Remediation of Crude Oil-Polluted Soil by the Bacterial Rhizosphere Community of Suaeda Salsa Revealed by 16S rRNA Genes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Designs
2.2. Soil Sampling
2.3. Soil DNA Extraction, Polymerase Chain Reaction (PCR), and Illumina MiSeq Sequencing
2.4. Statistical Analyses
3. Results and Discussion
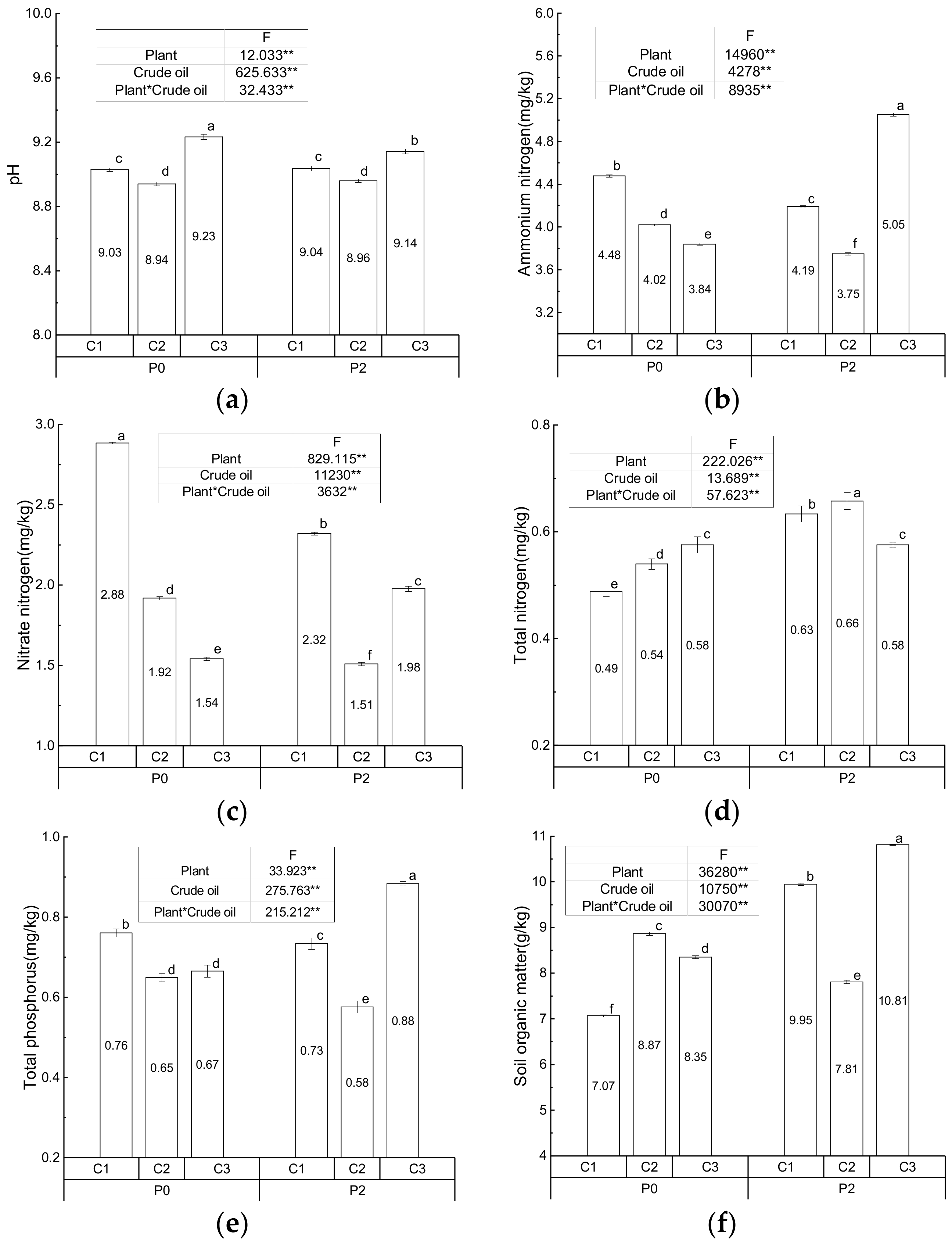
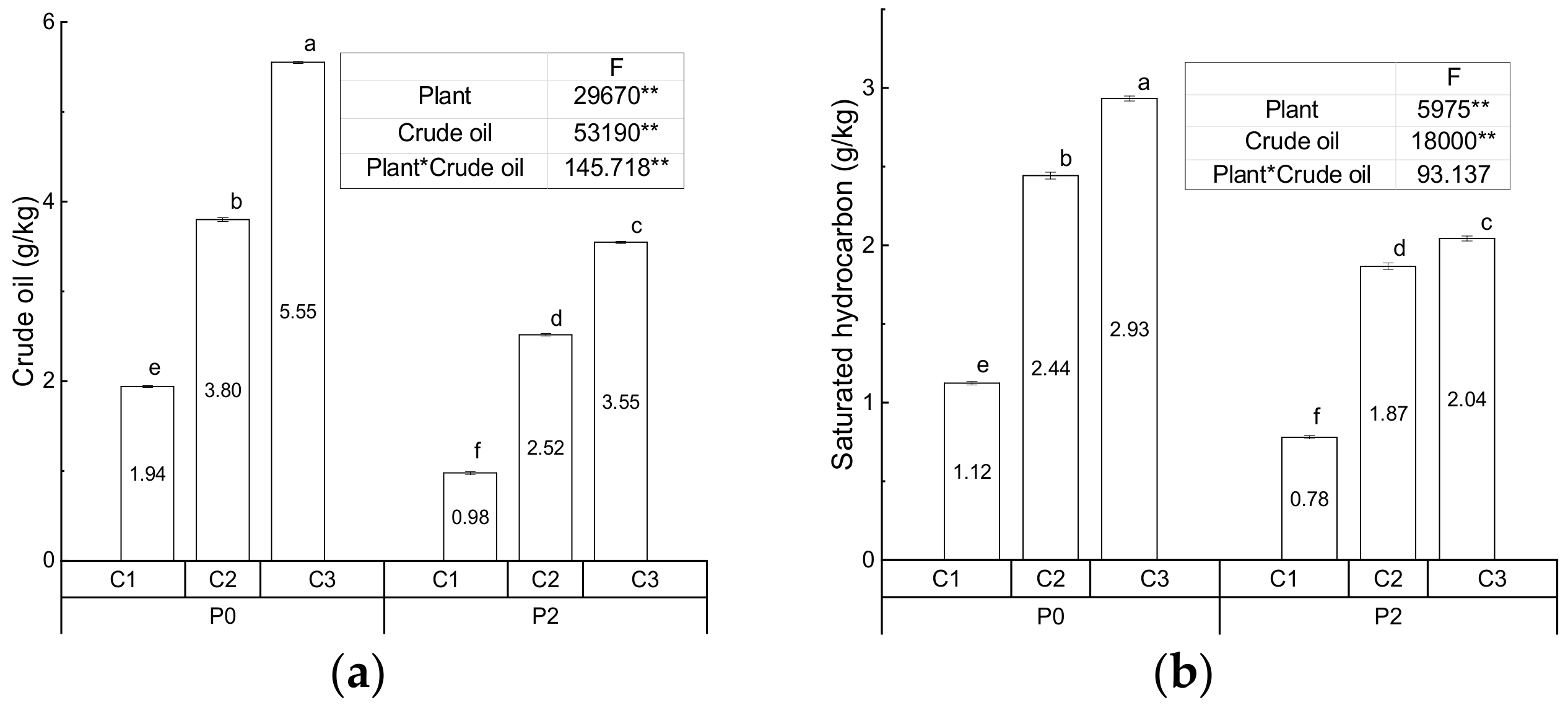
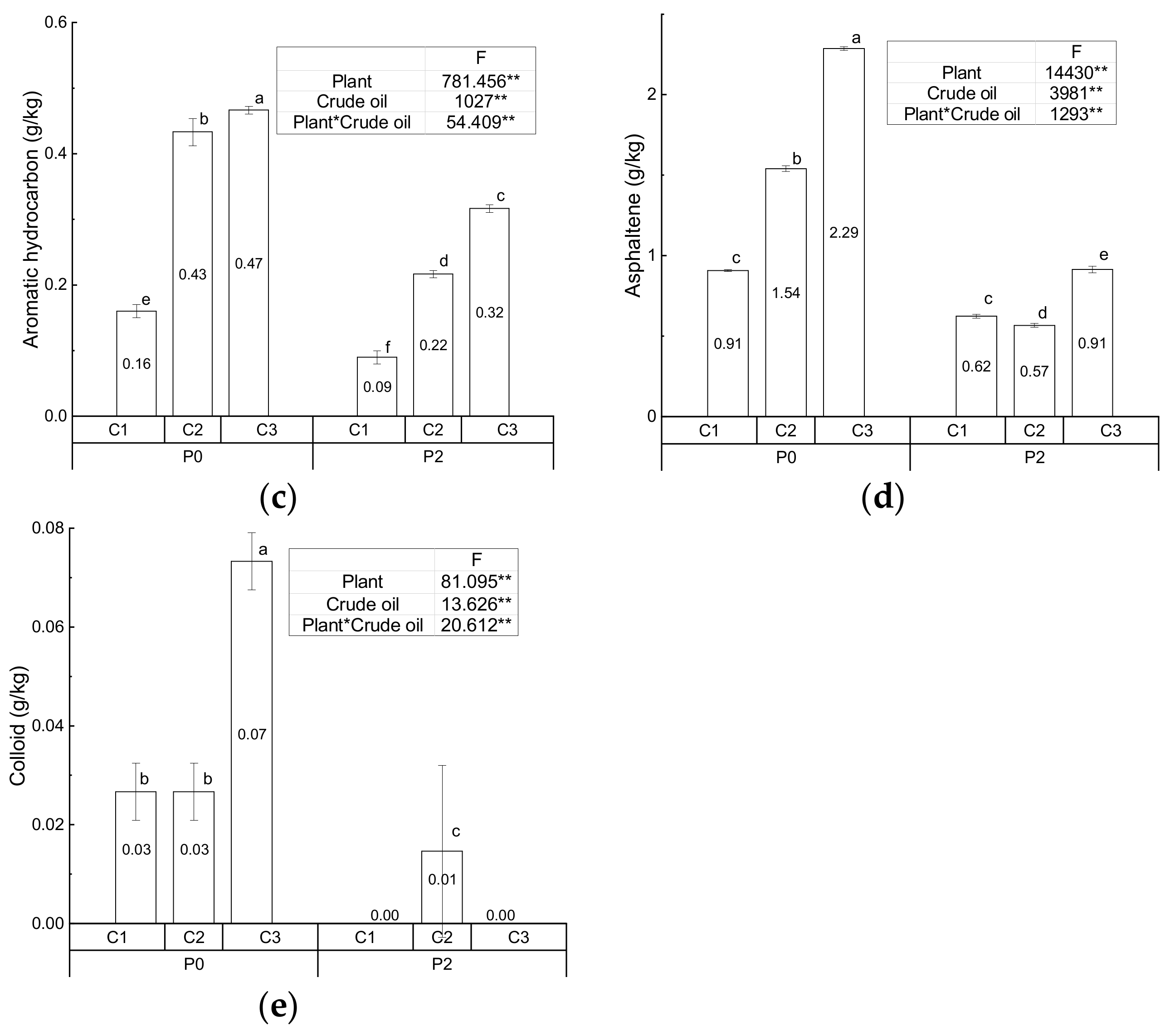
3.1. Physiochemical Characteristics of the Soil
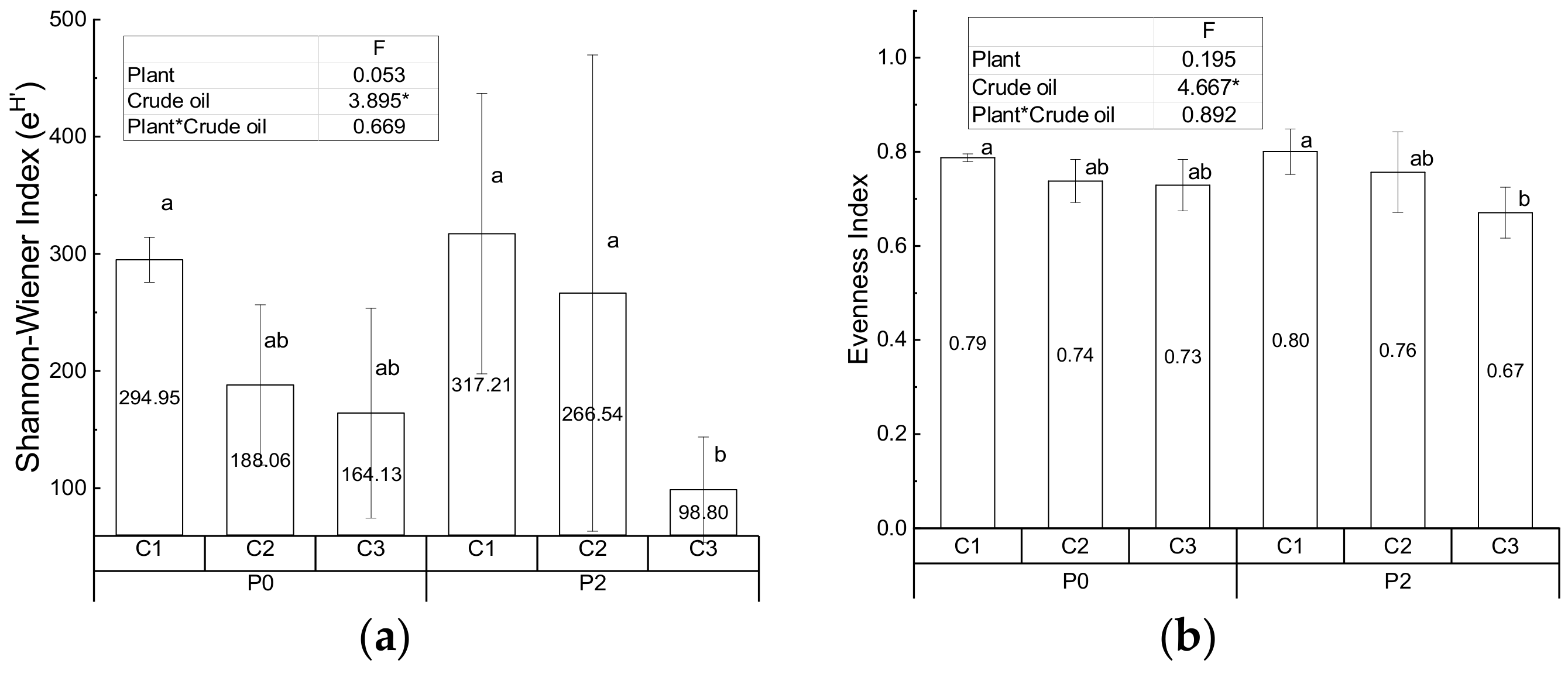
3.2. Diversity of the Bacterial Community
3.3. Taxonomic Compositions
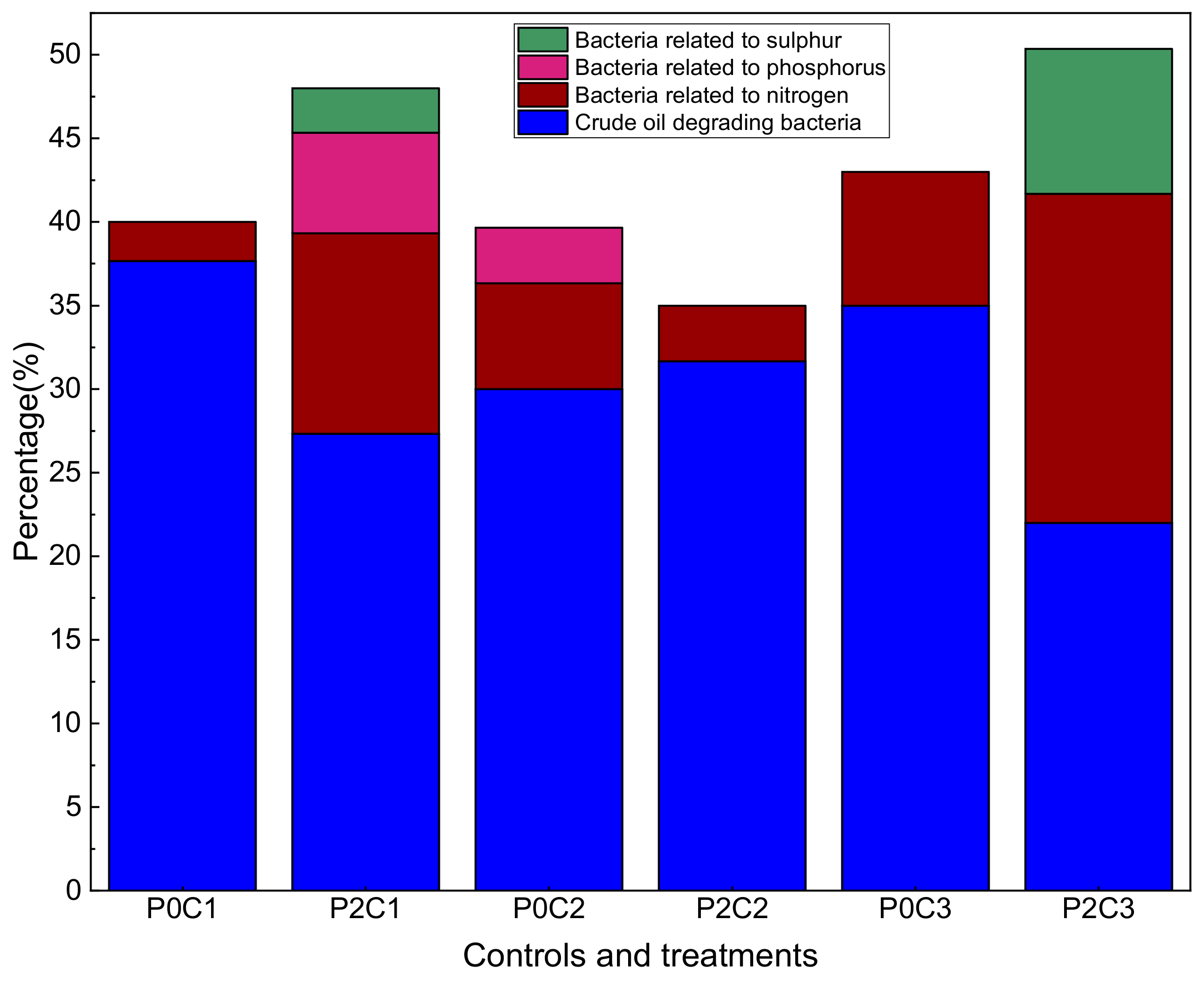
3.4. Functional Group Distribution
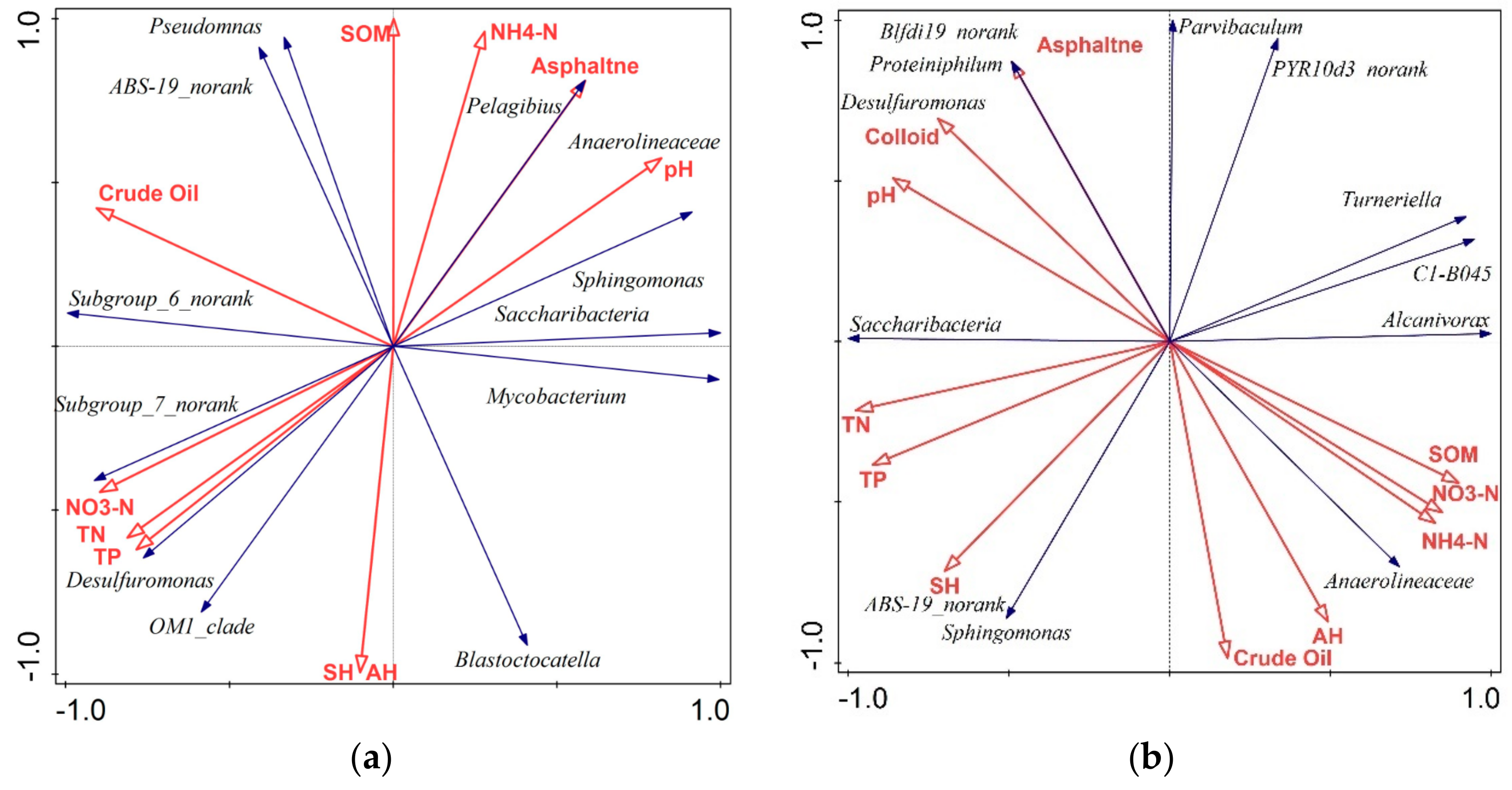
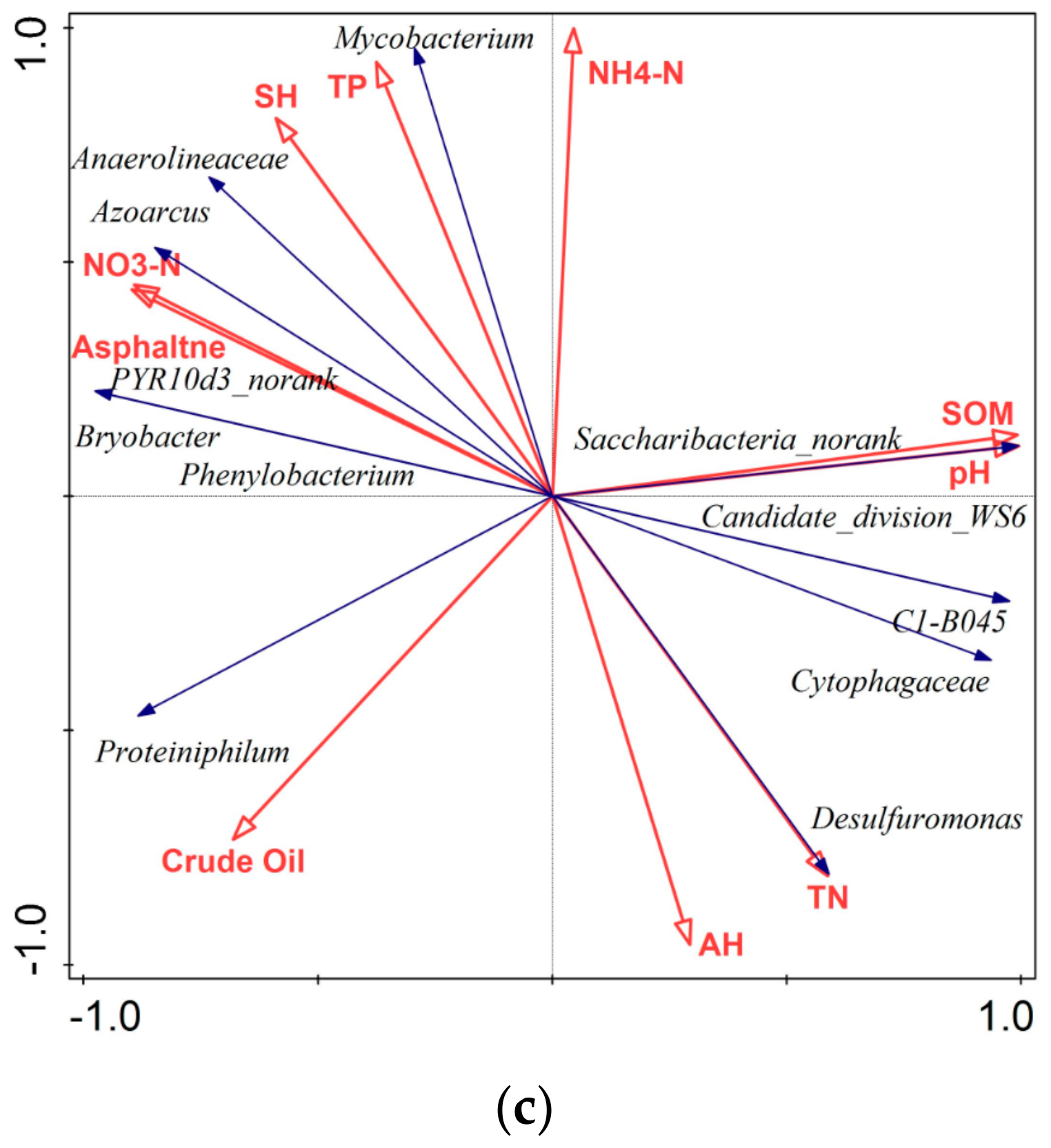
3.5. Relationship between Bacterial Groups and Soil Variables
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ian, S.; Riddle, M.J.; Stark, J.S.; Cole, C.M.; King, C.K.; Sabine, D.; Gore, D.B. Management and remediation of contaminated sites at Casey Station, Antarctica. Polar Rec. 2001, 37, 199–214. [Google Scholar]
- Wu, M.; Ye, X.; Chen, K.; Li, W.; Yuan, J.; Jiang, X. Bacterial community shift and hydrocarbon transformation during bioremediation of short-term petroleum-contaminated soil. Environ. Pollut. 2017, 223, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.F.; Yang, B.B.; Zhu, X.J. Study on the effect of different processing contaminated soil bioremediation technology. Biotechnol. Bull. 2016, 3, 68–72. (In Chinese) [Google Scholar]
- Zhang, D.; Ding, A.; Cui, S.; Hu, C.; Thornton, S.F.; Dou, J.; Sun, Y.; Huang, W.E. Whole cell bioreporter application for rapid detection and evaluation of crude oil spill in seawater caused by Dalian oil tank explosion. Water Res. 2013, 47, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Pokethitiyook, P. Phytoremediation of Petroleum-Contaminated Soil in Association with Soil Bacteria; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Sebiomo, A.; Bankole, S.A.; Awosanya, A.O. Determination of the ability of microorganisms isolated from mechanic soil to utilise lubricating oil as carbon source. Afr. J. Microbiol. Res. 2010, 4, 2257–2264. [Google Scholar]
