
Multifactorial Landscape Parses to Reveal a Predictive Model for Knee Osteoarthritis

 , and
, and
Abstract
:1. Introduction
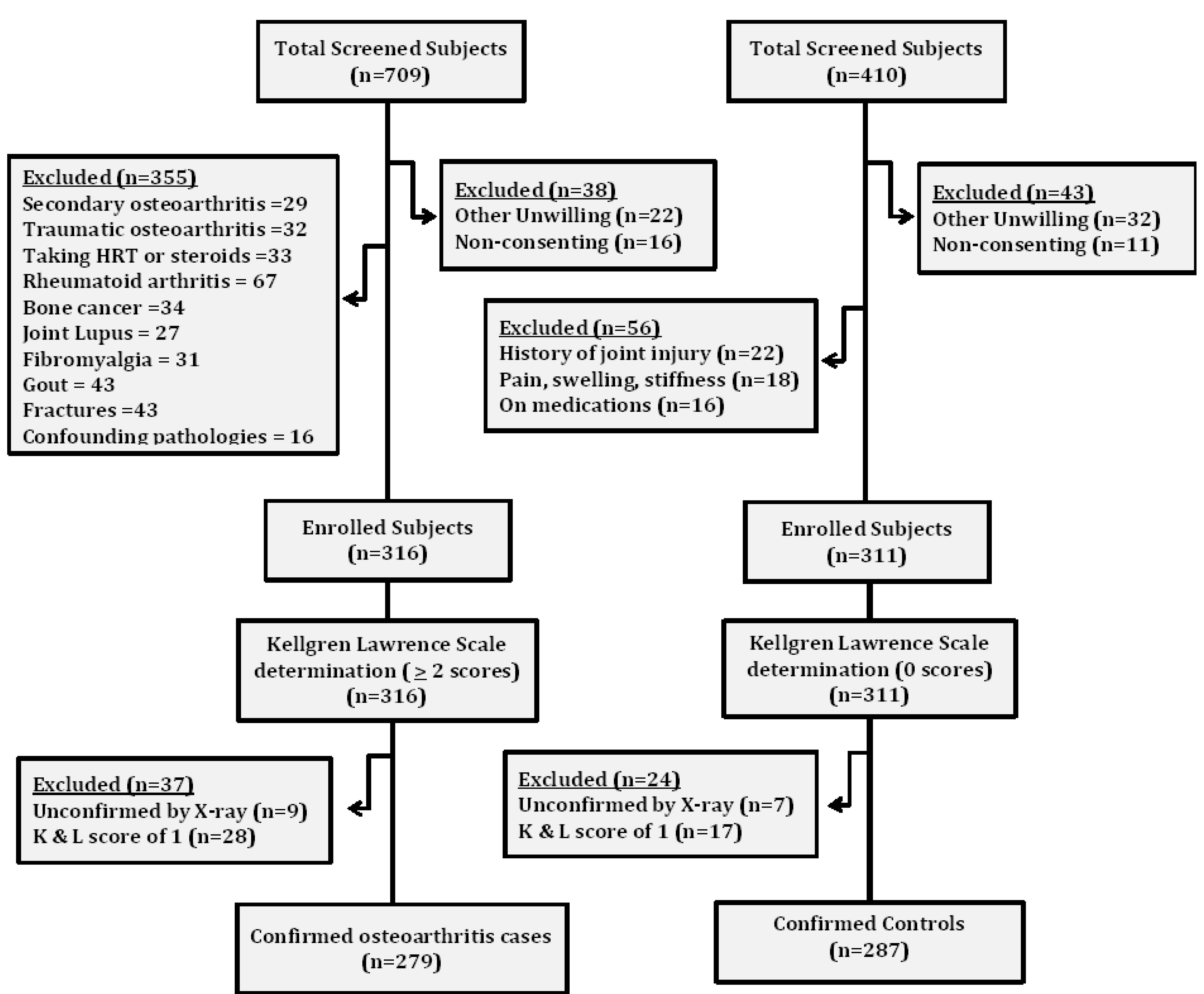
2. Subjects and Methods
2.1. Subjects
2.2. Sleep Quality and Biochemical Variables
2.3. SNP Selection and Genotyping
2.4. Statistical Power and Population Stratification
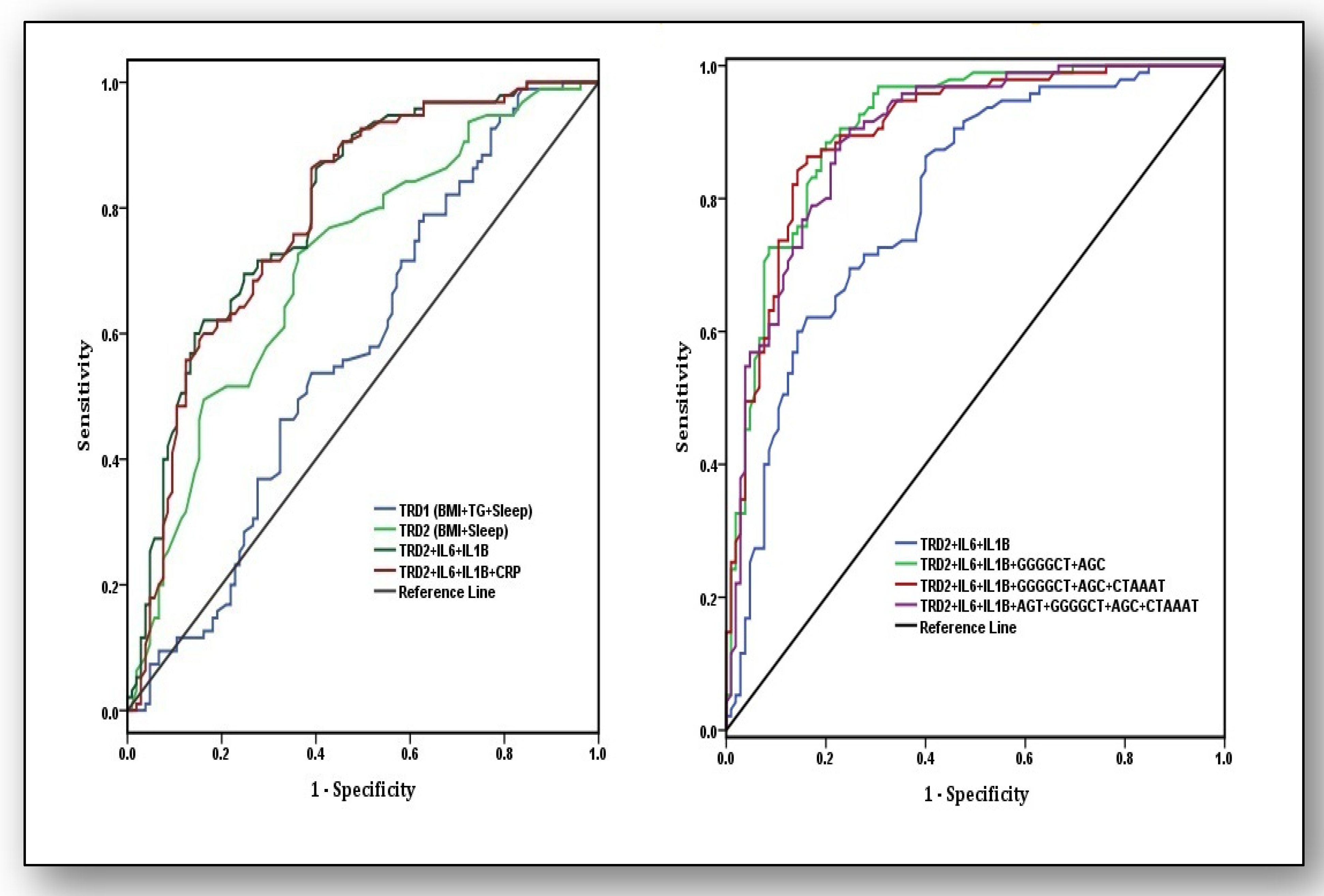
2.5. Receiver Operating Characteristic Curve Analysis
2.6. Statistical Analysis
3. Results
3.1. Variables at Baseline and Genetic Correlates
3.2. Detection of Independent Risk Predictors
3.3. SNP–SNP Cross Talks, Risky Traits, and Their Modes of Association
3.4. Haplotype Analysis, Their Risk and Best Fit Mode of Their Manifestation
3.5. Predictive Ability of Traditional Risk Factors, Biochemical Parameters, and Haplotypes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldgring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.P. Osteoarthritis. Nat. Rev. Dis. Primers 2016, 2, 16072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.-P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2010, 7, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Epstein, F.H.; Hamerman, D. The Biology of Osteoarthritis. N. Engl. J. Med. 1989, 320, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Kwan Tat, S.; Padrines, M.; Théoleyre, S.; Heymann, D.; Fortun, Y. IL-6, RANKL, TNF-alpha/IL-1: Interrelations in bone resorption pathophysiology. Cytokine Growth Factor Rev. 2004, 15, 49–60. [Google Scholar] [PubMed]
- Pollard, T.C.B.; Gwilym, S.E.; Carr, A.J. The assessment of early osteoarthritis. J. Bone Jt. Surg. Br. Vol. 2008, 90, 411–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Moreno, M.; Rego, I.; Carreira-Garcia, V.; Blanco, F.J. Genetics in osteoarthritis. Curr. Genom. 2008, 9, 542–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Mastana, S.; Singh, S.; Juneja, P.K.; Kaur, T.; Singh, P. Promoter polymorphisms in IL-6 gene influence pro-inflammatory cytokines for the risk of osteoarthritis. Cytokine 2020, 127, 154985. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, M.; Sheikh, N.; Suqaina, S.K.; Batool, A.; Fatima, N.; Mehmood, R.; Nazir, S. Vitamin D Receptor Gene Polymorphism: An Important Predictor of Arthritis Development. Biomed. Res. Int. 2019, 2019, 8326246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, A.R.; Di Cesare, P.E.; Vyas, P.; Attur, M.; Tzeng, E.; Billiar, T.R.; Stuchin, S.A.; Abramson, S.B. The expression and regulation of nitric oxide synthase in human osteoarthritis-affected chondrocytes: Evidence for up-regulated neuronal nitric oxide synthase. J. Exp. Med. 1995, 182, 2097–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remst, D.F.G.; Blom, A.B.; Vitters, E.L.; Bank, R.A.; Berg, W.B.V.D.; Davidson, E.N.B.; Van Der Kraan, P.M. Gene Expression Analysis of Murine and Human Osteoarthritis Synovium Reveals Elevation of Transforming Growth Factor β-Responsive Genes in Osteoarthritis-Related Fibrosis. Arthritis Rheumatol. 2014, 66, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Menashe, I.; Rosenberg, P.S.; Chen, B.E. PGA: Power calculator for case-control genetic association analyses. BMC Genet. 2008, 9, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Runesha, H.B.; Dvorkin, D.; Garbe, J.R.; Da, Y. Parallel and serial computing tools fortesting single-locus and epistatic SNP effects of quantitative traits in genome-wideassociationstudies. BMC Bioinform. 2008, 9, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel-Pelletier, J.; Raynauld, J.-P.; Caron, J.; Mineau, F.; Abram, F.; Dorais, M.; Haraoui, B.; Choquette, D. Decrease in serum level of matrix metalloproteinases is predictive of the disease-modifying effect of osteoarthritis drugs assessed by quantitative MRI in patients with knee osteoarthritis. Ann. Rheum. Dis. 2010, 69, 2095–2101. [Google Scholar] [CrossRef] [PubMed]
