Indoor Microbiome and Antibiotic Resistance on Floor Surfaces: An Exploratory Study in Three Different Building Types

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sampling Sites and Indoor Areas
2.2. Sample Collection and DNA Extraction
2.3. Detection of Antibiotic Resistance Genes and Microbial Source Tracking
2.4. Bacterial Community Analysis
2.5. Data Analyses
3. Results and Discussion
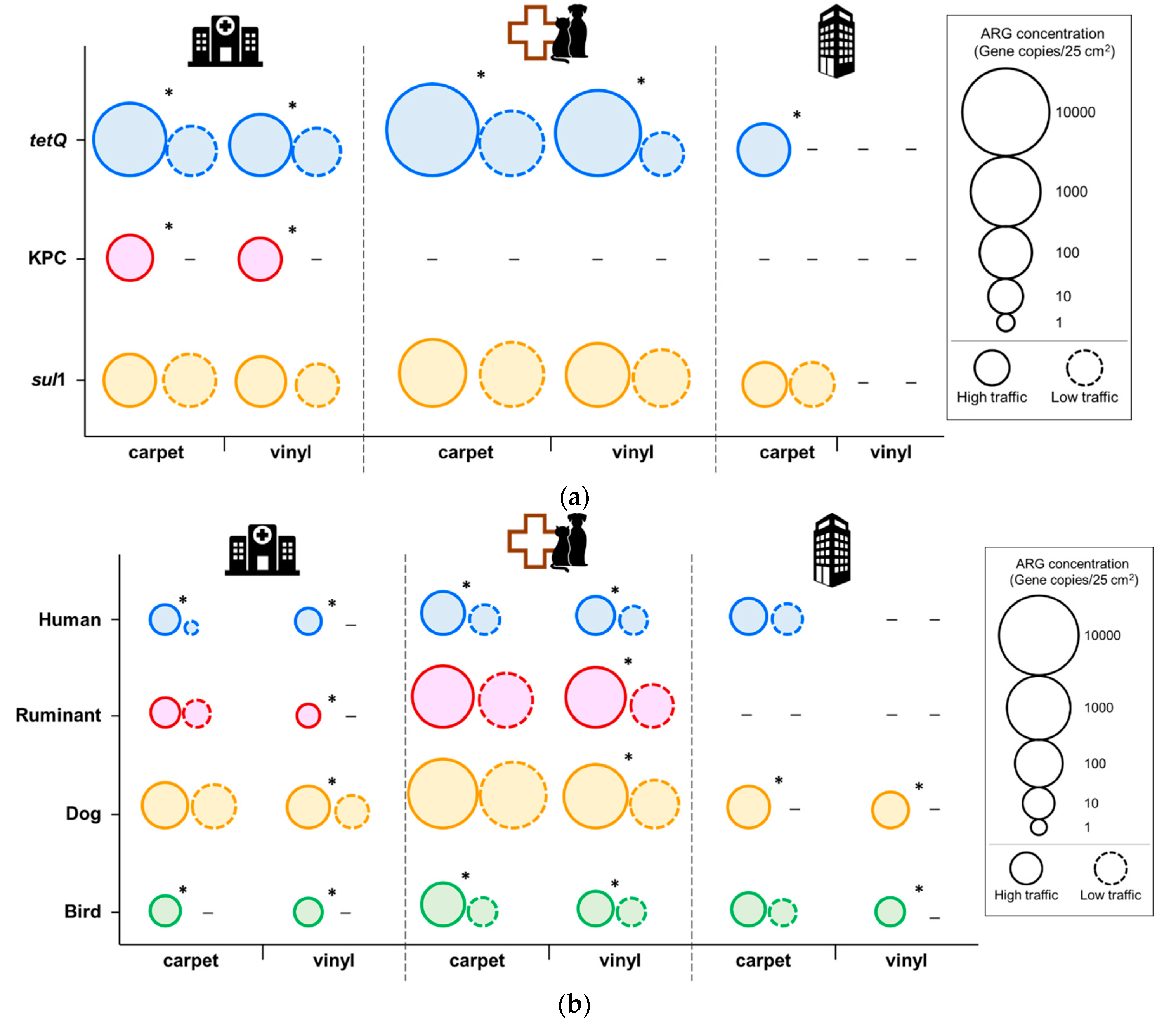
3.1. Antibiotic Resistance
3.2. Microbial Source Tracking
3.3. Bacterial Community
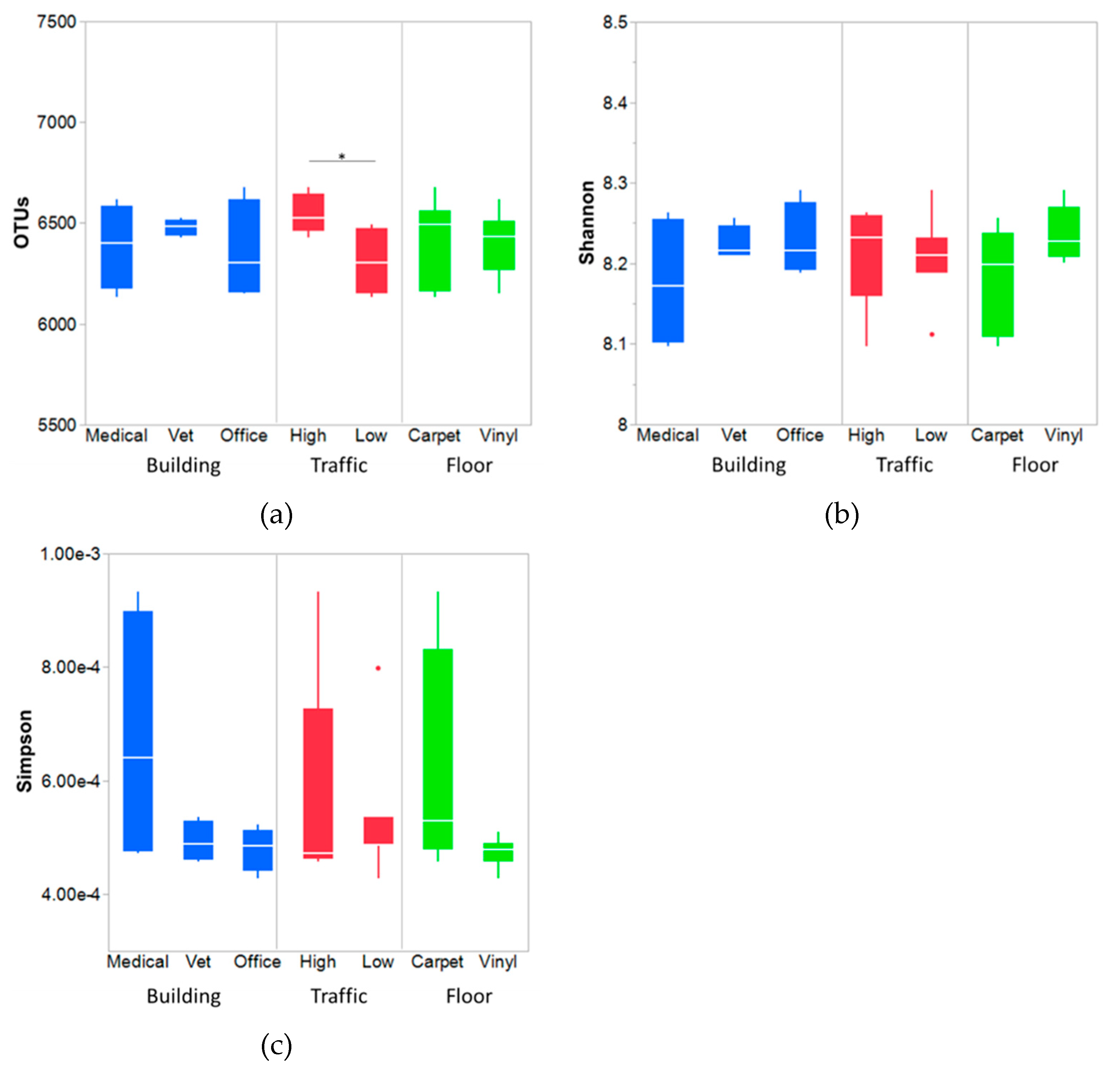
3.3.1. Bacterial Diversity and Richness
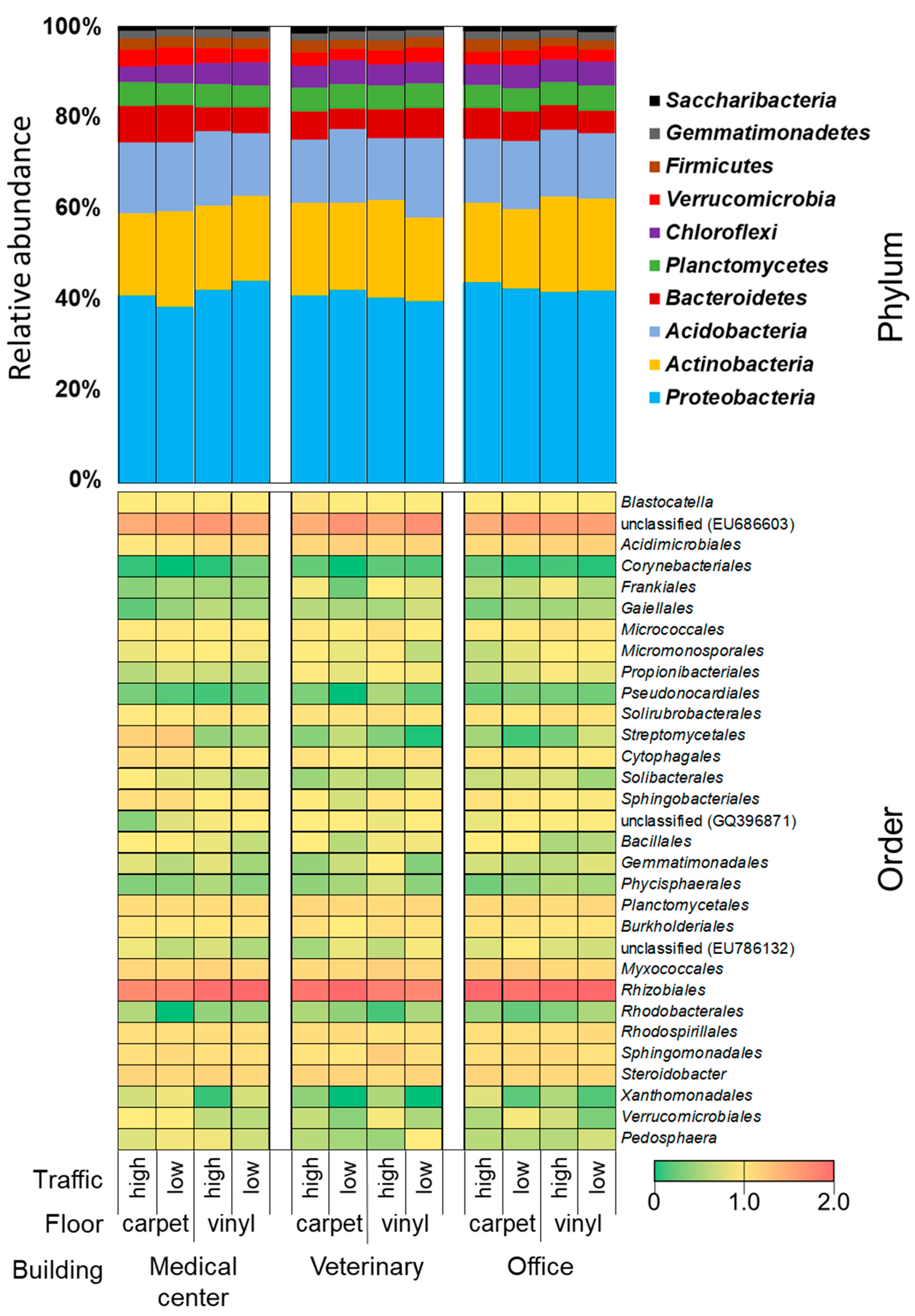
3.3.2. Bacterial Community Composition
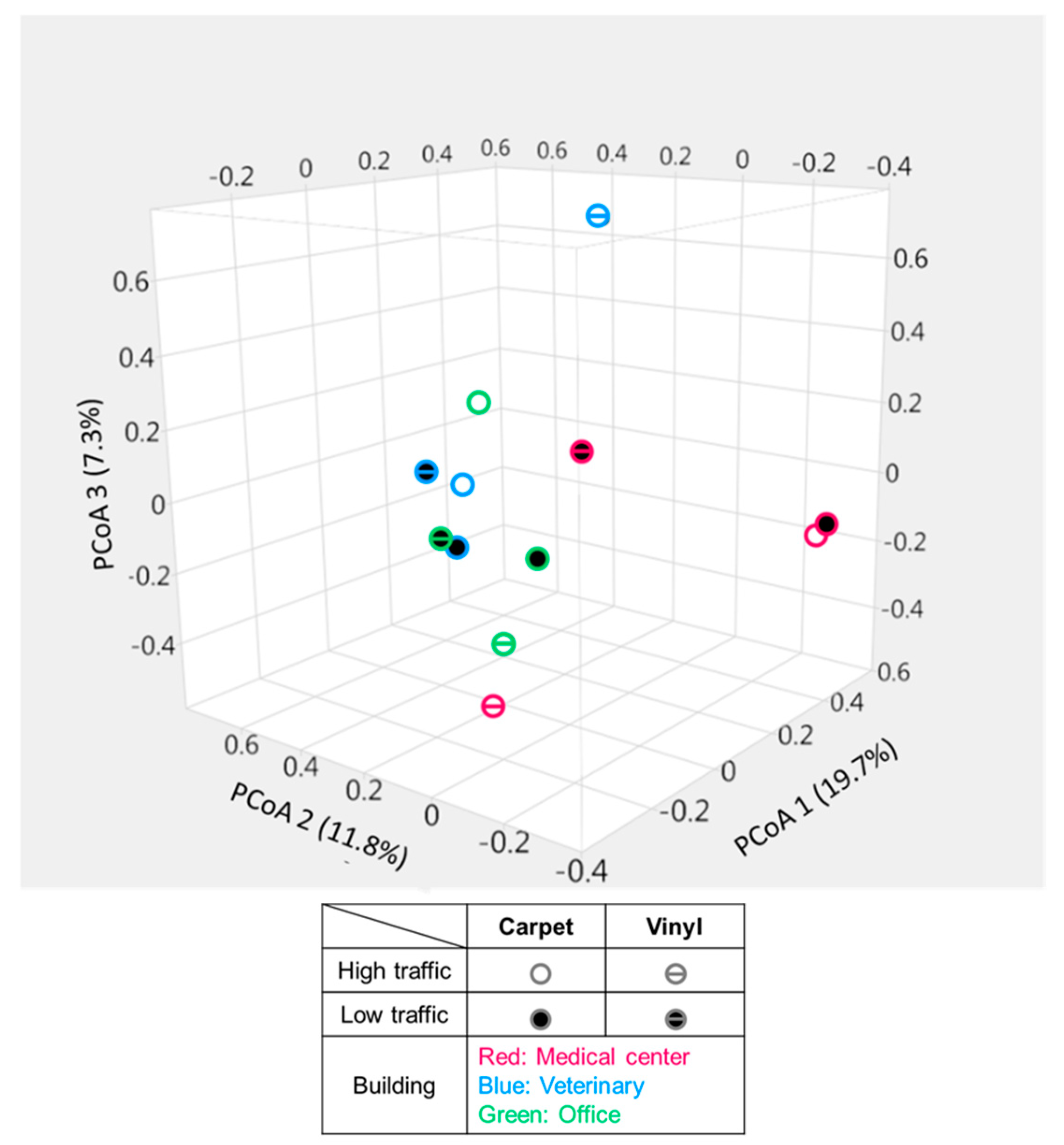
3.3.3. Bacterial Community Structure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, S.T.; Gilbert, J.A. Studying the microbiology of the indoor environment. Genome Bio. 2013, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- Prussin, A.J.; Marr, L.C. Sources of airborne microorganisms in the built environment. Microbiome 2015, 3, 78. [Google Scholar] [CrossRef] [PubMed]
- Tringe, S.G.; Zhang, T.; Liu, X.; Yu, Y.; Lee, W.H.; Yap, J.; Yao, F.; Suan, S.T.; Ing, S.K.; Haynes, M.; et al. The airborne metagenome in an indoor urban environment. PLoS ONE 2008, 3, e1862. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; Jones, E.; Kline, J.; Northcutt, D.; Stenson, J.; Womack, A.M.; Bohannan, B.J.; Brown, G.Z.; Green, J.L. Architectural design influences the diversity and structure of the built environment microbiome. ISME J. 2012, 6, 1469. [Google Scholar] [CrossRef]
