Management Options for Ixodes ricinus-Associated Pathogens: A Review of Prevention Strategies

 , and
, and 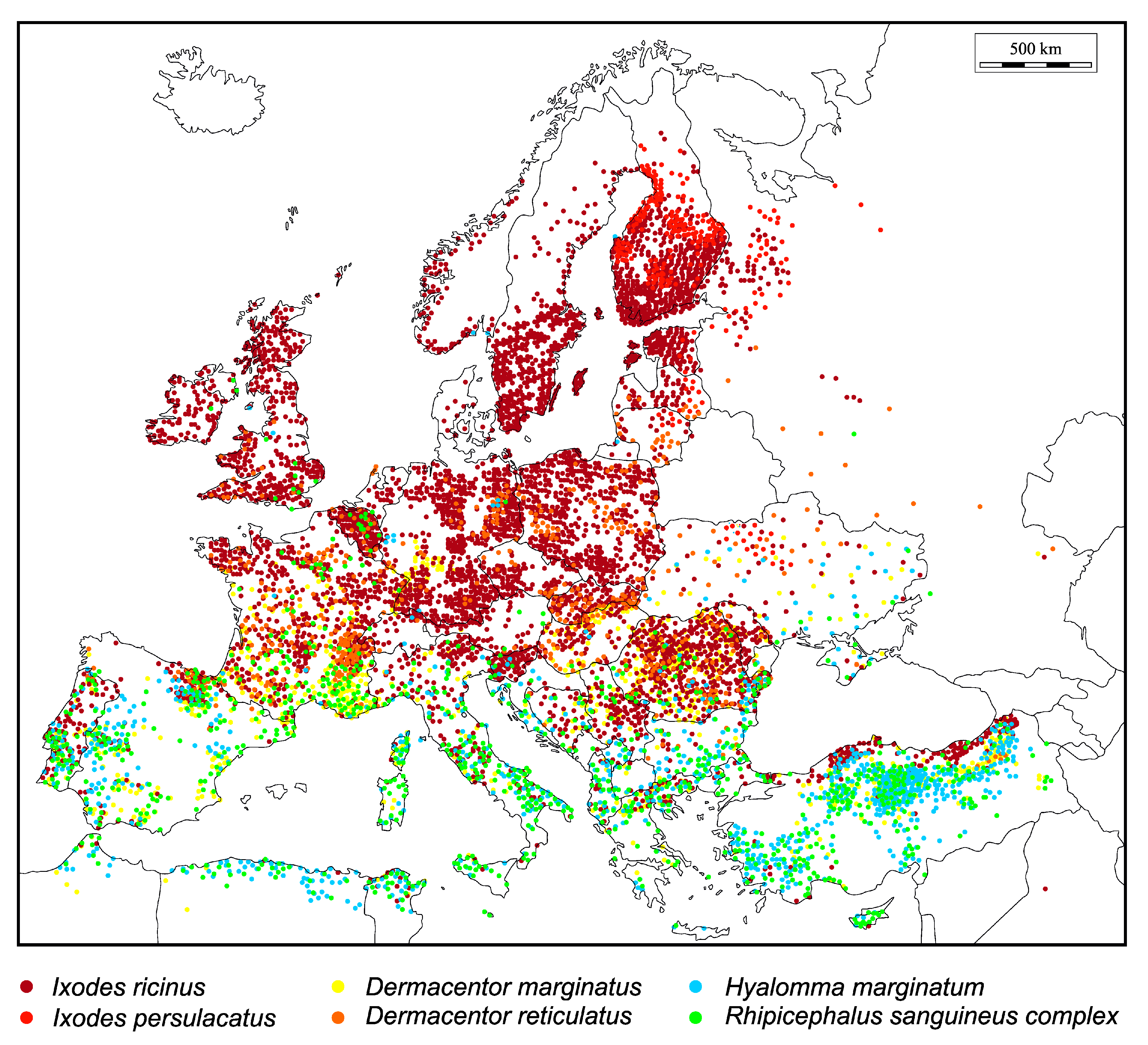{kind=link}
{kind=link}
Abstract
:1. Ecological Drivers Affecting Tick Activity
2. Anthropogenic Factors Affecting Tick Activity and Abundance of Tick-borne Diseases
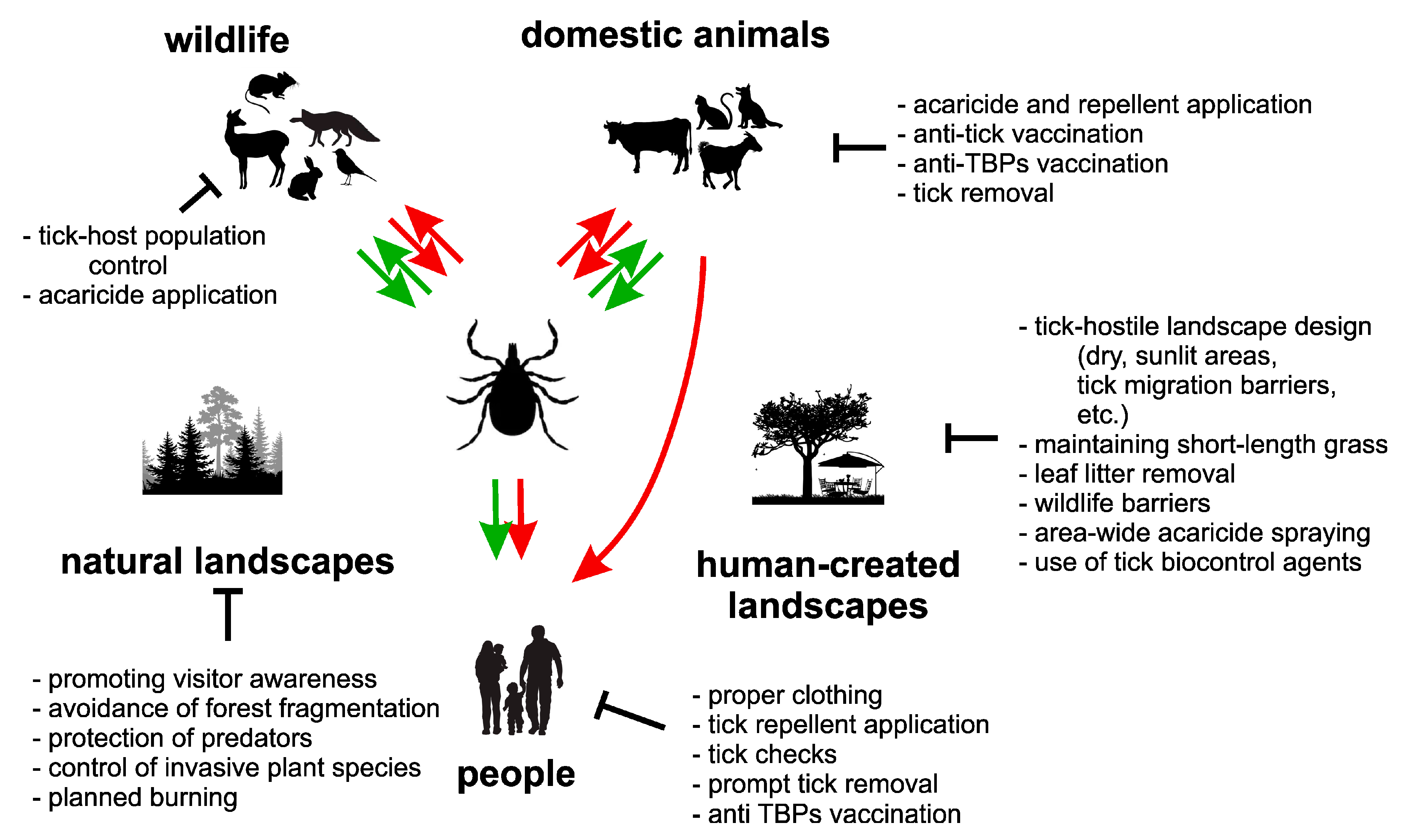
3. Tick Management Actions
3.1. Personal Protective Measures
3.2. Vaccination, Prophylaxis and Treatment
3.3. Tick Host Management
3.4. Landscape Management
3.5. Acaricides
3.6. Biological Agents
4. Tick Management in the USA in Comparison to Europe
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North. Africa—A Guide to Species Identification; Springer International Publishing: Cham, Switzerland, 2017; pp. XXI, 404. [Google Scholar]
- Beati, L.; Klompen, H. Phylogeography of Ticks (Acari: Ixodida). Annu. Rev. Entomol. 2019, 64, 379–397. [Google Scholar] [CrossRef]
- eCDC. Surveillance and disease data—Tick maps. Available online: https://www.ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/tick-maps (accessed on 10 October 2019).
- Abdullah, S.; Helps, C.; Tasker, S.; Newbury, H.; Wall, R. Ticks infesting domestic dogs in the UK: A large-scale surveillance programme. Parasit Vectors 2016, 9, 391. [Google Scholar] [CrossRef] [Green Version]
- Sprong, H.; Azagi, T.; Hoornstra, D.; Nijhof, A.M.; Knorr, S.; Baarsma, M.E.; Hovius, J.W. Control of Lyme borreliosis and other Ixodes ricinus-borne diseases. Parasit Vectors 2018, 11, 145. [Google Scholar] [CrossRef] [Green Version]
- Parola, P.; Raoult, D. Tick-borne bacterial diseases emerging in Europe. Clin. Microbiol. Infect. 2001, 7, 80–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barzon, L. Ongoing and emerging arbovirus threats in Europe. J. Clin. Virol. 2018, 107, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Karbowiak, G.; Biernat, B. The role of particular tick developmental stages in the circulation of tick-borne pathogens affecting humans in Central Europe. 2. Tick-borne encephalitis virus. Ann. Parasitol. 2016, 62, 3–9. [Google Scholar] [CrossRef]
- Gonzalez, J.P.; Camicas, J.L.; Cornet, J.P.; Faye, O.; Wilson, M.L. Sexual and transovarian transmission of Crimean-Congo haemorrhagic fever virus in Hyalomma truncatum ticks. Res. Virol. 1992, 143, 23–28. [Google Scholar] [CrossRef]
- Plowright, W.; Perry, C.T.; Greig, A. Sexual Transmission of African Swine Fever Virus in the Tick, Ornithodoros Moubata Porcinus, Walton. Res. Vet. Sci. 1974, 17, 106–113. [Google Scholar] [CrossRef]
- Hoby, S.; Mathis, A.; Doherr, M.G.; Robert, N.; Ryser-Degiorgis, M.P. Babesia capreoli infections in alpine chamois (Rupicapra r. Rupicapra), roe deer (Capreolus c. Capreolus) and red deer (Cervus elaphus) from Switzerland. J. Wildl. Dis. 2009, 45, 748–753. [Google Scholar] [CrossRef] [Green Version]
- Daniel, M.; Materna, J.; Honig, V.; Metelka, L.; Danielová, V.; Harcarik, J.; Kliegrová, S.; Grubhoffer, L. Vertical distribution of the tick Ixodes ricinus and tick-borne pathogens in the northern Moravian mountains correlated with climate warming (Jeseníky Mts., Czech Republic). Cent. Eur. J. Public Health 2009, 17, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Daniel, M.; Danielová, V.; Kříž, B.; Růžek, D.; Fialová, A.; Malý, M.; Materna, J.; Pejčoch, M.; Erhart, J. The occurrence of Ixodes ricinus ticks and important tick-borne pathogens in areas with high tick-borne encephalitis prevalence in different altitudinal levels of the Czech Republic Part I. Ixodes ricinus ticks and tick-borne encephalitis virus. Epidemiol. Mikrobiol. Imunol. 2016, 65, 118–128. [Google Scholar] [PubMed]