- Pearle, A.; Scanzello, C.; George, S.; Mandl, L.; DiCarlo, E.; Peterson, M.; Sculco, T.; Crow, M. Elevated high-sensitivity C-reactive protein levels are associated with local inflammatory findings in patients with osteoarthritis. Osteoarthr. Cartil. 2007, 15, 516–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerkhof, H.J.M.; Bierma-Zeinstra, S.M.A.; Castano-Betancourt, M.C.; De Maat, M.P.; Hofman, A.; Pols, H.A.P.; Rivadeneira, F.; Witteman, J.C.; Uitterlinden, A.G.; Van Meurs, J.B.J. Serum C reactive protein levels and genetic variation in the CRP gene are not associated with the prevalence, incidence or progression of osteoarthritis independent of body mass index. Ann. Rheum. Dis. 2010, 69, 1976–1982. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Sun, H.-J.; Wang, Y.-H.; Zhang, Z. Relationships of common polymorphisms in IL-6, IL-1A, and IL-1B genes with susceptibility to osteoarthritis: A meta-analysis. Clin. Rheumatol. 2014, 34, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Walston, J.; Arking, D.; Fallin, D.; Li, T.; Beamer, B.; Xue, Q.; Ferrucci, L.; Fried, L.; Chakravarti, A. IL-6 gene variation is not associated with increased serum levels of IL-6, muscle, weakness, or frailty in older women. Exp. Gerontol. 2005, 40, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Heidari, B.; Babaei, M. Therapeutic and Preventive Potential of Vitamin D Supplementation in Knee Osteoarthritis. ACR Open Rheumatol. 2019, 1, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.R.; Zhu, X.; Storfer-Isser, A.; Mehra, R.; Jenny, N.S.; Trcy, R.; Redline, S. Sleep duration and biomarkers of inflammation. Sleep 2009, 32, 200–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Variables | Cases (n = 279) | Controls (n = 287) | p Value (95%CI) |
|---|---|---|---|
| Age | 59.22 (9.41) | 58.57 (7.27) | 0.357 (0.74–2.04) |
| Gender (men/women) | 138/141 | 142/145 | 0.997 (0.72–1.39) |
| Systolic blood pressure (mmHg) | 135.44 (19.52) | 133.05 (22.22) | 0.175 (0.94–5.85) |
| Diastolic blood pressure (mmHg) | 86.75 (16.32) | 84.91 (14.42) | 0.155 (0.70–4.38) |
