
Wolbachia Impacts Anaplasma Infection in Ixodes scapularis Tick Cells

 and
and 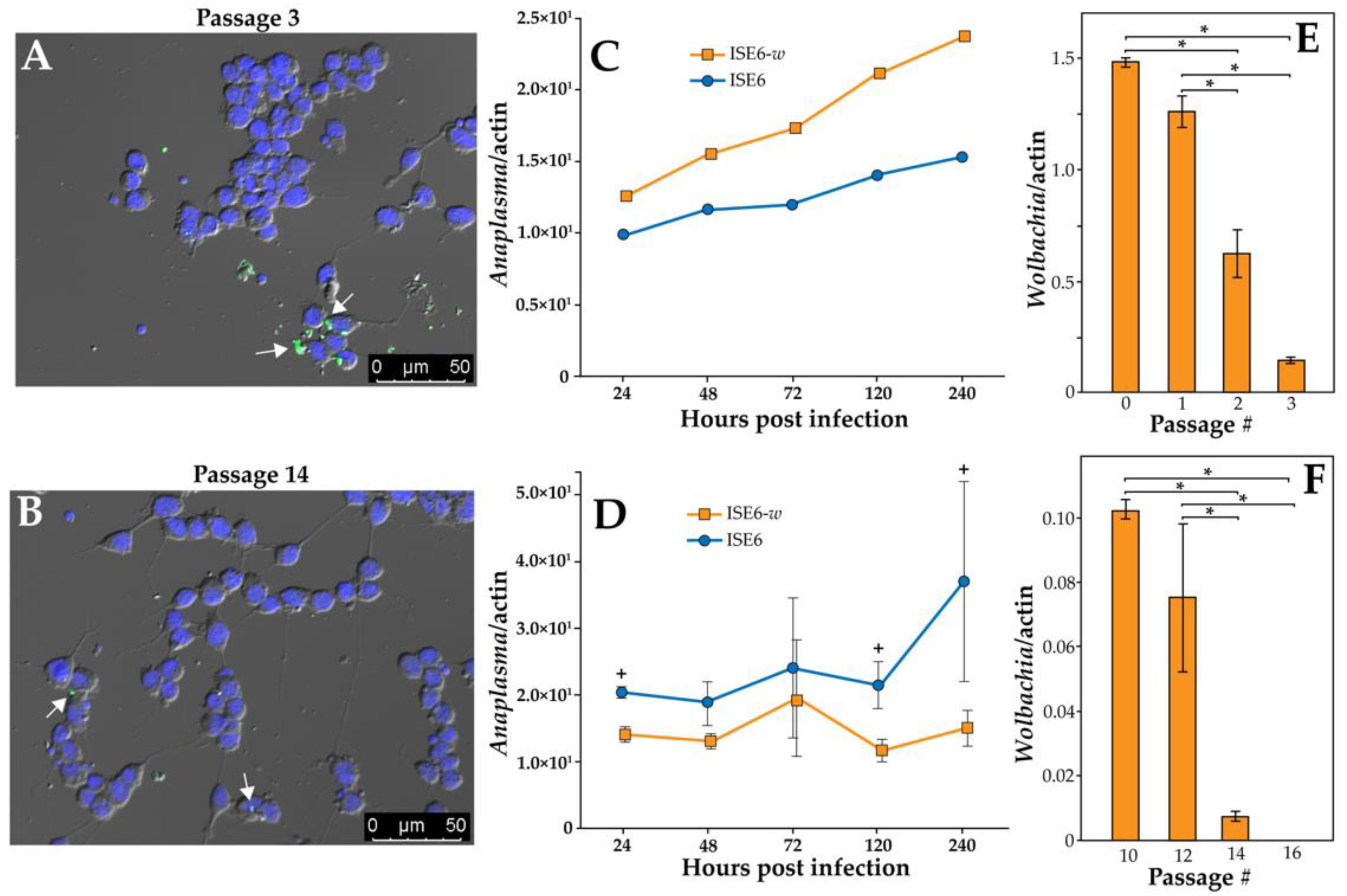{kind=link}
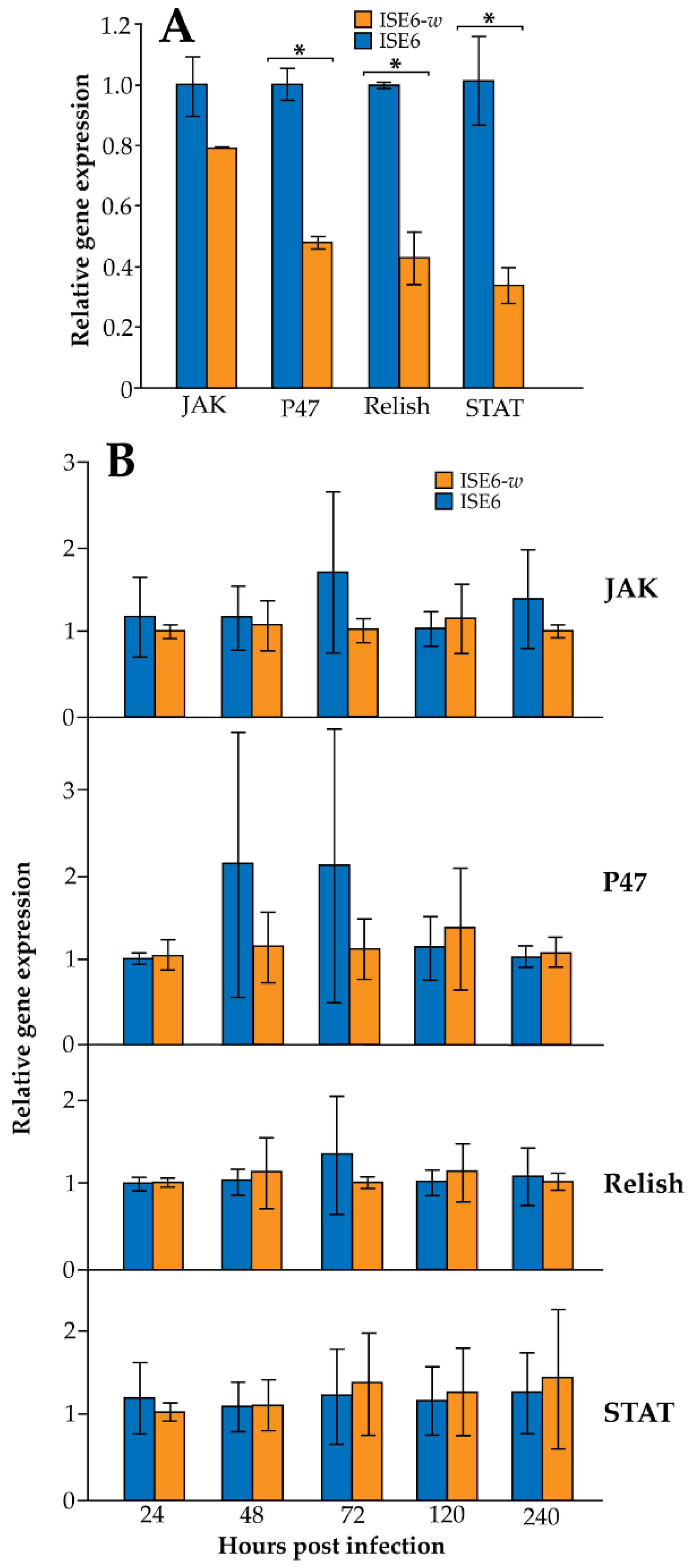{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. ISE6 Cell Transfection
2.2. Fluorescent In Situ Hybridization
2.3. Anaplasma phagocytophilum Propagation, ISE6 Cell Inoculation, and qPCR Quantification
2.4. Wolbachia Density
2.5. RNA Isolation and Immune Gene Expression
2.6. Statistical Analysis
3. Results
3.1. ISE6 Can Be Transfected with Wolbachia and Impact Anaplasma Transmission
3.2. Wolbachia Infection in ISE6 Cells Downregulates Immune Gene Expression but Is Dependent upon Wolbachia Density
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petersen, L.R.; Beard, C.B.; Visser, S.N. Combatting the Increasing Threat of Vector-Borne Disease in the United States with a National Vector-Borne Disease Prevention and Control System. Am. J. Trop. Med. Hyg. 2019, 100, 242–245. [Google Scholar] [CrossRef] [Green Version]
- Ahantarig, A.; Trinachartvanit, W.; Baimai, V.; Grubhoffer, L. Hard ticks and their bacterial endosymbionts (or would be pathogens). Folia Microbiol. 2013, 58, 419–428. [Google Scholar] [CrossRef]
- Budachetri, K.; Kumar, D.; Crispell, G.; Beck, C.; Dasch, G.; Karim, S. The tick endosymbiont Candidatus Midichloria mitochondrii and selenoproteins are essential for the growth of Rickettsia parkeri in the Gulf Coast tick vector. Microbiome 2018, 6, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, S.I.; Binetruy, F.; Hernández-Jarguín, A.M.; Duron, O. The Tick Microbiome: Why Non-pathogenic Microorganisms Matter in Tick Biology and Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7, 236. [Google Scholar] [CrossRef]
- Narasimhan, S.; Fikrig, E. Tick microbiome: The force within. Trends Parasitol. 2015, 31, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitsou, C.; Pal, U. Ixodes immune responses against Lyme disease pathogens. Front. Cell. Infect. Microbiol. 2018, 8, 176. [Google Scholar] [CrossRef]
