
Complete Genomic Characterization of Porcine Reproductive and Respiratory Syndrome Virus Strain HB-XL

Abstract
:1. Introduction
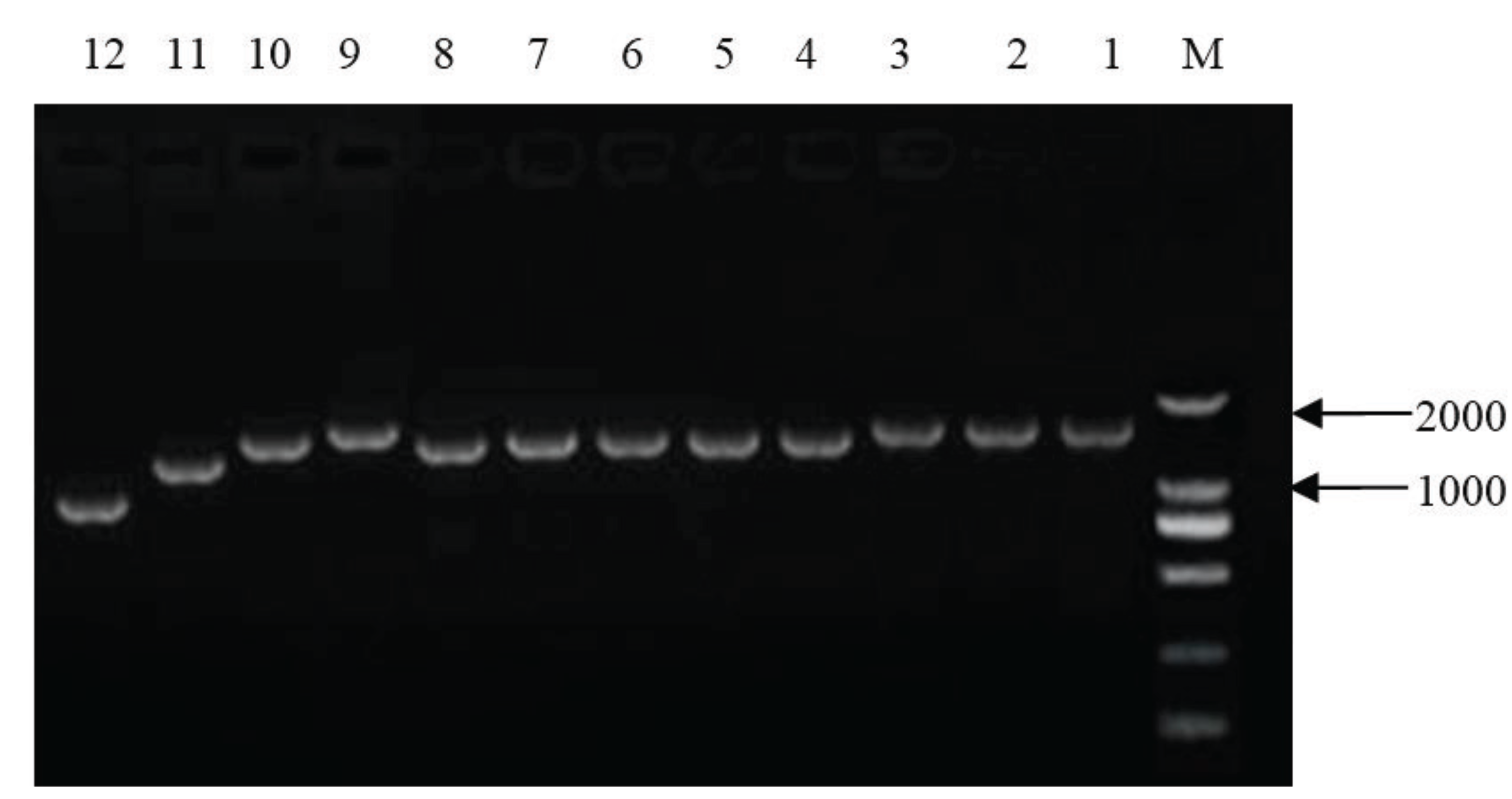
2. Materials and Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers Name | Sequence (5'–3') | Position a |
|---|---|---|
| PRRSV1F | ATGACGTATAGGTGTTGGCTCTATG | 1–1562 |
| PRRSV1R | AGGGTGGTCTCAAAGTTGGAAT | |
| PRRSV2F | ATTTCCGCCATCGTCAACCG | 1513–3040 |
| PRRSV2R | TCTGGTGCGTCAGCGTTGTTGT | |
| PRRSV3F | ACACCTATGAGTGAGCCCGTAC | 2827–4324 |
| PRRSV3R | CAACAATACCAAGCCTAAGCAA | |
| PRRSV4F | TGCTTAGGCTTGGCATTGTTG | 4304–5656 |
| PRRSV4R | TGACGGTGTTCAGTGAGGGCT | |
| PRRSV5F | CACCGCACCAGATGGAACCTACT | 5427–6890 |
| PRRSV5R | GTGCCTCGGACCTTATCAACCTGT | |
| PRRSV6F | AGTGCTTTGTTTCTGCGTCCAAC | 6770–8241 |
| PRRSV6R | GAATGGTCGGCACATACAACTCA | |
| PRRSV7F | CCTGCCTCACGCCTAATGCTA | 8233–9638 |
| PRRSV7R | CCCGCACATTCTGGACTTCTTC | |
| PRRSV8F | TTCCTGGCCCACCGTTCTTCTT | 9559–10861 |
| PRRSV8R | AGAGCCTGAGCAACCGTGATTT | |
| PRRSV9F | GCAAGACATGCTATCTTCGTGT | 10704–12151 |
| PRRSV9R | ATGGACACCAGAAATTCCGTGA | |
| PRRSV10F | CCCTGTCATTGAACCAACTTTA | 12035–13533 |
| PRRSV10R | AGCATGAGGAGGTCAGAAGAAT | |
| PRRSV11F | AACACCACCGCAGCATCAAACT | 13346–14685 |
| PRRSV11R | CCTAGCAAGCACAAACGGCATC | |
| PRRSV12F | TTCAGAGCACAAATAGGGTCGCG | 14565–15408 |
| PRRSV12R | TAATTGCGGCCGCATGGTTCTC |
3. Results
3.1. Genomic Characteristics on the HB-XL Strain

3.2. Homology Analyses
| No. | Name | Country | Year | Accession Number |
|---|---|---|---|---|
| 1 | CH-1a | China | 1996 | AY032626 |
| 2 | BJ-4 | China | 2000 | AF331831 |
| 3 | HB-1(sh)/2002 | China | 2002 | AY150312 |
| 4 | JXA1 | China | 2006 | EF112445 |
