
The Effects of Childhood Maltreatment on Non-Suicidal Self-Injury in Male Adolescents: The Moderating Roles of the Monoamine Oxidase A (MAOA) Gene and the Catechol-O-Methyltransferase (COMT) Gene


Abstract
:1. Introduction
1.1. Childhood Maltreatment and NSSI
1.2. Dopaminergic Genes and NSSI
1.3. Gene × Gene × Environment Interactions
1.4. Current Study
2. Materials and Methods
2.1. Participants and Procedures
2.2. Measures
2.2.1. Child Abuse and Neglect
2.2.2. Nonsuicidal Self-Injury (NSSI)
2.2.3. Covariables
2.2.4. Genotyping
2.3. Data Analyses
3. Results
3.1. Hardy-Weinberg Equilibrium Test
3.2. Descriptive Statistics and Correlations
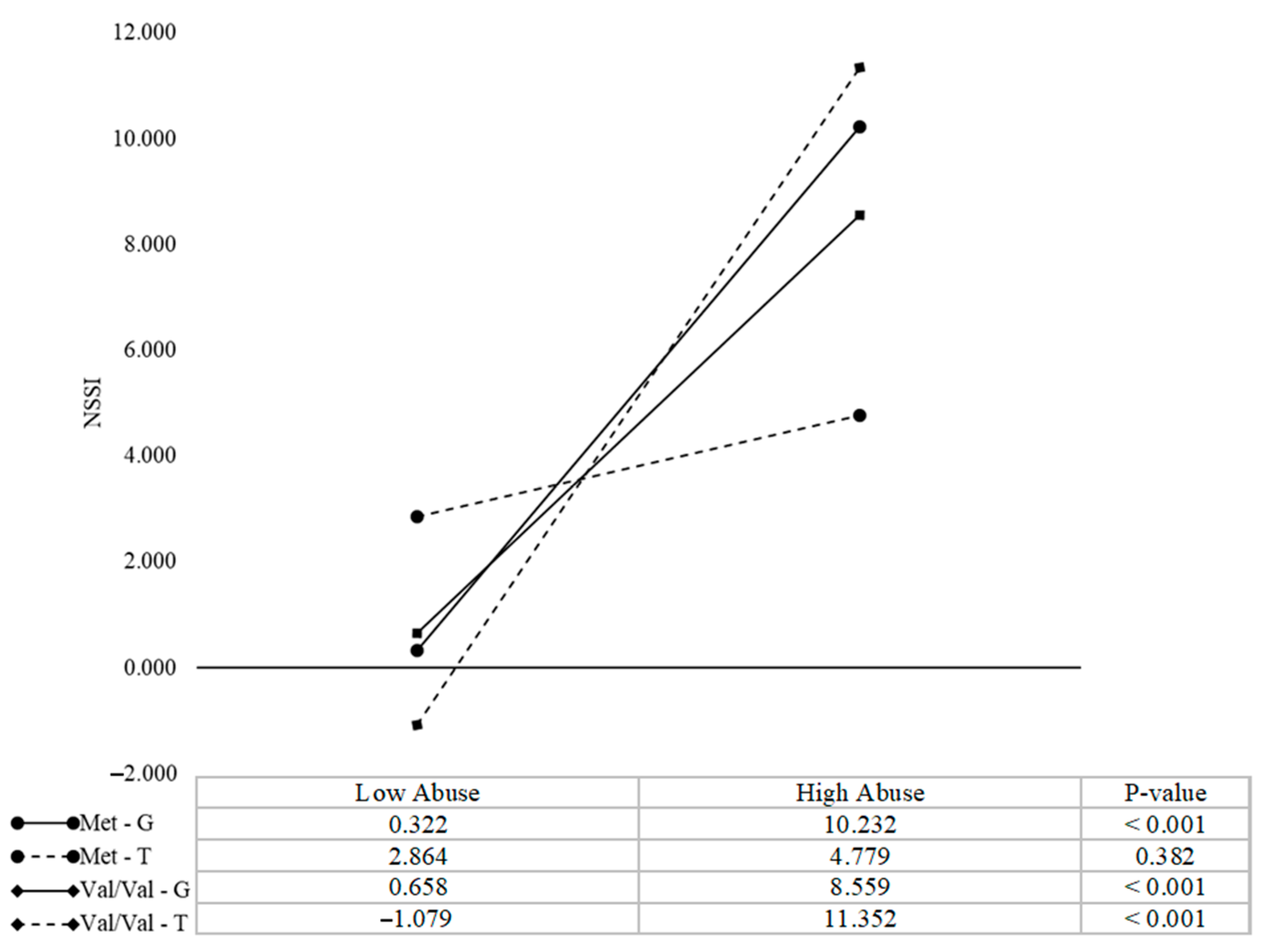
3.3. Interaction Effects of MAOA, COMT, and Child Abuse on NSSI
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nock, M.K. Why do people hurt themselves? New insights into the nature and functions of self-injury. Curr. Dir. Psychol. Sci. 2009, 18, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, C.M.; Gould, M. The epidemiology and phenomenology of non-suicidal self-injurious behavior among adolescents: A critical review of the literature. Arch. Suicide Res. 2007, 11, 129–147. [Google Scholar] [CrossRef]
- Swannell, S.V.; Martin, G.E.; Page, A.; Hasking, P.; St John, N.J. Prevalence of nonsuicidal self-injury in nonclinical samples: Systematic review, meta-analysis and meta-regression. Suicide Life Threat. Behav. 2014, 44, 273–303. [Google Scholar] [CrossRef]
- Tang, J.; Li, G.; Chen, B.; Huang, Z.; Zhang, Y.; Chang, H.; Wu, C.; Ma, X.; Wang, J.; Yu, Y. Prevalence of and risk factors for non-suicidal self-injury in rural china: Results from a nationwide survey in China. J. Affect. Disord. 2018, 226, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Plener, P.L.; Kaess, M.; Schmahl, C.; Pollak, S.; Fegert, J.M.; Brown, R.C. Nonsuicidal Self-Injury in Adolescents. Dtsch. Ärzteblatt Int. 2018, 115, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.D.; Franklin, J.C.; Fox, K.R.; Bentley, K.H.; Kleiman, E.M.; Chang, B.P.; Nock, M.K. Self-injurious thoughts and behaviors as risk factors for future suicide ideation, attempts, and death: A meta-analysis of longitudinal studies. Psychol. Med. 2016, 46, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Maciejewski, D.F.; Creemers, H.E.; Lynskey, M.T.; Madden, P.A.F.; Heath, A.C.; Statham, D.J.; Martin, N.G.; Verweij, K.J.H. Overlapping genetic and environmental influences on nonsuicidal self-injury and suicidal ideation: Different outcomes, same etiology? JAMA Psychiatry 2014, 71, 699. [Google Scholar] [CrossRef] [Green Version]
- Asselmann, E.; Hertel, J.; Beesdo-Baum, K.; Schmidt, C.; Homuth, G.; Nauck, M.; Grabe, H.; Pané-Farré, C.A. Interplay between COMT Val158Met, childhood adversities and sex in predicting panic pathology: Findings from a general population sample. J. Affect. Disord. 2018, 234, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Beach, S.R.H.; Brody, G.H.; Gunter, T.D.; Packer, H.; Wernett, P.; Philibert, R.A. Child maltreatment moderates the association of MAOA with symptoms of depression and antisocial personality disorder. J. Fam. Psychol. 2010, 24, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.T.; Scopelliti, K.M.; Pittman, S.K.; Zamora, A.S. Childhood maltreatment and non-suicidal self-injury: A systematic review and meta-analysis. Lancet Psychiatry 2018, 5, 51–64. [Google Scholar] [CrossRef]