- Smith, A.A.; Pal, U. Immunity-related genes in Ixodes scapularis--perspectives from genome information. Front Cell Infect. Microbiol. 2014, 4, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greay, T.L.; Gofton, A.W.; Paparini, A.; Ryan, U.M.; Oskam, C.L.; Irwin, P.J. Recent insights into the tick microbiome gained through next-generation sequencing. Parasites Vectors 2018, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Tsioka, K.; Kontana, A.; Papadopoulos, C.; Giadinis, N. Bacterial pathogens and endosymbionts in ticks. Ticks Tick-Borne Dis. 2017, 8, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Hartelt, K.; Oehme, R.; Frank, H.; Brockmann, S.O.; Hassler, D.; Kimmig, P. Pathogens and symbionts in ticks: Prevalence of Anaplasma phagocytophilum (Ehrlichia sp.), Wolbachia sp., Rickettsia sp., and Babesia sp. in Southern Germany. Int. J. Med. Microbiol. Suppl. 2004, 293, 86–92. [Google Scholar] [CrossRef]
- Benson, M.J.; Gawronski, J.D.; Eveleigh, D.E.; Benson, D.R. Intracellular symbionts and other bacteria associated with deer ticks (Ixodes scapularis) from Nantucket and Wellfleet, Cape Cod, Massachusetts. Appl. Environ. Microbiol. 2004, 70, 616–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoo, J.J.; Kurtti, T.J.; Husin, N.A.; Beliavskaia, A.; Lim, F.S.; Zulkifli, M.M.S.; Al-Khafaji, A.M.; Hartley, C.; Darby, A.C.; Hughes, G.L.; et al. Isolation and propagation of laboratory strains and a Novel Flea-Derived field strain of Wolbachia in tick cell lines. Microorganisms 2020, 8, 988. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Genet. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?--A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Rasgon, J. Population replacement strategies for controlling vector populations and the use of Wolbachia pipientis for genetic drive. J. Vis. Exp. 2007, e225. [Google Scholar] [CrossRef] [Green Version]
- Crain, P.R.; Mains, J.W.; Suh, E.; Huang, Y.; Crowley, P.H.; Dobson, S.L. Wolbachia infections that reduce immature insect survival: Predicted impacts on population replacement. BMC Evol. Biol. 2011, 11, 290. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, S.L. The use of Wolbachia by the world mosquito program to interrupt transmission of Aedes aegypti transmitted viruses. Adv. Exp. Med. Biol. 2018, 1062, 355–360. [Google Scholar]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.; McMeniman, C.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 2011, 476, 450–453. [Google Scholar] [CrossRef]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia Symbiont in Aedes aegypti Limits Infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 109, E23–E31. [Google Scholar] [CrossRef] [Green Version]