| 5 | HUB2 | China | 2006 | EF112446 |
| 6 | HuN4 | China | 2007 | EF635006 |
| 7 | SY0608 | China | 2007 | EU144079 |
| 8 | CH-1R | China | 2008 | EU807840 |
| 9 | HB-1 | China | 2008 | EU360130 |
| 10 | WUH4 | China | 2011 | JQ326271 |
| 11 | QYYZ | China | 2011 | JQ308798 |
| 12 | SD16 | China | 2012 | JX087437 |
| 13 | JXA1-R | China | 2012 | JQ804986 |
| 14 | HEB-2013 | China | 2013 | KJ591659 |
| 15 | HB-XL | China | 2013 | KP162169 |
| 16 | LMY | South Korea | 2002 | DQ473474 |
| 17 | VR-2332 | USA | 1992 | AY150564 |
| 18 | EuroPRRSV | USA | 1999 | AY366525 |
| 19 | RespPRRS MLV | USA | 1994 | AF066183 |
| 20 | Lelystad virus | Europe | 1991 | M96262 |
| HB-XL % | Identity to SH1211 | |||||
|---|---|---|---|---|---|---|
| HB-1(sh)/2002 | JXA1 | CH-1a | QYYZ | VR-2332 | LV | |
| Nucleotides (length) | ||||||
| °5'UTR (188) | 96.8 | 97.4 | 97.3 | 95.2 | 93.1 | 50.1 |
| °ORF1a (7422) | 96.6 | 99.1 | 94.0 | 85.2 | 87.5 | 59.3 |
| °ORF1b (4383) | 97.5 | 99.4 | 92.8 | 90.3 | 91.1 | 61.5 |
| °ORF2 (771) | 96.2 | 98.6 | 94.7 | 90.0 | 93.1 | 67.1 |
| °ORF3 (759) | 95.4 | 98.8 | 92.9 | 90.8 | 88.9 | 61.4 |
| °ORF4 (537) | 96.8 | 98.7 | 93.9 | 94.4 | 89.9 | 67.6 |
| °ORF5 (603) | 96.5 | 99.0 | 95.0 | 83.9 | 88.9 | 63.7 |
| °ORF6 (525) | 97.1 | 99.8 | 96.3 | 91.0 | 95.6 | 69.2 |
| °ORF7 (372) | 96.5 | 99.7 | 94.8 | 89.5 | 94.4 | 65.9 |
| °3'UTR (150) | 93.3 | 97.3 | 93.7 | 87.3 | 90.7 | 55.9 |
| Complete (15,323) | 96.9 | 99.1 | 95.0 | 87.7 | 89.5 | 61.7 |
| Amino acid (length) | ||||||
| °Nsp2 (950) | 91.1 | 97.1 | 84.9 | 74.2 | 74.8 | 30.0 |
| °GP2 (257) | 94.5 | 97.7 | 94.9 | 89.5 | 92.2 | 63.1 |
| °E (74) | 94.5 | 97.5 | 94.5 | 89.0 | 92.0 | 63.0 |
| °GP3 (253) | 92.9 | 93.8 | 89.3 | 87.4 | 88.6 | 54.9 |
