a basis for reporting. As study designs and outcome assessments varied, results are presented in a narrative way. Results are presented based on the PICOS criteria in Table 1 and PRISMA 2020 flow diagram in Figure 1 for new systematic reviews [13]. JBI Critical Appraisal Checklist was used to evaluate the risk of bias in the systemic reviews.
2.4. Results
Table 2 summarizes the characteristics of the included studies and the relevant results. Over time the quality of papers that reported on the effect of lifestyle on the gut microbiota has improved, but there remains an elevated level of heterogeneity in treatment outcomes making it difficult to generalize findings.
3. Microbiome
The microbiome is a collection of bacteria, fungi, and viruses that varies by location, age, health status, diet, and physical activity levels. The dominant bacterial phyla in the human microbiome are Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria, and Verrucomicrobia [44]. Most fungi are poor colonizers of the gastrointestinal tract (GIT), but seven taxa have been found: C. albicans, Saccharomyces cerevisiae, A. niger, Penicillium sp., Pichia, Aspergillus, and Mucor and are influenced by diet and alcohol consumption [45]. Candida is the most abundant genus in the first six weeks of life in infants, with colonization being greater in those delivered vaginally [46]. Bacteria reside in the gut, as well as other body sites (genital, skin, airway)—each with its own distinct population of archaea, viruses, and eukaryotes [47]. A healthy microbiome exhibits greater microbial diversity. Low microbial diversity is observed in disease states such as cardiovascular disease, cancer, obesity, and metabolic and immune disorders. The reduction in populations within the human microbial ecosystem or intestinal dysbiosis is associated with negative health effects [48].
The viruses (virobiota) and their communities (viromes) cross the IEB and are believed to have a cooperative evolution and symbiotic relationship with their host [49]. Viral colonization of the GI tract begins after birth and gradually increases with age [50]. Human endogenous retroviruses (HERVs) have entered host germ cells, eggs, and sperm, over successive generations of infecting our ancestors and make up approximately 8% of our genome [49]. HERVs can be coopted into protecting the host and participating in protective metabolic activities such as neuroprotection and embryonic development. Activation of HERVs can affect the expression of genes involved in immunity and inflammation. Researchers demonstrated that the fasting or overfed condition affected the expression of ERV-related genes in geese, suggesting that they are influential mediators in the development of non-alcoholic liver disease [51].
4. Diet
Individuals have unique gut microbial communities due to differences in host genetics, physical activity, aging, health, and dietary composition [52,53,54,55]. Food provides nutrients and other substrates for the bacteria that reside in the gut; in return, bacteria yields not only SCFAs, but significant amounts of vitamins K2 and B12, folate, riboflavin, thiamine, and other nutrients [56,57]. Interspecies bacterial competition for life-sustaining resources drives community composition [58]. To reduce dysbiosis and systemic inflammation, a better understanding of how strain diversity, their interactions, and the metabolism of various nutrients is crucial.
The habitual diet has wide-ranging effects on human health now and in the future. Telle-Hanson, Holven, and Ulven (2018) reviewed the literature on diet and its components, microbiota, and evidence of inflammation in dietary intervention trials to develop insight into the origin of cardiovascular disease [17]. Few conclusions were made due to the heterogeneity of factors and wide latitude in which the authors of the primary studies define a “healthy diet”. Data from the American Gut Project demonstrates that plant-based and flexitarian dietary patterns with higher Healthy Eating Index 2010 (HEI-2010) scores were associated with microbiome -diversity or the extent of change in the number of microbial communities [59]. Diets rich in inflammatory foods such as refined grains, processed and red meats, fried foods, and added sugars are major factors in the causation of chronic diseases. Tools such as the Dietary Inflammatory Index (DII) are used to evaluate the inflammatory potential of an individual’s dietary pattern [60]. Fruits, vegetables, whole grains, and legumes consumption are all linked to the reduction of systemic inflammation as well as microbial diversity [61]. Consumption of whole fruits, vegetables, and legumes in a Mediterranean dietary pattern has been shown to increase fecal short-chain fatty acid (SCFA) levels, mainly due to fermentation of insoluble fibers by the most abundant phyla—Firmicutes and Bacteroidetes [62,63]. Grape and red wine, with their high polyphenol content, have been associated with beneficial changes in GIT microbial composition Proteobacteria, Fusobacteria, Firmicutes, Bacteroidetes, and B. uniformis [19]. The response to the consumption of 100% fruit juice varies by type, but orange juice has a positive effect on the gut microbiome [64,65]. The data from a short-term human trial using cherry juice suggests that participants who consumed a Western-style diet showed different gut microbiota responses than other participants due to their reduced ability to metabolize polyphenols [66]. Healthy volunteers in a month-long trial of Montmorency tart cherry concentrate had no change in species richness or microbial composition [67]. Thus, habitual dietary pattern affects the ability of individuals to receive all of the health benefits from fruits and vegetables.
In a study of food frequency and related bacterial genera in 98 participants, the authors reported that the habitual diet was correlated with separate clusters, termed “enterotypes,” primarily dominated by Bacteroides, Prevotella, and Ruminococcus [15]. The resulting dietary-enterotype framework showed that Bacteroides entero-type was highly associated with a more Westernized diet high in animal protein and saturated fats, while the Prevotella enterotype was associated with a carbohydrate-dominated diet. Vegetarians showed a mixed pattern of Prevotella and Bacteroides. Investigators in this controlled feeding trial demonstrated that by changing the diet pattern on a short-term basis to either a plant-based or animal-based diet, microbial community structure recovered to baseline bacterial composition once participants resumed their usual diet [68].
Nutrient-dense foods are higher in nutrients and lower in calories and naturally contain vitamins, minerals, fiber, or resistant starches. The MAL-ED study (Etiology, Risk Factors, and Interactions of Enteric Infections and Malnutrition and the Consequences for Child Health) followed a cohort of 1283 children ages 9 to 15 months from eight low resource populations to determine their risk of anemia, low retinol, zinc, ferritin, and high transferrin receptor (TfR) [69]. The researchers assessed diet intake, nutrient density, micronutrient status, and markers of inflammation. The authors concluded that after accounting for dietary nutrient density, there was an independent association between biomarkers of intestinal permeability and micronutrient status for children with anemia, low ferritin, and retinol levels. These data suggest that a nutrient-dense diet may be useful in reducing systemic inflammation.
A trial of a Mediterranean diet with an additional green tea supplement and minimal amounts of meat demonstrated improvement in cardiometabolic risk, weight loss, and changes in microbial abundance driven by a small low abundant non-core change in taxonomic composition [70]. The addition of a polyphenol-rich green tea supplement to the diet enhanced the abundance of Prevotella and Bifidobacterium involved in the synthesis and degradation of branched-chain amino acids. The presence of the genera Prevotella and Bifidobacterium is common in non-Western societies, and some authors have suggested that it may be a biomarker for a healthy lifestyle [71].
4.1. Western Diet
The Western-style diet, an energy-dense and nutrient-poor pattern, consists of processed meats, full-fat dairy such as cheese and ice cream, refined grains, and added sugars, which promote inflammation and chronic disease. A systematic review of 46 studies that measured the effect of Western food choices on inflammation concluded that “balanced” diets reduce the risk for chronic disease [72]. Across the 46 studies meeting inclusion criteria, individuals consuming Western-type and meat-based diets had higher markers of low-grade chronic inflammation. Results from studies on the MedDiet, a diet rich in fruits and vegetables, demonstrate that individuals following this type of diet have lower markers of low-grade inflammation. As these studies were drawn from cross-sectional observational designs, the authors concluded that in order to confirm these results, prospective trials are needed.
4.1.1. Ultra-Processed Foods
One discriminating feature between traditional and Western dietary patterns is the consumption of ultra-processed foods. The Open Food Facts database, which gathers food product information from around the world, uses the NOVA classification system to score products [73]. This system of classification has been used in cross-sectional studies of diet intake and chronic disease [74]. The NOVA classification of food has four categories: unprocessed or minimally processed foods, processed culinary ingredients, processed foods, and ultra-processed food and drink products. A systematic review and meta-analysis of cross-sectional studies on the effect of exposure to ultra-processed foods (UPF) on health status using food frequency and 24 h recall data found an increased risk of overweight, hypertension, metabolic syndrome, and low high-density lipoprotein [75]. The authors also reviewed five quality prospective-cohort studies and found an increased risk of all-cause mortality (RR 1.25, 95% CI 1.14, 1.37; p < 0.00001) for individuals who consumed elevated levels of UPF.
Food processing adds sodium, sugar, and other additives to modify the flavor, texture, and/or color. The influence of high sodium diets (HSD) is attributed to aberrant T-cell activation resulting in essential hypertension and other autoimmune diseases [76]. Experimental evidence using a rat model suggests that dietary sodium also alters the composition of gut microbial taxa, specifically Christensenellaceae and families, as well as the Erwinia and the Anaerostipes genera [77]. HSD also elevated levels of proteinuria and produced hypertension, a finding in agreement with human studies [78]. A randomized, double-blind, placebo-controlled crossover trial of a reduced-sodium diet in 145 untreated hypertensive individuals ages 30 to 75 years demonstrated increased circulating levels of SCFAs and decreased blood pressure [79]. Results for women showed a significant difference in all eight SCFAs, but there was no difference by race. In another study, a high sodium diet consumed by healthy volunteers was associated with an increase in blood pressure and an increased abundance of Prevotella, Bacteroides, and Ruminococcaceae [80]. Since “enterotypes” associated with diet have been described as being either Prevotella or Bacteroides dominant, this may suggest that there is a transition point from healthy to unhealthy, making this shift a marker for wellness [71].
Components of a processed diet may include the use of additives, particularly titanium dioxide (TiO2) and emulsifiers. These components influence the integrity of the IEB, while food additives promote biofilm formation, altering microbial composition [81,82,83,84,85,86]. Another role of food processing is the enrichment of foods with vitamins, minerals, and nutraceuticals according to regulations or for product marketing. By increasing nutrient density, fortification promotes biofilm formation under these conditions and promotes bacterial persistence to antibiotics.
Consumption of ultra-processed foods may induce low-grade gut inflammation and increase an individual’s risk for cancer and cardiovascular disease. An online survey of consumers found that they readily identified ultra-processed foods as foods being ready-to-eat and containing additives, food colorings, preservatives, and stabilizers [87]. Low-income consumers choose foods that are ultra-processed for several reasons, such as cost, convenience, and taste [75,88]. Public health educators should focus on educating consumers about how to make smarter choices by reading the food label and choosing products with few additives and minimal processing.