- Narui, K.; Noguchi, N.; Matsunaga, N.; Namiki, Y.; Yamanaka, Y.; Kumaki, Y.; Suwa, J.; Nasu, Y.; Koyama, K.; Okuyama, K.; et al. Change in environmental bacterial flora in a new hospital building. J. Hosp. Infect. 2009, 73, 24–33. [Google Scholar] [CrossRef]
- Coughenour, C.; Stevens, V.; Stetzenbach, L.D. An Evaluation of Methicillin-Resistant Staphylococcus aureus Survival on Five Environmental Surfaces, Microb. Drug Resist. 2011, 17, 457–461. [Google Scholar] [CrossRef]
- Flores, G.E.; Bates, S.T.; Knights, D.; Lauber, C.L.; Stombaugh, J.; Knight, R.; Fierer, N. Microbial biogeography of public restroom surfaces. PLoS ONE 2011, 6, e28132. [Google Scholar] [CrossRef]
- Lax, S.; Smith, D.P.; Hampton-Marcell, J.; Owens, S.M.; Handley, K.M.; Scott, N.M.; Gibbons, S.M.; Larsen, P.; Shogan, B.D.; Weiss, S.; et al. Longitudinal analysis of microbial interaction between humans and the indoor environment. Science 2014, 345, 1048–1052. [Google Scholar] [CrossRef] [Green Version]
- Meadow, J.F.; Altrichter, A.E.; Kembel, S.W.; Moriyama, M.; O’Connor, T.K.; Womack, A.M.; Bohannan, B.J.M. Bacterial communities on classroom surfaces vary with human contact. Microbiome 2014, 2, 7. [Google Scholar] [CrossRef]
- Adams, R.I.; Bateman, A.C.; Bik, H.M.; Meadow, J.F. Microbiota of the indoor environment: A meta-analysis. Microbiome 2015, 13, 49. [Google Scholar] [CrossRef] [PubMed]
- Dunn, R.R.; Fierer, N.; Henley, J.B.; Leff, J.W.; Menninger, H.L. Home life: Factors structuring the bacterial diversity found within and between homes. PLoS ONE 2013, 8, e64133. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, S.M.; Schwartz, T.; Fouquier, J.; Mitchell, M.; Sangwan, N.; Gilbert, J.A.; Kelley, S.T. Ecological succession and viability of human-associated microbiota on restroom surfaces. Appl. Environ. Microbiol. 2015, 81, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Brent, S. What have we learned about the microbiomes of indoor environments? mSystems 2016, 1, e00083-16. [Google Scholar] [CrossRef]
