1. Introduction
Among the early environmental factors, parent–infant interaction modulates human development on a biological level, influencing social, emotional, and cognitive domains [
1]. Efficient parenting promotes an inclusive environment, fostering patterns for high-quality social relationships in childhood [
2,
3] which, in turn, can decrease behavioral and physiological distress in adulthood [
4]. Simultaneously, different long-term environmental factors can play an active role in the modulation of sociality across the human lifespan. For example, children are increasingly exposed to technological devices from the first years of their lives [
5] and begin to use mobile technologies before school age [
6]. This exposition may contribute to social competencies learning in online and offline worlds [
7]. As age limits can be easily deceived, younger and younger individuals can access social network sites (SNSs), where social exchanges between users are immediate, free, continuous, and pervasive [
8]. Among these sites, Instagram (IG) is a photo- and video-sharing platform where users can follow stories, publish or comment on posts, watch or upload content, and chat privately. Some evidence has demonstrated that increased Instagram activity might be associated with positive outcomes, acting as a motivator for a satisfying self-presentation and increasing self-confidence [
9]. However, it is also associated with negative consequences, such as a risk of developing of a depressed mood [
10] and a detrimental effect on mental well-being [
11].
The influence of environmental cues might not fully explain such a tangled phenomenon as social behavior. According to the biopsychosocial model of wellness and illness, it is more likely that genetic predispositions could be crucial in shaping the effect of early experiences on social conduct on SNSs. At the molecular level, individual behaviors result from genetic predispositions and environmental stressors or conditions, especially during critical periods [
12]. According to the
differential susceptibility model [
13], a given genetic predisposition is not uniquely “good” or “bad” in human behavior. Indeed, the level of susceptibility to life events is flexible and regulated by the allelic expression involved in the physiological and behavioral responses to a range of triggering environmental hazards. In the myriad of biological regulators, serotonin is a neurotransmitter implicated in the neural circuits of emotional regulation, social functioning, and social affiliation [
14,
15]. Serotonin levels may vary across brain regions and affect individuals differently [
16]. The serotonin transporter encoded by the serotonin transporter gene (SLC6A4) is the primary regulator of serotonin removal from the synaptic clefts. Specifically, the variable number of tandem repeats (VNTR), known as the promoter region of the serotonin transporter gene-linked polymorphic region (5-HTTLPR), and the A/G single-nucleotide polymorphism (SNP) located within the 5-HTTLPR repeats (rs25531) [
17,
18] have been investigated in association with perceived attachment and parental attitudes [
19,
20,
21,
22]. Concerning 5-HTTLPR, for instance, evidence from studies on animal models suggests that variations in the polymorphic region of this gene confer greater susceptibility to environmental factors. For instance, rhesus macaques carrying the short-allele variation benefited from a supportive and caring environment during early interactions, showing greater social competence later in life [
23]. As for human models, Truzzi et al. [
24] discovered that men with a higher predisposition (carriers of the short form of 5-HTTLPR) for sensitivity to maternal overprotection showed a distressed heart rate to female cry. Concerning rs25531, two allelic forms have been found: the substitution of thymine (T) and cytosine (C), with the paired nucleotides guanine (G) and adenine (A), shows differential susceptibility to stressors [
25,
26,
27,
28]. However, until now, literature has not been able to disentangle which variation is more associated with adaptive social responses [
29,
30]. Genetic vulnerability conferred by the A/A and T/T genotype, whose transcription processes are associated with a lower reuptake of serotonin, can be moderated by the social support of parents [
31]. Social support in childhood influences the risk of mental disorders in adulthood and facilitates a wide range of possible behavioral outcomes [
26,
28,
32,
33,
34]. For instance, people with a history of low-quality parental support in childhood could show maladaptive behaviors and benefit from others’ support in adulthood [
35,
36,
37,
38]. Conversely, individuals with the C or G allele showed enhanced serotonin degrees combined with lessened sensitivity to pain, traumatic events, and care [
27]. Within the social context, higher social avoidance and lower emotional engagement were related to a decrease in serotonin, whereas higher prosocial behavior and lower social anxiety from childhood were related to an increase in serotonin [
20,
26]. A piece of experimental research highlighted a positive association between avoidance towards peers and neural activity of the anterior prefrontal cortex in response to distress stimuli for C-carriers but not for T/T homozygotes [
39].
In summary, we could assert that sociability emerges from the combined effects of nature, driven by genetic and biological predispositions and nurture (e.g., social environment, social education across development, and social experiences with parents and peers) [
12]. However, the function of the “sociability genes” (i.e., rs25531) according to the early environment’s quality on the online social interaction of adults has not yet been investigated.
