
The Inflammation Response to DEHP through PPARγ in Endometrial Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatment
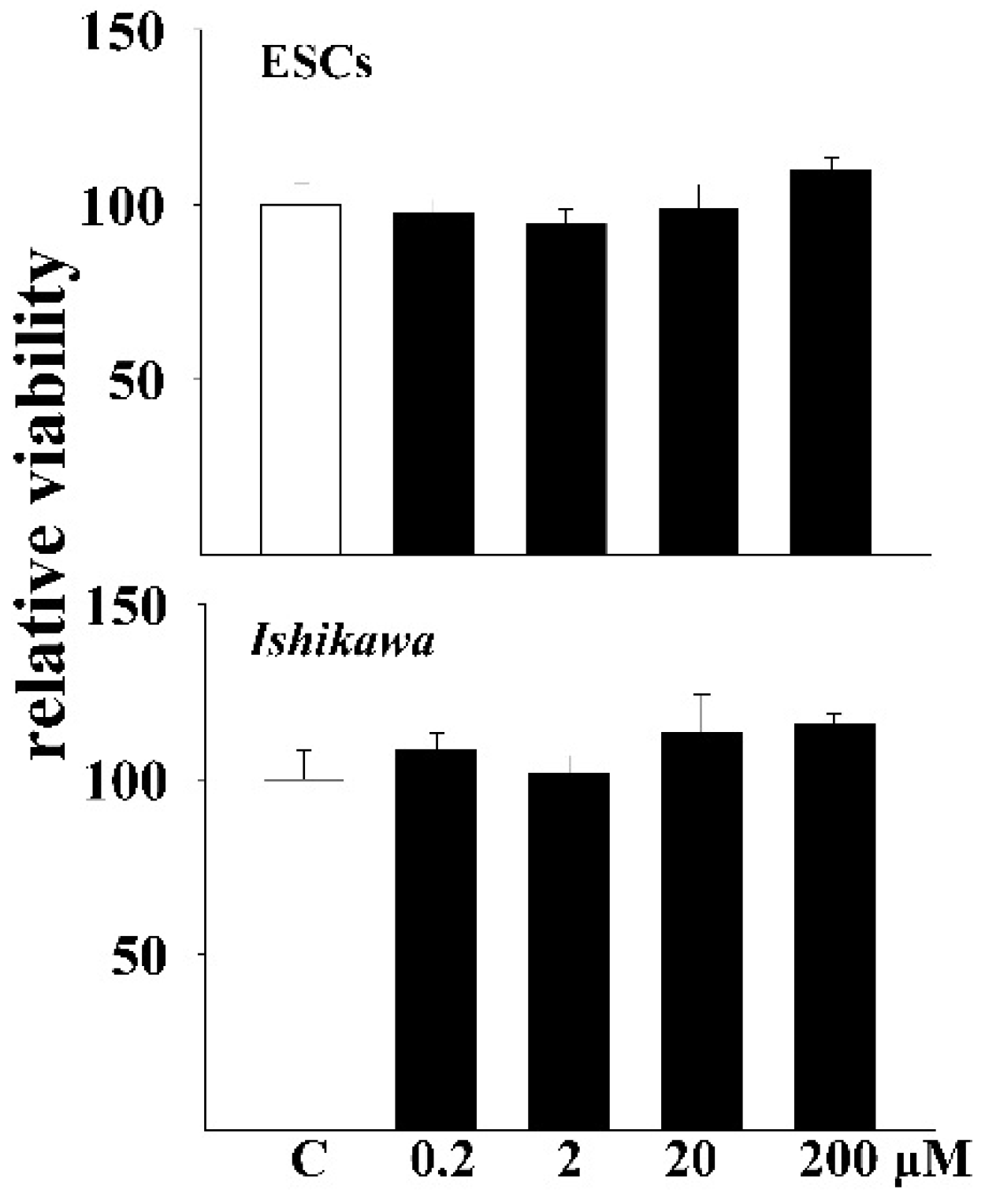
2.2. MTT Assays
2.3. RNA Isolation and SYBR Realtime RT-PCR
2.4. ELISA Assay
2.5. Western Blot Analysis
2.6. Data Analysis
3. Results
3.1. DEHP Did Not Inhibit the Proliferation of Endometrial Cells
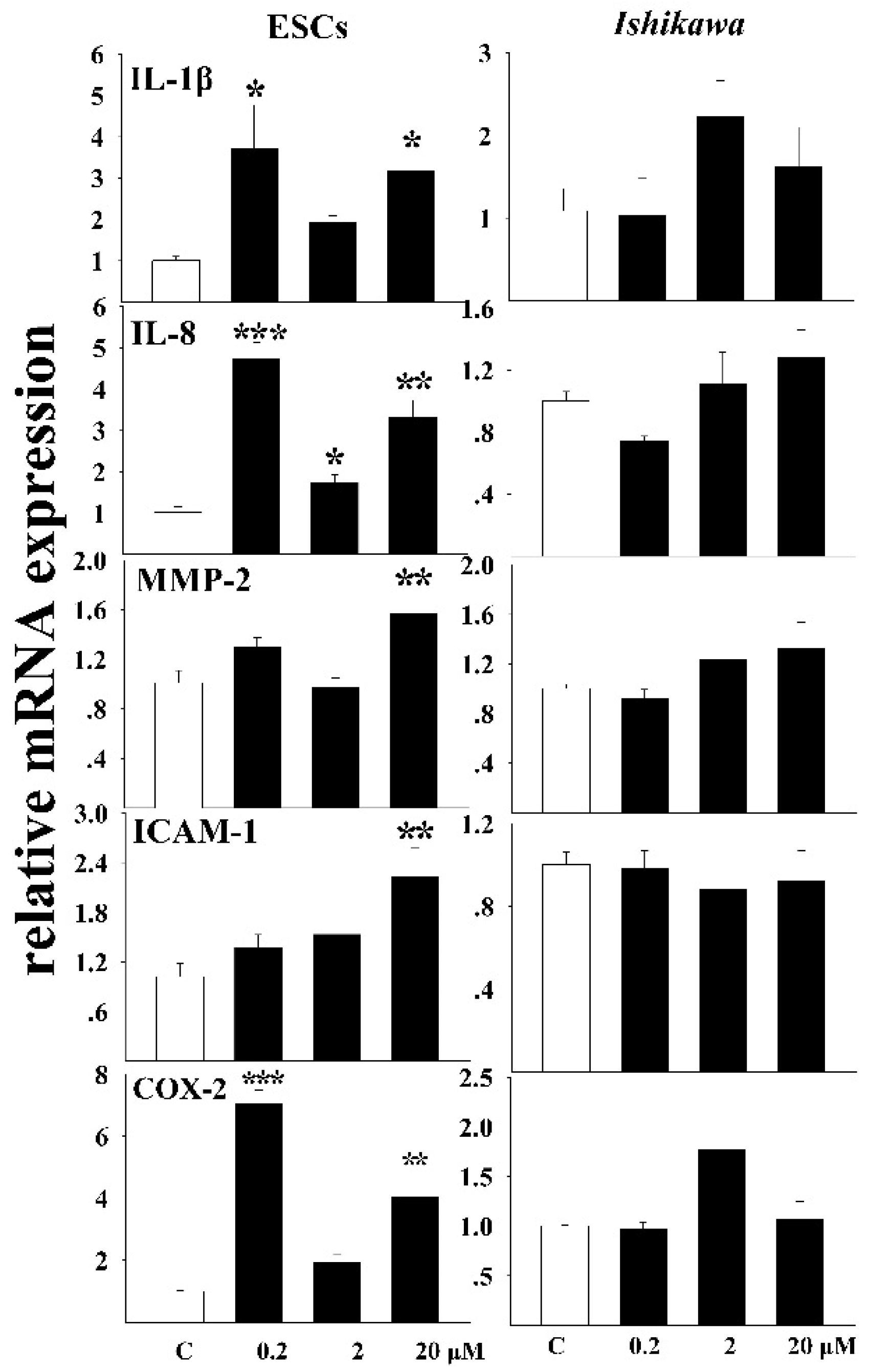
3.2. The Inflammatory Response of Cells to DEHP Exposure
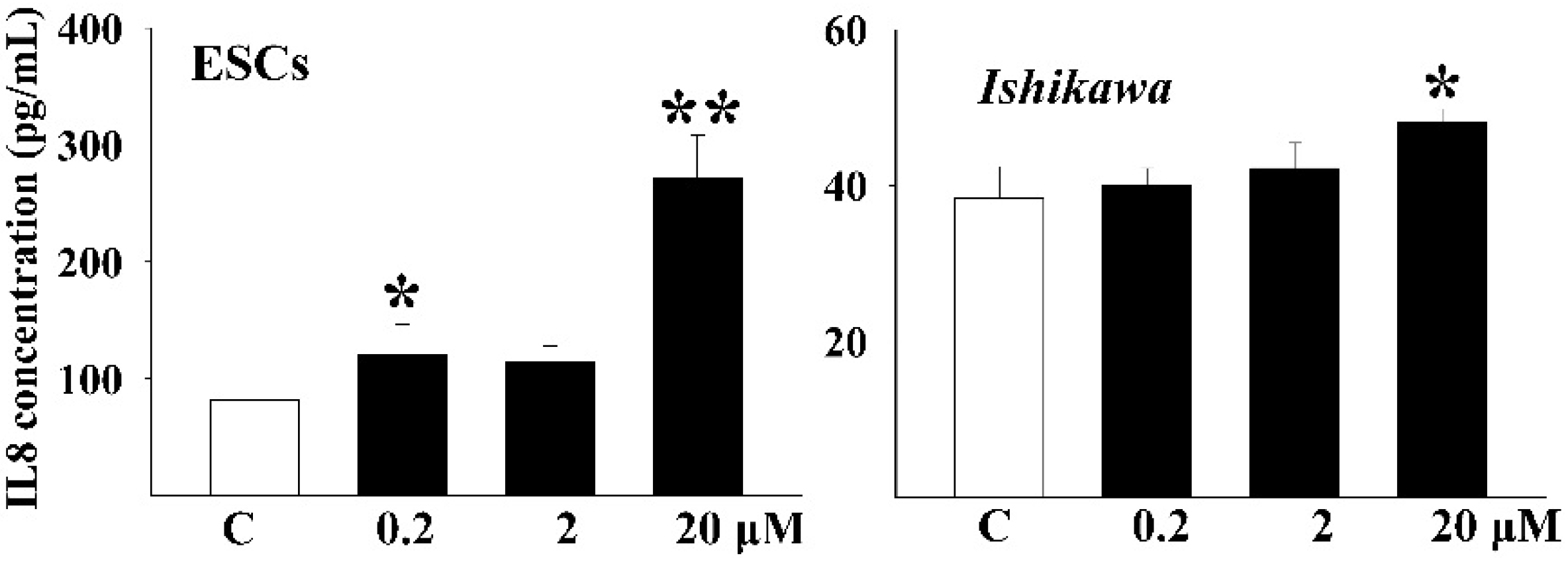
3.3. DEHP Induced the Secretion of Inflammatory Factors
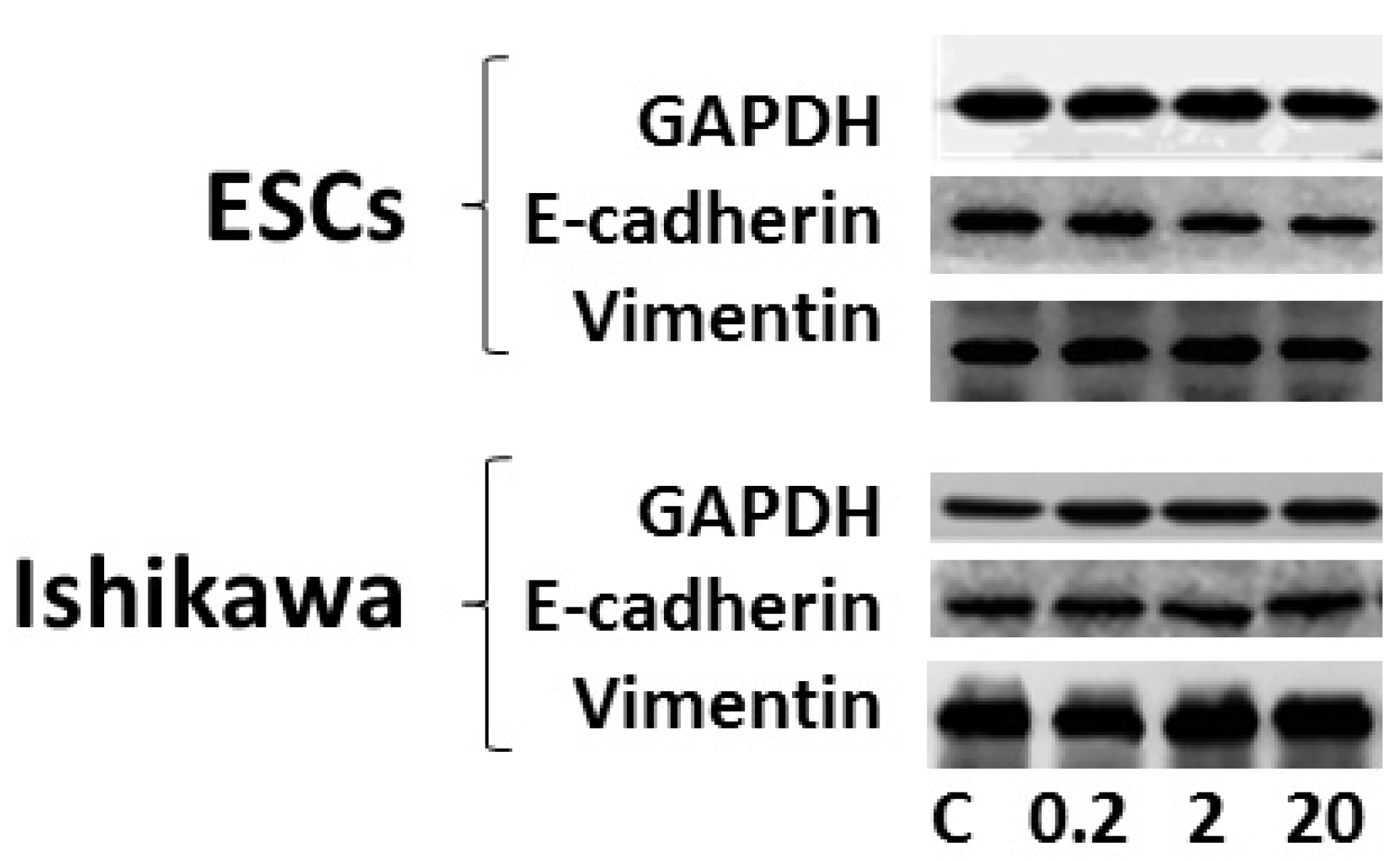
3.4. Effects of DEHP on Epithelial-Mesenchymal Transition (EMT)
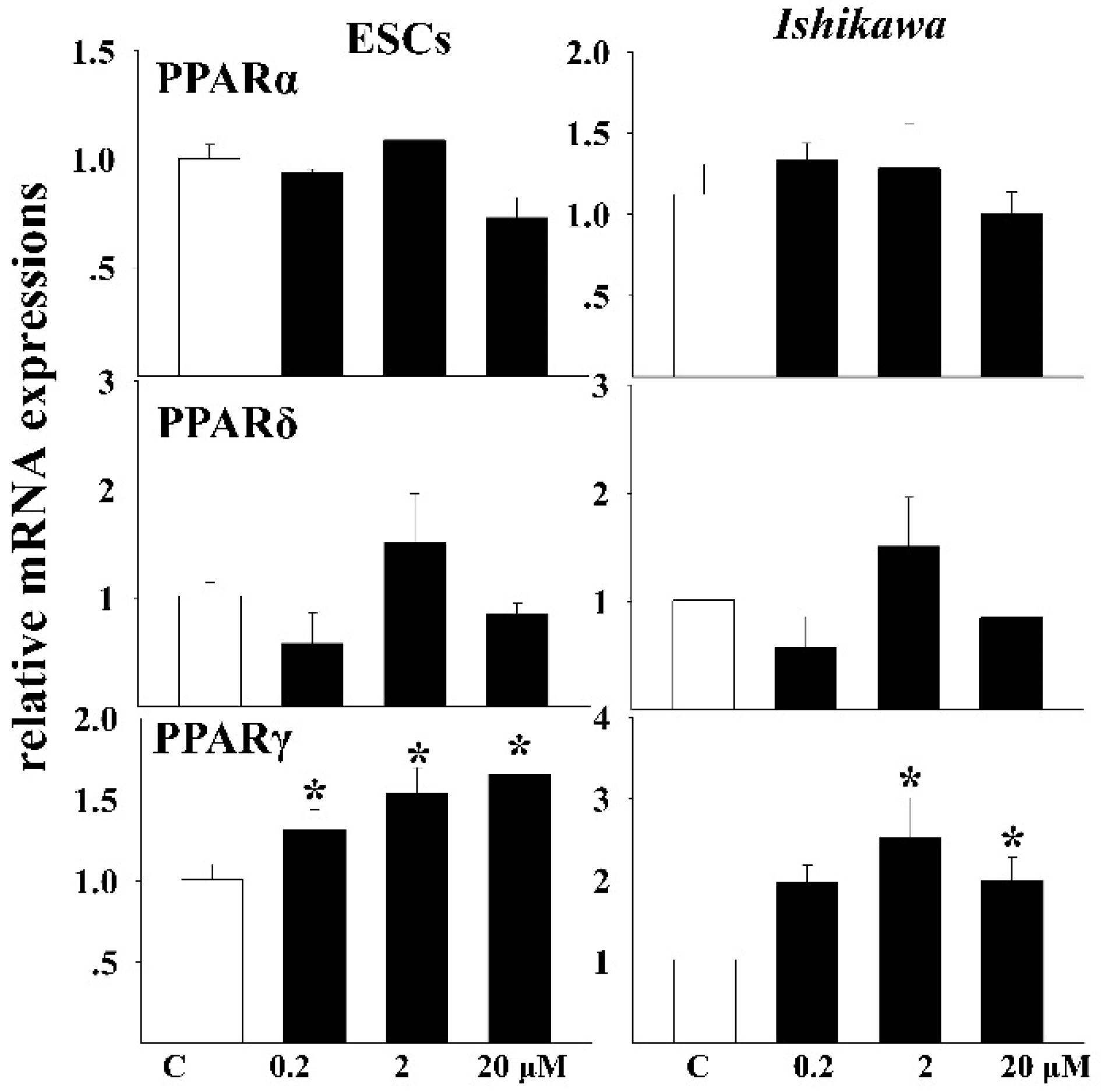
3.5. Effects of DEHP on the mRNA Expressions of Related Receptors
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Nucleotide Accession Number | Primer Sequences Used for qRT-PCR (5’ to 3’) |
|---|---|---|
| Glyceraldehyde-3-phosphate dehydrogenase(GAPDH) | NM_002046 | F: GGAGAAGGCTGGGGCTCAT |
| R: TGATGGCATGGACTGTGGTC | ||
| Interleukin 1 beta (IL-1β) | NM_000576 | F: GGACAAGCTGAGGAAGATGC |
| R: TCGTTATCCCATGTGTCGAA | ||
| Interleukin (IL-8) | NM_000584 | F: GCATAAAGACATACTCCAAACC |
| R: ACTTCTCCACAACCCTCTG | ||
| Intercellular cell adhesion molecule-1 (ICAM-1) | J03132 | F: CCGGAAGGTGTATGAACTGA |
| R: GGCAGCGTAGGGTAAGGTT | ||