| Body mass index (Kg/m2) | 29.40 (4.32) | 26.72 (4.45) | <0.001 (1.95–3.40) |
| Total cholesterol (mg/dL) | 168.82 (38.72) | 163.29 (39.40) | 0.093 (0.92–11.98) |
| Triglycerides (mg/dL) | 169.36 (22.80) | 155.72 (28.60) | <0.001 (9.36–17.92) |
| Low density lipoprotein (mg/dL) | 212.30 (35.55) | 152.34 (27.39) | <0.001 (54.72–65.19) |
| Current smokers | 92 (32.97) | 82 (28.57) | 0.256 (0.86–1.76) |
| Non-smokers | 187 (67.03) | 205 (71.43) | |
| Current alcohol drinkers | 93 (33.33) | 88 (30.66) | 0.500 (0.79–1.61) |
| Non-drinkers | 186 (66.67) | 199 (69.34) | |
| Subjects having good sleep | 81 (29.03) | 219 (76.31) | <0.001 (0.09–0.18) |
| Subjects having poor sleep | 198 (70.97) | 68 (23.69) | |
| Biochemical parameters | |||
| IL-6 levels (pg/mL) ǂ | 4.7 (1.1, 8.2) | 2.3 (0.9, 3.4) | <0.001 |
| IL-1β levels (pg/mL) ǂ | 3.8 (1.3, 6.4) | 1.5 (0.7, 2.4) | <0.001 |
| TNF-α levels (pg/mL) ǂ | 2.1 (1.0, 3.1) | 1.4 (0.6, 2.2) | 0.077 |
| hsCRP levels (mg/L) ǂ | 2.9 (0.9, 4.1) | 1.3 (0.7, 1.9) | 0.005 |
| CRP gene/SNPs | |||
| MAF; rs2794521 † | 0.25 ± 0.026 | 0.14 ± 0.021 | 0.001 |
| MAF; rs1800947 † | 0.15 ± 0.022 | 0.12 ± 0.022 | 0.335 |
| MAF; rs1130864 † | 0.29 ± 0.027 | 0.18 ± 0.023 | 0.002 |
| COL1AI gene/SNPs | |||
| MAF; rs1107946 † | 0.14 ± 0.021 | 0.21 ± 0.024 | 0.029 |
| MAF; rs1800012 † | 0.22 ± 0.023 | 0.16 ± 0.020 | 0.049 |
| IL-6 gene/SNPs | |||
| MAF; rs1800795 † | 0.19 ± 0.023 | 0.36 ± 0.028 | <0.001 |
| MAF; rs1800796 † | 0.20 ± 0.024 | 0.28 ± 0.026 | 0.024 |
| MAF; rs1800797 † | 0.19 ± 0.024 | 0.27 ± 0.026 | 0.024 |
| MAF; rs2069827 † | 0.14 ± 0.021 | 0.17 ± 0.022 | 0.325 |
| MAF; rs12700386 † | 0.24 ± 0.026 | 0.17 ± 0.022 | 0.040 |
| MAF; rs10499563 † | 0.16 ± 0.022 | 0.30 ± 0.020 | <0.001 |
| VDR gene/SNPs | |||
| MAF; rs2228570 † | 0.41 ± 0.029 | 0.31 ± 0.027 | 0.012 |
| MAF; rs1544410 † | 0.47 ± 0.030 | 0.35 ± 0.028 | 0.004 |
| MAF; rs731236 † | 0.43 ± 0.030 | 0.34 ± 0.028 | 0.029 |
| eNOS gene/SNPs | |||
| MAF; rs2070744 † | 0.23 ± 0.025 | 0.16 ± 0.021 | 0.032 |