This research explores how the interaction between the serotonin transporter gene and the individual’s relationship with caregivers during childhood modulates adults’ online social interactions on Instagram. Our attention is focused on Instagram, one of the most popular social networks, to unveil online social interaction’s fundamental mechanisms. Instagram users were firstly required to fill the Parental Bonding Instrument to measure their recalled parental bonding. Buccal mucosa samples were subsequently collected, and one 5-HTT single-nucleotide polymorphism (SNP: rs25531) was examined as a genetic factor. Finally, three social media indexes were assessed from the Instagram profile of each user: the number of (i) published posts as an index of the social productivity of the network, (ii) number of followed users (here called “followings”) as a measure of prosocial activity, (iii) number of followers as a measure of the social attraction of other users. A fourth Instagram index was obtained as a proxy of asymmetry of the social network, calculated through the ratio between the number of followers and the number of followings: (iv) the “Social Desirability Index” (SDI).
For each Instagram index as the dependent variable, we hypothesized an interaction effect between the genetic component and the attachment scores, independent of the gender. More precisely, we expected that adult Instagram users with a genetic risk factor (rs25531 T/T) who experienced a supportive and positive past relationship with parents (High Parental Care, Low Parental Overprotection) would show a greater online social activity compared to protective genetic carriers (rs25531 C-carriers). Although we formulated a directional hypothesis, we could not predict a priori on which Instagram variable potential effects would have been displayed. Given the new role of Instagram activity from a gene–environment perspective, we adopted an exploratory approach.
4. Discussion
In this study, we probed how alleles in rs25531 interact with recalled parental care and overprotection in the modulation of social behavior on Instagram. Specifically, we hypothesized one genotype*environment interaction (rs25531 SNP*parental bonding in childhood) on Instagram variables.
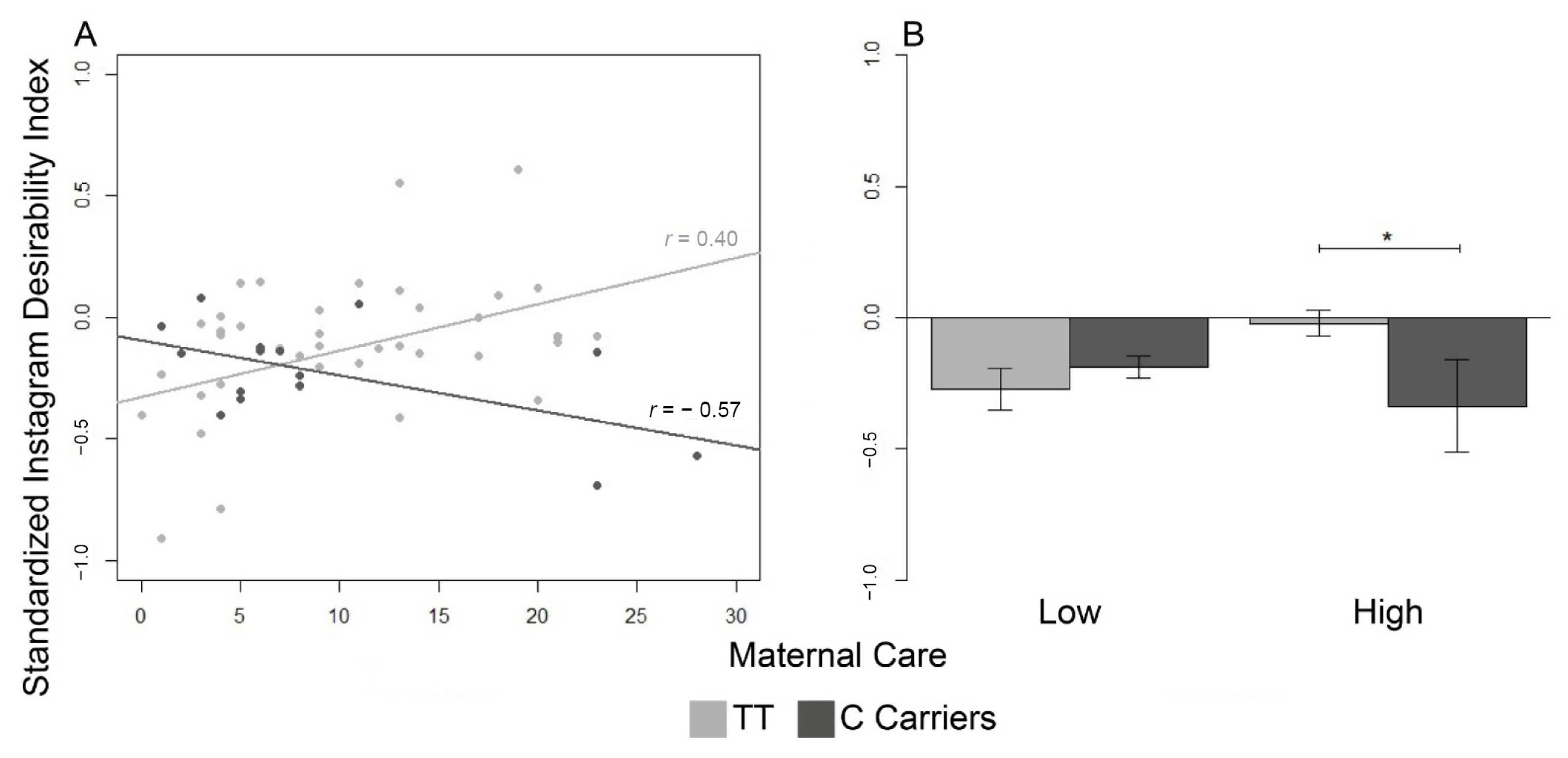
In accordance with the hypothesis, we discovered that adult Instagram users with genetic vulnerability (T/T homozygotes) showed varying Instagram social conduct related to the quality of the parental practices experienced in childhood. As expected, T/T homozygotes who reported high maternal care scores exhibited an increasing trend in the Instagram SDI. In contrast, those who reported low scores in the equivalent dimension determined a decreasing trend in the same index. Interestingly, C-carriers showed the opposite pattern: as maternal care scores were lower, Instagram SDI increased. In particular, vulnerable genetic carriers (T/T homozygotes) with a positive relationship with the mother displayed a higher Instagram SDI than protective genetic carriers (C-carriers).
This study investigated the
differential susceptibility model on rs25531 within a novel and innovative framework: the online social behavior on Instagram—a SNS. In line with the model [