| Matrix metalloproteinase-2 (mmp-2) | NM_001127891 | F: TGACGGTAAGGACGGACTC |
| R: ATACTTCACACGGACCACTTG | ||
| Cyclooxygenase-2 (COX-2) | M90100 | F: GCCTGAATGTGCCATAAGACTGAC |
| R: AAACCCACAGTGCTTGACACAGA | ||
| Peroxisome proliferator-activated receptor alpha (PPARα) | NM_001001928 | F: ACGGAAAGCCCACTCTGCCCCCTCTC |
| R: CTTGTCCCCGCAGATTCTACATTCG | ||
| Peroxisome proliferator-activated receptor beta/delta (PPARδ) | NM_001171818 | F: GGCCATCATTCTGTGTGGAGAC |
| R: CAGGATGGTGTCCTGGATAGC | ||
| Peroxisome proliferator-activated receptor (PPARγ) | NM_138712 | F: GCCATTTTCTCAAACGAGAGTCAGC |
| R: CCACGGAGCTGATCCCAAAGTT |

References
- Wittassek, M.; Koch, H.M.; Angerer, J.; Bruning, T. Assessing exposure to phthalates—The human biomonitoring approach. Mol. Nutr. Food Res. 2011, 55, 7–31. [Google Scholar] [CrossRef] [PubMed]
- Kay, V.R.; Chambers, C.; Foster, W.G. Reproductive and developmental effects of phthalate diesters in females. Crit. Rev. Toxicol. 2013, 43, 200–219. [Google Scholar] [CrossRef] [PubMed]
- Lyche, J.L.; Gutleb, A.C.; Bergman, A.; Eriksen, G.S.; Murk, A.J.; Ropstad, E. Reproductive and developmental toxicity of phthalates. J. Toxicol. Environ. Health B Crit. Rev. 2009, 12, 225–249. [Google Scholar] [CrossRef] [PubMed]