- Yates, T.M. The developmental psychopathology of self-injurious behavior: Compensatory regulation in posttraumatic adaptation. Clin. Psychol. Rev. 2004, 24, 35–74. [Google Scholar] [CrossRef] [PubMed]
- Asgeirsdottir, B.B.; Sigfusdottir, I.D.; Gudjonsson, G.H.; Sigurdsson, J.F. Associations between sexual abuse and family conflict/violence, self-injurious behavior, and substance use: The mediating role of depressed mood and anger. Child Abus. Negl. 2011, 35, 210–219. [Google Scholar] [CrossRef]
- Kang, N.; Jiang, Y.; Ren, Y.; Gong, T.; Liu, X.; Leung, F.; You, J. Distress intolerance mediates the relationship between child maltreatment and nonsuicidal self-injury among Chinese adolescents: A three-wave longitudinal study. J. Youth Adolesc. 2018, 47, 2220–2230. [Google Scholar] [CrossRef] [PubMed]
- Swannell, S.; Martin, G.; Page, A.; Hasking, P.; Hazell, P.; Taylor, A.; Protani, M. Child maltreatment, subsequent non-suicidal self-injury and the mediating roles of dissociation, alexithymia and self-blame. Child Abus. Negl. 2012, 36, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Wachter, T.; Murphy, S.; Kennerley, H.; Wachter, S. A preliminary study examining relationships between childhood maltreatment, dissociation, and self-injury in psychiatric outpatients. J. Trauma Dissociation 2009, 10, 261–275. [Google Scholar] [CrossRef]
- Serafini, G.; Canepa, G.; Adavastro, G.; Nebbia, J.; Murri, M.B.; Erbuto, D.; Pocai, B.; Fiorillo, A.; Pompili, M.; Flouri, E.; et al. The relationship between childhood maltreatment and non-suicidal self-injury: A systematic review. Front. Psychiatry 2017, 8, 149. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, B.S. Early Childhood Environment and Genetic Interactions: The Diathesis for Suicidal Behavior. Curr. Psychiatry Rep. 2016, 18, 86. [Google Scholar] [CrossRef]
- Chapman, A.L.; Gratz, K.L.; Brown, M.Z. Solving the puzzle of deliberate self-harm: The experiential avoidance model. Behav. Res. Ther. 2006, 44, 371–394. [Google Scholar] [CrossRef]
- Belmaker, R.H.; Agam, G. Mechanisms of disease: Major depressive disorder. N. Engl. J. Med. 2008, 358, 55–68. [Google Scholar] [CrossRef] [Green Version]
- Hamon, M.; Blier, P. Monoamine neurocircuitry in depression and strategies for new treatments. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 45, 54–63. [Google Scholar] [CrossRef]
- Melas, P.A.; Forsell, Y. Hypomethylation of MAOA’s first exon region in depression: A replication study. Psychiatry Res. 2015, 226, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Lindenberg, A.; Buckholtz, J.W.; Kolachana, B.; Hariri, A.R.; Pezawas, L.; Blasi, G.; Wabnitz, A.; Honea, R.; Verchinski, B.; Callicott, J.H.; et al. Neural mechanisms of genetic risk for impulsivity and violence in humans. Proc. Natl. Acad. Sci. USA 2006, 103, 6269–6274. [Google Scholar] [CrossRef] [Green Version]
- Caspi, A.; McClay, J.; Moffitt, T.; Mill, J.; Martin, J.; Craig, I.W.; Taylor, A.; Poulton, R. Role of genotype in the cycle of violence in maltreated children. Science 2002, 297, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Breakefield, X.O. Human monoamine oxidase A gene determines levels of enzyme activity. Am. J. Hum. Genet. 1991, 49, 383–392. [Google Scholar] [PubMed]