4.1.2. Protein
The degradation of dietary protein occurs through human and microbial proteases. Bacteria rely on carbohydrates and fiber for energy, but they are flexible and will use protein as an energy source when carbohydrate resources are low. Proteolytic fermentation produces phenols, branch-chain amino acids, sulfides, indoles, and ammonia as by-products [89]. Unlike the products of carbohydrate fermentation, colonic protein metabolites are often associated with colon cancer [90]. Researchers in one controlled human trial found that a three-week high protein diet had the effect of regulating the production of bacterial metabolites, causing an increase in the degradation of amino acids in the gut and modifying gut mucosal genes involved in cell cycle regulation [91]. Using a Human Intestinal Microbial Ecosystem (SHIME(R)) model researchers demonstrated that a high-protein diet (2.5 g/L casein) produces a different microbial community composition than a high-fiber (0.6 g/L casein) diet [92]. High-protein diets fed to mice lead to the development of obesity which was a major factor in shifting the gut microbial populations, but this effect was dependent on the diet’s protein-to-sucrose ratio [93].
One outcome of consuming diets rich in the essential amino acid tryptophan, such as dairy, poultry, and nuts, is that they favor the generation of indole and indole derivatives by colonic bacteria. Indole is a signaling molecule that regulates the host immune system by supporting epithelial barrier defense in a way that controls pathogens without producing an inflammatory response [94,95], thereby reducing chronic inflammation caused by the passage of bacteria and toxins from the gut into the bloodstream [96].
4.1.3. Fats
Trans-fat, or trans-fatty acids, are a by-product of the hydrogenation of vegetable oils to enhance shelf-life and other desirable food characteristics. Once considered to be a healthier alternative to saturated fat, there is now a consensus on the role trans fatty acids play in the development of heart disease and stroke by increasing LDL cholesterol and reducing HDL cholesterol. The association of high intakes of iTFA (industrial trans-fats) with cardiovascular disease has led to public health recommendations to reduce or eliminate their use. High intake of iTFA in mice has been shown to cause significant dysbiosis of gut microbiota [97]. A survey of the consumption of industrial trans-fatty acids reported that 22 out of 29 countries sampled found the intake of total trans-fat is currently below recommendations of 1% [98].
The ability of colonic bacteria to adapt to a fats-only source of energy for bacterial growth has been demonstrated in a fats-only medium through genes encoding for enzymes involved in fat degradation [99]. In the fats-only condition, Alistipes spp., Bilophila spp., and total Proteobacteria were favored, while SCFA-producing species, such as Bacteroides, Clostridium, and Eubacterium spp., declined. Through the use of an in vitro human gut simulator (HGS) model, the authors also demonstrated that in a fats-only medium fortified with micronutrients that lack protein or carbohydrate, bacterial communities responded with a change in community structure [99]. Colonic bacteria adaptation to a fats-only diet comes at a price to the host through a reduction in the absorption of antioxidants and an increase in the expression of bacterial virulence factors [100]. In a systematic review of the MyNewGut project data, researchers found that diets high in saturated fats degreased both richness and diversity, while those high in monounsaturated fatty acids (MUFA) produced a lower total number of bacteria, whereas a diet rich in polyunsaturated fatty acids (PUFA) resulted in no change [21].
4.1.4. Carbohydrates and Fiber
WPDs are comprised of highly refined sugars, particularly sucrose and fructose. It has been suggested that one of the causes of obesity may be an adaptation in microbial diversity due to one or more components in the Western diet [101]. Researchers using a mouse model reported that a diet high in fructose and/or sucrose, a pellet with 55% fructose-42% glucose ratio, alters the ratio of Bacteroidetes to Proteobacteria, favoring the profile associated with metabolic syndrome [102]. Metabolic syndrome increases an individual’s risk for cardiovascular disease and type 2 diabetes. Lifestyle intervention research using diet and physical activity approaches has only been partially successful in controlling the epidemic of obesity.
Diets high in fermentable carbohydrates and fiber alter the nutritional ecology of bacteria resulting in bacterial diversity, particular species that produce SCFAs such as butyrate, acetate, and propionate, as well as lactate and gasses (CO2, H2, and CH4) [89]. In a study of pregnant women, low fiber intake was associated with a gut microbiota profile favoring the fermentation of lactate and insulin resistance, while a diet rich in fruits and vegetables promoted bacteria that produce SCFAs through bacterial fermentation of plant polysaccharides [103]. Incorporating healthier eating behaviors into prenatal counseling may support a more beneficial gut microbiome.
Naturally sweet foods such as honey have long been known to have both antimicrobial and probiotic properties, although it is primarily glucose and fructose [104]. Honey promotes commensal strains such as Lactobacillus reuteri, Lactobacillus rhamnosus, and Bifidobacterium lactis while limiting the growth and adhesion of pathogenic bacteria in the host. North American maple syrup is rich in phytochemicals and oligosaccharides, as well as lignin [105,106]. Maple syrup improved insulin sensitivity and reduced non-alcoholic fatty liver in a diet-induced obese insulin-resistant rat model [107]. A randomized controlled trial in humans on the role of maple syrup on gut microbial diversity and metabolic syndrome in humans is currently underway [108].
Fermented foods are processed using techniques for “controlled microbial growth” to produce enzymes that alter food characteristics [109]. Food and beverage products have traditionally been either fermented, aged, or inoculated with bacteria and yeasts to preserve and enhance their flavor. Studies on the impact of fermented foods on the gut microbiome suggest they have a positive impact on the gut microbiome, but the quality of the evidence is unclear [110]. The generalizability of health benefits from fermentation is limited due to the undefined microbial content of starter cultures and the uniqueness of each processing plant’s bacteria and fungi communities [111]. The fermentation process has potential positive impacts, such as the release of bioactive peptides, biogenic amines, and phenolic compounds with increased antioxidant activity. Bioactive peptides and polyamines have beneficial effects on cardiovascular, immune and metabolic health, such as enhanced mineral absorption and reduced oxidative stress [112].
In addition to acting as the food source for intestinal bacteria-resistant starches, polyphenols and fiber have been shown experimentally to modulate the quantity and composition of microbiota by inducing prophages, virus-like-particles (VLPs) [113]. These bacteriophages protect the gut lining from pathogens and insert into the genome of their host’s chromosomal DNA as prophages. The bactericidal effects of VLPs are triggered by several common foods and products such as Tabasco sauce, vinegar, Kombucha, cinnamon, miso, oregano, coffee Arabica, and stevia. Although stevia appears to have positive effects on gut microbiota, it can induce heritable changes in gene expression in offspring. Experimental evidence of a recent study on the effect of maternal stevia consumption on obese rats fed a diet high in fat/sugar (HFS) combined with either aspartame or stevia resulted in second-generational effects of obesity and glucose intolerance in the offspring [114]. The proposed mechanism for the observed effect was the development of altered gene expression in the mesolimbic reward pathway associated with feeding behavior.
In the past, the mechanism behind the observed beneficial health effects of cultural superfoods, such as natto and kimchi, was unknown [115]. In the future, diets may be personalized to enhance the ecology of the gut microbiome through the targeted use of these foods.
4.1.5. Diet Supplementation
The quality of the diet shapes the health of the host and gut. For example, vitamin A deficiency has an effect on microbial community structure and gene expression [116]. The development of the gut microbiome in infancy and childhood is affected by the supply of micronutrients in the diet [117]. In regard to micronutrient supplementation, a study of Kenyan infants concluded that iron fortification adversely affects the gut microbiome by significantly increasing the population of pathogens, such as Clostridium and Escherichia/Shigella, with a reduction in beneficial Bifidobacterium which increases inflammation [118]. The authors of the study concluded that when iron is poorly absorbed by the small intestine, the resulting iron overload in the colon disrupts the bacterial population. The effect of iron and zinc fortified foods on gut microbiota was reviewed in one systematic review, and the authors reported no adverse effects in the five studies retrieved [119]. With the widespread fortification of food products to promote health and prevent nutrient deficiencies, more research on the effects of biofortification on gut bacterial populations is needed.
4.1.6. Water
Data from the American Gut project database suggests that drinking water source is associated with microbiota composition [120]. Fecal samples from individuals who consumed well water had greater β-diversity compared to those who drank bottled, filtered, or tap water. Dai et al. reported that microbes coming from drinking water supplies that have been disinfected versus non-disinfected water have a less diverse structure and function, favoring microbes that utilize fatty acids derived from microbial decomposition [121].
4.1.7. Plant-Based Diets: Vegetarian and Vegan
Vegetarian and vegan diets rich in cereals, nuts, fruit, vegetables, and legumes are the most studied in regard to gut microbial diversity and health effects. The long-term dietary pattern an individual chooses to follow will have an effect on the gut microbiota. Each dietary pattern has a different profile of gut microbiota producing distinct types of postbiotics, the compounds produced by bacteria residing in the gut that support health.
Consumption of a diet rich in plant-based foods rich in fiber, folate, and carotenoids improves resistance to inflammation and oxidative stress, promoting healthy cellular aging [122,123]. Data from the Nurses’ Health Study suggests that a diet rich in dietary fiber from cereals is positively associated with leukocyte telomere length [124]. The literature on nut consumption and the microbiome is inconsistent, with one review concluding no effect on β-diversity and increases in Clostridium, Lachnospira, and Rosburia, producers of SCFAs [27]. The Firmicutes/Bacteroidetes ratio is lower in vegans compared to omnivores and differs according to their metabolic profiles [33,125]. Meanwhile, another reviewed almonds, walnuts, hazelnuts, or pistachios, limiting the included papers to those with next-generation sequencing technology [29]. Almonds, hazelnuts, and pistachios had a small impact on microbial diversity, but walnuts were superior in changing both α- and β-diversity. The pulses are the edible seeds of the legume plant and include beans, lentils, and peas. A limited number of studies have looked at the effect of pulses on the microbiome, but those that do exist have found that whole pulses and pulse-derived flour have positive effects on diversity and richness [28]. Little is known about the components of each type of pulse and their prebiotics, fructo-oligosaccharides (FOS) and galacto-oligosaccharides (GOS) [126].