- Kavlock, R.; Boekelheide, K.; Chapin, R.; Cunningham, M.; Faustman, E.; Foster, P. NTP center for the evaluation of risks to human reproduction: Phthalates expert panel report on the reproductive and developmental toxicity of di-n-octyl phthalate. Reprod. Toxicol. 2002, 16, 721–734. [Google Scholar] [CrossRef]
- Wolff, M.S.; Engel, S.M.; Berkowitz, G.S.; Ye, X.; Silva, M.J.; Zhu, C. Prenatal phenol and phthalate exposures and birth outcomes. Environ. Health Perspect. 2008, 116, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Weuve, J.; Hauser, R.; Calafat, A.M.; Missmer, S.A.; Wise, L.A. Association of exposure to phthalates with endometriosis and uterine leiomyomata: findings from NHANES, 1999–2004. Environ. Health Perspect. 2010, 118, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Hannon, P.R.; Flaws, J.A. The effects of phthalates on the ovary. Front. Endocrinol. (Lausanne) 2015, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Barrett, E.S.; Parlett, L.E.; Wang, C.; Drobnis, E.Z.; Redmon, J.B.; Swan, S.H. Environmental exposure to di-2-ethylhexyl phthalate is associated with low interest in sexual activity in premenopausal women. Horm. Behav. 2014, 66, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Chun, S.; Jang, J.Y.; Chae, H.D.; Kim, C.H.; Kang, B.M. Increased plasma levels of phthalate esters in women with advanced-stage endometriosis: A prospective case-control study. Fert. Steril. 