- Danielová, V.; Daniel, M.; Schwarzová, L.; Materna, J.; Rudenko, N.; Golovchenko, M.; Holubová, J.; Grubhoffer, L.; Kilián, P. Integration of a tick-borne encephalitis virus and Borrelia burgdorferi sensu lato into mountain ecosystems, following a shift in the altitudinal limit of distribution of their vector, Ixodes ricinus (Krkonose mountains, Czech Republic). Vector Borne Zoonotic Dis. 2010, 10, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Stünzner, D.; Hubálek, Z.; Halouzka, J.; Wendelin, I.; Sixl, W.; Marth, E. Prevalence of Borrelia burgdorferi sensu lato in the tick Ixodes ricinus in the Styrian mountains of Austria. Wien. Klin Wochenschr 2006, 118, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Daniel, M.; Rudenko, N.; Golovchenko, M.; Danielová, V.; Fialová, A.; Kříž, B.; Malý, M. The occurrence of Ixodes ricinus ticks and important tick-borne pathogens in areas with high tick-borne encephalitis prevalence in different altitudinal levels of the Czech Republic Part II. Ixodes ricinus ticks and genospecies of Borrelia burgdorferi sensu lato complex. Epidemiol. Mikrobiol. Imunol. 2016, 65, 182–192. [Google Scholar] [PubMed]
- Soleng, A.; Edgar, K.S.; Paulsen, K.M.; Pedersen, B.N.; Okbaldet, Y.B.; Skjetne, I.E.B.; Gurung, D.; Vikse, R.; Andreassen, Å. Distribution of Ixodes ricinus ticks and prevalence of tick-borne encephalitis virus among questing ticks in the Arctic Circle region of northern Norway. Ticks Tick Borne Dis. 2018, 9, 97–103. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Brunner, J.L. Climate change and Ixodes tick-borne diseases of humans. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [Green Version]
- Semenza, J.C.; Suk, J.E. Vector-borne diseases and climate change: A European perspective. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Stone, B.L.; Tourand, Y.; Brissette, C.A. Brave New Worlds: The Expanding Universe of Lyme Disease. Vector Borne Zoonotic Dis. 2017, 17, 619–629. [Google Scholar] [CrossRef]
- Cayol, C.; Koskela, E.; Mappes, T.; Siukkola, A.; Kallio, E.R. Temporal dynamics of the tick Ixodes ricinus in northern Europe: Epidemiological implications. Parasit Vectors 2017, 10, 166. [Google Scholar] [CrossRef] [Green Version]
- Boeckmann, M.; Joyner, T.A. Old health risks in new places? An ecological niche model for I. ricinus tick distribution in Europe under a changing climate. Health Place 2014, 30, 70–77. [Google Scholar] [CrossRef]
- Ehrmann, S.; Liira, J.; Gärtner, S.; Hansen, K.; Brunet, J.; Cousins, S.A.O.; Deconchat, M.; Decocq, G.; De Frenne, P.; De Smedt, P.; et al. Environmental drivers of Ixodes ricinus abundance in forest fragments of rural European landscapes. BMC Ecol. 2017, 17, 31. [Google Scholar] [CrossRef] [PubMed]
- Alkishe, A.A.; Peterson, A.T.; Samy, A.M. Climate change influences on the potential geographic distribution of the disease vector tick Ixodes ricinus. PLoS ONE 2017, 12, e0189092. [Google Scholar] [CrossRef] [PubMed]
- Boehnke, D.; Gebhardt, R.; Petney, T.; Norra, S. On the complexity of measuring forests microclimate and interpreting its relevance in habitat ecology: The example of Ixodes ricinus ticks. Parasit Vectors 2017, 10, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furness, R.W.; Furness, E.N. Ixodes ricinus parasitism of birds increases at higher winter temperatures. J. Vector Ecol. 2018, 43, 59–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollack, K.; Sodoudi, S.; Névir, P.; Müller, K.; Richter, D. Influence of meteorological parameters during the preceding fall and winter on the questing activity of nymphal Ixodes ricinus ticks. Int. J. Biometeorol. 2017, 61, 1787–1795. [Google Scholar] [CrossRef]
- Kiewra, D.; Sobczyñski, M. Biometrical analysis of the common tick, Ixodes ricinus, in the Sleza Massif (Lower Silesia, Poland). J. Vector Ecol. 2006, 31, 239–244. [Google Scholar] [CrossRef]
- Daniel, M.; Kolár, J.; Zeman, P.; Pavelka, K.; Sádlo, J. Predictive map of Ixodes ricinus high-incidence habitats and a tick-borne encephalitis risk assessment using satellite data. Exp. Appl. Acarol. 1998, 22, 417–433. [Google Scholar] [CrossRef]