- Chase, J.; Fouquier, J.; Zare, M.; Sonderegger, D.L.; Knight, R.; Kelley, S.T.; Siegel, J.; Caporaso, J.G. Geography and location are the primary drivers of office microbiome composition. mSystems 2016, 1, e00022-16. [Google Scholar] [CrossRef]
- Hartmann, E.M.; Hickey, R.; Hsu, T.; Román, C.M.B.; Chen, J.; Schwager, R.; Kline, J.; Brown, G.Z.; Halden, R.U.; Huttenhower, C.; et al. Antimicrobial chemicals are associated with elevated antibiotic resistance genes in the indoor dust microbiome. Environ. Sci. Technol. 2016, 50, 9807–9815. [Google Scholar] [CrossRef]
- Heuer, H.; Schmitt, H.; Smalla, K. Antibiotic resistance gene spread due to manure application on agricultural fields. Curr. Opin. Microbiol. 2011, 14, 236–243. [Google Scholar] [CrossRef]
- Kummerer, K. Resistance in the environment. J. Antimicrob. Chemother. 2004, 54, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.S.; Yang, R.K.; Knapp, C.W.; Niu, Y.; Peak, N.; Hanfelt, M.M.; Galland, J.C.; Graham, D.W. Quantification of tetracycline resistance genes in feedlot lagoons by real-time PCR. Appl. Environ. Microbiol. 2004, 70, 7372–7377. [Google Scholar] [CrossRef]
- Armand-Lefevre, L.; Ruimy, R.; Andremont, A. Clonal comparison of Staphylococcus aureus isolates from healthy pig farmers, human controls, and pigs. Emerg. Infect. Dis. 2005, 11, 711–714. [Google Scholar] [CrossRef]