2011, 95, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Upson, K.; Sathyanarayana, S.; De Roos, A.J.; Thompson, M.L.; Scholes, D.; Dills, R. Phthalates and risk of endometriosis. Environ. Res. 2013, 126, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Buck Louis, G.M.; Peterson, C.M.; Chen, Z.; Croughan, M.; Sundaram, R.; Stanford, J. Bisphenol A and phthalates and endometriosis: The endometriosis: Natural history, diagnosis and outcomes study. Fert. Steril. 2013, 100, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Iwasaki, M.; Hanaoka, T.; Sasaki, H.; Tanaka, T.; Tsugane, S. Urinary phthalate monoesters and endometriosis in infertile Japanese women. Sci. Total Environ. 2009, 408, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Kim, S.H.; Lee, H.W.; Chae, H.D.; Kim, C.H.; Kang, B.M. Increased viability of endometrial cells by in vitro treatment with di-(2-ethylhexyl) phthalate. Fert. Steril. 2010, 94, 2413–2416. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Valbuena, D.; Krussel, J.; Bernal, A.; Murphy, C.R.; Shaw, T. Interleukin-1 receptor antagonist prevents embryonic implantation by a direct effect on the endometrial epithelium. Fert. Steril. 1998, 70, 896–906. [Google Scholar] [CrossRef]
- Guay, S.; Akoum, A. Stable inhibition of interleukin 1 receptor type II in Ishikawa cells augments secretion of matrix metalloproteinases: Possible role in endometriosis pathophysiology. Reproduction 2007, 134, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Ren, P.; Zhang, Y.; Huang, Y.; Yang, Y.; Jiang, M. Functions of peroxisome proliferator-activated receptor gamma (PPARgamma) in gynecologic disorders. Clin. Med. Insights Oncol. 2015, 9, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Knapp, P.; Chabowski, A.; Blachnio-Zabielska, A.; Jarzabek, K.; Wolczynski, S. Altered peroxisome-proliferator activated receptors expression in human endometrial cancer. PPAR Res. 2012, 2012, 471524. [Google Scholar] [CrossRef] [PubMed]
- Feige, J.N.; Gerber, A.; Casals-Casas, C.; Yang, Q.; Winkler, C.; Bedu, E. The pollutant diethylhexyl phthalate regulates hepatic energy metabolism via species-specific PPAR alpha-dependent mechanisms. Environ. Health Perspect. 2010, 118, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wu, R.F.; Su, L.; Zhou, W.D.; Zhu, M.B.; Chen, Q.H. Lipoxin A4 regulates expression of the estrogen receptor and inhibits 17beta-estradiol induced P38 mitogen-activated protein kinase phosphorylation in human endometriotic stromal cells. Fert. Steril. 2014, 10, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.; Akatsuka, Y.; Kurihara, R.; Matsuoka, S.; Nakatsukuri, M.; Kurokawa, Y. Evaluation of the Ishikawa cell line bioassay for the detection of estrogenic substances from sediment extracts. Environ. Toxicol. Chem. 2005, 24, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Frederiksen, H.; Kuiri-Hanninen, T.; Main, K.M.; Dunkel, L.; Sankilampi, U. A longitudinal study of urinary phthalate excretion in 58 full-term and 67 preterm infants from birth through 14 months. Environ. Health Perspect. 2014, 122, 998–1005. [Google Scholar] [CrossRef] [PubMed]
- Akingbemi, B.T.; Ge, R.S.; Klinefelter, G.R.; Zirkin, B.R.; Hardy, M.P. Phthalate-induced Leydig cell hyperplasia is associated with multiple endocrine disturbances. Proc. Natl. Acad. Sci. USA 2004, 101, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Craig, Z.R.; Basavarajappa, M.S.; Gupta, R.K.; Flaws, J.A. Di-(2-ethylhexyl) phthalate inhibits growth of mouse ovarian antral follicles through an oxidative stress pathway. Toxicol. Appl. Pharm. 2012, 258, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shang, L.; Wang, J.; Wu, N.; Wang, S. Effect of phthalate esters on the secretion of prostaglandins (F2alpha and E2) and oxytocin in cultured bovine ovarian and endometrial cells. Domest. Anim. Endocrinol. 2010, 39, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Kayisli, U.A.; Guzeloglu-Kayisli, O.; Arici, A. Endocrine-immune interactions in human endometrium. Ann. N.Y. Acad. Sci. 2004, 1034, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Campioli, E.; Martinez-Arguelles, D.B.; Papadopoulos, V. In utero exposure to the endocrine disruptor di-(2-ethylhexyl) phthalate promotes local adipose and systemic inflammation in adult male offspring. Nutr. Diabetes 2014, 4, e115. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, D.; Marques, C.; Pestana, D.; Faria, A.; Norberto, S.; Calhau, C. Effects of xenoestrogens in human M1 and M2 macrophage migration, cytokine release, and estrogen-related signaling pathways. Environ. Toxicol. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, J.; Iwahara, C.; Kawasaki, M.; Yoshizaki, F.; Nakayama, H.; Takamori, K. Di-(2-ethylhexyl) phthalate induces production of inflammatory molecules in human macrophages. Inflamm. Res. 2012, 61, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Ohashi, Y.; Ishikawa, K.; Yasue, A.; Kato, R.; Achiwa, Y. Expression of cyclooxygenase-2 in uterine endometrial cancer and anti-tumor effects of a selective COX-2 inhibitor. Int. J. Oncol. 2005, 26, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Novoa, J.M.; Nieto, M.A. Inflammation and EMT: An alliance towards organ fibrosis and cancer progression. EMBO Mol. Med. 2009, 1, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P. Epithelial-mesenchymal transitions in tumour progression. Nat. Rev. Cancer. 2002, 2, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Yun, Y.; Gao, R.; Li, G.; Sang, N. Winter polycyclic aromatic hydrocarbon-bound particulate matter from peri-urban north China promotes lung cancer cell metastasis. Environ. Sci. Technol. 2015, 49, 14484–14493. [Google Scholar] [CrossRef] [PubMed]