- Cat, J.; Beugnet, F.; Hoch, T.; Jongejan, F.; Prangé, A.; Chalvet-Monfray, K. Influence of the spatial heterogeneity in tick abundance in the modeling of the seasonal activity of Ixodes ricinus nymphs in Western Europe. Exp. Appl. Acarol. 2017, 71, 115–130. [Google Scholar] [CrossRef]
- Schulz, M.; Mahling, M.; Pfister, K. Abundance and seasonal activity of questing Ixodes ricinus ticks in their natural habitats in southern Germany in 2011. J. Vector Ecol. 2014, 39, 56–65. [Google Scholar] [CrossRef]
- Maaz, D.; Krücken, J.; Blümke, J.; Richter, D.; McKay-Demeler, J.; Matuschka, F.R.; Hartmann, S.; von Samson-Himmelstjerna, G. Factors associated with diversity, quantity and zoonotic potential of ectoparasites on urban mice and voles. PLoS ONE 2018, 13, e0199385. [Google Scholar] [CrossRef]
- Kriz, B.; Daniel, M.; Benes, C.; Maly, M. The role of game (wild boar and roe deer) in the spread of tick-borne encephalitis in the Czech Republic. Vector Borne Zoonotic Dis. 2014, 14, 801–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honig, V.; Carolan, H.E.; Vavruskova, Z.; Massire, C.; Mosel, M.R.; Crowder, C.D.; Rounds, M.A.; Ecker, D.J.; Ruzek, D.; Grubhoffer, L.; et al. Broad-range survey of vector-borne pathogens and tick host identification of Ixodes ricinus from Southern Czech Republic. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Qviller, L.; Viljugrein, H.; Loe, L.E.; Meisingset, E.L.; Mysterud, A. The influence of red deer space use on the distribution of Ixodes ricinus ticks in the landscape. Parasit Vectors 2016, 9, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labbé Sandelin, L.; Tolf, C.; Larsson, S.; Wilhelmsson, P.; Salaneck, E.; Jaenson, T.G.; Lindgren, P.E.; Olsen, B.; Waldenström, J. Candidatus Neoehrlichia mikurensis in Ticks from Migrating Birds in Sweden. PLoS ONE 2015, 10, e0133250. [Google Scholar] [CrossRef] [PubMed]
- Pajoro, M.; Pistone, D.; Varotto Boccazzi, I.; Mereghetti, V.; Bandi, C.; Fabbi, M.; Scattorin, F.; Sassera, D.; Montagna, M. Molecular screening for bacterial pathogens in ticks (Ixodes ricinus) collected on migratory birds captured in northern Italy. Folia Parasitol. (Praha) 2018, 65. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, A.; Rosà, R. Effect of host populations on the intensity of ticks and the prevalence of tick-borne pathogens: How to interpret the results of deer exclosure experiments. Parasitology 2008, 135, 1531–1544. [Google Scholar] [CrossRef]
- Slovák, M.; Kazimírová, M.; Siebenstichová, M.; Ustaníková, K.; Klempa, B.; Gritsun, T.; Gould, E.A.; Nuttall, P.A. Survival dynamics of tick-borne encephalitis virus in Ixodes ricinus ticks. Ticks Tick Borne Dis. 2014, 5, 962–969. [Google Scholar] [CrossRef]
- Achazi, K.; Růžek, D.; Donoso-Mantke, O.; Schlegel, M.; Ali, H.S.; Wenk, M.; Schmidt-Chanasit, J.; Ohlmeyer, L.; Rühe, F.; Vor, T.; et al. Rodents as sentinels for the prevalence of tick-borne encephalitis virus. Vector Borne Zoonotic Dis. 2011, 11, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Kurtenbach, K.; De Michelis, S.; Etti, S.; Schäfer, S.M.; Sewell, H.S.; Brade, V.; Kraiczy, P. Host association of Borrelia burgdorferi sensu lato--the key role of host complement. Trends Microbiol. 2002, 10, 74–79. [Google Scholar] [CrossRef]
- Kurtenbach, K.; Sewell, H.S.; Ogden, N.H.; Randolph, S.E.; Nuttall, P.A. Serum complement sensitivity as a key factor in Lyme disease ecology. Infect. Immun. 1998, 66, 1248–1251. [Google Scholar] [CrossRef] [Green Version]
- Kuo, M.M.; Lane, R.S.; Giclas, P.C. A comparative study of mammalian and reptilian alternative pathway of complement-mediated killing of the Lyme disease spirochete (Borrelia burgdorferi). J. Parasitol. 2000, 86, 1223–1228. [Google Scholar] [CrossRef]