- Byrd, A.L.; Manuck, S.B.; Hawes, S.W.; Vebares, T.J.; Nimgaonkar, V.; Chowdari, K.V.; Hipwell, A.E.; Keenan, K.; Stepp, S.D. The interaction between monoamine oxidase A (MAOA) and childhood maltreatment as a predictor of personality pathology in females: Emotional reactivity as a potential mediating mechanism. Dev. Psychopathol. 2019, 31, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Cicchetti, D.; Rogosch, F.A.; Sturge-Apple, M. Interactions of child maltreatment and serotonin transporter and monoamine oxidase A polymorphisms: Depressive symptomatology among adolescents from low socioeconomic status backgrounds. Dev. Psychopathol. 2007, 19, 1161–1180. [Google Scholar] [CrossRef]
- Kolla, N.J.; Meyer, J.; Sanches, M.; Charbonneau, J. Monoamine oxidase-A genetic variants and childhood abuse predict impulsiveness in borderline personality disorder. Clin. Psychopharmacol. Neurosci. 2017, 15, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Creveling, C.R. The role of catechol-O-methyltransferase in the inactivation of catecholestrogen. Cell. Mol. Neurobiol. 2003, 23, 289–291. [Google Scholar] [CrossRef]
- Lachman, H.M.; Papolos, D.F.; Saito, T.; Yu, Y.M.; Szumlanski, C.L.; Weinshilboum, R.M. Human catechol-O-methyltransferase pharmacogenetics: Description of a functional polymorphism and its potential application to neuropsychiatric disorders. Pharmacogenetics 1996, 6, 243–250. [Google Scholar] [CrossRef]
- Chen, J.; Lipska, B.K.; Halim, N.; Ma, Q.D.; Matsumoto, M.; Melhem, S.; Kolachana, B.S.; Hyde, T.M.; Herman, M.M.; Apud, J. Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): Effects on mRNA, protein, and enzyme activity in postmortem human brain. Am. J. Hum. Genet. 2004, 75, 807–821. [Google Scholar] [CrossRef] [Green Version]
- Drabant, E.M.; Hariri, A.R.; Meyer-Lindenberg, A.; Munoz, K.E.; Munoz, K.E.; Mattay, V.S.; Kolachana, B.S.; Egan, M.F.; Weinberger, D.R. Catechol O-methyltransferase Val158Met genotype and neural mechanisms related to affective arousal and regulation. Arch. Gen. Psychiatry 2006, 63, 1396–1406. [Google Scholar] [CrossRef]
- Crowell, S.E.; Beauchaine, T.P.; Linehan, M.M. A biosocial developmental model of borderline personality: Elaborating and extending Linehan’s theory. Psychol. Bull. 2009, 135, 495–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bresin, K.; Finy, M.S.; Verona, E. Childhood emotional environment and self-injurious behaviors: The moderating role of the BDNF Val66Met polymorphism. J. Affect. Disord. 2013, 150, 594–600. [Google Scholar] [CrossRef]
- Hankin, B.L.; Barrocas, A.L.; Young, J.F.; Haberstick, B.; Smolen, A. 5-HTTLPR × interpersonal stress interaction and nonsuicidal self-injury in general community sample of youth. Psychiatry Res. 2015, 225, 609–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napolitano, A.; Cesura, A.M.; Da Prada, M. The role of monoamine oxidase and catechol O-methyltransferase in dopaminergic neurotransmission. J. Neural Transm. 1995, 45, 35–45. [Google Scholar]
