
Hedgehog Signaling during Appendage Development and Regeneration

Abstract
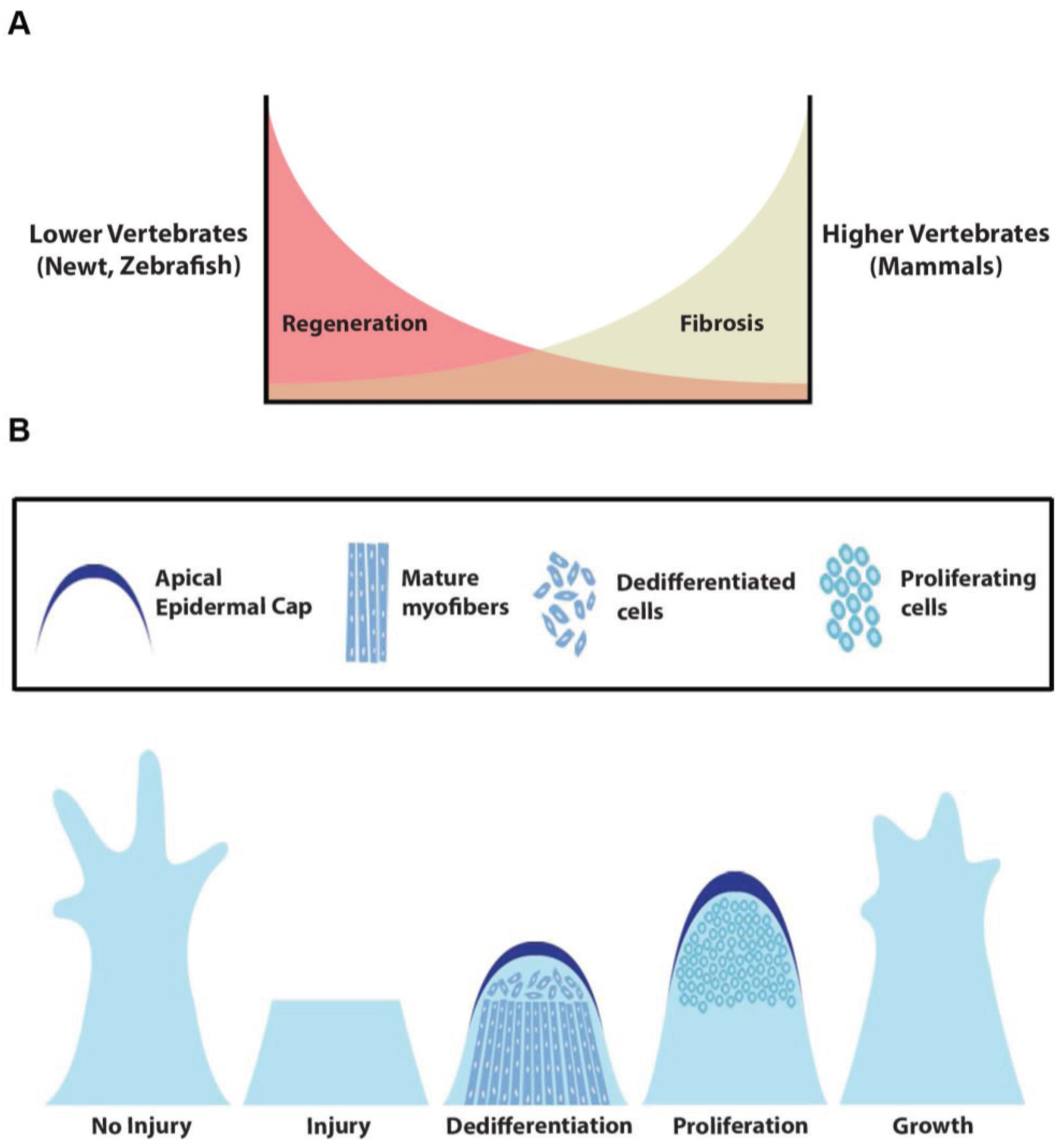
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
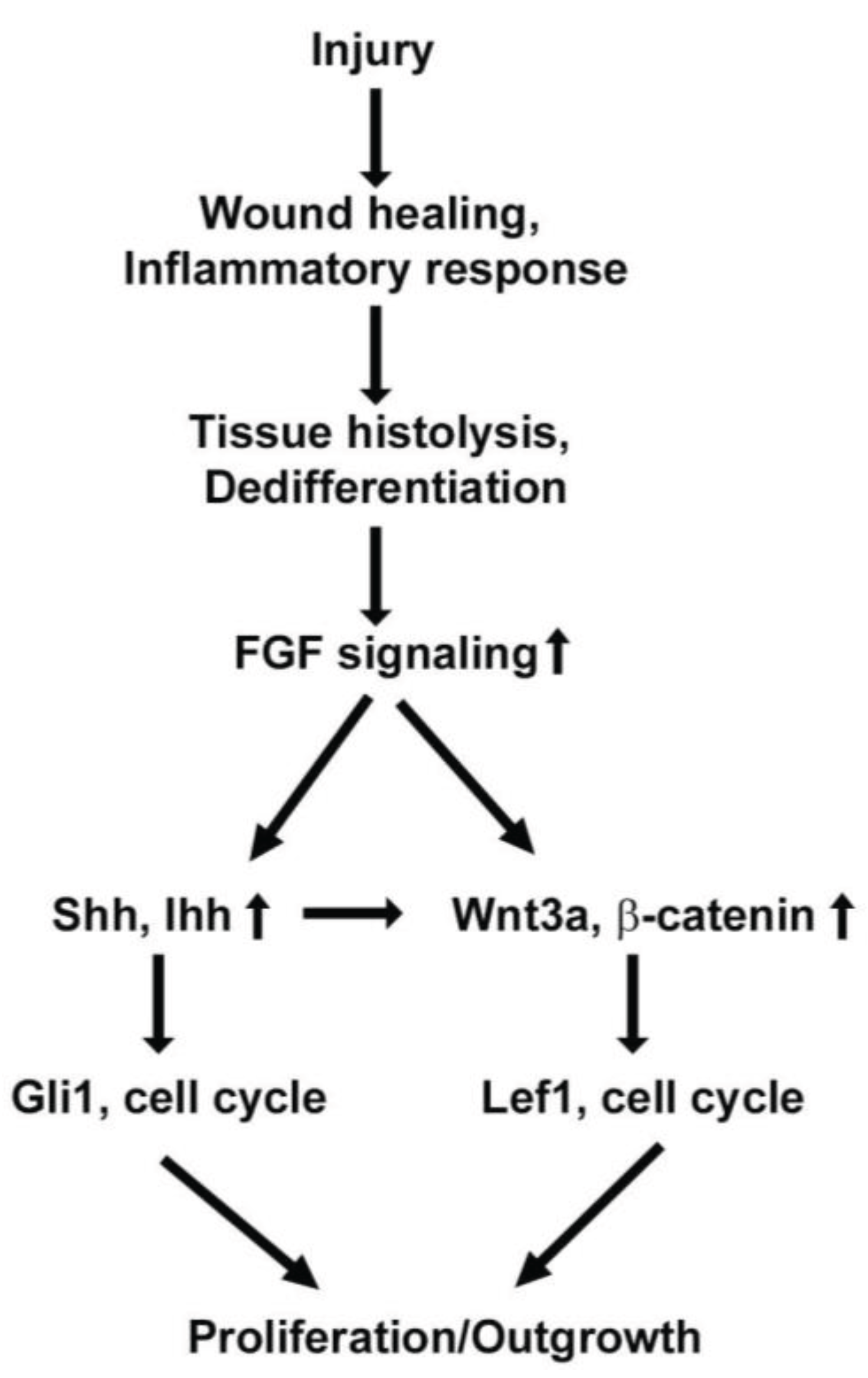{kind=link}
| Tissue | Species | Extent of Regeneration | Signaling Pathways |
|---|---|---|---|
| Tail | |||
| Mexican axolotl | WNT, BMP, NOTCH, SHH [8,10,11,12,13,14,15] | ||
| (Ambystoma mexicanum) | Complete | ||
| Xenopus | |||
| (Xenopus laevis) | |||
| Larval Stage | Complete | ||
| Newt | |||
| (Notophathalmus viridescencs) | Complete | ||
| Limb | |||
| Mexican axolotl | FGF, WNT, NOTCH, SHH, RA, BMP [16,17,18,19,20,21,22,23] | ||
| (Ambystoma mexicanum) | Complete | ||
| Xenopus | |||
| (Xenopus laevis) | |||
| Larval Stage | Complete | ||
| Adult Stage | Spike (Incomplete) | ||
| Newt | |||
| (Notophathalmus viridescencs) | Complete | ||
| Fin | |||
| Zebrafish (Danio rerio) | Complete | FGF, WNT, NOTCH, SHH, RA, BMP [17,24,25,26,27,28,29] | |
| Heart | |||
| Newt | FGF, NOTCH, RA [6,9,13,30,31,32] | ||
| (Notophathalmus viridescencs) | Complete | ||
| Zebrafish | |||
| (Danio rerio) | Complete | ||
| Mouse | |||
| (Mus musculus) | Complete | ||
| Neonatal Heart | Scar formation | ||
| Adult Heart |

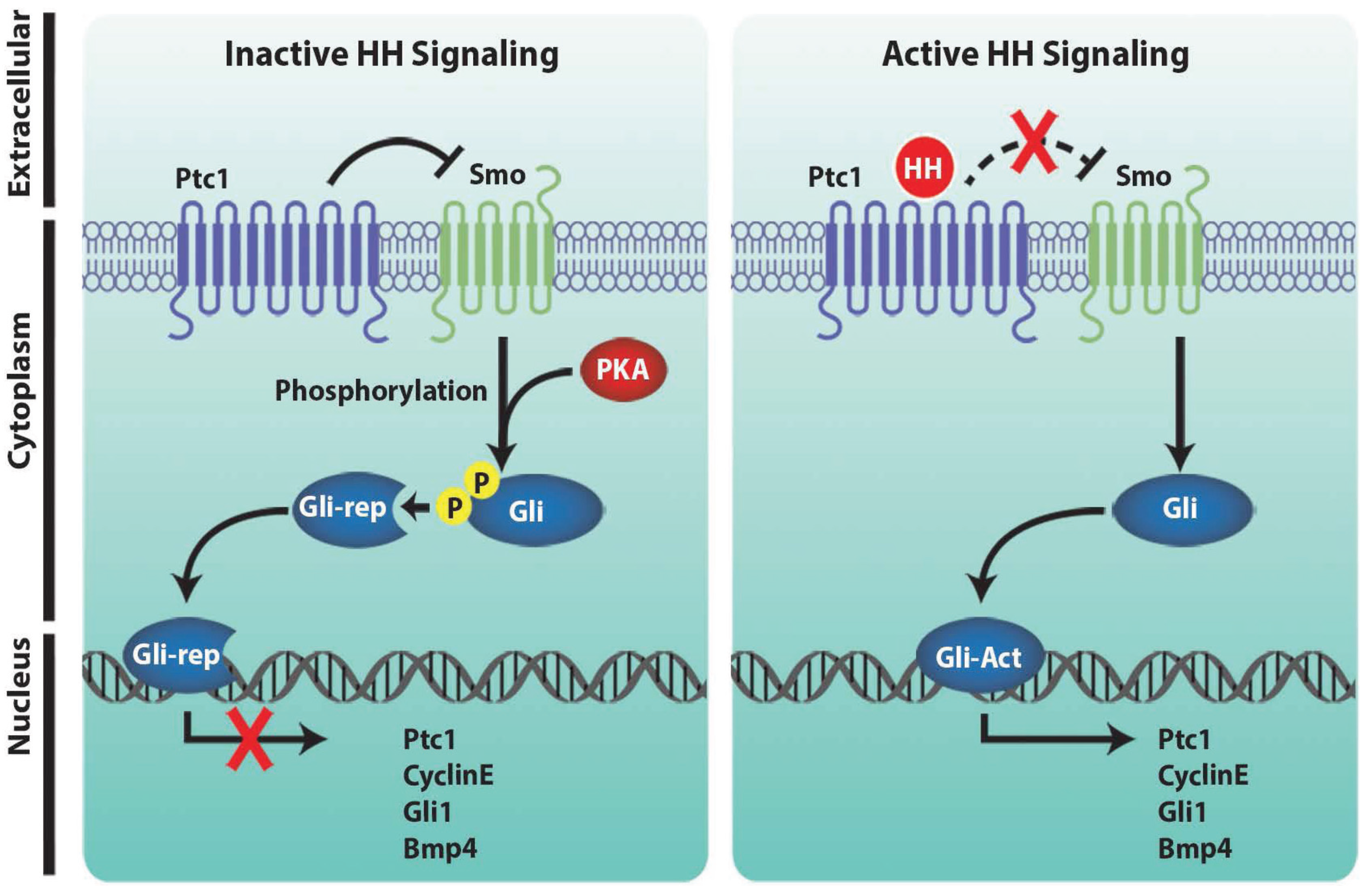
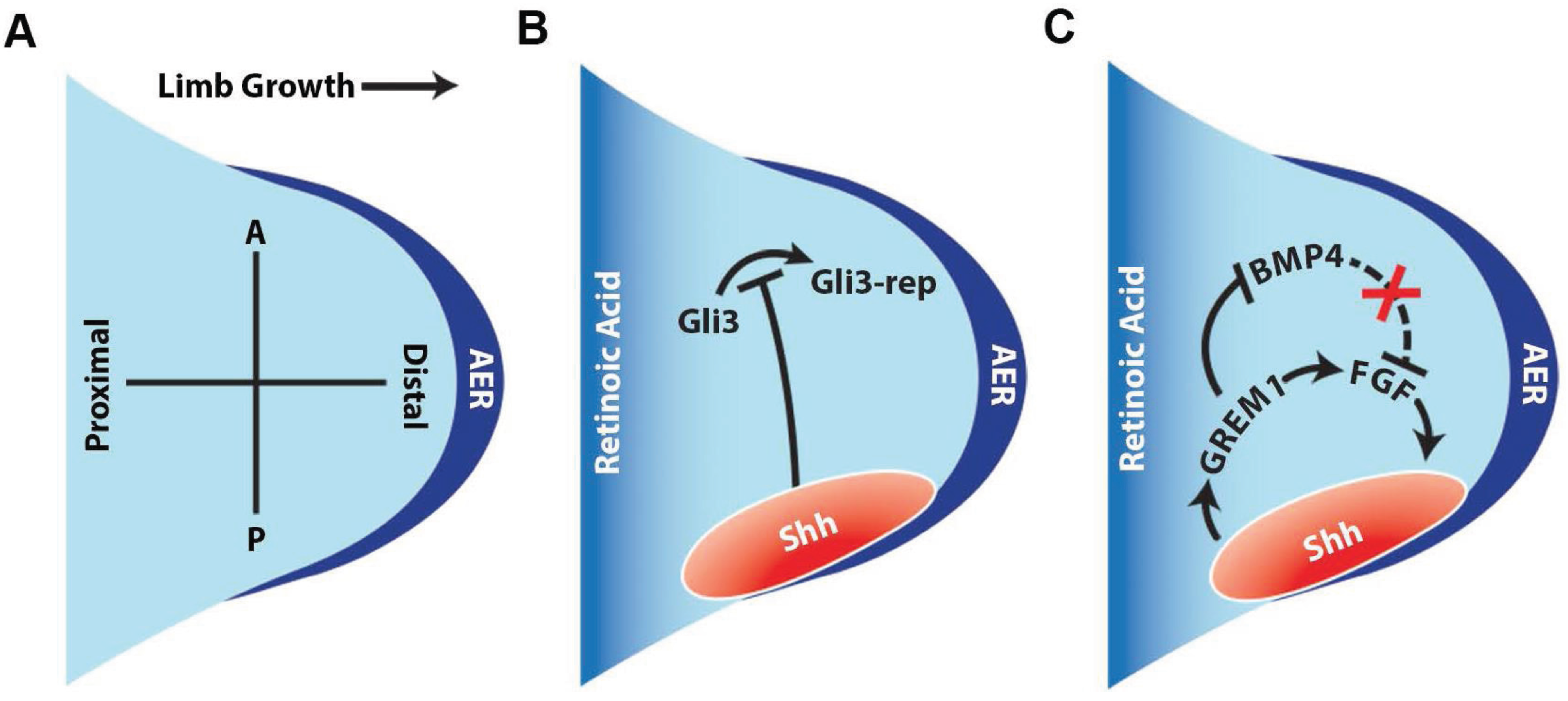
2. Hedgehog Signaling Pathways during Limb Development
| Genotype | Lethality | Phenotype |
|---|---|---|
| Shh-/- | Embryonic lethality (E11.5–E18.5) | Midline structural defects Defective distal structure Dorsoventral patterning defects [41,54] |
| Ihh-/- | Partial embryonic Lethality | Skeletal defects Mesenchymal loss Chondrocyte proliferation defects [41,55] |
| Smo-/- | Embryonic lethality (E9.5–E10.5) | Midline structural defects Cardio-vascular defects L/R asymmetry defects [41,67,68] |
| Shh-/-; Ihh-/- | Embryonic lethality (E9.5–E10.5) | Midline structural defects Cardio-vascular defects Abnormal forebrain Patterning defects [41,64,67,68] |
| Ptc1-/- | Embryonic lethality (E9.5–E10.5) | Open neural tube defects Cardiac morphogenesis defects [69] |
| Gli1-/- | Viable | No obvious phenotype [66] |
| Gli2-/- | Embryonic lethality (E15.5–E18.5) | Defective lung outgrowth [66] |
| Gli1-/-; Gli2-/- | Embryonic lethality (E15.5–E18.5) | Defective growth and patterning of lung lobes Notochord regression defects Defective spinal cord ventral midline [66] |
| Shh Morpholino (Zebrafish) | Reduced myoseptum Defective somitic patterning Partial cylopia [70] | |
| Msx2-Cre;SmoL/L | Lethality (birth) | Patterning defects [60,71] |
| Prx1-Cre;PtcL/L | Viable | Patterning defects [41,69] |


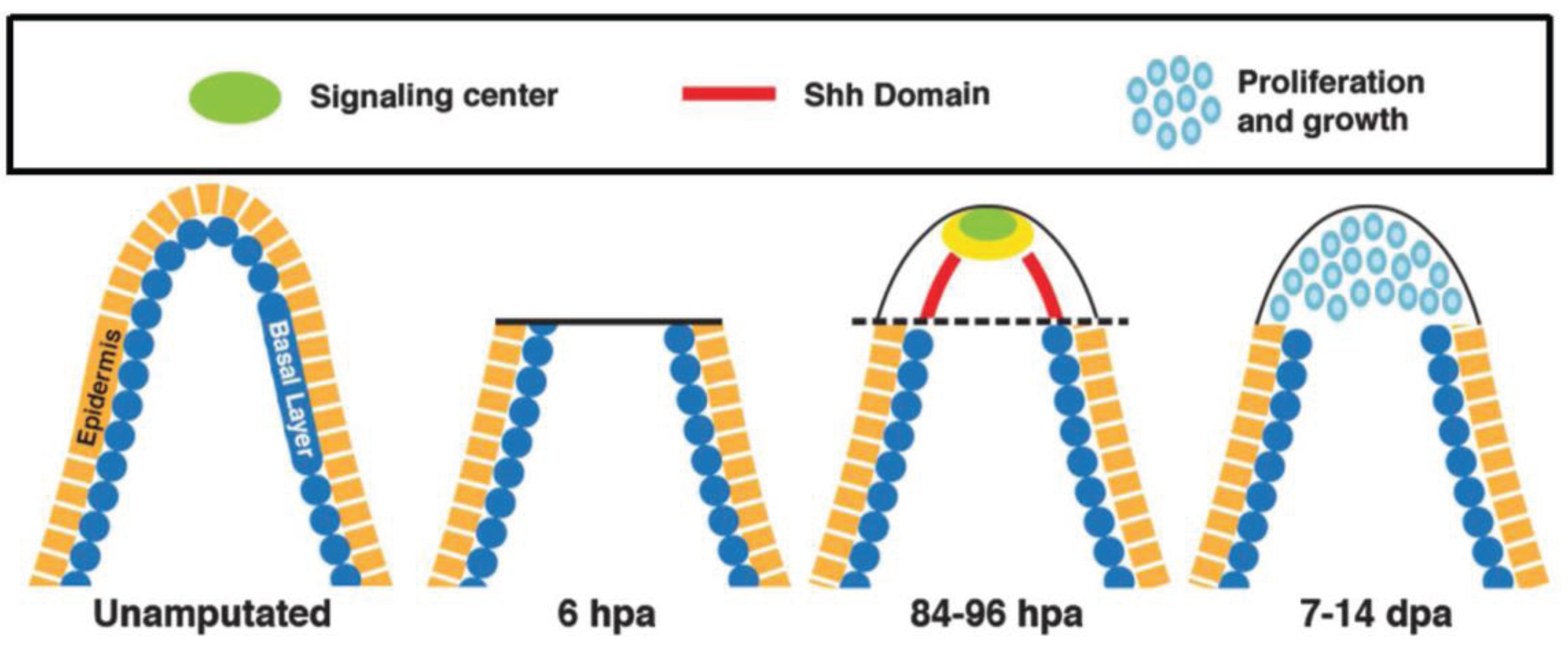
3. HH Signaling in Fin Regeneration

4. HH Signaling in Limb and Tail Regeneration

5. Concluding Remarks
Acknowledgment
Conflicts of Interest
References
- Slack, J.M. Regeneration research today. Dev. Dyn. 2003, 226, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Bely, A.E. Evolutionary loss of animal regeneration: Pattern and process. Integr. Comp. Biol. 2010, 50, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Hata, S.; Namae, M.; Nishina, H. Liver development and regeneration: From laboratory study to clinical therapy. Dev. Growth Differ. 2007, 49, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Garry, D.J. Muscle stem cells in development, regeneration, and disease. Genes Dev. 2006, 20, 1692–1708. [Google Scholar] [CrossRef] [PubMed]