- Lane, R.S.; Quistad, G.B. Borreliacidal factor in the blood of the western fence lizard (Sceloporus occidentalis). J. Parasitol. 1998, 84, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Hönig, V.; Svec, P.; Halas, P.; Vavruskova, Z.; Tykalova, H.; Kilian, P.; Vetiskova, V.; Dornakova, V.; Sterbova, J.; Simonova, Z.; et al. Ticks and tick-borne pathogens in South Bohemia (Czech Republic)--Spatial variability in Ixodes ricinus abundance, Borrelia burgdorferi and tick-borne encephalitis virus prevalence. Ticks Tick Borne Dis. 2015, 6, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Hubálek, Z.; Halouzka, J.; Juricová, Z. A simple method of transmission risk assessment in enzootic foci of Lyme borreliosis. Eur. J. Epidemiol. 1996, 12, 331–333. [Google Scholar] [CrossRef]
- L’Hostis, M.; Dumon, H.; Dorchies, B.; Boisdron, F.; Gorenflot, A. Seasonal incidence and ecology of the tick Ixodes ricinus (Acari: Ixodidae) on grazing pastures in western France. Exp. Appl. Acarol. 1995, 19, 211–220. [Google Scholar] [CrossRef]
- MILNE, A. The ecology of the sheep tick, Ixodes ricinus L.; host relationships of the tick; observations on hill and moorland grazings in northern England. Parasitology 1949, 39, 173–197. [Google Scholar] [CrossRef]
- Sumilo, D.; Bormane, A.; Asokliene, L.; Vasilenko, V.; Golovljova, I.; Avsic-Zupanc, T.; Hubalek, Z.; Randolph, S.E. Socio-economic factors in the differential upsurge of tick-borne encephalitis in Central and Eastern Europe. Rev. Med. Virol. 2008, 18, 81–95. [Google Scholar] [CrossRef]
- Gregory, R.D.; van Strien, A.; Vorisek, P.; Gmelig Meyling, A.W.; Noble, D.G.; Foppen, R.P.; Gibbons, D.W. Developing indicators for European birds. Philos Trans. R Soc. Lond. B Biol. Sci. 2005, 360, 269–288. [Google Scholar] [CrossRef] [Green Version]
- Simmons, B.I.; Balmford, A.; Bladon, A.J.; Christie, A.P.; De Palma, A.; Dicks, L.V.; Gallego-Zamorano, J.; Johnston, A.; Martin, P.A.; Purvis, A.; et al. Worldwide insect declines: An important message, but interpret with caution. Ecol. Evol. 2019, 9, 3678–3680. [Google Scholar] [CrossRef] [Green Version]
- Rizzoli, A.; Hauffe, H.; Carpi, G.; Vourc H, G.; Neteler, M.; Rosa, R. Lyme borreliosis in Europe. Euro Surveill 2011, 16. [Google Scholar]
- Randolph, S.E.; Asokliene, L.; Avsic-Zupanc, T.; Bormane, A.; Burri, C.; Gern, L.; Golovljova, I.; Hubalek, Z.; Knap, N.; Kondrusik, M.; et al. Variable spikes in tick-borne encephalitis incidence in 2006 independent of variable tick abundance but related to weather. Parasit Vectors 2008, 1, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randolph, S.E.; team, E.-T.S.-P. Human activities predominate in determining changing incidence of tick-borne encephalitis in Europe. Euro Surveill 2010, 15, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonenshine, D.E. Range Expansion of Tick Disease Vectors in North America: Implications for Spread of Tick-Borne Disease. Int. J. Environ. Res. Public Health 2018, 15, 478. [Google Scholar] [CrossRef] [Green Version]
- Due, C.; Fox, W.; Medlock, J.M.; Pietzsch, M.; Logan, J.G. Tick bite prevention and tick removal. BMJ 2013, 347, f7123. [Google Scholar] [CrossRef] [Green Version]
- Kjær, L.J.; Soleng, A.; Edgar, K.S.; Lindstedt, H.E.H.; Paulsen, K.M.; Andreassen, Å.; Korslund, L.; Kjelland, V.; Slettan, A.; Stuen, S.; et al. Predicting and mapping human risk of exposure to. Euro Surveill 2019, 24. [Google Scholar] [CrossRef] [Green Version]
- Kiewra, D.; Stefańska-Krzaczek, E.; Szymanowski, M.; Szczepańska, A. Local-scale spatio-temporal distribution of questing Ixodes ricinus L. (Acari: Ixodidae)-A case study from a riparian urban forest in Wrocław, SW Poland. Ticks Tick Borne Dis. 2017, 8, 362–369. [Google Scholar] [CrossRef]
- Slunge, D.; Boman, A. Learning to live with ticks? The role of exposure and risk perceptions in protective behaviour against tick-borne diseases. PLoS ONE 2018, 13, e0198286. [Google Scholar] [CrossRef]
- Pages, F.; Dautel, H.; Duvallet, G.; Kahl, O.; de Gentile, L.; Boulanger, N. Tick repellents for human use: Prevention of tick bites and tick-borne diseases. Vector Borne Zoonotic Dis. 2014, 14, 85–93. [Google Scholar] [CrossRef]