- Roberts, M.C. Update on acquired tetracycline resistance genes. FEMS Microbiol. Lett. 2005, 245, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Zhang, T. Occurrence, abundance, and diversity of tetracycline resistance genes in 15 sewage treatment plants across china and other global locations. Environ. Sci. Technol. 2011, 45, 2598–2604. [Google Scholar] [CrossRef] [PubMed]
- Ling, A.L.; Pace, N.R.; Hernandez, M.T.; LaPara, T.M. Tetracycline resistance and class 1 integron genes associated with indoor and outdoor aerosols. Environ. Sci. Technol. 2013, 47, 4046–4052. [Google Scholar] [CrossRef] [PubMed]
- Lerma, L.L.; Benomar, N.; Knapp, C.W.; Galeote, D.C.; Gálvez, A.; Abriouel, H. Diversity, distribution and quantification of antibiotic resistance genes in goat and lamb slaughterhouse surfaces and meat products. PLoS ONE 2014, 9, e114252. [Google Scholar] [CrossRef]
- Oberauner, L.; Zachow, C.; Lackner, S.; Högenauer, C.; Smolle, K.H.; Berg, G. The ignored diversity: Complex bacterial communities in intensive care units revealed by 16S pyrosequencing. Sci. Rep. 2013, 3, 1413. [Google Scholar] [CrossRef]
- Nikolich, M.P.; Hong, G.; Shoemaker, N.B.; Salyers, A.A. Evidence for natural horizontal transfer of tetQ between bacteria that normally colonize humans and bacteria that normally colonize livestock. Appl. Environ. Microbiol. 1994, 60, 3255–3260. [Google Scholar] [Green Version]
- Luo, Y.; Mao, D.; Rysz, M.; Zhou, Q.; Zhang, H.; Xu, L.; Alvarez, P.J.J. Trends in antibiotic resistance genes occurrence in the Haihe River, China. Environ. Sci. Technol. 2010, 44, 7220–7225. [Google Scholar] [CrossRef]
- Chmelnitsky, I.; Shklyar, M.; Leavitt, A.; Sadovsky, E.; Navon-Venezia, S.; Dalak, M.B.; Edgar, R.; Carmeli, Y. Mix and match of KPC-2 encoding plasmids in Enterobacteriaceae-comparative genomics. Diagn. Microbiol. Infect. Dis. 2014, 79, 255–260. [Google Scholar] [CrossRef]