- Janbandhu, A.; Fulekar, M.H. Biodegradation of phenanthrene using adapted microbial consortium isolated from petrochemical contaminated environment. J. Hazar. Mater. 2011, 187, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Prince, R.C.; Mcfarlin, K.M.; Butler, J.D.; Febbo, E.J.; Wang, F.C.; Nedwed, T.J. The primary biodegradation of dispersed crude oil in the sea. Chemosphere 2013, 90, 521–526. [Google Scholar] [CrossRef]
- Ebadi, A.; Khoshkholgh Sima, N.A.; Olamaee, M.; Hashemi, M.; Ghorbani, N.R. Remediation of saline soils contaminated with crude oil using the halophyte Salicornia persica in conjunction with hydrocarbon-degrading bacteria. J. Environ. Manag. 2018, 219, 260–268. [Google Scholar] [CrossRef]
- Jia, J.; Zong, S.; Hu, L.; Shi, S.; Zhai, X.; Wang, B.; Li, G.; Zhang, D. The Dynamic Change of Microbial Communities in Crude Oil-Contaminated Soils from Oil Fields in China. J. Soil Contam. 2017, 26, 171–183. [Google Scholar] [CrossRef]
- Dhote, M.; Kumar, A.; Jajoo, A.; Juwarkar, A. Assessment of hydrocarbon degradation potentials in plant-microbe interaction system with oil sludge contamination: A sustainable solution. Int. J. Phytoremediation 2017, 19, 1085–1092. [Google Scholar] [CrossRef]
- Aguilera, F.; Méndez, J.; Pásaro, E.; Laffon, B. Review on the effects of exposure to spilled oils on human health. J. Appl. Toxicol. 2010, 30, 291–301. [Google Scholar] [CrossRef]
- Robertson, L.W.; Hansen, L.G. PCBs: Recent Advances in Environmental Toxicology and Health Effects; The University Press of Kentucky: Lexington, KY, USA, 2015; pp. 511–512. [Google Scholar]
- Fatima, K.; Imran, A.; Naveed, M.; Afzal, M. Plant-bacteria synergism: An innovative approach for the remediation of crude oil-contaminated soils. Soil Environ. 2017, 36, 93–113. [Google Scholar] [CrossRef]
- Pinedo, J.; Ibáñez, R.; Lijzen, J.P.A.; Irabien, Á. Assessment of soil pollution based on total petroleum hydrocarbons and individual oil substances. J. Environ. Manag. 2013, 130, 72–79. [Google Scholar] [CrossRef]