- Eisen, L.; Rose, D.; Prose, R.; Breuner, N.E.; Dolan, M.C.; Thompson, K.; Connally, N. Bioassays to evaluate non-contact spatial repellency, contact irritancy, and acute toxicity of permethrin-treated clothing against nymphal Ixodes scapularis ticks. Ticks Tick Borne Dis. 2017, 8, 837–849. [Google Scholar] [CrossRef]
- Cisak, E.; Wójcik-Fatla, A.; Zając, V.; Dutkiewicz, J. Repellents and acaricides as personal protection measures in the prevention of tick-borne diseases. Ann. Agric. Environ. Med. 2012, 19, 625–630. [Google Scholar]
- Boffetta, P.; Desai, V. Exposure to permethrin and cancer risk: A systematic review. Crit. Rev. Toxicol. 2018, 48, 433–442. [Google Scholar] [CrossRef] [PubMed]
- BRENNAN, J.M. Preliminary report on some organic materials as tick repellents and toxic agents. Public Health Rep. 1947, 62, 1162–1165. [Google Scholar] [CrossRef] [PubMed]
- Ashitani, T.; Garboui, S.S.; Schubert, F.; Vongsombath, C.; Liblikas, I.; Pålsson, K.; Borg-Karlson, A.K. Activity studies of sesquiterpene oxides and sulfides from the plant Hyptis suaveolens (Lamiaceae) and its repellency on Ixodes ricinus (Acari: Ixodidae). Exp. Appl. Acarol. 2015, 67, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Mawela, K.G.; Luseba, D.; Magano, S.; Eloff, J.N. Repellent properties of Rotheca glabrum plant extracts against adults of Rhipicephalus appendiculatus. BMC Vet. Res. 2019, 15, 122. [Google Scholar] [CrossRef]
- Jaenson, T.G.; Pålsson, K.; Borg-Karlson, A.K. Evaluation of extracts and oils of tick-repellent plants from Sweden. Med. Vet. Entomol. 2005, 19, 345–352. [Google Scholar] [CrossRef]
- Galisteo Pretel, A.; Pérez Del Pulgar, H.; Olmeda, A.S.; Gonzalez-Coloma, A.; Barrero, A.F.; Quílez Del Moral, J.F. Novel Insect Antifeedant and Ixodicidal Nootkatone Derivatives. Biomolecules 2019, 9, 742. [Google Scholar] [CrossRef] [Green Version]
- Bharadwaj, A.; Stafford, K.C.; Behle, R.W. Efficacy and environmental persistence of nootkatone for the control of the blacklegged tick (Acari: Ixodidae) in residential landscapes. J. Med. Entomol. 2012, 49, 1035–1044. [Google Scholar] [CrossRef]
- Pitches, D.W. Removal of ticks: A review of the literature. Euro Surveill 2006, 11, E060817.060814. [Google Scholar] [CrossRef]
- de la Fuente, J.; Contreras, M.; Estrada-Peña, A.; Cabezas-Cruz, A. Targeting a global health problem: Vaccine design and challenges for the control of tick-borne diseases. Vaccine 2017, 35, 5089–5094. [Google Scholar] [CrossRef]
- de Silva, A.M.; Fish, D.; Burkot, T.R.; Zhang, Y.; Fikrig, E. OspA antibodies inhibit the acquisition of Borrelia burgdorferi by Ixodes ticks. Infect. Immun. 1997, 65, 3146–3150. [Google Scholar] [CrossRef] [Green Version]
- Lymerix. Lack of demand kills Lyme disease vaccine. Nursing 2002, 32, 18. [Google Scholar]
- Littman, M.P.; Goldstein, R.E.; Labato, M.A.; Lappin, M.R.; Moore, G.E. ACVIM small animal consensus statement on Lyme disease in dogs: Diagnosis, treatment, and prevention. J. Vet. Intern. Med. 2006, 20, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Guarino, C.; Asbie, S.; Rohde, J.; Glaser, A.; Wagner, B. Vaccination of horses with Lyme vaccines for dogs induces short-lasting antibody responses. Vaccine 2017, 35, 4140–4147. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, F.D.; Miller, R.J.; Pérez de León, A.A. Cattle tick vaccines: Many candidate antigens, but will a commercially viable product emerge? Int. J. Parasitol 2012, 42, 421–427. [Google Scholar] [CrossRef]
- Willadsen, P.; Bird, P.; Cobon, G.S.; Hungerford, J. Commercialisation of a recombinant vaccine against Boophilus microplus. Parasitology 1995, 110, S43–S50. [Google Scholar] [CrossRef]
- Sutton, D.; Spry, C. One Dose of Doxycycline for the Prevention of Lyme Disease: A Review of Clinical Effectiveness and Guidelines; Canadian Agency for Drugs and Technologies in Health: Ottawa, ON, Canada, 2019. Available online: https://www.ncbi.nlm.nih.gov/books/NBK545493/ (accessed on 12 December 2019).