- Bouma, E.M.C.; Riese, H.; Doornbos, B.; Ormel, J.; Oldehinkel, A.J. Genetically based reduced MAOA and COMT functioning is associated with the cortisol stress response: A replication study. Mol. Psychiatry 2012, 17, 119–121. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Wang, M.; Cao, Y.; Ji, L.; Zhang, W. The interactive effects of monoamine oxidase A (MAOA) gene and peer victimization on depressive symptoms in early adolescent boys: The moderating role of catechol-O-methyltransferase (COMT) gene. Acta Psychol. Sin. 2017, 49, 206–218. [Google Scholar] [CrossRef]
- Wang, M.; Li, H.; Deater-Deckard, K.; Zhang, W. Interacting effect of catechol-O-methyltransferase (COMT) and monoamine oxidase a (MAOA) gene polymorphisms, and stressful life events on aggressive behavior in Chinese male adolescents. Front. Psychol. 2018, 9, 1079. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Luo, Y. Effect of genetic polymorphisms on neural circuits of emotion regulation. Adv. Psychol. Sci. 2010, 18, 1440–1448. [Google Scholar]
- Carrel, L.; Willard, H.F. X-inactivation profile reveals extensive variability in X-linked gene expression in females. Nature 2005, 434, 400–404. [Google Scholar] [CrossRef]
- Bernstein, D.P.; Stein, J.A.; Newcomb, M.D.; Walker, E.; Pogge, D.; Ahluvalia, T.; Stokes, J.; Handelsman, L.; Medrano, M.; Desmond, D.; et al. Development and validation of a brief screening version of the childhood trauma questionnaire. Child Abus. Negl. 2003, 27, 169–190. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, Y.; Li, L.; Zhou, Y.; Yang, S. Reliability and validity of the Chinese version of childhood trauma questionnaire. Chin. J. Clin. Rehabil. 2005, 9, 105–107. [Google Scholar]
- Xie, P.; Wu, K.; Zheng, Y.; Guo, Y.; Yang, Y.; He, J.; Ding, Y.; Peng, H. Prevalence of childhood trauma and correlations between childhood trauma, suicidal ideation, and social support in patients with depression, bipolar disorder, and schizophrenia in southern china. J. Affect. Disord. 2018, 228, 41–48. [Google Scholar] [CrossRef]
- Jansen, K.; Cardoso, T.A.; Fries, G.R.; Branco, J.C.; Silva, R.A.; Kauer-Sant’Anna, M.; Kapczinski, F.; Magalhaes, P.V. Childhood trauma, family history, and their association with mood disorders in early adulthood. Acta Psychiatr. Scand. 2016, 134, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y. The Relation of Adolescents’ Self-Harm Behaviors, Individual Emotion Characteristics and Family Environment Factors. Master’s Thesis, Central China Normal University, Wuhan, China, 2008. [Google Scholar]
- Yu, L.; Ling, X.; Jiang, G. Impulsivity in non-suicidal self-injurious adolescents in china. Acta Psychol. Sin. 2013, 45, 320–335. [Google Scholar] [CrossRef]
- Gu, H.; Fu, D.; Lu, X.; Xia, T. The Relationship between Negative Life Events and Non-suicidal Self-injury among Junior Middle School Students: Moderated Mediation Effect. Psychol. Dev. Educ. 2018, 34, 229–238. [Google Scholar]
- Plener, P.L.; Schumacher, T.S.; Munz, L.M.; Groschwitz, R.C. The longitudinal course of non-suicidal self-injury and deliberate self-harm: A systematic review of the literature. Bord. Personal. Disord. Emot. Dysregulation 2015, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valencia-Agudo, F.; Burcher, G.C.; Ezpeleta, L.; Kramer, T. Nonsuicidal self-injury in community adolescents: A systematic review of prospective predictors, mediators and moderators. J. Adolesc. 