- Tara, N.; Afzal, M.; Ansari, T.M.; Tahseen, R.; Iqbal, S.; Khan, Q.M. Combined use of Alkane-Degrading and Plant Growth-Promoting Bacteria Enhanced Phytoremediation of Diesel Contaminated soil. Int. J. Phytoremediation 2014, 16, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Sharma, R.; Singh, K.; Sharma, A. Application of bioremediation technology in the environment contaminated with petroleum hydrocarbon. Ann. Microbiol. 2013, 63, 417–431. [Google Scholar] [CrossRef]
- Varjani, S.J.; Rana, D.P.; Jain, A.K.; Bateja, S.; Upasani, V.N. Synergistic ex-situ biodegradation of crude oil by halotolerant bacterial consortium of indigenous strains isolated from on shore sites of Gujarat, India. Int. Biodeterior. Biodegrad. 2015, 103, 116–124. [Google Scholar] [CrossRef]
- Abbasian, F.; Lockington, R.; Mallavarapu, M.; Naidu, R. A Comprehensive Review of Aliphatic Hydrocarbon Biodegradation by Bacteria. Appl. Biochem. Biotechnol. 2015, 176, 670–699. [Google Scholar] [CrossRef] [PubMed]
- Committee P.I.S.T.C.P.G.E.P.S. Analysis of Soluble Organic Matter and Crude Oil Component in Rock (SY/T5119-2008); Petroleum Industry Press: Beijing, China, 2008. [Google Scholar]
- Fatima, K.; Afzal, M.; Imran, A.; Khan, Q.M. Bacterial rhizosphere and endosphere populations associated with grasses and trees to be used for phytoremediation of crude oil contaminated soil. Bull. Environ. Contam. Toxicol. 2015, 94, 314–320. [Google Scholar] [CrossRef]
- Kuppusamy, S.; Palanisami, T.; Megharaj, M.; Venkateswarlu, K.; Naidu, R. Ex-Situ Remediation Technologies for Environmental Pollutants: A Critical Perspective. Rev. Environ. Contam. Toxicol. 2016, 236, 117–192. [Google Scholar]
- Ron, E.Z.; Rosenberg, E. Enhanced bioremediation of oil spills in the sea. Curr. Opin. Biotechnol. 2014, 27, 191–194. [Google Scholar] [CrossRef]
- Chao, C.; Qiu, L.; Liu, C.; Yu, J. Effect of different enrichment strategies on microbial community structure in petroleum-contaminated marine sediment in Dalian, China. Mar. Pollut. Bull. 2017, 117, 274. [Google Scholar]