- Sancho-Bru, P.; Najimi, M.; Caruso, M.; Pauwelyn, K.; Cantz, T.; Forbes, S.; Roskams, T.; Ott, M.; Gehling, U.; Sokal, E.; et al. Stem and progenitor cells for liver repopulation: Can we standardise the process from bench to bedside? Gut 2009, 58, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Olson, E.N.; Bassel-Duby, R. Mending broken hearts: cardiac development as a basis for adult heart regeneration and repair. Nat. Rev. Mol. Cell Biol. 2013, 14, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Maki, N.; Martinson, J.; Nishimura, O.; Tarui, H.; Meller, J.; Tsonis, P.A.; Agata, K. Expression profiles during dedifferentiation in newt lens regeneration revealed by expressed sequence tags. Mol. Vis. 2010, 16, 72–78. [Google Scholar] [PubMed]
- Love, N.R.; Chen, Y.; Bonev, B.; Gilchrist, M.J.; Fairclough, L.; Lea, R.; Mohun, T.J.; Paredes, R.; Zeef, L.A.; Amaya, E. Genome-wide analysis of gene expression during Xenopus tropicalis tadpole tail regeneration. BMC Dev. Biol. 2011, 11, 70. [Google Scholar] [CrossRef] [PubMed]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart regeneration in zebrafish. Science 2002, 298, 2188–2190. [Google Scholar] [CrossRef] [PubMed]
- Caubit, X.; Nicolas, S.; le Parco, Y. Possible roles for Wnt genes in growth and axial patterning during regeneration of the tail in urodele amphibians. Dev. Dyn. 1997, 210, 1–10. [Google Scholar] [CrossRef]
- Brockes, J.P.; Kumar, A. Plasticity and reprogramming of differentiated cells in amphibian regeneration. Nat. Rev. Mol. Cell Biol. 2002, 3, 566–574. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Slack, J.M. Requirement for Wnt and FGF signaling in Xenopus tadpole tail regeneration. Dev. Biol. 2008, 316, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Koyano-Nakagawa, N.; Garry, J.P.; Weaver, C.V. Heart of newt: A recipe for regeneration. J. Cardiovasc. Transl. Res. 2010, 3, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Globus, M.; Vethamany-Globus, S.; Lee, Y.C. Effect of apical epidermal cap on mitotic cycle and cartilage differentiation in regeneration blastemata in the newt, Notophthalmus viridescens. Dev. Biol. 1980, 75, 358–372. [Google Scholar] [CrossRef]
- Schnapp, E.; Kragl, M.; Rubin, L.; Tanaka, E.M. Hedgehog signaling controls dorsoventral patterning, blastema cell proliferation and cartilage induction during axolotl tail regeneration. Development 2005, 132, 3243–3253. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Yokoyama, H.; Tamura, K.; Ide, H. Shh expression in developing and regenerating limb buds of Xenopus laevis. Dev. Dyn. 1997, 209, 227–232. [Google Scholar] [CrossRef]