2018, 65, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Fendrich, M.; Weissman, M.M.; Warner, V. Screening for depressive disorder in children and adolescents: Validating the center for epidemiologic studies depression scale for children. Am. J. Epidemiol. 1990, 131, 538–551. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Drury, S.S.; Theall, K.P.; Smyke, A.T.; Keats, B.J.B.; Egger, H.L.; Nelson, C.A.; Fox, N.A.; Marshall, P.J.; Zeanah, C.H. Modification of depression by COMT val158met polymorphism in children exposed to early severe psychosocial deprivation. Child Abus. Negl. 2010, 34, 387–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Cao, C.; Wang, M.; Ji, L.; Cao, Y. Monoamine Oxidase A (MAOA) and Catechol-O-Methyltransferase (COMT) Gene Polymorphisms Interact with Maternal Parenting in Association with Adolescent Reactive Aggression but not Proactive Aggression: Evidence of Differential Susceptibility. J. Youth Adolesc. 2016, 45, 812–829. [Google Scholar] [CrossRef]
- Comasco, E.; Åslund, C.; Oreland, L.; Nilsson, K.W. Three-way interaction effect of 5-HTTLPR, BDNFVal66Met, and childhood adversity on depression: A replication study. Eur. Neuropsychopharmacol. 2013, 23, 1300–1306. [Google Scholar] [CrossRef]
- Starr, L.R.; Huang, M. HPA-axis multilocus genetic variation moderates associations between environmental stress and depressive symptoms among adolescents. Dev. Psychopathol. 2019, 31, 1339–1352. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, X.; Xia, G.; Liu, D.; Chen, P.; Zhang, W. Association between negative life events and early adolescents’ depression: The moderating effects of Catechol-O-methyltransferase (COMT) gene val158met polymorphism and parenting behavior. Acta Psychol. Sin. 2019, 51, 903–913. [Google Scholar]
- Zhou, Y.; Liang, Y.; Cheng, J.; Zheng, H.; Liu, Z. Child Maltreatment in Western China: Demographic Differences and Associations with Mental Health. Int. J. Environ. Res. Public Health 2019, 16, 3619. [Google Scholar] [CrossRef] [Green Version]
- Auerbach, R.P.; Kim, J.C.; Chango, J.M.; Spiro, W.J.; Cha, C.; Gold, J.; Esterman, M.; Nock, M.K. Adolescent nonsuicidal self-injury: Examining the role of child abuse, comorbidity, and disinhibition. Psychiatry Res. 2014, 220, 579–584. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, Y. Mediating effect of social support on relationship between childhood neglect and technical school students of self-injury. Chin. J. Sch. Health 2017, 38, 538–541. [Google Scholar]
- Brown, R.C.; Heines, S.; Witt, A.; Braehler, E.; Fegert, J.M.; Harsch, D.; Plener, P.L. The impact of child maltreatment on non-suicidal self-injury: Data from a representative sample of the general population. BMC Psychiatry 2018, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dannlowski, U.; Ohrmann, P.; Konrad, C.; Domschke, K.; Bauer, J.; Kugel, H.; Hohoff, C.; Schöning, S.; Kersting, A.; Baune, B.T.; et al. Reduced amygdala-prefrontal coupling in major depression: Association with MAOA genotype and illness severity. Int. J. Neuropsychopharmacol. 2009, 12, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Gong, P.; Xi, S.; Shen, G.; Li, S.; Zhang, P.; Cao, G.; Zhang, F.; Shen, Y.; Feng, T.; Ma, H. The effects of DBH, MAOA, and MAOB on attentional biases for facial expressions. J. Mol. Neurosci. 2013, 49, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.T.; Pearson, J.K.; Cicchetti, D.; Martin, M.J.; Cummings, E.M. Emotional insecurity as a mediator of the moderating role of dopamine genes in the association between interparental conflict and youth externalizing problems. Dev. Psychopathol. 2019, 31, 1111–1126. [Google Scholar] [CrossRef] [PubMed]