- Monsanto-Hearne, V.; Johnson, K.N. Wolbachia-mediated protection of Drosophila melanogaster against systemic infection with its natural viral pathogen Drosophila C virus does not involve changes in levels of highly abundant miRNAs. J. Gen. Virol. 2018, 99, 827–831. [Google Scholar] [CrossRef]
- Lu, P.; Bian, G.; Pan, X.; Xi, Z. Wolbachia induces density-dependent inhibition to dengue virus in mosquito cells. PLoS Negl. Trop. Dis. 2012, 6, e1754. [Google Scholar] [CrossRef] [Green Version]
- Frentiu, F.D.; Robinson, J.; Young, P.R.; McGraw, E.A.; O’Neill, S.L. Wolbachia-Mediated resistance to dengue virus infection and death at the cellular level. PLoS ONE 2010, 5, e13398. [Google Scholar] [CrossRef] [Green Version]
- Bian, G.; Joshi, D.; Dong, Y.; Lu, P.; Zhou, G.; Pan, X.; Xu, Y.; Dimopoulos, G.; Xi, Z. Wolbachia Invades Anopheles stephensi Populations and Induces Refractoriness to Plasmodium Infection. Science 2013, 340, 748–751. [Google Scholar] [CrossRef]
- Rasgon, J.L.; Gamston, C.E.; Ren, X. Survival of Wolbachia pipientis in Cell-Free Medium. Appl. Environ. Microbiol. 2006, 72, 6934–6937. [Google Scholar] [CrossRef] [Green Version]
- Munderloh, U.G.; Kurtti, T.J. Formulation of medium for tick cell culture. Exp. Appl. Acarol. 1989, 7, 219–229. [Google Scholar] [CrossRef]
- Dobson, S.L.; Marsland, E.J.; Veneti, Z.; Bourtzis, K.; O’Neill, S. Characterization of Wolbachia Host Cell Range via the In Vitro Establishment of Infections. Appl. Environ. Microbiol. 2002, 68, 656–660. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Jasperson, D.; Cohnstaedt, L.W.; Brelsfoard, C.L. Transfection of Culicoides sonorensis biting midge cell lines with Wolbachia pipientis. Parasites Vectors 2019, 12, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chávez, A.S.O.; Fairman, J.W.; Felsheim, R.F.; Nelson, C.M.; Herron, M.J.; Higgins, L.; Burkhardt, N.Y.; Oliver, J.D.; Markowski, T.W.; Kurtti, T.J.; et al. An O-Methyltransferase is required for infection of tick cells by Anaplasma Phagocytophilum. PLoS Pathog. 2015, 11, e1005248. [Google Scholar] [CrossRef] [Green Version]