| MAF; rs1799983 † | 0.22 ± 0.025 | 0.10 ± 0.018 | <0.001 |
| MAF; rs1800780 † | 0.45 ± 0.030 | 0.46 ±0.029 | 0.811 |
| MAF; rs3918181 † | 0.38 ± 0.029 | 0.33 ± 0.028 | 0.216 |
| MAF; rs891512 † | 0.24 ± 0.026 | 0.16 ± 0.022 | 0.019 |
| MAF; rs1808593 † | 0.21 ± 0.024 | 0.14 ± 0.020 | 0.025 |
| Variables | Univariate Analysis | Multivariate Analysis | ||||||
|---|---|---|---|---|---|---|---|---|
| Β | Exp (β) | 95%CI | p Value | Β | Exp (β) | 95%CI | p Value | |
| SBP (mmHg) | 0.718 | 2.05 | 0.93–3.15 | 0.596 | ---- | ----- | ----- | ---- |
| DBP (mmHg) | 0.559 | 1.75 | 0.87–2.84 | 0.791 | ---- | ----- | ----- | ---- |
| BMI (kg/m2) | 0.788 | 2.20 | 1.64–2.92 | 0.002 | 0.667 | 1.95 | 1.45–2.74 | 0.013 |
| TC (mg/dL) | 0.693 | 2.00 | 0.90–2.91 | 0.832 | ---- | ----- | ----- | ---- |
| LDL (mg/dL) | 0.615 | 1.85 | 1.37–2.75 | 0.034 | 0.451 | 1.57 | 0.96–2.38 | 0.058 |
| TG (mg/dL) | 1.019 | 2.74 | 1.45–3.15 | <0.001 | 0.891 | 2.44 | 1.31–2.98 | 0.007 |
| Sleep (global score) | 1.232 | 3.43 | 1.88–3.62 | <0.001 | 1.153 | 3.17 | 1.72–3.09 | 0.004 |
| IL-6 (pg/mL) | 1.175 | 3.24 | 1.73–3.16 | <0.001 | 1.098 | 3.00 | 1.62–3.01 | 0.002 |
| IL-1β (pg/mL) | 1.040 | 2.83 | 1.12–2.83 | 0.007 | 0.920 | 2.51 | 1.07–2.67 | 0.028 |
| TNF-α (pg/mL) | 0.625 | 1.87 | 0.84–2.85 | 0.572 | ----- | ----- | ----- | ----- |
| hsCRP(mg/L) | 0.577 | 1.78 | 1.23–4.16 | 0.003 | 0.501 | 1.65 | 1.15–3.75 | 0.021 |
| SNP | SNP | Trait | Test | Post | Pnost |  |
| rs1800795 | rs1800796 | IL-6 | DD | 0.0003 | 0.037 | |
| rs1800795 | rs1800797 | TG | I | 0.0021 | 0.135 | |
| rs1800795 | rs2228570 | hsCRP | DD | 0.0036 | 1.029 | |
| rs1800795 | rs12700386 | BMI | DA | 0.0013 | 0.558 | |
| rs1800795 | rs10499563 | IL-1β | I | 0.0025 | 0.782 | |
| rs1800795 | rs2794521 | Sleep | AA | 0.0017 | 0.139 | |
| rs1800795 | rs1130864 | IL-1β | AD | 0.0084 | 0.274 | |
| rs1800795 | rs891512 | Sleep | DA | 0.0020 | 0.038 | |
| rs1800795 | rs731236 | BMI | AD | 0.0043 | 0.293 | |