| °GP4 (179) | 97.8 | 96.1 | 97.8 | 94.9 | 90.4 | 70.9 |
| °GP5 (201) | 92.5 | 97.5 | 91.0 | 81.6 | 86.6 | 56.2 |
| °M (175) | 98.3 | 99.4 | 98.3 | 96.6 | 98.3 | 79.8 |
| °N (124) | 95.9 | 100 | 95.1 | 91.9 | 95.1 | 58.1 |
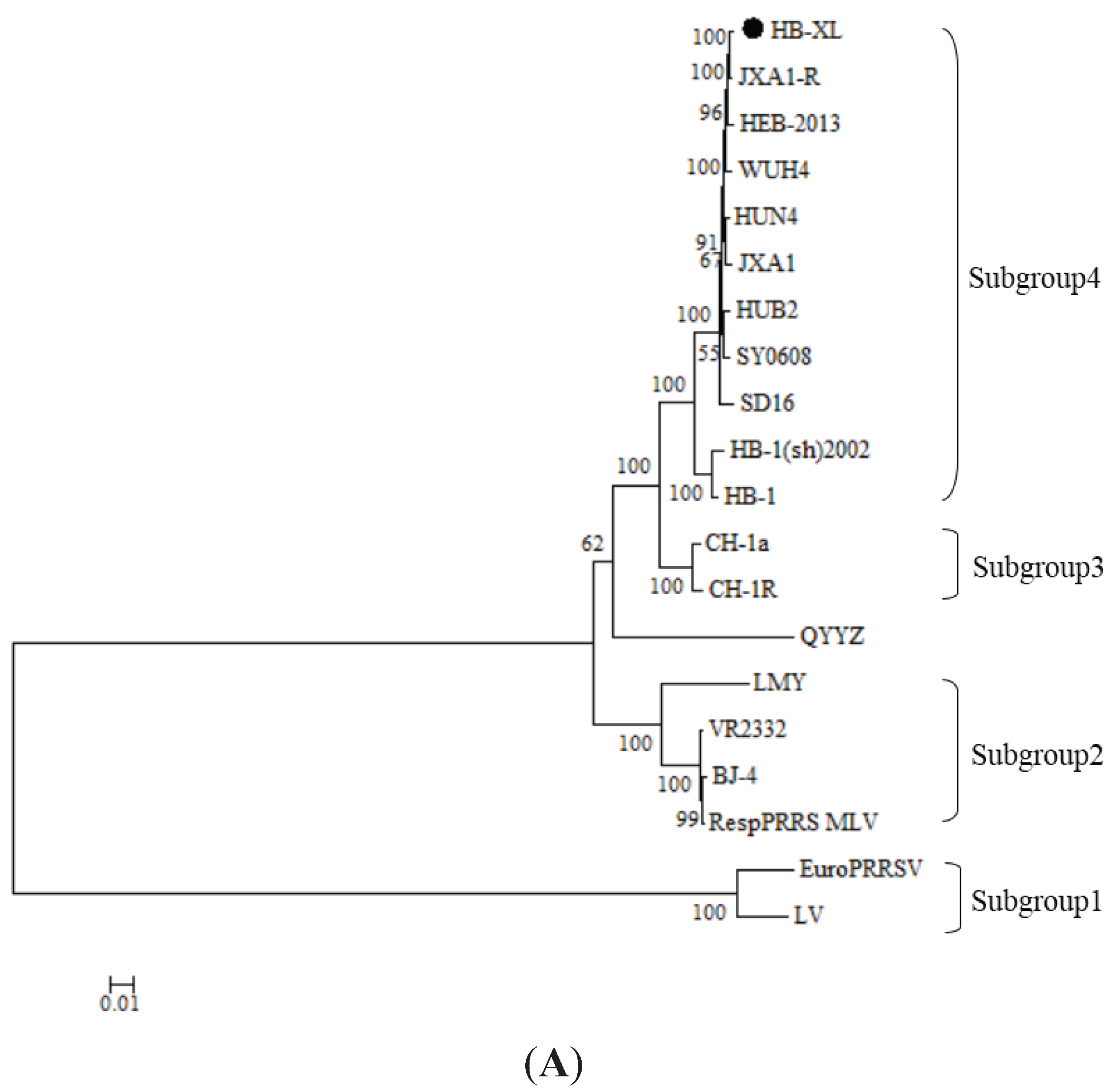
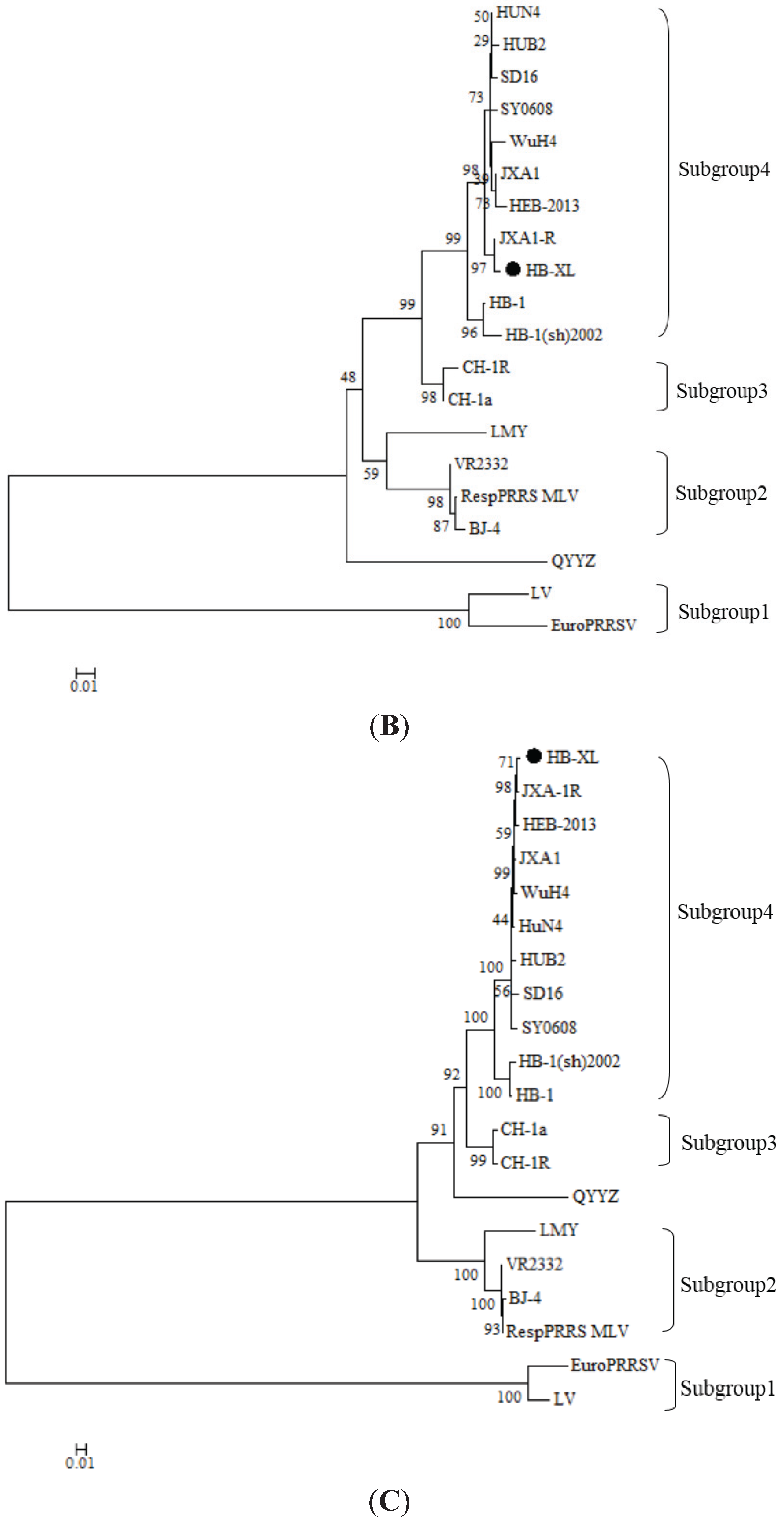
3.3. Phylogenetic Analyses


3.4. Variation Analyses

| Strain Name | N-Glycosylation Sites | ||||||
|---|---|---|---|---|---|---|---|
| 30 | 32 | 33 | 34 | 35 | 44 | 51 | |
| CH-1a | N | N | S | N | S | N | N |
| BJ-4 | N | S | N | D | S | N | N |
| HB-1(sh)/2002 | N | S | N | N | S | N | N |
| JXA1 | N | S | N | N | N | N | N |
| HUB2 | N | S | N | N | N | N | N |
| HuN4 | N | S | N | N | N | N | N |
| SY0608 | N | S | N | N | N | N | N |
| CH-1R | N | N | S | N | S | N | N |
| HB-1 | N | S | N | N | N | N | N |
| WUH4 | N | S | N | N | N | N | N |
| QYYZ | S | N | G | N | S | N | N |
| SD16 | N | S | N | N | N | N | N |
| JXA1-R | N | S | D | N | N | N | N |
| HEB-2013 | N | S | N | N | N | N | N |
| HB-XL | N | S | G | N | N | N | N |
| LMY | S | N | N | S | S | N | N |
| VR-2332 | N | S | N | D | S | N | N |
| EuroPRRSV | N | S | N | D | S | N | N |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gorbalenya, A.E.; Enjuanes, L.; Ziebuhr, J.; Snijder, E.J. Nidovirales: Evolving the largest RNA virus genome. Virus Res. 2006, 117, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Benfield, D.A.; Nelson, E.; Collins, J.E.; Harris, L.; Goyal, S.M.; Robison, D.; Christianson, W.T.; Morrison, R.B.; Gorcyca, D.; Chladek, D. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Investig. 1992, 4, 127–133. [Google Scholar] [CrossRef]