- Athar, H.U.; Ambreen, S.; Javed, M.; Hina, M.; Rasul, S.; Zafar, Z.U.; Manzoor, H.; Ogbaga, C.C.; Afzal, M.; Al-Qurainy, F. Influence of sub-lethal crude oil concentration on growth, water relations and photosynthetic capacity of maize (Zea mays L.) plants. Environ. Sci. Pollut. Res. 2016, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shehzadi, M.; Fatima, K.; Imran, A.; Mirza, M.S.; Khan, Q.M.; Afzal, M. Ecology of bacterial endophytes associated with wetland plants growing in textile effluent for pollutant-degradation and plant growth-promotion potentials. Giorn. Bot. Ital. 2015, 150, 1261–1270. [Google Scholar] [CrossRef]
- Fatima, K.; Imran, A.; Amin, I.; Khan, Q.M.; Afzal, M. Successful phytoremediation of crude-oil contaminated soil at an oil exploration and production company by plants-bacterial synergism. Int. J. Phytoremediation 2018, 20, 675. [Google Scholar] [CrossRef] [PubMed]
- Minoui, S.; Minai-Tehrani, D.; Shahriari, M.H. Phytoremediation of Crude Oil-Contaminated Soil by Medicago sativa (Alfalfa) and the Effect of Oil on its Growth; Springer: Berlin/Heidelberg, Germany, 2015; pp. 123–129. [Google Scholar]
- Almeida, R.; Mucha, A.P.; Teixeira, C.; Bordalo, A.A.; Almeida, C.M.R. Biodegradation of petroleum hydrocarbons in estuarine sediments: Metal influence. Biodegradation 2013, 24, 111–123. [Google Scholar] [CrossRef]
- Vaziri, A.; Panahpour, E.; Beni, M.H.M. Phytoremediation, a Method for Treatment of Petroleum Hydrocarbon Contaminated Soils. Int. J. Farm. Allied Sci. 2013, 2, 909–913. [Google Scholar]
- Jiao, S.; Chen, W.; Wei, G. Biogeography and ecological diversity patterns of rare and abundant bacteria in oil-contaminated soils. Mol. Ecol. 2017, 26, 5305–5317. [Google Scholar] [CrossRef]
- Nie, M.; Lu, M.; Yang, Q.; Zhang, X.D.; Xiao, M.; Jiang, L.F.; Yang, J.; Fang, C.M.; Chen, J.K.; Li, B. Plants’ use of different nitrogen forms in response to crude oil contamination. Environ. Pollut. 2011, 159, 157–163. [Google Scholar] [CrossRef]
- Fester, T.; Giebler, J.; Wick, L.Y.; Schlosser, D.; Kästner, M. Plant–microbe interactions as drivers of ecosystem functions relevant for the biodegradation of organic contaminants. Curr. Opin. Biotechnol. 2014, 27, 168–175. [Google Scholar] [CrossRef]