| rs1800797 | rs18008593 | IL-1β | AD | 0.0035 | 0.536 | |
| rs1800796 | rs1107946 | TG | DA | 0.0055 | 0.823 | |
| rs12700386 | rs1800947 | hsCRP | DD | 0.0036 | 0.178 | |
| rs2794521 | rs1544410 | TG | I | 0.0014 | 0.221 | |
| rs891512 | rs2070744 | BMI | AA | 0.0028 | 0.135 | |
| rs2228570 | rs1800012 | Sleep | AA | 0.0063 | 0.835 |
| Haplotype | Cases (n = 279) | Controls (n = 287) | PCor | Unadjusted OR (95%CI) | p Value | Adjusted OR (95%CI) a | p Value |
|---|---|---|---|---|---|---|---|
| CRP gene | |||||||
| AGC | 0.46 (130) | 0.54 (155) | 0.10 | Referent | ---- | Referent | ---- |
| GGT | 0.05 (14) | 0.07 (21) | 0.46 | 0.79 (0.39–1.63) | 0.65 | 0.77 (0.36–1.53) | 0.56 |
| AGT | 0.24 (68) | 0.09 (26) | <0.001 | 3.12 (1.88–5.19) | <0.001 | 2.73 (1.63–4.72) | 0.002 |
| GCT | 0.09 (26) | 0.07 (20) | 0.55 | 1.55 (0.83–2.90) | 0.22 | 1.31 (0.71–2.55) | 0.19 |
| GCC | 0.06 (18) | 0.08 (24) | 0.68 | 0.89 (0.46–1.72) | 0.87 | 0.77 (0.38–1.52) | 0.67 |
| COL1A1 gene | |||||||
| GG | 0.59 (165) | 0.57(171) | 0.99 | Referent | ---- | Referent | ---- |
| GT | 0.20 (55) | 0.13 (38) | 0.004 | 1.68 (1.04–2.70) | 0.04 | 1.54 (0.93–2.19) | 0.09 |
| TG | 0.08 (23) | 0.10 (30) | 0.65 | 0.79 (0.44–1.42) | 0.53 | 0.72 (0.38–1.26) | 0.47 |
| TT | 0.07(19) | 0.08 (24) | 0.81 | 0.82 (0.43–1.55) | 0.66 | 0.74 (0.36–1.38) | 0.52 |
| IL-6 gene | |||||||
| GGGGGT | 0.23 (65) | 0.25 (71) | 0.95 | Referent | -------- | Referent | ------ |
| CCGCGC | 0.08 (22) | 0.07 (19) | 0.88 | 1.26 (0.63–2.55) | 0.63 | 1.11 (0.54–2.31) | 0.57 |
| CTACAT | 0.06 (18) | 0.07 (21) | 0.95 | 0.94 (0.46–1.91) | 0.99 | 0.87 (0.43–1.82) | 0.82 |
| GGGGCT | 0.15 (43) | 0.07 (21) | <0.001 | 2.24 (1.20–4.16) | 0.02 | 2.10 (1.08–3.79) | 0.04 |
| CCGTAT | 0.09 (25) | 0.08 (24) | 0.99 | 1.14 (0.59–2.19) | 0.82 | 1.02 (0.49–2.00) | 0.71 |
| CTGCAC | 0.07 (19) | 0.07 (21) | 0.99 | 0.99 (0.49–2.00) | 0.88 | 0.79 (0.39–1.89) | 0.69 |
| CTGTAT | 0.08 (22) | 0.09 (27) | 0.84 | 0.89 (0.46–1.71) | 0.86 | 0.71 (0.37–1.43) | 0.62 |