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; Laak, E.A.T.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- An, T.Q.; Tian, Z.J.; Leng, C.L.; Peng, J.M.; Tong, G.Z. Highly pathogenic porcine reproductive and respiratory syndrome virus, Asia. Emerg. Infect. Dis. 2011, 17, 1782–1784. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Fang, L.; Guo, X.; Gao, J.; Song, T.; Bi, J.; He, K.; Chen, H.; Xiao, S. Epidemiology and evolutionary characteristics of the porcine reproductive and respiratory syndrome virus in China between 2006 and 2010. J. Clin. Microbiol. 2011, 49, 3175–3183. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.Q.; Yang, S.B.; Bounlom, D.; Yu, X.; Zhou, Z.; Song, J.; Khamphouth, V.; Vatthana, T.; Tian, K. Emergence and pathogenicity of highly pathogenic porcine reproductive and respiratory syndrome virus in Vientiane, Lao People’s Democratic Republic. J. Vet. Diagn. Investig. 2012, 24, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhou, R.; Zhang, J.; Zhou, L.; Jiang, Q.; Guo, X.; Ge, X.; Yang, H. Recombination analyses between two strains of porcine reproductive and respiratory syndrome virus in vivo. Virus Res. 2011, 155, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, N.; Han, W.; Cao, Z.; Deng, X.; Wang, L.; Yu, X.; Zhou, Z.; Li, X.; Shi, J.; et al. Complete genome sequence of an overattenuated highly Pathogenic porcine reproductive and respiratory syndrome virus. J. Virol. 2012, 86, 6381–6382. [Google Scholar] [CrossRef] [PubMed]
- Meulenberg, J.J.; Ruijter, J.N.B.; Wensvoort, G.; Moormann, R.J. An infectious cDNA clone of porcine reproductive and respiratory syndrome virus. Adv. Exp. Med. Biol. 1998, 440, 199–206. [Google Scholar] [PubMed]