- Hsu, T.T.D.; Mitsch, W.J.; Martin, J.F.; Lee, J. Towards sustainable protection of public health: The role of an urban wetland as a frontline safeguard of pathogen and antibiotic resistance spread. Ecol. Eng. 2017, 108, 547–555. [Google Scholar] [CrossRef]
- Haugland, R.A.; Varma, M.; Sivaganesan, M.; Kelty, C.; Peed, L.; Shanks, O.C. Evaluation of genetic markers from the 16S rRNA gene V2 region for use in quantitative detection of selected Bacteroidales species and human fecal waste by qPCR. Syst. Appl. Microbiol. 2010, 33, 348–357. [Google Scholar] [CrossRef]
- Mieszkin, S.; Yala, J.F.; Joubrel, R.; Gourmelon, M. Phylogenetic analysis of Bacteroidales 16S rRNA gene sequences from human and animal effluents and assessment of ruminant faecal pollution by real-time PCR. J. Appl. Microbiol. 2010, 108, 974–984. [Google Scholar] [CrossRef] [PubMed]
- Kildare, B.J.; Leutenegger, C.M.; McSwain, B.S.; Bambic, D.G.; Rajal, V.B.; Wuertz, S. 16S rRNA-based assays for quantitative detection of universal, human-, cow-, and dog-specific fecal Bacteroidales: A Bayesian approach. Water Res. 2007, 41, 3701–3715. [Google Scholar] [CrossRef] [PubMed]
- Green, H.C.; Dick, L.K.; Gilpin, B.; Samadpour, M.; Field, K.G. Genetic markers for rapid PCR-based identification of gull, Canada goose, duck, and chicken fecal contamination in water. Appl. Environ. Microbiol. 2012, 78, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Agidi, S.; Marion, J.W.; Lee, J. Arcobacter in Lake Erie beach waters: An emerging gastrointestinal pathogen linked with human-associated fecal contamination. Appl. Environ. Microbiol. 2012, 78, 5511–5519. [Google Scholar] [CrossRef]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef]