Adult whole milk consumption has been associated with an increased risk of chronic disease and reduced telomere length. Using NHANES data, a total of 3072 women and 2762 men were included in an analysis of milk consumption and telomere length [127]. Telomere length did not differ by how frequently men and women drank milk, but adults who drank full-fat or 2% milk had significantly shorter telomeres than those consuming nonfat or 1% milk, and that difference was clinically meaningful. This finding is consistent with the dietary guidelines, which encourage low-fat dairy and discourages the consumption of full-fat milk for adults. The effect of unfermented dairy on gut microbiota was a reduction in richness and diversity, while yogurt and kefir increased both Lactobacillus and Bifidobacterium [26]. Habitual vegans and vegetarians, when compared to omnivores, have a greater abundance of fiber-degrading species and lactic acid bacteria. Yet, when vegan and vegetarian diets were compared, no consistent difference in microbiota composition was found [18,32]. Both diets are rich in polyphenols that increase Bifidobacterium and Lactobacillus.
Diets excluding grains, dairy products, salt, and refined sugar, also known as ketogenic or modern paleolithic diets, have become popular in the past few years. A study comparing the gut microbiome profiles of 15 urban-dwelling Italians practicing a paleolithic diet to populations following a Mediterranean diet and traditional hunter-gatherer groups found major differences between groups, including a relative abundance of Bacteroides and other organisms in the paleo diet sample typically associated with high animal protein and saturated fat diets and an altered bile acid profile associated with inflammatory bowel and colon cancer [128]. In a review of RCTs, studies using whole-grain versus low-fiber diets found that fiber increases microbial diversity, abundance, and beneficial metabolites [20,23]. Gluten-free diets in healthy subjects accompanied by a reduction in resistant carbohydrate intake results in lower levels of Bifidobacterium, B. longum, and Lactobacillus and more unhealthy species [129,130]. Based on this information, dietary adherence to paleolithic diets over the long term would not be advised.
4.1.8. Mediterranean
The CARDIVEG (Cardiovascular Prevention with Vegetarian Diet) study, a three-month crossover design comparing the Mediterranean to a vegetarian diet, found that there was no statistical difference in microbial diversity and no change in the Firmicutes/Bacteroidetes ratio between the two diets [58]. Short-term dietary changes produced only small variations in gut microbiota composition. In another study, habitual Mediterranean diet and vegetarian diet consumption were associated with increased levels of fecal SCFAs, Prevotella and Firmicutes [10]. The effects of the Mediterranean diet on the gut microbiome, in general, have a positive impact on the Firmicutes/Bacteroides ratio, but results were not consistent across studies in a recent systematic review of prospective and cross-sectional studies [37]. The authors suggest that this may be due to differences in diet adherence between participants. The PREDIMED-Plus study looked at the influence of an energy-restricted Mediterranean diet along with physical activity on the microbiome of older adults [131]. Controls were given instruction on the Mediterranean diet with no calorie restrictions or physical activity guidance. First-year results reported that the physically active and calorie-restricted group had a significant increase in the Bacteriodetes/Firmicutes ratio and greater weight loss than controls. However, both groups saw a shift to more SCFA-producing genera, suggesting that the Mediterranean diet selectively enhances specific anaerobic gut bacteria responsible for the fermentation of complex carbohydrates.
Seafood is an important part of the Mediterranean, contributing to the intake of omega-3 fatty acids [132] but also influences the microbiome. In a crossover design study comparing the metabolic markers and fecal bacteria before and after a diet with and without lean seafood, lean seafood resulted in higher serum TMA and fecal TMA, presumably due to increased bacterial degradation of TMAO [133,134]. The fecal microbiota analysis showed an increase in Firmicutes and decreased Bacteroides, as well as higher amounts of Clostridium Clusters IV, producers of butyrate in the gut. Four weeks after the lean seafood intervention, there were significant improvements in cardiovascular risk factors, HDL, and a decrease in circulating triacylglycerol (TAG).
5. Physical Activity
In general, physical activity both lowers Bacteroidetes and raises Firmicutes. Moderate physical activity, defined as endurance activity performed 3 days a week for 30 to 60 min at 60% of heart rate reserve, increases both microbial diversity and richness, but authors concluded in one systematic review that the overall strength of the evidence is weak due to poor control over dietary patterns confounding the data interpretation [22]. A review of observational studies showed that only 5/9 showed that a higher level of physical activity or cardiovascular fitness was associated with greater α-diversity [24]. The effect of exercise, in general, produces an increase in Bifidobacterium, Lactobacilli, and Akkermansia [135]. Athletes have higher microbial diversity and produce an abundance of short-chain fatty acids (SCFA) [22]. Fecal and urine samples from elite cyclists participating in the 2016 Olympics were characterized by different training loads and amounts of time spent in static and dynamic activities [136]. Diet intake did not vary significantly between sports classification groups. The analysis demonstrated differences in microbial composition and SCFA metabolites detected in both feces and urine. A study of recreational and professional cyclists reported an expected abundance of Bacteroides, Faecalibacterium, and Eubacterium, along with increased levels of Methanobrevibacter smithii, indicating an upregulation of energy pathways using methane, a high-energy fuel [137]. This adaptation in athletes is beneficial because methanogens, such as Methanobrevibacter smithii, must produce methane to conserve energy for growth rather than produce it as a by-product of metabolism [138]. Mohr and colleagues have published a comprehensive review of the literature on the gut microbiota of various sports [135].
Estaki et al. examined the fitness levels of young adults as measured by VO2 peak and fecal microbiota and found that an individual’s fitness category was responsible for 20% of the variation associated with taxonomic richness [139]. The authors examined diet as a confounding variable and found that only one component was significant for beta diversity—protein. Protein intake and age accounted for an additional 7.9 and 2.3%, respectively, of the community beta diversity. A systematic review of 10 studies examining the influence of physical activity on the human gut microbiota composition concluded that most studies did not account for the influence of diet, and further studies of dietary protein levels are indicated to confirm this finding [31]. During periods of intense activity, the gut wall junctions lose their integrity, permitting materials from the lumen to translocate into the bloodstream resulting in increased inflammation due to IL-6 production [53]. Overall, the shift in the microbiome varies by the conditioning level of the athlete, with well-conditioned individuals benefiting from species involved in metabolic pathways producing amino acids and the metabolism of carbohydrates and fiber [53]. The reciprocal relationship between diet, activity, and microbial communities, which has been labeled the “nutrition-microbiota-physical activity triad”, represents a regulatory role for microbial communities [140].
Moderate to vigorous activity of less than an hour enhances anti-pathogen activity through the release of NK cells and CD8+ T lymphocytes, which over time improves the monitoring ability of the immune system to detect pathogens and tumors [141]. When conditioned athletes were compared to others who met the general recommendations for exercise, the less conditioned group showed no meaningful change in microbial richness or diversity [34,36]. A review of both interventional and observational studies found no consistency between trials for changes in microbial diversity, but there were decreases in pathogenic species and increases in beneficial taxa [9,35]. These studies support the conclusion that the benefits of exercise to the microbiome and its influence on host immune function are on a continuum, but the optimal has yet to be determined [142].
6. Discussion
6.1. Introduction
The aim of this scoping review was to identify lifestyle factors influencing the gut microbiome and host health. The microbiome is a dynamic system that affects not only the host’s internal environment but is also driven by external factors such as the genome and the health behaviors of the human host [143]. We have argued that resource availability is a major driver of microbial community composition and sustainability.
However, no one model can capture or predict all of the host–microbiome-lifestyle interactions. Stegan, Bottos, and Jansson have put forth a unified conceptual framework upon which to study time-dependent interactions, which may lead to manipulating microbes to support health and human performance [144]. Within this framework, an individual’s history of diet and physical activity influences the internal dynamics of the community structure and dynamics. External forces, such as resource availability, impact community density, and composition. The use of data-intensive modeling using the triad of history, external forces, and internal dynamics will expand our understanding of microbial communities and human health.
6.2. Limitations and Strengths of This Review
The authors were challenged by the pace of change in the emerging fields of microbiology within nutrition and exercise science but endeavored to find the highest level of evidence to begin integrating this knowledge for public health researchers and educators. The strength of this review is its recency, and its weakness is that the heterogeneity in the technologies and reported outcomes by the authors make data extraction for a meta-analysis impossible at this time. The difficulty in making general recommendations is that it was rare to find a physical activity paper within our search criteria that controlled for diet [31,131]. Animal studies demonstrate that exercise performance and conditioning that takes place in long-term interventions have positive effects on both the microbiome and host metabolism [145]. Transferring this model to human studies will prove challenging because of the need to maintain a controlled research diet over the length of the study. The “captive” environment needed to perform research in animals may reduce the diversity of the microbiome by limiting exposure to other mammals and the environment [146]. Thus, systematic reviews of studies using natural experimental designs on well-defined human sub-populations may yield the best available evidence.
7. Conclusions
Promoting a healthy gut microbiome is crucial to protecting and supporting the health of an individual. Food and activity choices can have a dramatic impact on the gut microbiome. In turn, lactate acid bacteria may have a positive influence on sports performance and recovery through the conversion of lactate to propionate. Diets that are based on the Healthy Eating Index, such as the Mediterranean diet, promote the development of gut microbial diversity, which provides beneficial postbiotics such as SCFAs and butyrate. Polyphenols, found in abundance in plant foods, yield microbial metabolites that reduce inflammation and promote human health. Including prebiotics and probiotics as a functional food-based approach to manipulating the microbiome may prevent or reduce the progression of diet-related diseases. When making food choices, individuals should follow a diet that is rich in fruits, vegetables, whole grains, legumes, nuts, and seeds and low in processed foods, refined grains, and meats. Future versions of the Physical Activity and Dietary Guidelines for Americans should also review the literature on the impact of current recommendations for exercise on the abundance and diversity of the microbiome [147,148]. Our current state of knowledge on how changes in the microbiota determine long-term health and the aging process is lacking. Researchers manipulating the microbiome with a targeted probiotic approach should consider how microbial dynamics interplay with diet and physical activity in the study design.