- Dea, S.; Gagnon, C.A.; Mardassi, H.; Pirzadeh, B.; Rogan, D. Current knowledge on the structural proteins of porcine reproductive and respiratory syndrome (PRRS) virus: Comparison of the North American and European isolates. Arch. Virol. 2000, 145, 659–688. [Google Scholar] [CrossRef] [PubMed]
- Ropp, S.L.; Wees, C.E.; Fang, Y.; Nelson, E.A.; Rossow, K.D.; Bien, M.; Arndt, B.; Preszler, S.; Steen, P.; Christopher-Hennings, J.; et al. Characterization of emerging European-like porcine reproductive and respiratory syndrome virus isolates in the United States. J. Virol. 2004, 78, 3684–3703. [Google Scholar] [CrossRef] [PubMed]
- Stadejek, T.; Oleksiewicz, M.B.; Potapchuk, D.; Podgórska, K. Porcine reproductive and respiratory syndrome virus strains of exceptional diversity in Eastern Europe support the definition of new genetic subtypes. J. Gen. Virol. 2006, 87, 1835–1841. [Google Scholar] [CrossRef] [PubMed]
- Nelsen, C.J.; Murtaugh, M.P.; Faaberg, K.S. Porcine reproductive and respiratory syndrome virus comparison: Divergent evolution on two continents. J. Virol. 1999, 73, 270–280. [Google Scholar] [PubMed]
- Fan, B.; Wang, H.; Bai, J.; Zhang, L.; Jiang, P. A Novel isolate with deletion in GP3 gene of porcine reproductive and respiratory syndrome virus from Mid-Eastern China. Biomed. Res. Int. 2014. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lam, T.T.; Hon, C.C.; Murtaugh, M.P.; Davies, P.R.; Hui, R.K.; Li, J.; Wong, L.T.; Yip, C.W.; Jiang, J.W.; et al. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef] [PubMed]