- Yu, J.; Kan, X.; Wang, X.; Han, G.; Guan, B.; Xie, W.; Lin, Q. Simulation of the effects of oil pollution in the Yellow River Delta on the growth of reeds and Suaeda salsa seedlings. Geogr. Sci. 2012, 10, 1254–1261. (In Chinese) [Google Scholar]
- Wang, J.; Mu, C.; Liu, F. Collection of Environmental Impact Assessment of Oilfield Development; China Environmental Science Press: Beijing, China, 1989. (In Chinese) [Google Scholar]
- Zhao, S.; Ye, J. Vegetation of Gudao Oil Production Plant; Shandong Provincial Map Press: Jinan, China, 1995. (In Chinese) [Google Scholar]
- Liu, Q.; Liu, G.; Li, H. Contents and Changes of Soil Petroleum Substances in Shengtuo and Gudong Oilfields. Soil Bull. 2003, 34, 592–593. (In Chinese) [Google Scholar]
- He, Q.; Xu, J.; Zhang, B.; Ye, X. Geological Environment and Sustainable Development of the Yellow River Delta; Geological Publishing House: Beijing, China, 2006. (In Chinese) [Google Scholar]
- Wang, C.; Yang, C.; Sun, Z.; Yang, Y.; Qu, C.; Wang, Y. Relationship between soil petroleum pollution and physical and chemical properties in the ecological zone of the Yellow River Delta. J. Soil Water Conserv. 2010, 24, 214–217. [Google Scholar]
- He, Q.; Cui, B.; Bertness, M.D.; An, Y. Testing the importance of plant strategies on facilitation using congeners in a coastal community. Ecology 2012, 93, 2023–2029. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Altieri, A.H.; Cui, B. Herbivory drives zonation of stress-tolerant marsh plants. Ecology 2016, 96, 1318–1328. [Google Scholar] [CrossRef] [Green Version]
- Bao, S. Soil Agrochemical Analysis, 3rd ed.; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Pielou, E.C.; Levandowsky, M. Ecological Diversity; John Wiley & Sons: New York, NY, USA, 1975. [Google Scholar]
- Liao, J.; Wang, J.; Huang, Y. Bacterial Community Features Are Shaped by Geographic Location, Physicochemical Properties, and Oil Contamination of Soil in Main Oil Fields of China. Microb. Ecol. 2015, 70, 1–10. [Google Scholar] [CrossRef]