Author Contributions
Conceptualization, P.G.-F. and H.F.; formal analysis, P.G.-F., M.D.I. and H.F.; data curation, P.G.-F.; writing—original draft preparation, P.G.-F., M.D.I. and H.F.; writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Williams, A. Shining a Light on the Resource Curse: An Empirical Analysis of the Relationship Between Natural Resources, Transparency, and Economic Growth. World Dev. 2011, 39, 490–505. [Google Scholar] [CrossRef]
- Swain Ewald, H.A.; Ewald, P.W. Natural Selection, The Microbiome, and Public Health. Yale J. Biol. Med. 2018, 91, 445–455. [Google Scholar]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 2016, 167, 1339–1353.e1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, M.E.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilg, H.; Moschen, A.R. Food, immunity, and the microbiome. Gastroenterology 2015, 148, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Warburton, D.E.R.; Bredin, S.S.D. Health benefits of physical activity: A systematic review of current systematic reviews. Curr. Opin. Cardiol. 2017, 32, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Jollet, M.; Nay, K.; Chopard, A.; Bareille, M.P.; Beck, A.; Ollendorff, V.; Vernus, B.; Bonnieu, A.; Mariadassou, M.; Rué, O.; et al. Does Physical Inactivity Induce Significant Changes in Human Gut Microbiota? New Answers Using the Dry Immersion Hypoactivity Model. Nutrients 2021, 13, 3865. [Google Scholar] [CrossRef] [PubMed]
- Bressa, C.; Bailen-Andrino, M.; Perez-Santiago, J.; Gonzalez-Soltero, R.; Perez, M.; Montalvo-Lominchar, M.G.; Mate-Munoz, J.L.; Dominguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cronin, O.; Barton, W.; Skuse, P.; Penney, N.C.; Garcia-Perez, I.; Murphy, E.F.; Woods, T.; Nugent, H.; Fanning, A.; Melgar, S.; et al. A Prospective Metagenomic and Metabolomic Analysis of the Impact of Exercise and/or Whey Protein Supplementation on the Gut Microbiome of Sedentary Adults. mSystems 2018, 3, e00044-18. [Google Scholar] [CrossRef] [Green Version]
- Aromataris, E.; Munn, Z. JBI Manual for Evidence Synthesis; JBI: Adelaide, Australia, 2020. [Google Scholar]
- Peters, M.D.J.; Marnie, C.; Tricco, A.C.; Pollock, D.; Munn, Z.; Alexander, L.; McInerney, P.; Godfrey, C.M.; Khalil, H. Updated methodological guidance for the conduct of scoping reviews. JBI Evid. Implement. 2021, 19, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Mirzayi, C.; Renson, A.; Genomic Standards, C.; Massive, A.; Quality Control, S.; Zohra, F.; Elsafoury, S.; Geistlinger, L.; Kasselman, L.J.; Eckenrode, K.; et al. Reporting guidelines for human microbiome research: The STORMS checklist. Nat. Med. 2021, 27, 1885–1892. [Google Scholar] [CrossRef] [PubMed]
- White, R.G.; Hakim, A.J.; Salganik, M.J.; Spiller, M.W.; Johnston, L.G.; Kerr, L.; Kendall, C.; Drake, A.; Wilson, D.; Orroth, K.; et al. Strengthening the Reporting of Observational Studies in Epidemiology for respondent-driven sampling studies: "STROBE-RDS" statement. J. Clin. Epidemiol. 2015, 68, 1463–1471. [Google Scholar] [CrossRef] [Green Version]
- Little, J.; Higgins, J.P.; Ioannidis, J.P.; Moher, D.; Gagnon, F.; von Elm, E.; Khoury, M.J.; Cohen, B.; Davey-Smith, G.; Grimshaw, J.; et al. Strengthening the reporting of genetic association studies (STREGA): An extension of the STROBE statement. Eur. J. Epidemiol. 2009, 24, 37–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telle-Hansen, V.H.; Holven, K.B.; Ulven, S.M. Impact of a Healthy Dietary Pattern on Gut Microbiota and Systemic Inflammation in Humans. Nutrients 2018, 10, 1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trefflich, I.; Jabakhanji, A.; Menzel, J.; Blaut, M.; Michalsen, A.; Lampen, A.; Abraham, K.; Weikert, C. Is a vegan or a vegetarian diet associated with the microbiota composition in the gut? Results of a new cross-sectional study and systematic review. Crit. Rev. Food. Sci. Nutr. 2019, 60, 2990–3004. [Google Scholar] [CrossRef]
- Nash, V.; Ranadheera, C.S.; Georgousopoulou, E.N.; Mellor, D.D.; Panagiotakos, D.B.; McKune, A.J.; Kellett, J.; Naumovski, N. The effects of grape and red wine polyphenols on gut microbiota—A systematic review. Food Res. Int. 2018, 113, 277–287. [Google Scholar] [CrossRef] [PubMed]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [Green Version]
- Wolters, M.; Ahrens, J.; Romani-Perez, M.; Watkins, C.; Sanz, Y.; Benitez-Paez, A.; Stanton, C.; Gunther, K. Dietary fat, the gut microbiota, and metabolic health—A systematic review conducted within the MyNewGut project. Clin. Nutr. 2019, 38, 2504–2520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, C.M.; Davy, B.M.; Hulver, M.W.; Neilson, A.P.; Bennett, B.J.; Davy, K.P. Does Exercise Alter Gut Microbial Composition? A Systematic Review. Med. Sci. Sports Exerc. 2019, 51, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, A.; Adolphus, K. The Effects of Intact Cereal Grain Fibers, Including Wheat Bran on the Gut Microbiota Composition of Healthy Adults: A Systematic Review. Front. Nutr. 2019, 6, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz-Alvarez, L.; Xu, H.; Martinez-Tellez, B. Influence of Exercise on the Human Gut Microbiota of Healthy Adults: A Systematic Review. Clin. Transl. Gastroenterol. 2020, 11, e00126. [Google Scholar] [CrossRef] [PubMed]
- Tzemah Shahar, R.; Koren, O.; Matarasso, S.; Shochat, T.; Magzal, F.; Agmon, M. Attributes of Physical Activity and Gut Microbiome in Adults: A Systematic Review. Int. J. Sports Med. 2020, 41, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Aslam, H.; Marx, W.; Rocks, T.; Loughman, A.; Chandrasekaran, V.; Ruusunen, A.; Dawson, S.L.; West, M.; Mullarkey, E.; Pasco, J.A.; et al. The effects of dairy and dairy derivatives on the gut microbiota: A systematic literature review. Gut Microbes 2020, 12, 1799533. [Google Scholar] [CrossRef]
- Creedon, A.C.; Hung, E.S.; Berry, S.E.; Whelan, K. Nuts and their Effect on Gut Microbiota, Gut Function and Symptoms in Adults: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Nutrients 2020, 12, 2347. [Google Scholar] [CrossRef] [PubMed]
- Marinangeli, C.P.F.; Harding, S.V.; Zafron, M.; Rideout, T.C. A systematic review of the effect of dietary pulses on microbial populations inhabiting the human gut. Benef. Microbes 2020, 11, 457–468. [Google Scholar] [CrossRef]
- Fitzgerald, E.; Lambert, K.; Stanford, J.; Neale, E.P. The effect of nut consumption (tree nuts and peanuts) on the gut microbiota of humans: A systematic review. Br. J. Nutr. 2021, 125, 508–520. [Google Scholar] [CrossRef]
- Aya, V.; Flórez, A.; Perez, L.; Ramírez, J.D. Association between physical activity and changes in intestinal microbiota composition: A systematic review. PLoS ONE 2021, 16, e0247039. [Google Scholar] [CrossRef] [PubMed]
- Dorelli, B.; Gallè, F.; De Vito, C.; Duranti, G.; Iachini, M.; Zaccarin, M.; Preziosi Standoli, J.; Ceci, R.; Romano, F.; Liguori, G.; et al. Can Physical Activity Influence Human Gut Microbiota Composition Independently of Diet? A Systematic Review. Nutrients 2021, 13, 1890. [Google Scholar] [CrossRef]
- Gibiino, G.; De Siena, M.; Sbrancia, M.; Binda, C.; Sambri, V.; Gasbarrini, A.; Fabbri, C. Dietary Habits and Gut Microbiota in Healthy Adults: Focusing on the Right Diet. A Systematic Review. Int. J. Mol. Sci. 2021, 22, 6728. [Google Scholar] [CrossRef] [PubMed]
- Losno, E.A.; Sieferle, K.; Perez-Cueto, F.J.A.; Ritz, C. Vegan Diet and the Gut Microbiota Composition in Healthy Adults. Nutrients 2021, 13, 2402. [Google Scholar] [CrossRef] [PubMed]
- Pinart, M.; Dotsch, A.; Schlicht, K.; Laudes, M.; Bouwman, J.; Forslund, S.K.; Pischon, T.; Nimptsch, K. Gut Microbiome Composition in Obese and Non-Obese Persons: A Systematic Review and Meta-Analysis. Nutrients 2021, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.; Gibson, G.R.; Walton, G.E.; Magistro, D.; Kinnear, W.; Hunter, K. Systematic Review of the Effects of Exercise and Physical Activity on the Gut Microbiome of Older Adults. Nutrients 2022, 14, 674. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, S.; Bonavolontà, V.; Poli, L.; Clemente, F.M.; De Candia, M.; Carvutto, R.; Silva, A.F.; Badicu, G.; Greco, G.; Fischetti, F. The Relationship between Physical Activity, Physical Exercise, and Human Gut Microbiota in Healthy and Unhealthy Subjects: A Systematic Review. Biology 2022, 11, 479. [Google Scholar] [CrossRef]