- Weiss, E.M.; Stadelmann, E.; Kohler, C.G.; Brensinger, C.M.; Nolan, K.A.; Oberacher, H.; Parson, W.; Pitterl, F.; Niederstätter, H.; Kemmler, G.; et al. Differential effect of catechol-O-methyltransferase Val158Met genotype on emotional recognition abilities in healthy men and women. J. Int. Neuropsychol. Soc. 2007, 13, 881–887. [Google Scholar] [CrossRef]
- Klucken, T.; Kruse, O.; Wehrum-Osinsky, S.; Hennig, J.; Schweckendiek, J.; Stark, R. Impact of COMT Val158Met-polymorphism on appetitive conditioning and amygdala/prefrontal effective connectivity. Hum. Brain Mapp. 2015, 36, 1093–1101. [Google Scholar] [CrossRef]
- Andersen, S.L.; Teicher, M.H. Stress, sensitive periods and maturational events in adolescent depression. Trends Neurosci. 2008, 31, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Ban, J.; Oh, I. Mediating effects of teacher and peer relationships between parental abuse/neglect and emotional/behavioral problems. Child Abus. Negl. 2018, 61, 35–42. [Google Scholar] [CrossRef]
- Liu, R.T. Childhood maltreatment and impulsivity: A meta-analysis and recommendations for future study. J. Abnorm. Child Psychol. 2019, 47, 221–243. [Google Scholar] [CrossRef]
- Mills, R.; Scott, J.; Alati, R.; O’Callaghan, M.; Najman, J.M.; Strathearn, L. Child maltreatment and adolescent mental health problems in a large birth cohort. Child Abus. Negl. 2013, 37, 292–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glassman, L.H.; Weierich, M.R.; Hooley, J.M.; Deliberto, T.L.; Nock, M.K. Child maltreatment, non-suicidal self-injury, and the mediating role of self-criticism. Behav. Res. Ther. 2007, 45, 2483–2490. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.; Lee, O. Relationships between parental maltreatment and adolescents’ school adjustment: Mediating roles of self-esteem and peer attachment. J. Child Fam. Stud. 2017, 26, 393–404. [Google Scholar] [CrossRef]
- Belsky, J.; Pluess, M. Beyond diathesis stress: Differential susceptibility to environmental influences. Psychol. Bull. 2009, 135, 885–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, K.W.; Comasco, E.; Hodgins, S.; Oreland, L.; Åslund, C. Genotypes do not confer risk for delinquency but rather alter susceptibility to positive and negative environmental factors: Gene-environment interactions of BDNF Val66Met, 5-HTTLPR, and MAOA-uVNTR. Int. J. Neuropsychopharmacol. 2014, 18, pyu107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Lin, X.; Chen, L.; Ji, L.; Zhang, W. The catechol-O-methyltransferase and dopamine transporter genes moderated the impact of peer relationships on adolescent depressive symptoms: A gene–gene–environment study. J. Youth Adolesc. 2018, 47, 2468–2480. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.C.N.; Donat, J.; Brunnet, A.E.; Silva, T.G.; Silva, G.R.; Kristensen, C.H. Cognitive, neurobiological and psychopathological alterations associated with child maltreatment: A review of systematic reviews. Child Indic. Res. 2016, 9, 389–406. [Google Scholar] [CrossRef]