- Liu, R.; Yu, Z.; Ran, D. Comparison of archaeal and bacterial community structures in heavily oil-contaminated and pristine soils. J. Biosci. Bioeng. 2009, 108, 400–407. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination; Version 5.0; Biometris: New York, NY, USA, 2012. [Google Scholar]
- Marinescu, M.; Toti, M.; Tanase, V.; Carabulea, V. An assessment of the effects of crude oil pollution on soil properties. Ann. Food Sci. Technol. 2010, 11, 94–99. [Google Scholar]
- Ji, Z. Study on Physicochemical Properties and Enzymes of Oil-Contaminated Soil by Suaeda salsa-Microorganism-Sericulture; Dalian Ocean University: Dalian City, China, 2016. (In Chinese) [Google Scholar]
- Leahy, J.G.; Colwell, R.R. Microbial degradation of hydrocarbons in the environment. Microbiol. Mol. Biol. Rev. 1990, 54, 305–315. [Google Scholar] [CrossRef]
- Kuo, H.-C.; Juang, D.-F.; Yang, L.; Kuo, W.-C.; Wu, Y.-M. Phytoremediation of soil contaminated by heavy oil with plants colonized by mycorrhizal fungi. Int. J. Environ. Sci. Technol. 2014, 11, 1661–1668. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, D.; Dussan, J. Landfarmed oil sludge as a carbon source for Canavalia ensiformis during phytoremediation. Int. J. Environ. Sci. Technol. 2014, 11, 1197–1206. [Google Scholar] [CrossRef] [Green Version]
- Asemoloye, M.D.; Ahmad, R.; Jonathan, S.G. Synergistic action of rhizospheric fungi with Megathyrsus maximus root speeds up hydrocarbon degradation kinetics in oil polluted soil. Chemosphere 2017, 187, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sutton, N.B.; Maphosa, F.; Morillo, J.A.; Al-Soud, W.A.; Langenhoff, A.A.; Grotenhuis, T.; Rijnaarts, H.H.; Smidt, H. Impact of long-term diesel contamination on soil microbial community structure. Appl. Environ. Microbiol. 2013, 79, 619–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mnif, S.; Sayadi, S.; Chamkha, M. Biodegradative potential and characterization of a novel aromatic-degrading bacterium isolated from a geothermal oil field under saline and thermophilic conditions. Int. Biodeterior. Biodegrad. 2014, 86, 258–264. [Google Scholar] [CrossRef]