- Leenders, A.C. Single-dose doxycycline for the prevention of Lyme disease. N. Engl. J. Med. 2001, 345, 1349, author reply 1349–1350. [Google Scholar]
- Sanchez, E.; Vannier, E.; Wormser, G.P.; Hu, L.T. Diagnosis, Treatment, and Prevention of Lyme Disease, Human Granulocytic Anaplasmosis, and Babesiosis: A Review. JAMA 2016, 315, 1767–1777. [Google Scholar] [CrossRef]
- Torbahn, G.; Hofmann, H.; Rücker, G.; Bischoff, K.; Freitag, M.H.; Dersch, R.; Fingerle, V.; Motschall, E.; Meerpohl, J.J.; Schmucker, C. Efficacy and Safety of Antibiotic Therapy in Early Cutaneous Lyme Borreliosis: A Network Meta-analysis. JAMA Dermatol. 2018, 154, 1292–1303. [Google Scholar] [CrossRef]
- Vial, H.J.; Gorenflot, A. Chemotherapy against babesiosis. Vet. Parasitol. 2006, 138, 147–160. [Google Scholar] [CrossRef]
- Eyer, L.; Valdés, J.J.; Gil, V.A.; Nencka, R.; Hřebabecký, H.; Šála, M.; Salát, J.; Černý, J.; Palus, M.; De Clercq, E.; et al. Nucleoside inhibitors of tick-borne encephalitis virus. Antimicrob. Agents Chemother. 2015, 59, 5483–5493. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.; Mahaffy, J.; Vaidya, N.K. Modeling transmission dynamics of lyme disease: Multiple vectors, seasonality, and vector mobility. Infect. Dis Model. 2019, 4, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Foley, J.E.; Clark, K.L.; Anderson, J.F.; Durden, L.A.; Manord, J.M.; Smith, M.L. Established Population of Blacklegged Ticks with High Infection Prevalence for the Lyme Disease Bacterium. Int. J. Med. Sci. 2016, 13, 881–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, S.C.; Stafford, K.C.; Molaei, G.; Linske, M.A. Integrated Control of Nymphal Ixodes scapularis: Effectiveness of White-Tailed Deer Reduction, the Entomopathogenic Fungus Metarhizium anisopliae, and Fipronil-Based Rodent Bait Boxes. Vector Borne Zoonotic Dis. 2018, 18, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Hofmeester, T.R.; Sprong, H.; Jansen, P.A.; Prins, H.H.T.; van Wieren, S.E. Deer presence rather than abundance determines the population density of the sheep tick, Ixodes ricinus, in Dutch forests. Parasit Vectors 2017, 10, 433. [Google Scholar] [CrossRef]
- Gilbert, L.; Maffey, G.L.; Ramsay, S.L.; Hester, A.J. The effect of deer management on the abundance of Ixodes ricinus in Scotland. Ecol. Appl 2012, 22, 658–667. [Google Scholar] [CrossRef]
- Kugeler, K.J.; Jordan, R.A.; Schulze, T.L.; Griffith, K.S.; Mead, P.S. Will Culling White-Tailed Deer Prevent Lyme Disease? Zoonoses Public Health 2016, 63, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapron, G.; Kaczensky, P.; Linnell, J.D.; von Arx, M.; Huber, D.; Andrén, H.; López-Bao, J.V.; Adamec, M.; Álvares, F.; Anders, O.; et al. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 2014, 346, 1517–1519. [Google Scholar] [CrossRef] [Green Version]
- Daniels, T.J.; Fish, D.; Falco, R.C. Evaluation of host-targeted acaricide for reducing risk of Lyme disease in southern New York state. J. Med. Entomol. 1991, 28, 537–543. [Google Scholar] [CrossRef]
- Hofmeester, T.R.; Jansen, P.A.; Wijnen, H.J.; Coipan, E.C.; Fonville, M.; Prins, H.H.T.; Sprong, H.; van Wieren, S.E. Cascading effects of predator activity on tick-borne disease risk. Proc. Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [Green Version]
- Hoen, A.G.; Rollend, L.G.; Papero, M.A.; Carroll, J.F.; Daniels, T.J.; Mather, T.N.; Schulze, T.L.; Stafford, K.C.; Fish, D. Effects of tick control by acaricide self-treatment of white-tailed deer on host-seeking tick infection prevalence and entomologic risk for Ixodes scapularis-borne pathogens. Vector Borne Zoonotic Dis. 2009, 9, 431–438. [Google Scholar] [CrossRef] [Green Version]
- Wong, T.J.; Schramm, P.J.; Foster, E.; Hahn, M.B.; Schafrick, N.H.; Conlon, K.C.; Cameron, L. The Effectiveness and Implementation of 4-Poster Deer Self-Treatment Devices for Tick-borne Disease Prevention—A Potential Component of an Integrated Tick Management Program. Available online: https://www.cdc.gov/climateandhealth/docs/Effectiveness4PosterDevices_508.pdf (accessed on 2 February 2020).
- Baráková, I.; Derdáková, M.; Selyemová, D.; Chvostáč, M.; Špitalská, E.; Rosso, F.; Collini, M.; Rosà, R.; Tagliapietra, V.; Girardi, M.; et al. Tick-borne pathogens and their reservoir hosts in northern Italy. Ticks Tick Borne Dis. 2018, 9, 164–170. [Google Scholar] [CrossRef]
- Skotarczak, B. Canine borreliosis—Epidemiology and diagnostics. Ann. Agric. Environ. Med. 2002, 9, 137–140. [Google Scholar] [PubMed]
- Almazan, C.; Tipacamu, G.A.; Rodriguez, S.; Mosqueda, J.; Perez de Leon, A. Immunological control of ticks and tick-borne diseases that impact cattle health and production. Front. Biosci. (Landmark Ed.) 2018, 23, 1535–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, J.A.; Mather, T.N.; Tanner, P.; Gallo, G.; Jacobson, R.H. Efficacy of a nonadjuvanted, outer surface protein A, recombinant vaccine in dogs after challenge by ticks naturally infected with Borrelia burgdorferi. Vet. Ther. 2000, 1, 96–107. [Google Scholar] [PubMed]
- Geurden, T.; Becskei, C.; Grace, S.; Strube, C.; Doherty, P.; Liebenberg, J.; Mahabir, S.P.; Slootmans, N.; Lloyd, A.; Six, R.H. Efficacy of a novel oral formulation of sarolaner (Simparica™) against four common tick species infesting dogs in Europe. Vet. Parasitol. 2016, 222, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Cavalleri, D.; Murphy, M.; Gorbea, R.L.; Seewald, W.; Drake, J.; Nanchen, S. Laboratory evaluations of the immediate and sustained effectiveness of lotilaner (Credelio™) against three common species of ticks affecting dogs in Europe. Parasit Vectors 2017, 10, 527. [Google Scholar] [CrossRef] [Green Version]
- Dorko, E.; Hockicko, J.; Rimárová, K.; Bušová, A.; Popaďák, P.; Popaďáková, J.; Schréter, I. Milk outbreaks of tick-borne encephalitis in Slovakia, 2012–2016. Cent. Eur. J. Public Health 2018, 26, S47–S50. [Google Scholar] [CrossRef] [Green Version]
- Růžek, D.; Yakimenko, V.V.; Karan, L.S.; Tkachev, S.E. Omsk haemorrhagic fever. Lancet 2010, 376, 2104–2113. [Google Scholar] [CrossRef]
- Annen, K.; Friedman, K.; Eshoa, C.; Horowitz, M.; Gottschall, J.; Straus, T. Two cases of transfusion-transmitted Anaplasma phagocytophilum. Am. J. Clin. Pathol. 2012, 137, 562–565. [Google Scholar] [CrossRef] [Green Version]
- Ngo, V.; Civen, R. Babesiosis acquired through blood transfusion, California, USA. Emerg Infect. Dis. 2009, 15, 785–787. [Google Scholar] [CrossRef]
- Gleim, E.R.; Conner, L.M.; Berghaus, R.D.; Levin, M.L.; Zemtsova, G.E.; Yabsley, M.J. The phenology of ticks and the effects of long-term prescribed burning on tick population dynamics in southwestern Georgia and northwestern Florida. PLoS ONE 2014, 9, e112174. [Google Scholar] [CrossRef] [PubMed]
- Scasta, J.D. Fire and Parasites: An Under-Recognized Form of Anthropogenic Land Use Change and Mechanism of Disease Exposure. Ecohealth 2015, 12, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Stafford, K.C.; Ward, J.S.; Magnarelli, L.A. Impact of controlled burns on the abundance of Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 1998, 35, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Mather, T.N.; Duffy, D.C.; Campbell, S.R. An unexpected result from burning vegetation to reduce Lyme disease transmission risks. J. Med. Entomol. 1993, 30, 642–645. [Google Scholar] [CrossRef] [Green Version]
- Morlando, S.; Schmidt, S.J.; LoGiudice, K. Reduction in Lyme Disease Risk as an Economic Benefit of Habitat Restoration. Restoration Ecol. 2011. [Google Scholar] [CrossRef]
- Lerman, S.B.; D’Amico, V. Lawn mowing frequency in suburban areas has no detectable effect on Borrelia spp. vector Ixodes scapularis (Acari: Ixodidae). PLoS ONE 2019, 14, e0214615. [Google Scholar] [CrossRef] [Green Version]
- Stafford, K.C. Tick Management Handbook; An Integrated Guide for Homeowners, Pest Control Operators, and Public Health Officials for the Prevention of Tick-Associated Disease; Connecticut Agricultural Experiment Station: New Haven, CT, USA, 2004. Available online: https://stacks.cdc.gov/view/cdc/11444 (accessed on 12 December 2019).