13], a T versus C allele’s presence implicated a conditional variation in the online social response: T/T homozygotes who benefited from efficacious maternal care displayed a higher Instagram SDI than C-carriers. In this condition, Instagram users carrying the C allele were less susceptible than T/T homozygotes to supportive maternal care during childhood. Compared with T/T genotype, the C allele conferred lower vulnerability to adverse conditions linked to neglectful caregiving and less advantage from a favorable relationship with the mother. Consequently, users with the C allele exhibited an Instagram social response unconcerned about the advantages promoted by appropriate maternal care [
27,
58]. Otherwise, users with the T/T genotype took advantage of a protective and comfortable maternal relationship, exhibiting higher asymmetry between the number of followers and followings than less-sensitive ones [
59]. As a result of developmental flexibility, the T allele predisposes individuals to be more vulnerable or plastic according to the functional or dysfunctional attachment with the mother [
60,
61,
62].
The present research extends previous evidence of genetic influence on the frequency of online social behavior [
63,
64,
65]. Regarding rs25531, a previous study found that carriers of susceptibility genetic factors (T/T homozygotes) presented a higher Instagram number of followings than nonsensitive ones (C-carriers) when they reported a high level of confidence toward others [
66]. As pointed out by another work [
44], in the current study, users genetically susceptible to environmental influences displayed higher SDI when they had previously experienced positive maternal bonding. However, recent work on rs25531 disclosed an association between avoidance in adult attachment and Instagram number of followings independent of genetic predispositions [
67].
4.1. Possible Implications in Brain Mechanisms
The increased vulnerability conferred by rs25531 could influence the regulation of 5-HTTLPR expression and, in turn, processes mediated by serotonin and serotonergic pathways, such as sensory processing of socially relevant stimuli and emotional decision making [
68]. Serotonin neurons are particularly dense in the prefrontal cortex (PFC), suggesting that it acts as a major modulator of the functions carried out by this area [
69]. The PFC is closely linked to the amygdala and is involved in processing information related to social perception and experience. A targeted region of the PFC, namely, the orbitofrontal cortex (OFC), is involved in operations requiring social flexibility, such as evaluating rewards and risks when responding to a social situation and processes of salient social cues [
70,
71]. In fact, OFC integrates information regarding environmental context, emotions, and memory [
72]. OFC is also shown to be involved in mental activity aimed at suppressing negative thoughts, which is strictly linked to the perception of parental care received at the early stages of life [
73]. Other than the PFC, serotonin is involved in the circuits of the nucleus accumbens (NAc) related to social rewards [
74] and neural mechanisms of the medial preoptic area (MPOA) of the hypothalamus [
75] that support maternal behavior [
76]. As such, serotonin-related regulations appear to be widely involved in planning emotional and behavioral responses to environmental social stimuli [
39,
72,
77], especially when the social incentives are mediated by the expectations of the self and of others [
28].