- Kawakami, Y.; Rodriguez Esteban, C.; Raya, M.; Kawakami, H.; Marti, M.; Dubova, I.; Izpisua Belmonte, J.C. Wnt/beta-catenin signaling regulates vertebrate limb regeneration. Genes Dev. 2006, 20, 3232–3237. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Doyle, M.J.; Weaver, C.V.; Koyano-Nakagawa, N.; Garry, D.J. Hedgehog and Wnt coordinate signaling in myogenic progenitors and regulate limb regeneration. Dev. Biol. 2012, 371, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Rose, S.M. Epidermal dedifferentiation during blastema formation in regenerating limbs of Triturus viridescens. J. Exp. Zool. 1948, 108, 337–361. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.M.; Endo, T.; Bryant, S.V. The molecular basis of amphibian limb regeneration: integrating the old with the new. Semin. Cell Dev. Biol. 2002, 13, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Stoick-Cooper, C.L.; Moon, R.T.; Weidinger, G. Advances in signaling in vertebrate regeneration as a prelude to regenerative medicine. Genes Dev. 2007, 21, 1292–1315. [Google Scholar] [CrossRef] [PubMed]
- Yakushiji, N.; Suzuki, M.; Satoh, A.; Ide, H.; Tamura, K. Effects of activation of hedgehog signaling on patterning, growth, and differentiation in Xenopus froglet limb regeneration. Dev. Dyn. 2009, 238, 1887–1896. [Google Scholar] [CrossRef] [PubMed]
- Yakushiji, N.; Suzuki, M.; Satoh, A.; Sagai, T.; Shiroishi, T.; Kobayashi, H.; Sasaki, H.; Ide, H.; Tamura, K. Correlation between Shh expression and DNA methylation status of the limb-specific Shh enhancer region during limb regeneration in amphibians. Dev. Biol. 2007, 312, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Chablais, F.; Jazwinska, A. IGF signaling between blastema and wound epidermis is required for fin regeneration. Development 2010, 137, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Iovine, M.K. Conserved mechanisms regulate outgrowth in zebrafish fins. Nat. Chem. Biol. 2007, 3, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Knopf, F.; Hammond, C.; Chekuru, A.; Kurth, T.; Hans, S.; Weber, C.W.; Mahatma, G.; Fisher, S.; Brand, M.; Schulte-Merker, S.; et al. Bone regenerates via dedifferentiation of osteoblasts in the zebrafish fin. Dev. Cell 2011, 20, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Padhi, B.K.; Joly, L.; Tellis, P.; Smith, A.; Nanjappa, P.; Chevrette, M.; Ekker, M.; Akimenko, M.A. Screen for genes differentially expressed during regeneration of the zebrafish caudal fin. Dev. Dyn. 2004, 231, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Poss, K.D.; Keating, M.T.; Nechiporuk, A. Tales of regeneration in zebrafish. Dev. Dyn. 2003, 226, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Avaron, F.; Guay, D.; Padhi, B.K.; Akimenko, M.A. Inhibition of BMP signaling during zebrafish fin regeneration disrupts fin growth and scleroblasts differentiation and function. Dev. Biol. 2006, 299, 438–454. [Google Scholar] [CrossRef] [PubMed]
- Haubner, B.J.; Adamowicz-Brice, M.; Khadayate, S.; Tiefenthaler, V.; Metzler, B.; Aitman, T.; Penninger, J.M. Complete cardiac regeneration in a mouse model of myocardial infarction. Aging 2012, 4, 966–977. [Google Scholar] [PubMed]
