
Characterization and Comparison of the Two Mitochondrial Genomes in the Genus Rana

1
Laboratory of Amphibian Diversity Investigation, College of Ecology, Lishui University, Lishui 323000, China
2
College of Life and Environmental Sciences, Hangzhou Normal University, Hangzhou 311121, China
3
School of Life Sciences, Xiamen University, Xiamen 361005, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Genes 2023, 14(9), 1786; https://doi.org/10.3390/genes14091786
Submission received: 1 August 2023
/
Revised: 29 August 2023
/
Accepted: 7 September 2023
/
Published: 11 September 2023
(This article belongs to the Special Issue Advanced Research on Mitochondrial Genome)
Abstract
:The mitochondrial genome (mitogenome) possesses several invaluable attributes, including limited recombination, maternal inheritance, a fast evolutionary rate, compact size, and relatively conserved gene arrangement, all of which make it particularly useful for applications in phylogenetic reconstruction, population genetics, and evolutionary research. In this study, we aimed to determine the complete mitogenomes of two morphologically similar Rana species (Rana hanluica and Rana longicrus) using next-generation sequencing. The entire circular mitogenome was successfully identified, with a length of 19,395 bp for R. hanluica and 17,833 bp for R. longicrus. The mitogenomes of both species contained 37 genes, including 13 protein-coding genes (PCGs), two ribosomal RNA genes, 22 transfer RNA genes, and one control region; mitogenome size varied predominantly with the length of the control region. The two synonymous codon usages in 13 PCGs showed that T and A were used more frequently than G and C. The ratios of non-synonymous to synonymous substitutions of all 13 PCGs were <1 in the Rana species, indicating that the PCGs were under purifying selection. Finally, phylogenetic relationship analyses suggested that R. hanluica and R. longicrus were classified in the R. japonica group. Our study provides valuable reference material for the taxonomy of the genus Rana.
1. Introduction
The mitochondrial genome (mitogenome) possesses several valuable characteristics (e.g., limited recombination, maternal inheritance, rapid evolutionary rate, small size, and relatively conserved gene arrangement) for utilization in phylogenetic reconstruction, population genetics, and evolutionary studies [1,2]. Understanding the factors driving mitogenomic evolution may offer crucial insights into these benefits [2]. As anticipated, the mitogenomes of Anura are double-stranded circular molecules with a length of 16–24 kbp [3], encompassing two ribosomal RNA (12S and 16S rRNAs) genes, 22 transfer RNA (tRNA) genes, 13 protein-coding genes (PCGs), and one control region (CR; also known as the D-loop) [1,4,5,6]. The mitogenome is extensively employed in evolutionary studies of diverse species [7], and the evolution of tRNA genes and novel gene orders in the mitogenome can aid in determining phylogenetic relationships [5].
True frogs of the genus Rana Linnaeus, 1758 (Ranidae) have a wide distribution across Eurasia and the Americas [8]. Owing to their body coloration and habitat preferences, species within this genus are colloquially called brown frogs or wood frogs [9,10]. The taxonomy of Rana (brown frogs) has been the subject of intense debate over the past 20 years [8,11].
A recent phylogenetic analysis indicated that Rana sensu lato may consist of nine clades, namely the genus Rana [12]. At present, the well-established genus Rana comprises 54 species globally [13], with 26 species documented in over 30 provinces in China [8]. To date, complete mitogenome sequences of only 14 Rana species were deposited in GenBank.
In the genus Rana, one of the major species groups of Chinese brown frogs is Rana longicrus, and the majority of southern Chinese brown frogs fall within this species group [14]. Previous studies have proposed that the Long-legged Brown Frog (R. longicrus) should also be classified under this species group, owing to morphological and nucleotide differences [13,15]. However, prior research indicated that R. longicrus belongs to the R. japonica group [15,16]. Alongside taxonomic revisions, recent investigations on this subgenus have unveiled several new species in China, underscoring the incomplete understanding of the subgenus Rana [9,11,17].