- Thomassin, K.; Shaffer, A.; Madden, A.; Londino, D.L. Specificity of childhood maltreatment and emotion deficit in nonsuicidal self-injury in an inpatient sample of youth. Psychiatry Res. 2016, 244, 103–108. [Google Scholar] [CrossRef]


{kind=link}
| Variables | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1. Age | 1 | |||||||||
| 2. Father’s education level | −0.23 *** | 1 | ||||||||
| 3. Mother’s education level | −0.24 *** | 0.46 *** | 1 | |||||||
| 4. Household income | −0.13 * | 0.38 *** | 0.34 *** | 1 | ||||||
| 5. Depressive symptoms | 0.06 | −0.11 | −0.10 | −0.10 | 1 | |||||
| 6. MAOA T941G | 0.02 | −0.00 | 0.05 | −0.10 | −0.04 | 1 | ||||
| 7. COMT Val158Met | −0.16 * | 0.06 | 0.11 | 0.07 | 0.04 | −0.01 | 1 | |||
| 8. Abuse | −0.01 | −0.07 | −0.02 | −0.07 | 0.47 *** | −0.01 | −0.13 | 1 | ||
| 9. Neglect | 0.16 ** | −0.14 * | −0.09 | −0.11 | 0.35 *** | 0.08 | −0.11 | 0.42 *** | 1 | |
| 10. NSSI | 0.04 | −0.09 | −0.03 | −0.04 | 0.38 *** | 0.01 | −0.01 | 0.54 *** | 0.18 ** | 1 |
| M | 14.72 | − | − | − | 38.71 | − | − | 20.20 | 17.73 | 4.37 |
| SD | 0.92 | − | − | − | 9.84 | − | − | 6.15 | 6.44 | 8.66 |
| Predictors | ΔR2 | β | p | p(i) |
|---|---|---|---|---|
| Model 1 (Abuse) | ||||
| Step 1: Age | 0.18 | −0.03 | 0.574 | 0.050 |
| Depressive symptoms | 0.17 | 0.010 | 0.022 | |
| Step 2: Abuse | 0.19 | 0.11 | 0.383 | 0.044 |
| MAOA | 0.08 | 0.346 | 0.033 | |
| COMT | 0.08 | 0.377 | 0.039 | |
| Step 3: Abuse × MAOA | 0.02 | 0.35 | 0.003 | 0.017 |
| Abuse × COMT | 0.42 | <0.001 | 0.006 | |
| MAOA × COMT | −0.10 | 0.331 | 0.028 | |
| Step 4: Abuse × MAOA × COMT | 0.03 | −0.34 | 0.002 | 0.011 |
| Model 2 (Neglect) | ||||
| Step 1: Age | 0.18 | −0.01 | 0.936 | 0.050 |
| Depressive symptoms | 0.43 | <0.001 | 0.006 | |
| Step 2: Neglect | 0.00 | −0.13 | 0.302 | 0.033 |
| MAOA | 0.12 | 0.230 | 0.022 | |
| COMT | 0.05 | 0.635 | 0.044 | |
| Step 3: Neglect × MAOA | 0.01 | 0.21 | 0.103 | 0.011 |
| Neglect × COMT | 0.08 | 0.554 | 0.039 | |
| MAOA × COMT | −0.14 | 0.264 | 0.028 | |
| Step 4: Neglect × MAOA × COMT | 0.01 | −0.18 | 0.182 | 0.017 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Xiong, Y.; Liu, X.; Wang, H. The Effects of Childhood Maltreatment on Non-Suicidal Self-Injury in Male Adolescents: The Moderating Roles of the Monoamine Oxidase A (MAOA) Gene and the Catechol-O-Methyltransferase (COMT) Gene. Int. J. Environ. Res. Public Health 2021, 18, 2598. https://doi.org/10.3390/ijerph18052598
Gao Y, Xiong Y, Liu X, Wang H. The Effects of Childhood Maltreatment on Non-Suicidal Self-Injury in Male Adolescents: The Moderating Roles of the Monoamine Oxidase A (MAOA) Gene and the Catechol-O-Methyltransferase (COMT) Gene. International Journal of Environmental Research and Public Health. 2021; 18(5):2598. https://doi.org/10.3390/ijerph18052598
Chicago/Turabian StyleGao, Yemiao, Yuke Xiong, Xia Liu, and Hui Wang. 2021. "The Effects of Childhood Maltreatment on Non-Suicidal Self-Injury in Male Adolescents: The Moderating Roles of the Monoamine Oxidase A (MAOA) Gene and the Catechol-O-Methyltransferase (COMT) Gene" International Journal of Environmental Research and Public Health 18, no. 5: 2598. https://doi.org/10.3390/ijerph18052598