- Kikuchi, K.; Holdway, J.E.; Major, R.J.; Blum, N.; Dahn, R.D.; Begemann, G.; Poss, K.D. Retinoic acid production by endocardium and epicardium is an injury response essential for zebrafish heart regeneration. Dev. Cell 2011, 20, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Borikova, A.L.; Ben-Yair, R.; Guner-Ataman, B.; MacRae, C.A.; Lee, R.T.; Burns, C.G.; Burns, C.E. Notch signaling regulates cardiomyocyte proliferation during zebrafish heart regeneration. Proc. Natl. Acad. Sci. USA 2014, 111, 1403–1408. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.; Montserrat, N.; Zacchigna, S.; Nivet, E.; Hishida, T.; Krause, M.N.; Kurian, L.; Ocampo, A.; Vazquez-Ferrer, E.; Rodriguez-Esteban, C.; et al. In vivo activation of a conserved microRNA program induces mammalian heart regeneration. Cell Stem Cell 2014, 15, 589–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zgheib, C.; Allukian, M.W.; Xu, J.; Morris, M.W., Jr.; Caskey, R.C.; Herdrich, B.J.; Hu, J.; Gorman, J.H., 3rd; Gorman, R.C.; Liechty, K.W. Mammalian fetal cardiac regeneration after myocardial infarction is associated with differential gene expression compared with the adult. Ann. Thorac. Surg. 2014, 97, 1643–1650. [Google Scholar] [CrossRef] [PubMed]
- Muneoka, K.; Holler-Dinsmore, G.; Bryant, S.V. Intrinsic control of regenerative loss in Xenopus laevis limbs. J. Exp. Zool. 1986, 240, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Brockes, J.P. Amphibian limb regeneration: rebuilding a complex structure. Science 1997, 276, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Brockes, J.P.; Kumar, A. Appendage regeneration in adult vertebrates and implications for regenerative medicine. Science 2005, 310, 1919–1923. [Google Scholar] [CrossRef] [PubMed]
- Hay, E.D.; Fischman, D.A. Origin of the blastema in regenerating limbs of the newt Triturus viridescens. An autoradiographic study using tritiated thymidine to follow cell proliferation and migration. Dev. Biol. 1961, 3, 26–59. [Google Scholar] [CrossRef]
- Jopling, C.; Sleep, E.; Raya, M.; Marti, M.; Raya, A.; Izpisua Belmonte, J.C. Zebrafish heart regeneration occurs by cardiomyocyte dedifferentiation and proliferation. Nature 2010, 464, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Casimir, C.M.; Gates, P.B.; Patient, R.K.; Brockes, J.P. Evidence for dedifferentiation and metaplasia in amphibian limb regeneration from inheritance of DNA methylation. Development 1988, 104, 657–668. [Google Scholar] [PubMed]
- Zeller, R.; Lopez-Rios, J.; Zuniga, A. Vertebrate limb bud development: Moving towards integrative analysis of organogenesis. Nat. Rev. Genet. 2009, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Teran, M.; Ros, M.A. The Apical Ectodermal Ridge: Morphological aspects and signaling pathways. Int. J. Dev. Biol. 2008, 52, 857–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandoski, M.; Sun, X.; Martin, G.R. Fgf8 signalling from the AER is essential for normal limb development. Nat. Genet. 2000, 26, 460–463. [Google Scholar] [PubMed]