| CGAGGC | 0.07 (21) | 0.14 (49) | 0.007 | 0.57 (0.31–1.07) | 0.11 | 0.63 (0.51–1.12) | 0.93 |
| VDR gene | |||||||
| GGT (baT) | 0.48 (133) | 0.51 (147) | 0.70 | Referent | -------- | Referent | ------ |
| ATC (BAt) | 0.09 (24) | 0.09 (27) | 0.97 | 0.98 (0.54–1.79) | 0.92 | 0.83 (0.4301.54) | 0.83 |
| GTT (bAT) | 0.10 (29) | 0.09 (26) | 0.90 | 1.23 (0.69–2.20) | 0.57 | 1.11 (0.57–2.00) | 0.47 |
| AGC (Bat) | 0.14 (38) | 0.06 (17) | <0.001 | 2.47 (1.33–4.58) | 0.005 | 2.10 (1.26–3.93) | 0.01 |
| ATT (BAT) | 0.08 (23) | 0.07 (19) | 0.78 | 1.34 (0.70–2.57) | 0.48 | 1.13 (0.60–2.28) | 0.33 |
| GTC (bAt) | 0.07 (19) | 0.05 (16) | 0.86 | 1.31 (0.65–2.66) | 0.56 | 1.10 (0.52–2.38) | 0.31 |
| eNOS gene | |||||||
| TTAGGG | 0.14 (40) | 0.19 (55) | 0.19 | Referent | -------- | Referent | ------ |
| TTGGGG | 0.13 (36) | 0.14 (41) | 0.93 | 1.21 (0.66–2.21) | 0.65 | 1.10 (0.55–1.98) | 0.52 |
| CGAAGG | 0.12 (33) | 0.13 (38) | 0.91 | 1.19 (0.64–2.22) | 0.69 | 1.00 (0.47–1.87) | 0.48 |
| CTAAAT | 0.25 (69) | 0.08 (24) | 1 × 10−11 | 3.95 (2.13–7.33) | <0.001 | 3.12 (1.99–6.72) | 0.006 |
| CGGAGG | 0.10 (28) | 0.09 (26) | 0.96 | 1.48 (0.76–2.90) | 0.33 | 1.27 (0.65–2.45) | 0.28 |
| CTGGAT | 0.09 (24) | 0.09 (27) | 0.97 | 1.22 (0.62–2.42) | 0.69 | 1.09 (0.46–2.00) | 0.57 |
| TTAGAT | 0.08 (22) | 0.06 (19) | 0.88 | 1.59 (0.76–3.33) | 0.29 | 1.33 (0.63–2.88) | 0.26 |
| CRP Haplotype AGT | |||||
|---|---|---|---|---|---|
| Model | ǂ β (SE) | Wald Test | p Value | R2h | AIC |
| Dominant | 0.48 (0.53) | 0.90 | 0.510 | 0.5972 | 4210.38 |
| Recessive | 2.11(0.76) | 2.77 | <0.001 | 1.0000 | 2791.39 |
| Multiplicative | −0.31 (0.79) | −0.39 | 0.389 | 0.8817 | 3912.30 |
| General (0 copy) | −0.09 (0.37) | −0.24 | 0.560 | 0.9280 | 4142.49 |
| General (1 copy) | 0.70 (0.89) | 0.78 | 0.021 | 0.9663 | 3358.40 |
| IL-6 Haplotype GGGGCT | |||||
| Dominant | 0.52(0.64) | 0.81 | 0.629 | 0.8213 | 6571.11 |
| Recessive | 2.75 (0.59) | 4.66 | <0.001 | 0.9922 | 2785.31 |