- Allende, R.; Kutish, G.F.; Laegreid, W.; Lu, Z.; Lewis, T.L.; Rock, D.L.; Friesen, J.; Galeota, J.A.; Doster, A.R.; Osorio, F.A. Mutations in the genome of porcine reproductive and respiratory syndrome virus responsible for the attenuation phenotype. Arch. Virol. 2000, 145, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wei, C.; Yang, X.Y.; Hou, X.L.; Dai, A.L.; Li, X.H.; Wei, M.K.; Pan, X.Z. Genetic diversity and evolutionary characterization of Chinese porcine reproductive and respiratory syndrome viruses based on NSP2 and ORF5. Arch. Virol. 2013, 158, 1811–1816. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Benfield, D.; Rowland, R.R. Porcine reproductive and respiratory syndrome virus: An update on an emerging and re-emerging viral disease of swine. Virus Res. 2010, 154, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.P.; Elam, M.R.; Kakach, L.T. Comparison of the structural protein coding sequences of the VR-2332 and Lelystad virus strains of the PRRS virus. Arch. Virol. 1995, 140, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.H. Immunodominant epitopes in Nsp2 of porcine reproductive and respiratory syndrome virus are dispensable for replication, but play an important role in modulation of the host immune response. J. Gen. Virol. 2010, 91, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, Y.; Faaberg, K.S. Complete genome analysis of RFLP 184 isolates of porcine reproductive and respiratory syndrome virus. Virus Res. 2006, 122, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L. The 30-amino-acid deletion in the Nsp2 of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in China is not related to its virulence. J. Virol. 2009, 83, 5156–5167. [Google Scholar] [CrossRef] [PubMed]