- Mariani, F.V.; Ahn, C.P.; Martin, G.R. Genetic evidence that FGFs have an instructive role in limb proximal-distal patterning. Nature 2008, 453, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Wylie, J.N.; Galceran, J.; Arkhitko, O.; Li, C.; Deng, C.; Grosschedl, R.; Bruneau, B.G. Tbx5 is essential for forelimb bud initiation following patterning of the limb field in the mouse embryo. Development 2003, 130, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Koshiba-Takeuchi, K.; Takeuchi, J.K.; Arruda, E.P.; Kathiriya, I.S.; Mo, R.; Hui, C.C.; Srivastava, D.; Bruneau, B.G. Cooperative and antagonistic interactions between Sall4 and Tbx5 pattern the mouse limb and heart. Nat. Genet. 2006, 38, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Rallis, C.; del Buono, J.; Logan, M.P. Tbx3 can alter limb position along the rostrocaudal axis of the developing embryo. Development 2005, 132, 1961–1970. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Capdevila, J.; Buscher, D.; Itoh, T.; Rodriguez Esteban, C.; Izpisua Belmonte, J.C. WNT signals control FGF-dependent limb initiation and AER induction in the chick embryo. Cell 2001, 104, 891–900. [Google Scholar] [CrossRef]
- Cohen, M.M., Jr. The hedgehog signaling network. Am. J. Med. Genet. A. 2003, 123A, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Pan, A.; Chang, L.; Nguyen, A.; James, A.W. A review of hedgehog signaling in cranial bone development. Front. Physiol. 2013, 4, 61. [Google Scholar] [CrossRef] [PubMed]
- Robbins, D.J.; Fei, D.L.; Riobo, N.A. The Hedgehog signal transduction network. Sci. Signal. 2012, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D. Hedgehog signalling: Emerging evidence for non-canonical pathways. Cell Signal. 2009, 21, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Parmantier, E.; Lynn, B.; Lawson, D.; Turmaine, M.; Namini, S.S.; Chakrabarti, L.; McMahon, A.P.; Jessen, K.R.; Mirsky, R. Schwann cell-derived Desert hedgehog controls the development of peripheral nerve sheaths. Neuron 1999, 23, 713–724. [Google Scholar] [CrossRef]
- Chiang, C.; Litingtung, Y.; Lee, E.; Young, K.E.; Corden, J.L.; Westphal, H.; Beachy, P.A. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 1996, 383, 407–413. [Google Scholar] [CrossRef] [PubMed]
- St-Jacques, B.; Hammerschmidt, M.; McMahon, A.P. Indian hedgehog signaling regulates proliferation and differentiation of chondrocytes and is essential for bone formation. Genes Dev. 1999, 13, 2072–2086. [Google Scholar] [CrossRef] [PubMed]
- Tukachinsky, H.; Lopez, L.V.; Salic, A. A mechanism for vertebrate Hedgehog signaling: recruitment to cilia and dissociation of SuFu-Gli protein complexes. J. Cell Biol. 2010, 191, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Keady, B.T.; Samtani, R.; Tobita, K.; Tsuchya, M.; San Agustin, J.T.; Follit, J.A.; Jonassen, J.A.; Subramanian, R.; Lo, C.W.; Pazour, G.J. IFT25 links the signal-dependent movement of Hedgehog components to intraflagellar transport. Dev. Cell 2012, 22, 940–951. [Google Scholar] [CrossRef] [PubMed]
- Ashe, A.; Butterfield, N.C.; Town, L.; Courtney, A.D.; Cooper, A.N.; Ferguson, C.; Barry, R.; Olsson, F.; Liem, K.F., Jr.; Parton, R.G.; et al. Mutations in mouse Ift144 model the craniofacial, limb and rib defects in skeletal ciliopathies. Hum. Mol. Genet. 2012, 21, 1808–1823. [Google Scholar] [CrossRef] [PubMed]