4.2. The Importance of Social Desirability
We focused our attention on the SDI because it represents the distribution of contacts for each user on Instagram. This index attributes a size to each user’s network structure, where the number of followers increases at the cost of the number of followings. Maximizing the SDI means enlarging the network structure to achieve greater approval and benefit from the masses [
78,
79]. Within this frame, we argue that higher maternal care could determine the basis of new prosocial behavior in the offline environment, later combined with a greater need for social desirability in the online environment [
9,
44,
80]. An Instagram user sensitive to early maternal experiences could, in turn, respond more efficiently to the request of others and support social exchanges in response to their followers’ expectations [
55,
81,
82]. This outcome suggests that the perception of maternal warmth could favor or reduce needs and abilities related to self-management [
83], influencing the capacity to correspond to one’s followers’ expectations [
84]. SDI resents the user’s online activity in terms of frequency, minutes, or hours of usage per day, and time passed since the subscription to the platform; these parameters have not been investigated in the present study, but future research could elucidate the contribution of time-related variables to the SDI. In fact, SDI presents the potential for a solid link to the posting or usage activity since they might reinforce one another. Additionally, if the user is not competent or familiar with the usage of Instagram, the profile indexes might display lower numbers of following or followers. In comparison, more expert users could interact more efficiently, reaching more significant numbers of following and attracting more users’ requests for connection. A limit of the SDI is that numbers of following and followers can be easily manipulated by the profile owner, for instance, by deleting following accounts to maximize the impact of the number of followers.
6. Conclusions
This research is not exempt from some limitations. Firstly, the low sample size was constrained by data collection of multiple variables in a fixed period. Data collection was circumscribed to young adults between 18 and 25 years old from the same cultural context. Even though the 1000 genomes project found no significant differences in the allele variability of rs25531 (T > C) between Asian populations (1000 Genomes project, BioSample: SAMN07486024, dbSNP (Short Genetic Variations), 2017), an unbalanced sample composition should be noted (e.g.,
N = 56 Chinese and 1 Indian). At a cultural level, Singapore is a multiethnic country, where citizens of different ethnic backgrounds (e.g., Chinese, Indian, Malay) share a sense of belonging to the community [
88]. Moreover, an unbalanced rate in the ethnicity of Singaporean participants may reflect the demographic distribution of the country since Chinese citizens represent over 75% of Singapore’s population [
89].
Overall, future studies could overcome these potential limitations. For instance, collecting data from a cross-cultural sample (e.g., Western- vs. Eastern-oriented) could allow for inspecting differences in the genetic predispositions, caregiving propensities, and Instagram behaviors. Instagram responses could also be explored to prove that online communication frequency and duration could vary between age ranges (e.g., adolescence, early adulthood, adulthood, old age).
The participants’ socioeconomic status (SES) was also not evaluated. Users who live in a situation of economic security could increase their Instagram activity to prove their ideal prosperity to their followers [
90]. The year of first registration on Instagram was also not collected; thus, the indexes’ quantities were not controlled for the duration of online activity. Furthermore, the analysis was performed on no more than three Instagram parameters and one combined index that did not discriminate “bot accounts” or “artificial” followers that can be purchased instead of being earned through social capital [
91,
92]. Future investigations should discriminate and analyze data only from real followed members and no other reel, advertisement, paid promotions, or other content suggested automatically by the social media algorithm. Finally, the recalled parental bonding was measured with a self-reported instrument.
Future studies should consider a more extensive and multicultural sample to probe potential cross-cultural differences in Instagram behavior and a gender-balanced sample including participants of different ages. The assessment of plausible moderators of Instagram usage such as the SES and the sense of membership with peer groups may also be considered. The increase in the number of posts, followers, and followings should be traced in relation to the year of the first registration on Instagram. Novel Instagram variables and indexes able to detect the variety of the shared content may also be taken into account, such as Instagram number of stories and tagged posts or the time spent using Instagram actively (i.e., chatting, posting) and passively (i.e., viewing, scrolling). Additionally, a replication of the study on data extracted from different social media platforms, such as Twitter or TikTok, may be conducted to verify the generalizability of reported findings. Furthermore, to detect the long-term impact of early life, a longitudinal study to analyze the development of online social skills as an effect of genes and experiences is recommended. Moreover, physiological measures—such as electrocardiography, electromyography, skin conductance, or recordings of neural activity—from peripheral and central nervous systems could be adopted to discover potential differences between online behaviors and offline social responses [
93,
94]. Moreover, sentiment analysis techniques could be adopted to assess Instagram posts in relation to attachment patterns to predict emotional suffering or mental health conditions [
95,
96]. Finally, as rs25531 and 5-HTTLPR result in two independent loci for analysis [
18], this model could be implemented on 5-HTTLPR.
In conclusion, serotonin transporter gene polymorphisms (rs25531) and early caregiving behaviors play an interactive role in developing online social behavior in Instagram users. Combined with previous evidence, these results offer a contribution to identifying an online social marker in the interplay between genetic and environmental components.


 ,
, 

{kind=link}