- Kimble, R.; Gouinguenet, P.; Ashor, A.; Stewart, C.; Deighton, K.; Matu, J.; Griffiths, A.; Malcomson, F.C.; Joel, A.; Houghton, D.; et al. Effects of a mediterranean diet on the gut microbiota and microbial metabolites: A systematic review of randomized controlled trials and observational studies. Crit. Rev. Food Sci. Nutr. 2022, 1–22. [Google Scholar] [CrossRef]
- Lo, C.K.; Mertz, D.; Loeb, M. Newcastle-Ottawa Scale: Comparing reviewers’ to authors’ assessments. BMC Med. Res. Methodol. 2014, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.; Green, S. Cochrane Handbook for Systematic Reviews of Interventions; John Wiley & Sons Ltd.: Chichester, UK, 2011. [Google Scholar]
- Clark, H.D.; Wells, G.A.; Huet, C.; McAlister, F.A.; Salmi, L.R.; Fergusson, D.; Laupacis, A. Assessing the quality of randomized trials: Reliability of the Jadad scale. Control Clin. Trials 1999, 20, 448–452. [Google Scholar] [CrossRef]
- The Joanna Briggs Institute Levels of Evidence and Grades of Recommendation Working Party 2014. Joanna Briggs Institute Levels of Evidence and Grades of Recommendation; The Joanna Briggs Institute: Adelaide, Australia, 2015. [Google Scholar]
- Handu, D.; Moloney, L.; Wolfram, T.; Ziegler, P.; Acosta, A.; Steiber, A. Academy of Nutrition and Dietetics Methodology for Conducting Systematic Reviews for the Evidence Analysis Library. J. Acad. Nutr. Diet. 2016, 116, 311–318. [Google Scholar] [CrossRef]
- Sterne, J.A.; Hernán, M.A.; Reeves, B.C.; Savović, J.; Berkman, N.D.; Viswanathan, M.; Henry, D.; Altman, D.G.; Ansari, M.T.; Boutron, I.; et al. ROBINS-I: A tool for assessing risk of bias in non-randomised studies of interventions. BMJ 2016, 355, i4919. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.R.; Pop, M.; Deboy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, J. Food Habit Associated Mycobiota Composition and Their Impact on Human Health. Front. Nutr. 2021, 8, 773577. [Google Scholar] [CrossRef]
- Henderickx, J.G.E.; de Weerd, H.; Groot Jebbink, L.J.; van Zoeren-Grobben, D.; Hemels, M.A.C.; van Lingen, R.A.; Knol, J.; Belzer, C. The first fungi: Mode of delivery determines early life fungal colonization in the intestine of preterm infants. Microbiome Res. Rep. 2022, 1, 7. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The healthy human microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriss, M.; Hazleton, K.Z.; Nusbacher, N.M.; Martin, C.G.; Lozupone, C.A. Low diversity gut microbiota dysbiosis: Drivers, functional implications and recovery. Curr. Opin. Microbiol. 2018, 44, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Luganini, A.; Gribaudo, G. Retroviruses of the Human Virobiota: The Recycling of Viral Genes and the Resulting Advantages for Human Hosts During Evolution. Front. Microbiol. 2020, 11, 1140. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Bushman, F.D. The human virome: Assembly, composition and host interactions. Nat. Rev. Microbiol. 2021, 19, 514–527. [Google Scholar] [CrossRef]
- Liu, T.; Xing, Y.; Fan, X.; Chen, Z.; Zhao, C.; Liu, L.; Zhao, M.; Hu, X.; Dong, B.; Wang, J.; et al. Fasting and overfeeding affect the expression of the immunity- or inflammation-related genes in the liver of poultry via endogenous retrovirus. Poult. Sci. 2021, 100, 973–981. [Google Scholar] [CrossRef]
- Kurilshikov, A.; Medina-Gomez, C.; Bacigalupe, R.; Radjabzadeh, D.; Wang, J.; Demirkan, A.; Le Roy, C.I.; Raygoza Garay, J.A.; Finnicum, C.T.; Liu, X.; et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 2021, 53, 156–165. [Google Scholar] [CrossRef]
- Clauss, M.; Gérard, P.; Mosca, A.; Leclerc, M. Interplay Between Exercise and Gut Microbiome in the Context of Human Health and Performance. Front. Nutr. 2021, 8, 305. [Google Scholar] [CrossRef] [PubMed]
- Wilmanski, T.; Diener, C.; Rappaport, N.; Patwardhan, S.; Wiedrick, J.; Lapidus, J.; Earls, J.C.; Zimmer, A.; Glusman, G.; Robinson, M.; et al. Gut microbiome pattern reflects healthy ageing and predicts survival in humans. Nat. Metab. 2021, 3, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Polo, A.; Arora, K.; Ameur, H.; Di Cagno, R.; De Angelis, M.; Gobbetti, M. Gluten-free diet and gut microbiome. J. Cereal Sci. 2020, 95, 103058. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Brancaccio, M.; Mennitti, C.; Cesaro, A.; Fimiani, F.; Vano, M.; Gargiulo, B.; Caiazza, M.; Amodio, F.; Coto, I.; D’Alicandro, G.; et al. The Biological Role of Vitamins in Athletes’ Muscle, Heart and Microbiota. Int. J. Environ. Res. Public Health 2022, 19, 1249. [Google Scholar] [CrossRef] [PubMed]
- Patnode, M.L.; Beller, Z.W.; Han, N.D.; Cheng, J.; Peters, S.L.; Terrapon, N.; Henrissat, B.; Le Gall, S.; Saulnier, L.; Hayashi, D.K.; et al. Interspecies Competition Impacts Targeted Manipulation of Human Gut Bacteria by Fiber-Derived Glycans. Cell 2019, 179, 59–73.e13. [Google Scholar] [CrossRef]
- Cotillard, A.; Cartier-Meheust, A.; Litwin, N.S.; Chaumont, S.; Saccareau, M.; Lejzerowicz, F.; Tap, J.; Koutnikova, H.; Lopez, D.G.; McDonald, D.; et al. A posteriori dietary patterns better explain variations of the gut microbiome than individual markers in the American Gut Project. Am. J. Clin. Nutr. 2022, 115, 432–443. [Google Scholar] [CrossRef]
- Tabung, F.K.; Steck, S.E.; Zhang, J.; Ma, Y.; Liese, A.D.; Agalliu, I.; Hingle, M.; Hou, L.; Hurley, T.G.; Jiao, L.; et al. Construct validation of the dietary inflammatory index among postmenopausal women. Ann. Epidemiol. 2015, 25, 398–405. [Google Scholar] [CrossRef] [Green Version]
- Asnicar, F.; Berry, S.E.; Valdes, A.M.; Nguyen, L.H.; Piccinno, G.; Drew, D.A.; Leeming, E.; Gibson, R.; Le Roy, C.; Khatib, H.A.; et al. Microbiome connections with host metabolism and habitual diet from 1098 deeply phenotyped individuals. Nat. Med. 2021, 27, 321–332. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.S.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut 2020, 69, 1218–1228. [Google Scholar] [CrossRef] [Green Version]
- Perri, M.R.; Romano, C.; Marrelli, M.; Zicarelli, L.; Toma, C.C.; Basta, D.; Conforti, F.; Statti, G. Beneficial Role of Fruits, Their Juices, and Freeze-Dried Powders on Inflammatory Bowel Disease and Related Dysbiosis. Plants 2021, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.C.D.; Cecatti, C.; Fidelix, M.P.; Adorno, M.A.T.; Sakamoto, I.K.; Cesar, T.B.; Sivieri, K. Effect of Daily Consumption of Orange Juice on the Levels of Blood Glucose, Lipids, and Gut Microbiota Metabolites: Controlled Clinical Trials. J. Med. Food 2019, 22, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Mayta-Apaza, A.C.; Pottgen, E.; De Bodt, J.; Papp, N.; Marasini, D.; Howard, L.; Abranko, L.; Van de Wiele, T.; Lee, S.O.; Carbonero, F. Impact of tart cherries polyphenols on the human gut microbiota and phenolic metabolites in vitro and in vivo. J. Nutr. Biochem. 2018, 59, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Hillman, A.R.; Chrismas, B.C.R. Thirty Days of Montmorency Tart Cherry Supplementation Has No Effect on Gut Microbiome Composition, Inflammation, or Glycemic Control in Healthy Adults. Front. Nutr. 2021, 8, 733057. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- McCormick, B.J.J.; Murray-Kolb, L.E.; Lee, G.O.; Schulze, K.J.; Ross, A.C.; Bauck, A.; Lima, A.A.M.; Maciel, B.L.L.; Kosek, M.N.; Seidman, J.C.; et al. Intestinal permeability and inflammation mediate the association between nutrient density of complementary foods and biochemical measures of micronutrient status in young children: Results from the MAL-ED study. Am. J. Clin. Nutr. 2019, 110, 1015–1025. [Google Scholar] [CrossRef] [PubMed]
- Rinott, E.; Meir, A.Y.; Tsaban, G.; Zelicha, H.; Kaplan, A.; Knights, D.; Tuohy, K.; Scholz, M.U.; Koren, O.; Stampfer, M.J.; et al. The effects of the Green-Mediterranean diet on cardiometabolic health are linked to gut microbiome modifications: A randomized controlled trial. Genome Med. 2022, 14, 29. [Google Scholar] [CrossRef]
- Gorvitovskaia, A.; Holmes, S.P.; Huse, S.M. Interpreting Prevotella and Bacteroides as biomarkers of diet and lifestyle. Microbiome 2016, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Barbaresko, J.; Koch, M.; Schulze, M.B.; Nothlings, U. Dietary pattern analysis and biomarkers of low-grade inflammation: A systematic literature review. Nutr. Rev. 2013, 71, 511–527. [Google Scholar] [CrossRef]
- Open Food Facts. Nova Groups for Food Processing. Available online: https://world.openfoodfacts.org/nova (accessed on 17 November 2019).