- Hansford, K.M.; Fonville, M.; Gillingham, E.L.; Coipan, E.C.; Pietzsch, M.E.; Krawczyk, A.I.; Vaux, A.G.C.; Cull, B.; Sprong, H.; Medlock, J.M. Ticks and Borrelia in urban and peri-urban green space habitats in a city in southern England. Ticks Tick Borne Dis. 2017, 8, 353–361. [Google Scholar] [CrossRef]
- Kowalec, M.; Szewczyk, T.; Welc-Falęciak, R.; Siński, E.; Karbowiak, G.; Bajer, A. Ticks and the city—Are there any differences between city parks and natural forests in terms of tick abundance and prevalence of spirochaetes? Parasit Vectors 2017, 10, 573. [Google Scholar] [CrossRef]
- Winkel, K.T.; Ribeiro, P.B.; Antunes, L.O.; Cárcamo, M.C.; Vianna, E.E. Rhipicephalus sanguineus sensu lato (Ixodidae) in synantropic rodents in Rio Grande do Sul, Brazil. Rev. Bras. Parasitol. Vet. 2014, 23, 276–279. [Google Scholar] [CrossRef] [Green Version]
- Godfrey, E.R.; Randolph, S.E. Economic downturn results in tick-borne disease upsurge. Parasit. Vectors 2011, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Košnik, I.G.; Lah, A.K. A campaign to increase the vaccination rate in a highly endemic tick-borne encephalitis region of Slovenia. Vaccine 2013, 31, 732–734. [Google Scholar] [CrossRef] [PubMed]
- Bergmeier, E.; Petermann, J.; Schröder, E. Geobotanical survey of wood-pasture habitats in Europe: Diversity, threats and conservation. Biodiversity Conserv. 2010, 19, 2995–3014. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Vivas, R.I.; Jonsson, N.N.; Bhushan, C. Strategies for the control of Rhipicephalus microplus ticks in a world of conventional acaricide and macrocyclic lactone resistance. Parasitol. Res. 2018, 117, 3–29. [Google Scholar] [CrossRef] [Green Version]
- Banumathi, B.; Vaseeharan, B.; Rajasekar, P.; Prabhu, N.M.; Ramasamy, P.; Murugan, K.; Canale, A.; Benelli, G. Exploitation of chemical, herbal and nanoformulated acaricides to control the cattle tick, Rhipicephalus (Boophilus) microplus—A review. Vet. Parasitol. 2017, 244, 102–110. [Google Scholar] [CrossRef]
- Stafford, K.C. Pesticide use by licensed applicators for the control of Ixodes scapularis (Acari: Ixodidae) in Connecticut. J. Med. Entomol. 1997, 34, 552–558. [Google Scholar] [CrossRef]
- Petermann, J.; Cauquil, L.; Hurlin, J.C.; Gaia, H.; Hüe, T. Survey of cattle tick, Riphicephalus (Boophilus) microplus, resistance to amitraz and deltamethrin in New Caledonia. Vet. Parasitol. 2016, 217, 64–70. [Google Scholar] [CrossRef]
- Guerrero, F.D.; Lovis, L.; Martins, J.R. Acaricide resistance mechanisms in Rhipicephalus (Boophilus) microplus. Rev. Bras. Parasitol. Vet. 2012, 21, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Rajput, Z.I.; Hu, S.H.; Chen, W.J.; Arijo, A.G.; Xiao, C.W. Importance of ticks and their chemical and immunological control in livestock. J. Zhejiang Univ. Sci. B 2006, 7, 912–921. [Google Scholar] [CrossRef]
- Jordan, R.A.; Schulze, T.L. Availability and Nature of Commercial Tick Control Services in Three Lyme Disease Endemic States. J. Med. Entomol. 2019. [Google Scholar] [CrossRef]
- Kunz, S.E.; Kemp, D.H. Insecticides and acaricides: Resistance and environmental impact. Rev. Sci. Tech. 1994, 13, 1249–1286. [Google Scholar] [CrossRef]
- Sánchez-Bayoa, F.; Wyckhuysbcd, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Canale, A.; Mehlhorn, H. Tick repellents and acaricides of botanical origin: A green roadmap to control tick-borne diseases? Parasitol. Res. 2016, 115, 2545–2560. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L.; Dolan, M.C. Evidence for Personal Protective Measures to Reduce Human Contact With Blacklegged Ticks and for Environmentally Based Control Methods to Suppress Host-Seeking Blacklegged Ticks and Reduce Infection with Lyme Disease Spirochetes in Tick Vectors and Rodent Reservoirs. J. Med. Entomol. 2016, 53, 1063–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stafford, K.C.; Denicola, A.J.; Magnarelli, L.A. Presence of Ixodiphagus hookeri (Hymenoptera: Encyrtidae) in two Connecticut populations of Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 1996, 33, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Samish, M.; Glazer, I. Entomopathogenic nematodes for the biocontrol of ticks. Trends Parasitol. 2001, 17, 368–371. [Google Scholar] [CrossRef]
- Lipa, J.; Eilenberg, J.; Bresciani, J.; Frandsen, F. Some observations on a newly recorded mermithid parasite of Ixodes ricinus L. (Acarina: Ixodidae). Acta Parasitol. 1997, 42, 109–114. [Google Scholar]
- Vega, F.E. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Fernandes, E.K.; Bittencourt, V.R. Entomopathogenic fungi against South American tick species. Exp. Appl. Acarol. 2008, 46, 71–93. [Google Scholar] [CrossRef]
- Fernandes, É.; Bittencourt, V.R.; Roberts, D.W. Perspectives on the potential of entomopathogenic fungi in biological control of ticks. Exp. Parasitol. 2012, 130, 300–305. [Google Scholar] [CrossRef]
- Samish, M.; Alekseev, E. Arthropods as predators of ticks (Ixodoidea). J. Med. Entomol. 2001, 38, 1–11. [Google Scholar] [CrossRef]