- Dagoneau, N.; Goulet, M.; Genevieve, D.; Sznajer, Y.; Martinovic, J.; Smithson, S.; Huber, C.; Baujat, G.; Flori, E.; Tecco, L.; et al. DYNC2H1 mutations cause asphyxiating thoracic dystrophy and short rib-polydactyly syndrome, type III. Am. J. Hum. Genet. 2009, 84, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Ingham, P.W.; Nakano, Y.; Seger, C. Mechanisms and functions of Hedgehog signalling across the metazoa. Nat. Rev. Genet. 2011, 12, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, N.C.; Metzis, V.; McGlinn, E.; Bruce, S.J.; Wainwright, B.J.; Wicking, C. Patched 1 is a crucial determinant of asymmetry and digit number in the vertebrate limb. Development 2009, 136, 3515–3524. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, L.V.; Milenkovic, L.; Higgins, K.M.; Scott, M.P. Altered neural cell fates and medulloblastoma in mouse patched mutants. Science 1997, 277, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Vokes, S.A.; Ji, H.; Wong, W.H.; McMahon, A.P. A genome-scale analysis of the cis-regulatory circuitry underlying sonic hedgehog-mediated patterning of the mammalian limb. Genes Dev. 2008, 22, 2651–2663. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.W.; Chuang, P.T. Mechanism and evolution of cytosolic Hedgehog signal transduction. Development 2010, 137, 2079–2094. [Google Scholar] [CrossRef] [PubMed]
- Laufer, E.; Nelson, C.E.; Johnson, R.L.; Morgan, B.A.; Tabin, C. Sonic hedgehog and Fgf-4 act through a signaling cascade and feedback loop to integrate growth and patterning of the developing limb bud. Cell 1994, 79, 993–1003. [Google Scholar] [CrossRef]
- Park, H.L.; Bai, C.; Platt, K.A.; Matise, M.P.; Beeghly, A.; Hui, C.C.; Nakashima, M.; Joyner, A.L. Mouse Gli1 mutants are viable but have defects in SHH signaling in combination with a Gli2 mutation. Development 2000, 127, 1593–1605. [Google Scholar] [PubMed]
- Zhang, X.M.; Ramalho-Santos, M.; McMahon, A.P. Smoothened mutants reveal redundant roles for Shh and Ihh signaling including regulation of L/R asymmetry by the mouse node. Cell 2001, 105, 781–792. [Google Scholar] [CrossRef]
- Chiang, C.; Litingtung, Y.; Harris, M.P.; Simandl, B.K.; Li, Y.; Beachy, P.A.; Fallon, J.F. Manifestation of the limb prepattern: limb development in the absence of sonic hedgehog function. Dev. Biol. 2001, 236, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Bruce, S.J.; Butterfield, N.C.; Metzis, V.; Town, L.; McGlinn, E.; Wicking, C. Inactivation of Patched1 in the mouse limb has novel inhibitory effects on the chondrogenic program. J. Biol. Chem. 2010, 285, 27967–27981. [Google Scholar] [CrossRef] [PubMed]
- Nasevicius, A.; Ekker, S.C. Effective targeted gene 'knockdown' in zebrafish. Nat. Genet. 2000, 26, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Bouldin, C.M.; Gritli-Linde, A.; Ahn, S.; Harfe, B.D. Shh pathway activation is present and required within the vertebrate limb bud apical ectodermal ridge for normal autopod patterning. Proc. Natl. Acad. Sci. USA 2010, 107, 5489–5494. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Litingtung, Y.; Chiang, C. Cholesterol modification restricts the spread of Shh gradient in the limb bud. Proc. Natl. Acad. Sci. USA 2006, 103, 6548–6553. [Google Scholar] [CrossRef] [PubMed]
- Harfe, B.D.; Scherz, P.J.; Nissim, S.; Tian, H.; McMahon, A.P.; Tabin, C.J. Evidence for an expansion-based temporal Shh gradient in specifying vertebrate digit identities. Cell 2004, 118, 517–528. [Google Scholar] [CrossRef] [PubMed]