- Li, B. Immunogenicity of the highly pathogenic porcine reproductive and respiratory syndrome virus GP5 protein encoded by a synthetic ORF5 gene. Vaccine 2009, 27, 1957–1963. [Google Scholar] [CrossRef] [PubMed]
- An, T.Q. Genetic diversity and phylogenetic analysis of glycoprotein 5 of PRRSV isolates in mainland China from 1996 to 2006: Coexistence of two NA-subgenotypes with great diversity. Vet. Microbiol. 2007, 123, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, M.; Galeota, J.A.; Jar, A.M.; Platt, K.B.; Osorio, F.A.; Lopez, O.J. Identification of neutralizing and nonneutralizing epitopes in the porcine reproductive and respiratory syndrome virus GP5 ectodomain. J. Virol. 2002, 76, 4241–4250. [Google Scholar] [CrossRef] [PubMed]
- Faaberg, K.S. Neutralizing antibody responses of pigs infected with natural GP5 N-glycan mutants of porcine reproductive and respiratory syndrome virus. Viral Immunol. 2006, 19, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.S.; Yang, X.; Wang, H.N. Molecular characterization of a complete genome and 12 Nsp2 genes of PRRSV of southwestern China. Food Environ. Virol. 2012, 4, 102–114. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, Y.; Yuan, W.; Sun, J. Complete Genomic Characterization of Porcine Reproductive and Respiratory Syndrome Virus Strain HB-XL. Genes 2015, 6, 672-684. https://doi.org/10.3390/genes6030672
Zuo Y, Yuan W, Sun J. Complete Genomic Characterization of Porcine Reproductive and Respiratory Syndrome Virus Strain HB-XL. Genes. 2015; 6(3):672-684. https://doi.org/10.3390/genes6030672
Chicago/Turabian StyleZuo, Yi, Wanzhe Yuan, and Jiguo Sun. 2015. "Complete Genomic Characterization of Porcine Reproductive and Respiratory Syndrome Virus Strain HB-XL" Genes 6, no. 3: 672-684. https://doi.org/10.3390/genes6030672