- Samish, M.; Ginsberg, H.; Glazer, I. Biological control of ticks. Parasitology 2004, 129, S389–S403. [Google Scholar] [CrossRef] [PubMed]
- Nunn, C.L.; Ezenwa, V.O.; Arnold, C.; Koenig, W.D. Mutualism or parasitism? Using a phylogenetic approach to characterize the oxpecker-ungulate relationship. Evolution 2011, 65, 1297–1304. [Google Scholar] [CrossRef] [PubMed]
- Samish, M.; Rehacek, J. Pathogens and predators of ticks and their potential in biological control. Annu. Rev. Entomol. 1999, 44, 159–182. [Google Scholar] [CrossRef]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, V.V.S.; Navegantes-Lima, K.C.; de Lemos, A.B.; da Silva, G.L.; de Souza Gomes, R.; Reis, J.F.; Rodrigues Junior, L.C.; da Silva, O.S.; Romão, P.R.T.; Monteiro, M.C. -Chikungunya Virus Interaction: Key Role of Vector Midguts Microbiota and Its Saliva in the Host Infection. Front. Microbiol. 2019, 10, 492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegde, S.; Rasgon, J.L.; Hughes, G.L. The microbiome modulates arbovirus transmission in mosquitoes. Curr. Opin. Virol. 2015, 15, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terradas, G.; McGraw, E.A. Wolbachia-mediated virus blocking in the mosquito vector Aedes aegypti. Curr. Opin. Insect. Sci. 2017, 22, 37–44. [Google Scholar] [CrossRef]
- Zhang, X.; Norris, D.E.; Rasgon, J.L. Distribution and molecular characterization of Wolbachia endosymbionts and filarial nematodes in Maryland populations of the lone star tick (Amblyomma americanum). FEMS Microbiol. Ecol. 2011, 77, 50–56. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Hayes, S.F.; Mavros, A.J. Nonpathogenic rickettsiae in Dermacentor andersoni: A limiting factor for the distribution of Rickettsia rickettsii. In Rickettsiae and Rickettsial Diseases; Burgdorfer, W., Anacker, R.L., Eds.; Academic Press: New York, NY, USA, 1981. [Google Scholar]
- Macaluso, K.R.; Sonenshine, D.E.; Ceraul, S.M.; Azad, A.F. Rickettsial infection in Dermacentor variabilis (Acari: Ixodidae) inhibits transovarial transmission of a second Rickettsia. J. Med. Entomol. 2002, 39, 809–813. [Google Scholar] [CrossRef] [Green Version]
- Narasimhan, S.; Fikrig, E. Tick microbiome: The force within. Trends Parasitol. 2015, 31, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Flores, H.A.; O’Neill, S.L. Controlling vector-borne diseases by releasing modified mosquitoes. Nat. Rev. Microbiol. 2018, 16, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Mead, P.; Hook, S.; Niesobecki, S.; Ray, J.; Meek, J.; Delorey, M.; Prue, C.; Hinckley, A. Risk factors for tick exposure in suburban settings in the Northeastern United States. Ticks Tick Borne Dis. 2018, 9, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Connally, N.P.; Durante, A.J.; Yousey-Hindes, K.M.; Meek, J.I.; Nelson, R.S.; Heimer, R. Peridomestic Lyme disease prevention: Results of a population-based case-control study. Am. J. Prev. Med. 2009, 37, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Zintl, A.; Mulcahy, G.; Skerrett, H.E.; Taylor, S.M.; Gray, J.S. Babesia divergens, a bovine blood parasite of veterinary and zoonotic importance. Clin. Microbiol. Rev. 2003, 16, 622–636. [Google Scholar] [CrossRef] [Green Version]
- Mysterud, A.; Jore, S.; Østerås, O.; Viljugrein, H. Emergence of tick-borne diseases at northern latitudes in Europe: A comparative approach. Sci. Rep. 2017, 7, 16316. [Google Scholar] [CrossRef] [Green Version]
- CDC. Ticks. Available online: https://www.cdc.gov/ticks/index.html (accessed on 10 October 2019).
- Burtis, J.C.; Pflueger, C. Interactions between soil-dwelling arthropod predators and Ixodes scapularis under laboratory and field conditions. Ecosphere 2017, 8, e01914. [Google Scholar] [CrossRef]
- Fouet, C.; Kamdem, C. Integrated Mosquito Management: Is Precision Control a Luxury or Necessity? Trends Parasitol. 2019, 35, 85–95. [Google Scholar] [CrossRef]
- Benelli, G.; Beier, J.C. Current vector control challenges in the fight against malaria. Acta Trop. 2017, 174, 91–96. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Černý, J.; Lynn, G.; Hrnková, J.; Golovchenko, M.; Rudenko, N.; Grubhoffer, L. Management Options for Ixodes ricinus-Associated Pathogens: A Review of Prevention Strategies. Int. J. Environ. Res. Public Health 2020, 17, 1830. https://doi.org/10.3390/ijerph17061830
Černý J, Lynn G, Hrnková J, Golovchenko M, Rudenko N, Grubhoffer L. Management Options for Ixodes ricinus-Associated Pathogens: A Review of Prevention Strategies. International Journal of Environmental Research and Public Health. 2020; 17(6):1830. https://doi.org/10.3390/ijerph17061830
Chicago/Turabian StyleČerný, Jiří, Geoffrey Lynn, Johana Hrnková, Maryna Golovchenko, Natalia Rudenko, and Libor Grubhoffer. 2020. "Management Options for Ixodes ricinus-Associated Pathogens: A Review of Prevention Strategies" International Journal of Environmental Research and Public Health 17, no. 6: 1830. https://doi.org/10.3390/ijerph17061830