- Kopantzev, E.P.; Vayshlya, N.A.; Kopantseva, M.R.; Egorov, V.I.; Pikunov, M.; Zinovyeva, M.V. Cellular and molecular phenotypes of proliferating stromal cells from human carcinomas. Br. J. Cancer 2010, 102, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Daynes, R.A.; Jones, D.C. Emerging roles of PPARs in inflammation and immunity. Nat. Rev. Immunol. 2002, 2, 748–759. [Google Scholar] [CrossRef] [PubMed]




© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Q.; Zhang, H.; Chen, Y.-J.; Chi, Y.-L.; Dong, S. The Inflammation Response to DEHP through PPARγ in Endometrial Cells. Int. J. Environ. Res. Public Health 2016, 13, 318. https://doi.org/10.3390/ijerph13030318
Huang Q, Zhang H, Chen Y-J, Chi Y-L, Dong S. The Inflammation Response to DEHP through PPARγ in Endometrial Cells. International Journal of Environmental Research and Public Health. 2016; 13(3):318. https://doi.org/10.3390/ijerph13030318
Chicago/Turabian StyleHuang, Qiansheng, Huanteng Zhang, Ya-Jie Chen, Yu-Lang Chi, and Sijun Dong. 2016. "The Inflammation Response to DEHP through PPARγ in Endometrial Cells" International Journal of Environmental Research and Public Health 13, no. 3: 318. https://doi.org/10.3390/ijerph13030318