The Hanlui Brown Frog (Rana hanluica), named after its breeding season, which is around the “Cold Dew Festival”, falling in one of the 24 solar terms of the Chinese lunar calendar, is endemic to China. Its type locality is in Shuangpai, Hunan Province [18]. While most Rana species distributed in southern China are part of the R. japonica group [17,19], Fei et al. [13] classified six Rana species from southern China, including R. hanluica, under the R. longicrus group. The uncertain systematic studies of the genus Rana have hindered a comprehensive understanding of species diversity and biogeographic patterns within the genus [12]. Therefore, further investigations based on complete mitogenome data might enhance our comprehension of the phylogenetic relationships among these Ranidae species [20].
In this study, we successfully determined the complete mitogenome sequences of the two Rana species via Illumina sequencing and compared them with those of related species based on mitochondrial structure. We also analyzed the phylogenetic position and relationships of R. longicrus and R. hanluica by reference to the available complete mitogenomes of the family Ranidae. Our findings offer valuable insights for future research on the evolutionary genetics, species delineation, and phylogeny of the genus Rana.
2. Materials and Methods
2.1. Sample Collection
The specimen of R. longicrus (voucher number: LSU20210423MXZY001) was collected from the Fujian Junzifeng National Nature Reserve, Mingxi, Fujian Province, China (E 117.4977°, N 26.3357°), in April 2021. The specimen of R. hanluica (voucher number: LSU20201009LSLD001) was collected from the Liandu Fengyuan Nature Reserve, Lishui, Zhejiang Province, China (E 119.8266°, N 28.1934°). Both specimens were deposited in 75% ethanol and stored in the Museum of the Laboratory of Amphibian Diversity Investigation at Lishui University (Lishui, Zhejiang, China). Total genomic DNAs of the two specimens were extracted from the muscle using the DNA easy Tissue Kit (TransGen Biotech Co., Beijing, China). The quantity and quality of extracted DNA samples were determined using a NanoDrop2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and agarose gel electrophoresis, respectively.
2.2. Assembly and Annotation
Sequencing libraries of the two Rana species were generated using an Illumina Truseq™ DNA Sample Preparation Kit (Illumina, San Diego, CA, USA). The prepared libraries were loaded onto an Illumina HiSeq 6000 platform for paired-end 150 bp sequencing (Novogene Bioinformatics Technology Co., Ltd., Tianjin, China). Complete mitogenomes were assembled de novo based on clean data using the GetOrganelle toolkit [21] and annotated using MitoZ v2.4 [22]. Initial sequencing data were aligned to the mitogenome using Burrows–Wheeler alignment, resulting in two SAM alignment files. The SAM files were subsequently transformed into the BAM format and organized using Samtools to optimize the efficiency of the subsequent image processing steps. Coverage statistics were determined for each position along the mitogenome by analyzing the sorted BAM files of the same depth. The outcome was a text file containing coverage details. To further analyze this information, the coverage files were imported into R and processed using the tidyverse package. Within R, the data were organized according to position, allowing the calculation of the average coverage per position. Notably, the entire circular mitogenomes of the two target species were fully covered by the reference sequence. The reference sequence comprehensively covered the entire circular mitogenome of the two target species, and this thorough coverage resulted in an average read coverage depth of 181.66× in R. hanluica and 160.51× in R. longicrus (Figure S1). This indicated that the sequencing data possessed high quality and reliability, enabling the provision of more accurate genomic information.
2.3. Bioinformatic Analysis
Graphical maps of the R. longicrus and R. hanluica mitogenomes were produced using the CGView Server (http://cgview.ca/, accessed on 12 December 2021). The base composition and relative synonymous codon usage (RSCU) were obtained using MEGA X [23]. Besides R. hanluica and R. longicrus, 22 mitogenomes of other 18 species in Ranidae were also collected, including two Amolops species, two Odorrana species, and 14 Rana species (Table S1). The skew in nucleotide composition was calculated by the GC and AT skew, which was measured according to the following formulas [24]: AT skew = (A − T)/(A + T); and GC skew = (G − C)/(G + C). To evaluate the divergence of the paralogous genes, we analyzed synonymous (Ks) and non-synonymous (Ka) substitutions by using DnaSP 5.1 [25]. To evaluate possible evolutionary patterns across the 13 PCGs, the Ka/Ks ratios (namely the ω value) were calculated among the 16 representative Rana species. If a selection does not affect fitness, Ka = Ks and ω = 1. If non-synonymous mutations are deleterious, purifying selection will reduce their fixation rate, such that Ka < Ks and ω < 1. If non-synonymous mutations are favored by Darwinian selection, it results in Ka > Ks and ω > 1 [26].