- te Welscher, P.; Zuniga, A.; Kuijper, S.; Drenth, T.; Goedemans, H.J.; Meijlink, F.; Zeller, R. Progression of vertebrate limb development through SHH-mediated counteraction of GLI3. Science 2002, 298, 827–830. [Google Scholar] [CrossRef] [PubMed]
- Kraus, P.; Fraidenraich, D.; Loomis, C.A. Some distal limb structures develop in mice lacking Sonic hedgehog signaling. Mech. Dev. 2001, 100, 45–58. [Google Scholar] [CrossRef]
- Montavon, T.; Le Garrec, J.F.; Kerszberg, M.; Duboule, D. Modeling Hox gene regulation in digits: reverse collinearity and the molecular origin of thumbness. Genes Dev. 2008, 22, 346–359. [Google Scholar] [CrossRef] [PubMed]
- Niswander, L.; Jeffrey, S.; Martin, G.R.; Tickle, C. A positive feedback loop coordinates growth and patterning in the vertebrate limb. Nature 1994, 371, 609–612. [Google Scholar] [CrossRef] [PubMed]
- Verheyden, J.M.; Sun, X. An Fgf/Gremlin inhibitory feedback loop triggers termination of limb bud outgrowth. Nature 2008, 454, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Scherz, P.J.; Harfe, B.D.; McMahon, A.P.; Tabin, C.J. The limb bud Shh-Fgf feedback loop is terminated by expansion of former ZPA cells. Science 2004, 305, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Selever, J.; Liu, W.; Lu, M.F.; Behringer, R.R.; Martin, J.F. Bmp4 in limb bud mesoderm regulates digit pattern by controlling AER development. Dev. Biol. 2004, 276, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Dahn, R.D.; Fallon, J.F. Interdigital regulation of digit identity and homeotic transformation by modulated BMP signaling. Science 2000, 289, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Akimenko, M.A.; Mari-Beffa, M.; Becerra, J.; Geraudie, J. Old questions, new tools, and some answers to the mystery of fin regeneration. Dev. Dyn. 2003, 226, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, Y.; Yoshizato, K. Expression of Sonic hedgehog gene in regenerating newt limb blastemas recapitulates that in developing limb buds. Proc. Natl. Acad. Sci. USA 1997, 94, 9159–9164. [Google Scholar] [CrossRef] [PubMed]
- Scadding, S.R.; Maden, M. Retinoic acid gradients during limb regeneration. Dev. Biol. 1994, 162, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Quint, E.; Smith, A.; Avaron, F.; Laforest, L.; Miles, J.; Gaffield, W.; Akimenko, M.A. Bone patterning is altered in the regenerating zebrafish caudal fin after ectopic expression of sonic hedgehog and bmp2b or exposure to cyclopamine. Proc. Natl. Acad. Sci. USA 2002, 99, 8713–8718. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Grill, S.; Sanchez, A.; Murphy-Ryan, M.; Poss, K.D. Fgf signaling instructs position-dependent growth rate during zebrafish fin regeneration. Development 2005, 132, 5173–5183. [Google Scholar] [CrossRef] [PubMed]