- Monteiro, C.A.; Cannon, G.; Moubarac, J.C.; Levy, R.B.; Louzada, M.L.C.; Jaime, P.C. The UN Decade of Nutrition, the NOVA food classification and the trouble with ultra-processing. Public Health Nutr. 2018, 21, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Pagliai, G.; Dinu, M.; Madarena, M.P.; Bonaccio, M.; Iacoviello, L.; Sofi, F. Consumption of ultra-processed foods and health status: A systematic review and meta-analysis. Br. J. Nutr. 2021, 125, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Rucker, A.J.; Rudemiller, N.P.; Crowley, S.D. Salt, Hypertension, and Immunity. Annu. Rev. Physiol. 2018, 80, 283–307. [Google Scholar] [CrossRef] [PubMed]
- Bier, A.; Braun, T.; Khasbab, R.; Di Segni, A.; Grossman, E.; Haberman, Y.; Leibowitz, A. A High Salt Diet Modulates the Gut Microbiota and Short Chain Fatty Acids Production in a Salt-Sensitive Hypertension Rat Model. Nutrients 2018, 10, 1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamler, J.; Chan, Q.; Daviglus, M.L.; Dyer, A.R.; Van Horn, L.; Garside, D.B.; Miura, K.; Wu, Y.; Ueshima, H.; Zhao, L.; et al. Relation of Dietary Sodium (Salt) to Blood Pressure and Its Possible Modulation by Other Dietary Factors: The INTERMAP Study. Hypertension 2018, 71, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, F.J.; Dong, Y.; Huang, Y.; Wang, C.; Harshfield, G.A.; Zhu, H. Modest Sodium Reduction Increases Circulating Short-Chain Fatty Acids in Untreated Hypertensives: A Randomized, Double-Blind, Placebo-Controlled Trial. Hypertension 2020, 76, 73–79. [Google Scholar] [CrossRef]
- Ferguson, J.F.; Aden, L.A.; Barbaro, N.R.; Van Beusecum, J.P.; Xiao, L.; Simmons, A.J.; Warden, C.; Pasic, L.; Himmel, L.E.; Washington, M.K.; et al. High dietary salt-induced dendritic cell activation underlies microbial dysbiosis-associated hypertension. JCI Insight 2019, 5, e126241. [Google Scholar] [CrossRef] [Green Version]
- Dudefoi, W.; Moniz, K.; Allen-Vercoe, E.; Ropers, M.H.; Walker, V.K. Impact of food grade and nano-TiO2 particles on a human intestinal community. Food Chem. Toxicol. 2017, 106, 242–249. [Google Scholar] [CrossRef]
- Guo, Z.; Martucci, N.J.; Moreno-Olivas, F.; Tako, E.; Mahler, G.J. Titanium Dioxide Nanoparticle Ingestion Alters Nutrient Absorption in an In Vitro Model of the Small Intestine. Nano Impact 2017, 5, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Everard, A. Keeping gut lining at bay: Impact of emulsifiers. Trends Endocrinol. Metab. 2015, 26, 273–274. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Changes in intestinal tight junction permeability associated with industrial food additives explain the rising incidence of autoimmune disease. Autoimmun. Rev. 2015, 14, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Wheildon, N.; Ishikawa, S. Food Additive P-80 Impacts Mouse Gut Microbiota Promoting Intestinal Inflammation, Obesity and Liver Dysfunction. SOJ Microbiol. Infect. Dis. 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lock, J.Y.; Carlson, T.L.; Wang, C.M.; Chen, A.; Carrier, R.L. Acute Exposure to Commonly Ingested Emulsifiers Alters Intestinal Mucus Structure and Transport Properties. Sci. Rep. 2018, 8, 10008. [Google Scholar] [CrossRef] [PubMed]
- Ares, G.; Vidal, L.; Allegue, G.; Gimenez, A.; Bandeira, E.; Moratorio, X.; Molina, V.; Curutchet, M.R. Consumers’ conceptualization of ultra-processed foods. Appetite 2016, 105, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Hawk, T.; Aggarwal, A.; Drewnowski, A. Characterizing Ultra-Processed Foods by Energy Density, Nutrient Density, and Cost. Front. Nutr. 2019, 6, 70. [Google Scholar] [CrossRef] [Green Version]
- Hamer, H.M.; De Preter, V.; Windey, K.; Verbeke, K. Functional analysis of colonic bacterial metabolism: Relevant to health? Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1-9. [Google Scholar] [CrossRef]
- Windey, K.; De Preter, V.; Verbeke, K. Relevance of protein fermentation to gut health. Mol. Nutr. Food Res. 2012, 56, 184–196. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Liu, H.; Brown, M.A.; Qiao, S. Dietary Protein and Gut Microbiota Composition and Function. Curr. Protein Pept. Sci. 2019, 20, 145–154. [Google Scholar] [CrossRef]
- Marzorati, M.; Vilchez-Vargas, R.; Bussche, J.V.; Truchado, P.; Jauregui, R.; El Hage, R.A.; Pieper, D.H.; Vanhaecke, L.; Van de Wiele, T. High-fiber and high-protein diets shape different gut microbial communities, which ecologically behave similarly under stress conditions, as shown in a gastrointestinal simulator. Mol. Nutr. Food Res. 2017, 61, 1600150. [Google Scholar] [CrossRef] [Green Version]
- Kiilerich, P.; Myrmel, L.S.; Fjaere, E.; Hao, Q.; Hugenholtz, F.; Sonne, S.B.; Derrien, M.; Pedersen, L.M.; Petersen, R.K.; Mortensen, A.; et al. Effect of a long-term high-protein diet on survival, obesity development, and gut microbiota in mice. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E886–E899. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.H. Immune regulation by microbiome metabolites. Immunology 2018, 154, 220–229. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, J. Indole as an intercellular signal in microbial communities. FEMS Microbiol. Rev. 2010, 34, 426–444. [Google Scholar] [CrossRef] [PubMed]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 19376–19387. [Google Scholar] [CrossRef]
- Ge, Y.; Liu, W.; Tao, H.; Zhang, Y.; Liu, L.; Liu, Z.; Qiu, B.; Xu, T. Effect of industrial trans-fatty acids-enriched diet on gut microbiota of C57BL/6 mice. Eur. J. Nutr. 2019, 58, 2625–2638. [Google Scholar] [CrossRef]
- Wanders, A.J.; Zock, P.L.; Brouwer, I.A. Trans Fat Intake and Its Dietary Sources in General Populations Worldwide: A Systematic Review. Nutrients 2017, 9, 840. [Google Scholar] [CrossRef] [PubMed]
- Agans, R.; Gordon, A.; Kramer, D.L.; Perez-Burillo, S.; Rufian-Henares, J.A.; Paliy, O. Dietary Fatty Acids Sustain the Growth of the Human Gut Microbiota. Appl. Environ. Microbiol. 2018, 84, e01525-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the gut to the peripheral tissues: The multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, A.N.; Chassard, C.; Lacroix, C. Gut microbial adaptation to dietary consumption of fructose, artificial sweeteners and sugar alcohols: Implications for host-microbe interactions contributing to obesity. Obes. Rev. 2012, 13, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Do, M.H.; Lee, E.; Oh, M.J.; Kim, Y.; Park, H.Y. High-Glucose or -Fructose Diet Cause Changes of the Gut Microbiota and Metabolic Disorders in Mice without Body Weight Change. Nutrients 2018, 10, 761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Arango, L.F.; Barrett, H.L.; Wilkinson, S.A.; Callaway, L.K.; McIntyre, H.D.; Morrison, M.; Nitert, M.D. Low dietary fiber intake increases Collinsella abundance in the gut microbiota of overweight and obese pregnant women. Gut Microbes 2017, 9, 189–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, A.; Quek, S.-Y.; Gutierrez-Maddox, N.; Gao, Y.; Shu, Q. Effect of honey in improving the gut microbial balance. Food Qual. Saf. 2017, 1, 107–115. [Google Scholar] [CrossRef]
- Perkins, T.D.; van den Berg, A.K. Maple syrup-production, composition, chemistry, and sensory characteristics. Adv. Food Nutr. Res. 2009, 56, 101–143. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Seeram, N.P. Further investigation into maple syrup yields 3 new lignans, a new phenylpropanoid, and 26 other phytochemicals. J. Agric. Food Chem. 2011, 59, 7708–7716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valle, M.; St-Pierre, P.; Pilon, G.; Anhê, F.F.; Varin, T.; Marette, A. Effects of various natural sweeteners on insulin resistance, inflammation and liver steatosis in a rat model of diet-induced obesity. FASEB J. 2016, 30, lb650. [Google Scholar] [CrossRef]
- Marette, A. Impact of Free Sugar Replacement by Maple Syrup on Prevention of Metabolic Disorders Associated With Overweight in Humans: Role of Gut Microbiota; ClinicalTrials.gov: Washington, DC, USA, 2019.
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligne, B.; Ganzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented Foods: Definitions and Characteristics, Impact on the Gut Microbiota and Effects on Gastrointestinal Health and Disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, V.; Ferrao, J.; Pimentel, L.; Pintado, M.; Fernandes, T. One Health, Fermented Foods, and Gut Microbiota. Foods 2018, 7, 195. [Google Scholar] [CrossRef] [Green Version]
- Pessione, E.; Cirrincione, S. Bioactive Molecules Released in Food by Lactic Acid Bacteria: Encrypted Peptides and Biogenic Amines. Front. Microbiol. 2016, 7, 876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boling, L.; Cuevas, D.A.; Grasis, J.A.; Kang, H.S.; Knowles, B.; Levi, K.; Maughan, H.; McNair, K.; Rojas, M.I.; Sanchez, S.E.; et al. Dietary prophage inducers and antimicrobials: Toward landscaping the human gut microbiome. Gut Microbes 2020, 11, 721–734. [Google Scholar] [CrossRef] [Green Version]