2.4. Phylogenetic Analyses
Eighteen previously published Rana mitogenomes [27,28,29,30,31,32,33,34,35,36] along with the newly determined sequences of the two species, were used in the phylogenetic analyses to discuss the relationships within the family Ranidae. Four other species (Amolops ricketti, Amolops wuyiensis, Odorrana graminea, and Odorrana schmackeri) were used as outgroups [37,38,39]. The genomes were aligned using MAFFT v7.388 [40]. The concatenated set of nucleotide sequences, including 13 PCGs and 2 rRNA genes, was used for phylogenetic analyses, which were performed following the Bayesian inference (BI) and Maximum Likelihood (ML) methods in MrBayes v3.2.7 and IQ-TREE2 v2.1.2, respectively. In the BI analysis, the best-fit substitution model (GTR + G + I) was selected, and the following settings were applied: two simultaneous runs of 10,000,000 generations were conducted for the matrix; the number of Markov chain Monte Carlo generations was set to ten million; the sampling frequency was set to 1000, and the burn-in was set to 1000. The Bayesian runs achieved sufficient convergence when the average standard deviation of the split frequencies was <0.01. ML analyses were performed using the best-fitting nucleotide substitution model (TPM2 + F + I + G4), which was selected by ModelFinder [41,42]. Support for the inferred ML tree was inferred by bootstrapping with 1000 replicates.
3. Results and Discussion
3.1. General Features of Mitogenomes of Two Rana Species
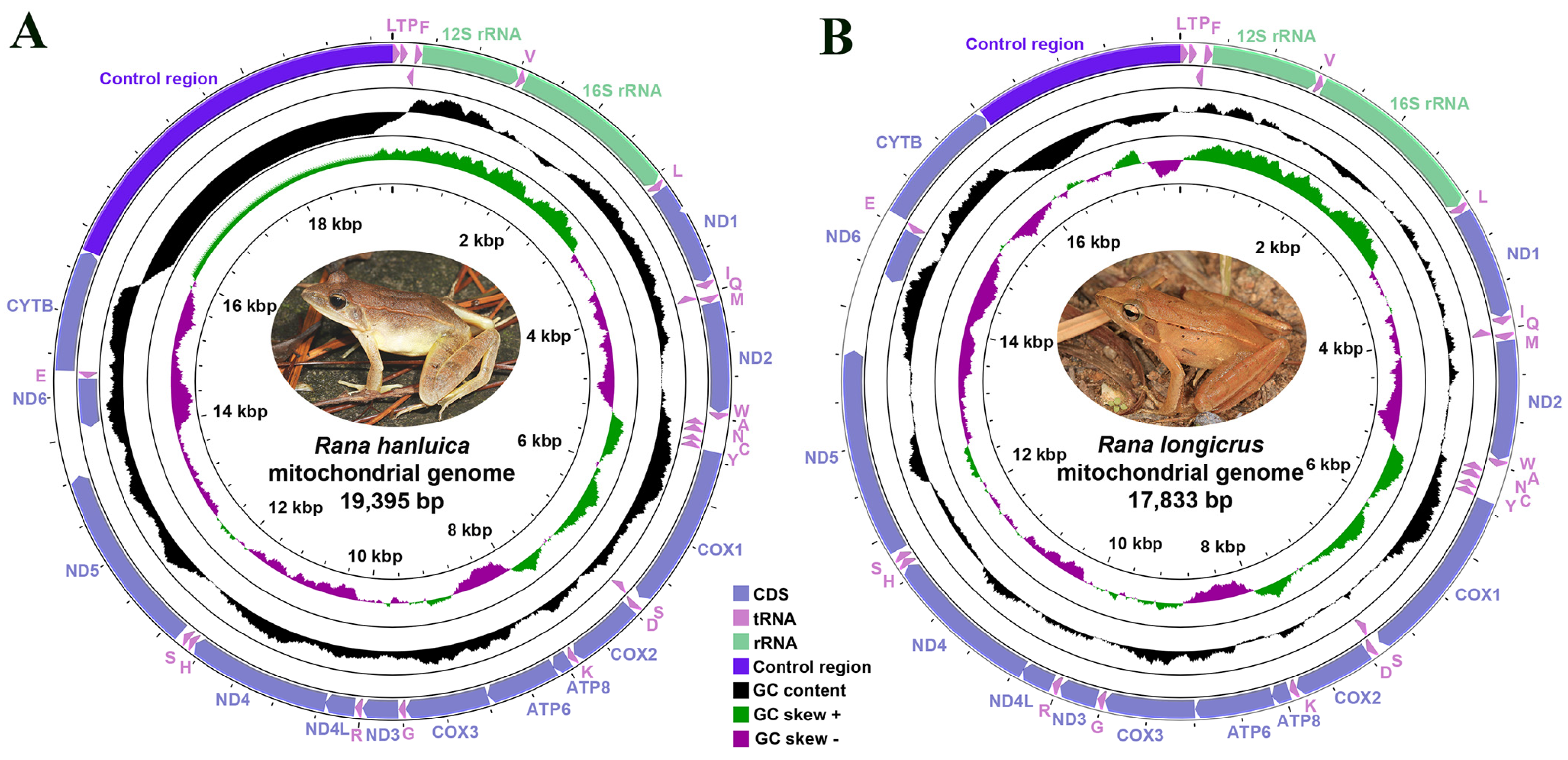
As in most ranid frogs, the two Rana species had circular mitogenomes (Figure 1). The size of the mitogenome was larger in R. hanluica (19,395 bp) than in R. longicrus (17,833 bp) (Table 1), and both comprised 13 PCGs, two rRNA genes, 22 tRNA genes, and a CR (Table S2), the length of which was inconsistent between the two species (Figure 1). Differences in mitogenome length between the species were predominantly attributed to the overall length of the CR, which in turn differed in both replicates and lengths of various short-repeat sequences within it [43]. The mitochondrial genomes of R. longicrus