- Varjani, S.J. Microbial degradation of petroleum hydrocarbons. Bioresour. Technol. 2017, 223, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.; Chen, W.; Wang, J.; Zhang, L.; Yang, F.; Lin, Y.; Wei, G. Plant growth and oil contamination after the diversity and compositon of bacterial communities in agricultural soils across China. Land Degrad. Dev. 2018, 29, 1660–1671. [Google Scholar] [CrossRef]
- Peng, M.; Zi, X.; Wang, Q. Bacterial Community Diversity of Oil-Contaminated Soils Assessed by High Throughput Sequencing of 16S rRNA Genes. Int. J. Environ. Res. Public Health 2015, 12, 12002–12015. [Google Scholar] [CrossRef] [Green Version]
- Weyens, N.; van der Lelie, D.; Taghavi, S.; Vangronsveld, J. Phytoremediation: Plant-endophyte partnerships take the challenge. Curr. Opin. Biotechnol. 2009, 20, 248–254. [Google Scholar] [CrossRef]
- Sessitsch, A.; Kuffner, M.; Kidd, P.; Vangronsveld, J.; Wenzel, W.W.; Fallmann, K.; Puschenreiter, M. The role of plant-associated bacteria in the mobilization and phytoextraction of trace elements in contaminated soils. Soil Biol. Biochem. 2013, 60, 182–194. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. Using soil bacteria to facilitate phytoremediation. Biotechnol. Adv. 2010, 28, 367–374. [Google Scholar] [CrossRef]
- Kuiper, I.; Lagendijk, E.L.; Bloemberg, G.V.; Lugtenberg, B.J. Rhizoremediation: A beneficial plant-microbe interaction. Mol. Plant-Mic. Interact. 2004, 17, 6. [Google Scholar] [CrossRef] [Green Version]
- Alkhatib, M.A.F.; Alam, Z.; Muyibi, S.A.; Husain, A. An isolated bacterial consortium for crude oil biodegradation. Afr. J. Biotechnol. 2011, 10, 18763–18767. [Google Scholar]
- Mukred, A.M.; Hamid, A.A.; Hamzah, A.; Yusoff, W.M.W. Development of Three Bacteria Consortium for the Bioremediation of Crude Petroleum-oil in Contaminated Water. Online J. Biol. Sci. 2008, 2008, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Chen, Y.P.; Jin, P.K.; Wang, X.C. Bacterial communities in a crude oil gathering and transferring system (China). Anaerobe 2009, 15, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340. [Google Scholar] [CrossRef]