- Layton, D.W.; Beamer, P.I. Migration of contaminated soil and airborne particulates to indoor dust. Environ. Sci. Technol. 2009, 43, 8199–8205. [Google Scholar] [CrossRef]
- Chao, A. Non-parametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Hunt, A.; Johnson, D.L.; Griffith, D.A. Mass transfer of soil indoors by track-in on footwear. Sci. Total Environ. 2006, 370, 360–371. [Google Scholar] [CrossRef]
- Lax, S.; Hampton-Marcell, J.T.; Gibbons, S.M.; Colares, G.B.; Smith, D.; Eisen, J.A.; Gilbert, J.A. Forensic analysis of the microbiome of phones and shoes. Microbiome 2015, 3, 21. [Google Scholar] [CrossRef]
- Rintala, H.; Pitkäranta, M.; Toivola, M.; Paulin, L.; Nevalainen, A. Diversity and seasonal dynamics of bacterial community in indoor environment. BMC Microbiol. 2008, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Cytryn, E. The soil resistome: The anthropogenic, the native, and the unknown. Soil Biol. Biochem. 2013, 63, 18–23. [Google Scholar] [CrossRef]
- Hewitt, K.M.; Gerba, C.P.; Maxwell, S.L.; Kelley, S.T. Office space bacterial abundance and diversity in three metropolitan areas. PLoS ONE 2012, 7, e37849. [Google Scholar] [CrossRef] [PubMed]
- Da Fonseca, T.A.P.; Pessôa, R.; Felix, A.C.; Sanabani, S.S. Diversity of bacterial communities on four frequently used surfaces in a large Brazilian teaching hospital. Int. J. Environ. Res. Public Health 2016, 13, 152. [Google Scholar] [CrossRef] [PubMed]