- Nettleton, J.E.; Cho, N.A.; Klancic, T.; Nicolucci, A.C.; Shearer, J.; Borgland, S.L.; Johnston, L.A.; Ramay, H.R.; Noye Tuplin, E.; Chleilat, F.; et al. Maternal low-dose aspartame and stevia consumption with an obesogenic diet alters metabolism, gut microbiota and mesolimbic reward system in rat dams and their offspring. Gut 2020, 69, 1807–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chilton, S.N.; Burton, J.P.; Reid, G. Inclusion of fermented foods in food guides around the world. Nutrients 2015, 7, 390–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibberd, M.C.; Wu, M.; Rodionov, D.A.; Li, X.; Cheng, J.; Griffin, N.W.; Barratt, M.J.; Giannone, R.J.; Hettich, R.L.; Osterman, A.L.; et al. The effects of micronutrient deficiencies on bacterial species from the human gut microbiota. Sci. Transl. Med. 2017, 9, eaal4069. [Google Scholar] [CrossRef] [Green Version]
- Biesalski, H.K. Nutrition meets the microbiome: Micronutrients and the microbiota. Ann. N. Y. Acad. Sci. 2016, 1372, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Jaeggi, T.; Kortman, G.A.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef]
- Juste Contin Gomes, M.; Stampini Duarte Martino, H.; Tako, E. Effects of Iron and Zinc Biofortified Foods on Gut Microbiota In Vivo (Gallus gallus): A Systematic Review. Nutrients 2021, 13, 189. [Google Scholar] [CrossRef] [PubMed]
- Vanhaecke, T.; Bretin, O.; Poirel, M.; Tap, J. Drinking Water Source and Intake Are Associated with Distinct Gut Microbiota Signatures in US and UK Populations. J. Nutr. 2022, 152, 171–182. [Google Scholar] [CrossRef]
- Dai, Z.; Sevillano-Rivera, M.C.; Calus, S.T.; Bautista-de Los Santos, Q.M.; Eren, A.M.; van der Wielen, P.; Ijaz, U.Z.; Pinto, A.J. Disinfection exhibits systematic impacts on the drinking water microbiome. Microbiome 2020, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Leung, C.W.; Fung, T.T.; McEvoy, C.T.; Lin, J.; Epel, E.S. Diet Quality Indices and Leukocyte Telomere Length Among Healthy US Adults: Data From the National Health and Nutrition Examination Survey, 1999–2002. Am. J. Epidemiol. 2018, 187, 2192–2201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous-Bou, M.; Molinuevo, J.L.; Sala-Vila, A. Plant-Rich Dietary Patterns, Plant Foods and Nutrients, and Telomere Length. Adv. Nutr. 2019, 10, S296–S303. [Google Scholar] [CrossRef]
- Cassidy, A.; De Vivo, I.; Liu, Y.; Han, J.; Prescott, J.; Hunter, D.J.; Rimm, E.B. Associations between diet, lifestyle factors, and telomere length in women. Am. J. Clin. Nutr. 2010, 91, 1273–1280. [Google Scholar] [CrossRef] [Green Version]
- Franco-de-Moraes, A.C.; de Almeida-Pititto, B.; da Rocha Fernandes, G.; Gomes, E.P.; da Costa Pereira, A.; Ferreira, S.R.G. Worse inflammatory profile in omnivores than in vegetarians associates with the gut microbiota composition. Diabetol. Metab. Syndr. 2017, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Swanson, K.S.; de Vos, W.M.; Martens, E.C.; Gilbert, J.A.; Menon, R.S.; Soto-Vaca, A.; Hautvast, J.; Meyer, P.D.; Borewicz, K.; Vaughan, E.E.; et al. Effect of fructans, prebiotics and fibres on the human gut microbiome assessed by 16S rRNA-based approaches: A review. Benef. Microbes 2020, 11, 101–129. [Google Scholar] [CrossRef] [PubMed]
- Tucker, L.A. Milk Fat Intake and Telomere Length in U.S. Women and Men: The Role of the Milk Fat Fraction. Oxidative Med. Cell Longev. 2019, 2019, 1574021. [Google Scholar] [CrossRef]
- Barone, M.; Turroni, S.; Rampelli, S.; Soverini, M.; D’Amico, F.; Biagi, E.; Brigidi, P.; Troiani, E.; Candela, M. Gut microbiome response to a modern Paleolithic diet in a Western lifestyle context. PLoS ONE 2019, 14, e0220619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Palma, G.; Nadal, I.; Collado, M.C.; Sanz, Y. Effects of a gluten-free diet on gut microbiota and immune function in healthy adult human subjects. Br. J. Nutr. 2009, 102, 1154–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz, Y. Effects of a gluten-free diet on gut microbiota and immune function in healthy adult humans. Gut Microbes 2010, 1, 135–137. [Google Scholar] [CrossRef] [Green Version]
- Muralidharan, J.; Moreno-Indias, I.; Bullo, M.; Lopez, J.V.; Corella, D.; Castaner, O.; Vidal, J.; Atzeni, A.; Fernandez-Garcia, J.C.; Torres-Collado, L.; et al. Effect on gut microbiota of a 1-y lifestyle intervention with Mediterranean diet compared with energy-reduced Mediterranean diet and physical activity promotion: PREDIMED-Plus Study. Am. J. Clin. Nutr. 2021, 114, 1148–1158. [Google Scholar] [CrossRef] [PubMed]
- Roman, G.C.; Jackson, R.E.; Gadhia, R.; Roman, A.N.; Reis, J. Mediterranean diet: The role of long-chain omega-3 fatty acids in fish; polyphenols in fruits, vegetables, cereals, coffee, tea, cacao and wine; probiotics and vitamins in prevention of stroke, age-related cognitive decline, and Alzheimer disease. Rev. Neurol. 2019, 175, 724–741. [Google Scholar] [CrossRef] [PubMed]
- Schmedes, M.; Brejnrod, A.D.; Aadland, E.K.; Kiilerich, P.; Kristiansen, K.; Jacques, H.; Lavigne, C.; Graff, I.E.; Eng, O.; Holthe, A.; et al. The Effect of Lean-Seafood and Non-Seafood Diets on Fecal Metabolites and Gut Microbiome: Results from a Randomized Crossover Intervention Study. Mol. Nutr. Food Res. 2019, 63, e1700976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, R.; Merz, B.; Rist, M.J.; Ferrario, P.G.; Bub, A.; Kulling, S.E.; Watzl, B. Associations of current diet with plasma and urine TMAO in the KarMeN study: Direct and indirect contributions. Mol. Nutr. Food Res. 2017, 61, 1700363. [Google Scholar] [CrossRef] [PubMed]
- Mohr, A.E.; Jager, R.; Carpenter, K.C.; Kerksick, C.M.; Purpura, M.; Townsend, J.R.; West, N.P.; Black, K.; Gleeson, M.; Pyne, D.B.; et al. The athletic gut microbiota. J. Int. Soc. Sports Nutr. 2020, 17, 24. [Google Scholar] [CrossRef]
- O’Donovan, C.M.; Madigan, S.M.; Garcia-Perez, I.; Rankin, A.; O’Sullivan, O.; Cotter, P.D. Distinct microbiome composition and metabolome exists across subgroups of elite Irish athletes. J. Sci. Med. Sport 2020, 23, 63–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, L.M.; Bautista, E.J.; Nguyen, H.; Hanson, B.M.; Chen, L.; Lek, S.H.; Sodergren, E.; Weinstock, G.M. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome 2017, 5, 98. [Google Scholar] [CrossRef] [PubMed]
- Buan, N.R. Methanogens: Pushing the boundaries of biology. Emerg. Top Life Sci. 2018, 2, 629–646. [Google Scholar] [CrossRef] [Green Version]
- Estaki, M.; Pither, J.; Baumeister, P.; Little, J.P.; Gill, S.K.; Ghosh, S.; Ahmadi-Vand, Z.; Marsden, K.R.; Gibson, D.L. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome 2016, 4, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisseau, N.; Barnich, N.; Koechlin-Ramonatxo, C. The Nutrition-Microbiota-Physical Activity Triad: An Inspiring New Concept for Health and Sports Performance. Nutrients 2022, 14, 924. [Google Scholar] [CrossRef]
- Nieman, D.C.; Wentz, L.M. The compelling link between physical activity and the body’s defense system. J. Sport Health Sci. 2019, 8, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Bermon, S.; Petriz, B.; Kajeniene, A.; Prestes, J.; Castell, L.; Franco, O.L. The microbiota: An exercise immunology perspective. Exerc. Immunol. Rev. 2015, 21, 70–79. [Google Scholar]
- Gonze, D.; Coyte, K.Z.; Lahti, L.; Faust, K. Microbial communities as dynamical systems. Curr. Opin. Microbiol. 2018, 44, 41–49. [Google Scholar] [CrossRef]
- Stegen, J.C.; Bottos, E.M.; Jansson, J.K. A unified conceptual framework for prediction and control of microbiomes. Curr. Opin. Microbiol. 2018, 44, 20–27. [Google Scholar] [CrossRef]
- Yang, W.; Liu, Y.; Yang, G.; Meng, B.; Yi, Z.; Yang, G.; Chen, M.; Hou, P.; Wang, H.; Xu, X. Moderate-Intensity Physical Exercise Affects the Exercise Performance and Gut Microbiota of Mice. Front. Cell Infect. Microbiol. 2021, 11, 712381. [Google Scholar] [CrossRef]
- Diaz, J.; Reese, A.T. Possibilities and limits for using the gut microbiome to improve captive animal health. Anim. Microbiome 2021, 3, 89. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Health and Human Services. Physical Activity Guidelines for Americans, 2nd ed.; U.S. Department of Health and Human Services: Washington, DC, USA, 2018.
- U.S. Department of Agriculture; U.S. Department of Health and Human Services. Dietary Guidelines for Americans, 2020–2025, 9th ed.; U.S. Department of Agriculture: Washington, DC, USA, 2020.
Figure 1.
PRISMA-ScR flow diagram.


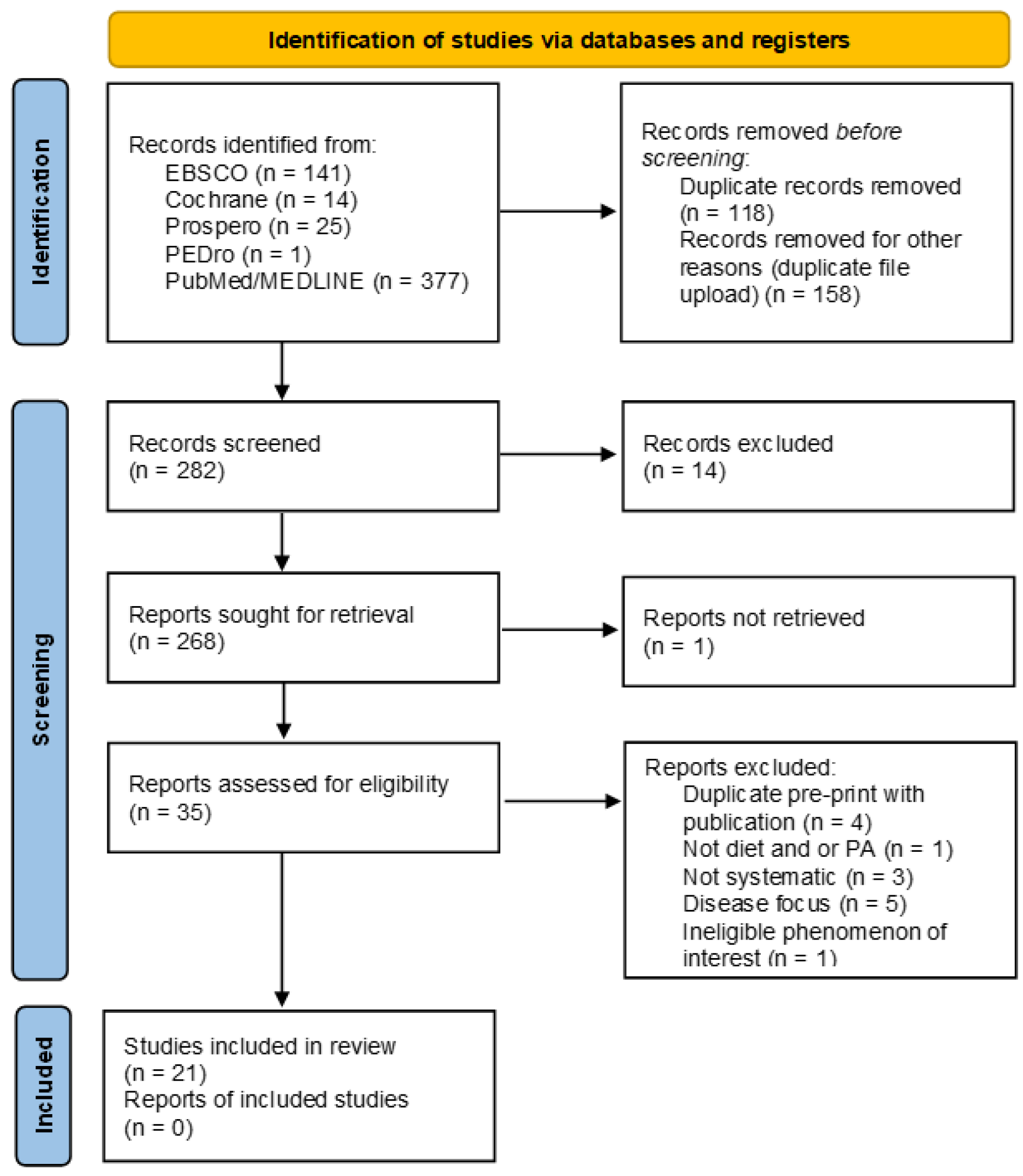{kind=link}
Table 1.