- Nacke, H.; Thürmer, A.; Wollherr, A.; Will, C.; Hodac, L.; Herold, N.; Schöning, I.; Schrumpf, M.; Daniel, R. Pyrosequencing-based assessment of bacterial community structure along different management types in German forest and grassland soils. PLoS ONE 2011, 6, e17000. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, J.; Liao, J.; Xie, S.; Huang, Y. Abundance and diversity of soil petroleum hydrocarbon-degrading microbial communities in oil exploring areas. Appl. Microbiol. Biotechnol. 2015, 99, 1935–1946. [Google Scholar] [CrossRef]
- Powell, S.M.; Bowman, J.P.; Snape, I.; Stark, J.S. Microbial community variation in pristine and polluted nearshore Antarctic sediments. FEMS Microbiol. Ecol. 2003, 45, 135–145. [Google Scholar] [CrossRef]
- Paerl, H.W.; Dyble, J.; Moisander, P.H.; Noble, R.T.; Piehler, M.F.; Pinckney, J.L.; Steppe, T.F.; Twomey, L.; Valdes, L.M. Microbial indicators of aquatic ecosystem change: Current applications to eutrophication studies. FEMS Microbiol. Ecol. 2003, 46, 233–246. [Google Scholar] [CrossRef]
- Arslan, M.; Imran, A.; Khan, Q.M.; Afzal, M. Plant–bacteria partnerships for the remediation of persistent organic pollutants. Environ. Sci. Pollut. Res. 2017, 24, 4322–4336. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Zhang, Y.; Zhao, N.; Guo, J.; Xu, W.; Ma, M.; Li, X. Remediation of Crude Oil-Polluted Soil by the Bacterial Rhizosphere Community of Suaeda Salsa Revealed by 16S rRNA Genes. Int. J. Environ. Res. Public Health 2020, 17, 1471. https://doi.org/10.3390/ijerph17051471
Yu Y, Zhang Y, Zhao N, Guo J, Xu W, Ma M, Li X. Remediation of Crude Oil-Polluted Soil by the Bacterial Rhizosphere Community of Suaeda Salsa Revealed by 16S rRNA Genes. International Journal of Environmental Research and Public Health. 2020; 17(5):1471. https://doi.org/10.3390/ijerph17051471
Chicago/Turabian StyleYu, Yilei, Yinghua Zhang, Nana Zhao, Jia Guo, Weigang Xu, Muyuan Ma, and Xiaoxia Li. 2020. "Remediation of Crude Oil-Polluted Soil by the Bacterial Rhizosphere Community of Suaeda Salsa Revealed by 16S rRNA Genes" International Journal of Environmental Research and Public Health 17, no. 5: 1471. https://doi.org/10.3390/ijerph17051471