- Avaron, F.; Hoffman, L.; Guay, D.; Akimenko, M.A. Characterization of two new zebrafish members of the hedgehog family: atypical expression of a zebrafish indian hedgehog gene in skeletal elements of both endochondral and dermal origins. Dev. Dyn. 2006, 235, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jeradi, S.; Strahle, U.; Akimenko, M.A. Laser ablation of the sonic hedgehog-a-expressing cells during fin regeneration affects ray branching morphogenesis. Dev. Biol. 2012, 365, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Hami, D.; de Val, S.; Kagermeier-Schenk, B.; Wills, A.A.; Black, B.L.; Weidinger, G.; Poss, K.D. Maintenance of blastemal proliferation by functionally diverse epidermis in regenerating zebrafish fins. Dev. Biol. 2009, 331, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.J.; Crews, C.M. Wound epidermis formation and function in urodele amphibian limb regeneration. Cell Mol. Life Sci. 2008, 65, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Lentz, T.L. Cytological studies of muscle dedifferentiation and differentiation during limb regeneration of the newt Triturus. Am. J. Anat. 1969, 124, 447–479. [Google Scholar] [CrossRef] [PubMed]
- Echeverri, K.; Clarke, J.D.; Tanaka, E.M. In vivo imaging indicates muscle fiber dedifferentiation is a major contributor to the regenerating tail blastema. Dev. Biol. 2001, 236, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.I.; Loof, S.; He, P.; Simon, A. Salamander limb regeneration involves the activation of a multipotent skeletal muscle satellite cell population. J. Cell. Biol. 2006, 172, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Guzman, T.; Wang, H.; Khattak, S.; Schuez, M.; Roensch, K.; Nacu, E.; Tazaki, A.; Joven, A.; Tanaka, E.M.; Simon, A. Fundamental differences in dedifferentiation and stem cell recruitment during skeletal muscle regeneration in two salamander species. Cell Stem Cell 2014, 14, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Kragl, M.; Knapp, D.; Nacu, E.; Khattak, S.; Maden, M.; Epperlein, H.H.; Tanaka, E.M. Cells keep a memory of their tissue origin during axolotl limb regeneration. Nature 2009, 460, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, L.V.; Johnson, R.L.; Milenkovic, L.; McMahon, J.A.; Scott, M.P. Conservation of the hedgehog/patched signaling pathway from flies to mice: induction of a mouse patched gene by Hedgehog. Genes Dev. 1996, 10, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Ochoa, B.; Syn, W.K.; Delgado, I.; Karaca, G.F.; Jung, Y.; Wang, J.; Zubiaga, A.M.; Fresnedo, O.; Omenetti, A.; Zdanowicz, M.; et al. Hedgehog signaling is critical for normal liver regeneration after partial hepatectomy in mice. Hepatology 2010, 51, 1712–1723. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, B.N.; Koyano-Nakagawa, N.; Donaldson, A.; Weaver, C.V.; Garry, M.G.; Garry, D.J. Hedgehog Signaling during Appendage Development and Regeneration. Genes 2015, 6, 417-435. https://doi.org/10.3390/genes6020417
Singh BN, Koyano-Nakagawa N, Donaldson A, Weaver CV, Garry MG, Garry DJ. Hedgehog Signaling during Appendage Development and Regeneration. Genes. 2015; 6(2):417-435. https://doi.org/10.3390/genes6020417
Chicago/Turabian StyleSingh, Bhairab N., Naoko Koyano-Nakagawa, Andrew Donaldson, Cyprian V. Weaver, Mary G. Garry, and Daniel J. Garry. 2015. "Hedgehog Signaling during Appendage Development and Regeneration" Genes 6, no. 2: 417-435. https://doi.org/10.3390/genes6020417