PICOS criteria for inclusion.
| PICOS Format | Description |
|---|---|
| Population | Healthy subjects Adult humans aged 18 years or older |
| Intervention or exposure | Diet (Western-style, plant-based, vegan); and/or Physical activity or exercise |
| Comparisons | Diet (omnivore, Western-type, vegetarians, vegans) and/or Physical Activity level; reviews of interventions examining probiotics solely and non-interventional reviews were excluded |
| Outcome | Gut microbiota composition through fecal samples; abundance composition or abundance of specific intestinal bacteria |
| Study design | Systematic reviews of cross-sectional, prospective cohort studies, randomized-controlled trials of either parallel or crossover design; and reviews or studies for background information on food or physical activity |
Table 2.
Summary of systematic reviews and metanalyses on the effect of diet or exercise on the microbiome.
Table 2.
Summary of systematic reviews and metanalyses on the effect of diet or exercise on the microbiome.
| Aim and Design of Studies | Number of Studies | Quality | Effect on Microbiome | First Author/Study Name |
|---|---|---|---|---|
| Influence of a healthy diet pattern on the microbiome and inflammatory markers Interventional human trials | 18 | Critical appraisal not reported | Due to heterogeneity in study design and type of subjects, no conclusions could be made | Telle-Hansen (2018) [17] |
| Vegan and vegetarian diet association with gut microbiota composition Cross-sectional/cohort/RCT | 37 | a Newcastle–Ottawa scale NOS: 4.6 out of 10 points | No consistent association between a vegan or vegetarian diet and microbiota composition | Trefflich (2019) [18] |
| Effect of wine and grape polyphenols on human gut microbiota. Meta-analysis RCTs | 7 | b Cochrane Risk of Bias Low (5) (1) unclear (1) high risk | Increased Proteobacteria, Fusobacteria, Firmicutes, Bacteroidetes, and B. uniformis after red wine intake; Decrease in dysbiosis-associated species: Clostridum, Eubacterium, and Bacteroides | Nash (2018) [19] |
| Dietary fiber intervention on microbiome Meta-analysis of RCTs | 64-reviewed 58-retained for meta-analysis 12 studies focused on whole-grain diet versus a low-fiber diet | b Cochrane Risk of Bias Low to moderate risk of bias (n = 64) | Dietary fiber intervention compared to placebo/low fiber diet did not significantly increase α-diversity, but increased abundance of Bifidobacterium spp. No difference in Lactobacillus spp. Abundance with food intervention, but significant in fiber supplement group | So (2018) [20] |
| Dietary fat and gut microbiota Cross-sectional, cohort; interventional studies and randomized controlled trials | 16 | b Cochrane Risk of Bias 14 RCT-low risk; 2 RCT-high-risk a Newcastle-Ottawa NOS: 3 = 8, 3 = 7, 1 + 6, and 2 = 5 | n3, n6 PUFA increase beneficial bacteria; high fat/saturated fat diets reduced richness and diversity and had negative metabolic health outcomes; observational studies show an association between fat and health outcomes | Wolters (2019) [21] |
| Association between exercise and gut microbial composition in mammals RCT, cross-sectional, and cohort studies | Human—20 Animal—5 | b Cochrane Risk of Bias–unclear Low Quality Lack of appraisal tools for heterogeneous models | Exercise was associated with changes in gut microbial composition, an increase in butyrate-producing bacteria, and fecal butyrate | Mitchell (2019) [22] |
| Effects of intact cereal grain fibers on microbiome RCT, RCT crossover, non-randomized | 40 | Critical appraisal not provided | Cereal fiber (6–8 g) increases diversity and abundance; increase in bacterial metabolites | Jefferson (2019) [23] |
| Influence of exercise on the human gut microbiota in healthy adults Observational and case-control | 18 | d PEDro 18—Medium | 4/9 observational studies showed higher levels of physical activity or cardiorespiratory fitness were positively associated with α-diversity | Ortez-Alvarez (2020) [24] |
| Influence of endurance training intervention and gut microbiome Interventional studies > 4 weeks duration | 5 | d PEDro 4 studies score ≥ 4—Fair quality 1 study was rated Poor | PA significantly lowers abundance of Bacteroidetes and increases Firmicutes and β diversity in some studies | Shahar (2020) [25] |
| Effects of dairy and dairy derivatives on the gut microbiota (Bovine, yogurt, soy) | 8 | b Cochrane risk-of-bias 2—low 5—some concerns 1—high risk | Richness and diversity declined in all types of milk, Lactobacillus increased in bovine milk; fermented yogurt and kefir increased Lactobacillus and Bifidobacterium | Aslam (2020) [26] |
| Effect of nut consumption on gut microbiome and gut function RCTs | 8 | b Cochrane risk-of-bias No studies were low risk of bias; variable across categories of analysis | Meta-analysis found no effect on β-diversity; no effect of nut type, dose, duration of intervention; increased abundances of Clostridium, Lachnospira and Roseburia | Creedon (2020) [27] |
| Effect of dietary pulses on microbial populations RCT-C, (cross-over); Interventions with control or placebo group | 5 | Critical appraisal not provided | Bacteroides fragilis OUT↓ i for navy bean pulse flour; No difference in Shannon index for diet with chickpeas; lupin fiber consumption decreased abundance of Bacteroides-Prevotella | Marinangeli (2020) [28] |
| Effect of nut consumption on gut microbiome RCT-C, (cross-over); RCT parallel design, and pre/post-test studies | 8 | f Quality Criteria Checklist and g Risk of Bias Assessment Tool 6/8 positive quality 2 neutral | Nuts in general, but especially walnuts, had an impact on gut microbial composition | Fitzgerald (2021) [29] |
| Association between physical activity and changes in intestinal microbiota composition Cross-sectional and longitudinal studies | 17 | h ROBINS-I 6—low 7—moderate 1—serious 3—not reported | Increase in SCFAs concentration after the training period in lean athletes only; composition and diversity differ by sport | Aya (2021) [30] |
| Physical activity influences on human gut microbiota independent of diet observational | 10 4/10 studies controlled for diet | e JBI Critical Appraisal Checklist-criteria met b Cochrane Risk of Bias −2/20 some concerns | Variability is affected by dietary factors and physical characteristics; use of high protein diets contributes to greater variability among athletes | Dorelli (2021) [31] |
| Dietary habits and gut microbiota in healthy adults cross-sectional and RCT Diet regimen studies | 16 | a Newcastle-Ottawa scale Mean score for cross-sectional studies was 5/10; b Cochrane Collaboration tool risk of bias-Low | Significant impact on some bacterial genera from a rich and varied omnivore diet, such as Mediterranean | Gibiino (2021) [32] |
| Vegan diet and gut microbiota Cross-sectional studies | 9 | a Newcastle-Ottawa scale Most studies scored “medium” quality | Firmicutes/Bacteroidetes ratio is lower in vegans compared to omnivores; Abundance of Bacteriodetes and Prevotella in vegans | Losno (2021) [33] |
| a- and β-diversity in obese and non-obese adults Intervention studies and RCT | 32 22 reported Shannon Index (diversity) 25 studies investigated diversity; 2 did not; 5 did not stratify by BMI | h Adapted ROBINS-I Serious risk in one domain: 22 Moderate: 10 | Higher levels of PA and cardiorespiratory fitness are associated with greater α-diversity and increases in some phyla and certain short-chain fatty acids | Pinart (2021) [34] |
| Effects of exercise and physical activity on the gut microbiome in older adults Observational and interventional studies | 7 | Critical appraisal not provided | PA had beneficial impact on the gut microbial composition of older adults | Ramos (2022) [35] |
| Physical activity and human gut microbiota in healthy and unhealthy subjects Observational and interventional studies | 25 | h ROBINS-I 8 studies scored 4–5 c Jadad Scale 4/5 studies: Moderate e JBI Critical Appraisal Checklist for Analytical Cross-Sectional Studies 12/12 Included | No significant change in richness and diversity in gut microbiota for minimum PA recommendations Microbial diversity is associated with aerobic exercise | Cataldi (2022) [36] |
| Effect of MedDiet on microbiota and metabolites RCT and Observational studies | 34 17-RCT 17-Observ | b Cochrane (RCT) Mixed Quality a Newcastle-Ottawa scale (Observational) Prospective Studies: High 2 Moderate 1 Low 2 Cross-sectional High 6 Moderate 6 Low 2 | Overall positive impact of Mediterranean diet on Firmicutes/Bacteroidetes ratio. but effects are not consistent between studies due to adherence differences and fewer species that utilize oligosaccharides and simple sugars | Kimble (2022) [37] |
a Newcastle–Ottawa scale (NOS) [38]; b Cochrane risk of bias [39]; c Jadad Scale or Oxford Quality Scoring System Scale [40]; d PEDro scale (Maher, 2003); e JBI Critical Appraisal Checklist for Analytical Cross-Sectional Studies [41]; f Quality Criteria Checklist; g Risk of Bias Assessment Tool [42], h ROBINS-I [43] and i ↓ indicates a decrease.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Grace-Farfaglia, P.; Frazier, H.; Iversen, M.D. Essential Factors for a Healthy Microbiome: A Scoping Review. Int. J. Environ. Res. Public Health 2022, 19, 8361. https://doi.org/10.3390/ijerph19148361
AMA Style
Grace-Farfaglia P, Frazier H, Iversen MD. Essential Factors for a Healthy Microbiome: A Scoping Review. International Journal of Environmental Research and Public Health. 2022; 19(14):8361. https://doi.org/10.3390/ijerph19148361
Chicago/Turabian StyleGrace-Farfaglia, Patricia, Heather Frazier, and Maura Daly Iversen. 2022. "Essential Factors for a Healthy Microbiome: A Scoping Review" International Journal of Environmental Research and Public Health 19, no. 14: 8361. https://doi.org/10.3390/ijerph19148361
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.