- Suihko, M.L.; Priha, O.; Alakomi, H.L.; Thompson, P.; Mälarstig, B.; Stott, R.; Richardson, M. Detection and molecular characterization of filamentous actinobacteria and thermoactinomycetes present in water-damaged building materials. Indoor Air 2009, 19, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Shafer, J.; Jackel, U.; Kampfer, P. Analysis of Actinobacteria from mould-colonized water damaged building material. Syst. Appl. Microbiol. 2010, 33, 260–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locations | Areas | Floor | Traffic |
|---|---|---|---|
| Medical center | Main entrance | Carpet | High |
| Office | Carpet | Low | |
| Main entrance | Vinyl | High | |
| Patient room | Vinyl | Low | |
| Veterinary hospital | Small animal office | Carpet | High |
| Large farm animal clinic | Carpet | Low | |
| Small animal clinic | Vinyl | High | |
| Large farm animal clinic office | Vinyl | Low | |
| Office/classroom building | Classroom | Carpet | High |
| Office | Carpet | Low | |
| Laboratory room | Vinyl | Low | |
| Laboratory | Vinyl | Low |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, M.; Lee, S.; Bisesi, M.; Lee, J. Indoor Microbiome and Antibiotic Resistance on Floor Surfaces: An Exploratory Study in Three Different Building Types. Int. J. Environ. Res. Public Health 2019, 16, 4160. https://doi.org/10.3390/ijerph16214160
Gupta M, Lee S, Bisesi M, Lee J. Indoor Microbiome and Antibiotic Resistance on Floor Surfaces: An Exploratory Study in Three Different Building Types. International Journal of Environmental Research and Public Health. 2019; 16(21):4160. https://doi.org/10.3390/ijerph16214160
Chicago/Turabian StyleGupta, Mridula, Seungjun Lee, Michael Bisesi, and Jiyoung Lee. 2019. "Indoor Microbiome and Antibiotic Resistance on Floor Surfaces: An Exploratory Study in Three Different Building Types" International Journal of Environmental Research and Public Health 16, no. 21: 4160. https://doi.org/10.3390/ijerph16214160