| Multiplicative | 0.20 (0.43) | 0.46 | 0.372 | 0.7661 | 4559.90 |
| General (0 copy) | 0.19 (0.32 | 0.59 | 0.624 | 0.8317 | 5216.29 |
| General (1 copy) | −0.07 (0.51) | −0.14 | 0.721 | 0.8922 | 5888.91 |
| VDR Haplotype AGC | |||||
| Dominant | 2.35 (0.65) | 3.61 | <0.001 | 1.000 | 4635.58 |
| Recessive | 0.28 (0.53) | 0.53 | 0.412 | 0.780 | 5128.44 |
| Multiplicative | 0.72 (0.39) | 1.85 | 0.782 | 0.458 | 5012.48 |
| General (0 copy) | −0.62 (0.41) | −1.51 | 0.212 | 0.673 | 5344.25 |
| General (1 copy) | 0.007 (0.24) | 0.029 | 0.991 | 0.887 | 4989.16 |
| eNOS Haplotype CTAAAT | |||||
| Dominant | 0.33 (0.67) | 0.49 | 0.562 | 0.667 | 7571.20 |
| Recessive | 0.58 (0.63) | 0.92 | 0.411 | 0.540 | 7255.23 |
| Multiplicative | 1.89 (0.52) | 3.63 | <0.001 | 0.975 | 6965.78 |
| General (0 copy) | −0.42 (0.33) | −1.27 | 0.211 | 0.521 | 7349.90 |
| General (1 copy) | 0.83 (0.34) | 2.44 | 0.689 | 0.634 | 7136.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, M.; Valecha, S.; Khinda, R.; Kumar, N.; Singh, S.; Juneja, P.K.; Kaur, T.; Di Napoli, M.; Minhas, J.S.; Singh, P.; et al. Multifactorial Landscape Parses to Reveal a Predictive Model for Knee Osteoarthritis. Int. J. Environ. Res. Public Health 2021, 18, 5933. https://doi.org/10.3390/ijerph18115933
Singh M, Valecha S, Khinda R, Kumar N, Singh S, Juneja PK, Kaur T, Di Napoli M, Minhas JS, Singh P, et al. Multifactorial Landscape Parses to Reveal a Predictive Model for Knee Osteoarthritis. International Journal of Environmental Research and Public Health. 2021; 18(11):5933. https://doi.org/10.3390/ijerph18115933
Chicago/Turabian StyleSingh, Monica, Srishti Valecha, Rubanpal Khinda, Nitin Kumar, Surinderpal Singh, Pawan K. Juneja, Taranpal Kaur, Mario Di Napoli, Jatinder S. Minhas, Puneetpal Singh, and et al. 2021. "Multifactorial Landscape Parses to Reveal a Predictive Model for Knee Osteoarthritis" International Journal of Environmental Research and Public Health 18, no. 11: 5933. https://doi.org/10.3390/ijerph18115933