- Carroll, E.E.M.; Wang, X.; Shaw, D.K.; O’Neal, A.J.; Chávez, A.S.O.; Brown, L.J.; Boradia, V.M.; Hammond, H.L.; Pedra, J.H.F. p47 licenses activation of the immune deficiency pathway in the tick Ixodes scapularis. Proc. Natl. Acad. Sci. USA 2018, 116, 205–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsheim, R.F.; Herron, M.J.; Nelson, C.M.; Burkhardt, N.Y.; Barbet, A.F.; Kurtti, T.J.; Munderloh, U.G. Transformation of Anaplasma phagocytophilum. BMC Biotechnol. 2006, 6, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Resistance of Vectors of Disease to Pesticides: Fifth Report of the WHO Expert Committee in Vector Biology and Control; World Health Organization Technical Report Series; World Health Organization: Geneva, Switzerland, 1980; pp. 1–82.
- Kotsarenko, K.; Vechtova, P.; Lieskovska, J.; Füssy, Z.; Cabral-De-Mello, D.C.; Rego, R.; Alberdi, P.; Collins, M.; Bell-Sakyi, L.; Sterba, J.; et al. Karyotype changes in long-term cultured tick cell lines. Sci. Rep. 2020, 10, 13443. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Xu, Y.; Lu, P.; Xie, Y.; Xi, Z. The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti. PLoS Pathog. 2010, 6, e1000833. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skinner, K.M.; Underwood, J.; Ghosh, A.; Oliva Chavez, A.S.; Brelsfoard, C.L. Wolbachia Impacts Anaplasma Infection in Ixodes scapularis Tick Cells. Int. J. Environ. Res. Public Health 2022, 19, 1051. https://doi.org/10.3390/ijerph19031051
Skinner KM, Underwood J, Ghosh A, Oliva Chavez AS, Brelsfoard CL. Wolbachia Impacts Anaplasma Infection in Ixodes scapularis Tick Cells. International Journal of Environmental Research and Public Health. 2022; 19(3):1051. https://doi.org/10.3390/ijerph19031051
Chicago/Turabian StyleSkinner, Kalin M., Jacob Underwood, Arnab Ghosh, Adela S. Oliva Chavez, and Corey L. Brelsfoard. 2022. "Wolbachia Impacts Anaplasma Infection in Ixodes scapularis Tick Cells" International Journal of Environmental Research and Public Health 19, no. 3: 1051. https://doi.org/10.3390/ijerph19031051