and R. hanluica were compact with only a few intergenic spaces and overlapping gene junctions. Furthermore, among the 13 mitochondrial PCGs, only one gene (ND6) was encoded by the L-strand, which is the typical arrangement of 13 PCGs [33]. The remaining 12 genes were encoded by H-strands (Table S2). Other species in the genera Rana and Odorrana [44,45] also have similar distribution characteristics [33,46], as the arrangement of genes within vertebrate mitogenomes is highly conserved [6,47,48,49].
The mitogenomes of R. longicrus and R. hanluica were biased toward AT nucleotides, ranging from 55.6% (R. longicrus) to 59.1% (R. hanluica) (Table 1). This A-T-rich pattern reflects typical sequence features of the vertebrate mitochondrial genome [50]. Furthermore, in the two Rana mitogenomes, AT-skews and GC-skews were both negative, indicating that the mitogenomes of both species contained a higher percentage of AT vs. GC nucleotides, which is consistent with the lowest frequency of G content in typical amphibian mitogenomes [3,48,51]. Generally, amphibian mitogenomes show a strong bias in nucleotide composition, with third codon positions being the most strongly affected by nucleotide compositional bias and skew [48].
3.2. Protein-Coding Genes and Codon Usage
A summary of the genes constituting the R. longicrus and R. hanluica mitogenomes is given in Table S2. The total length of all the PCGs in R. longicrus (11,288 bp) exceeded that in R. hanluica (11,279 bp). The longest PCG was ND5, and the shortest was ATP8 in the two Rana mitogenomes. Although gene rearrangements are frequently observed in amphibians [6], they were not detected in the two Rana species. Ten of the thirteen PCGs used the standard ATG as the start codon in the two Rana species, except for ND1, COX1, and ND4L, which were initiated by the GTG codon. Although TTG is an uncommon start codon among PCGs, it is often found in amphibians [51]. However, similar results were not found in the two Rana species. Four typical stop codons were found, TAG, AGG, AGA, and TAA, and incomplete terminal codons (TA-, T-) were also present in the two Rana mitogenomes.
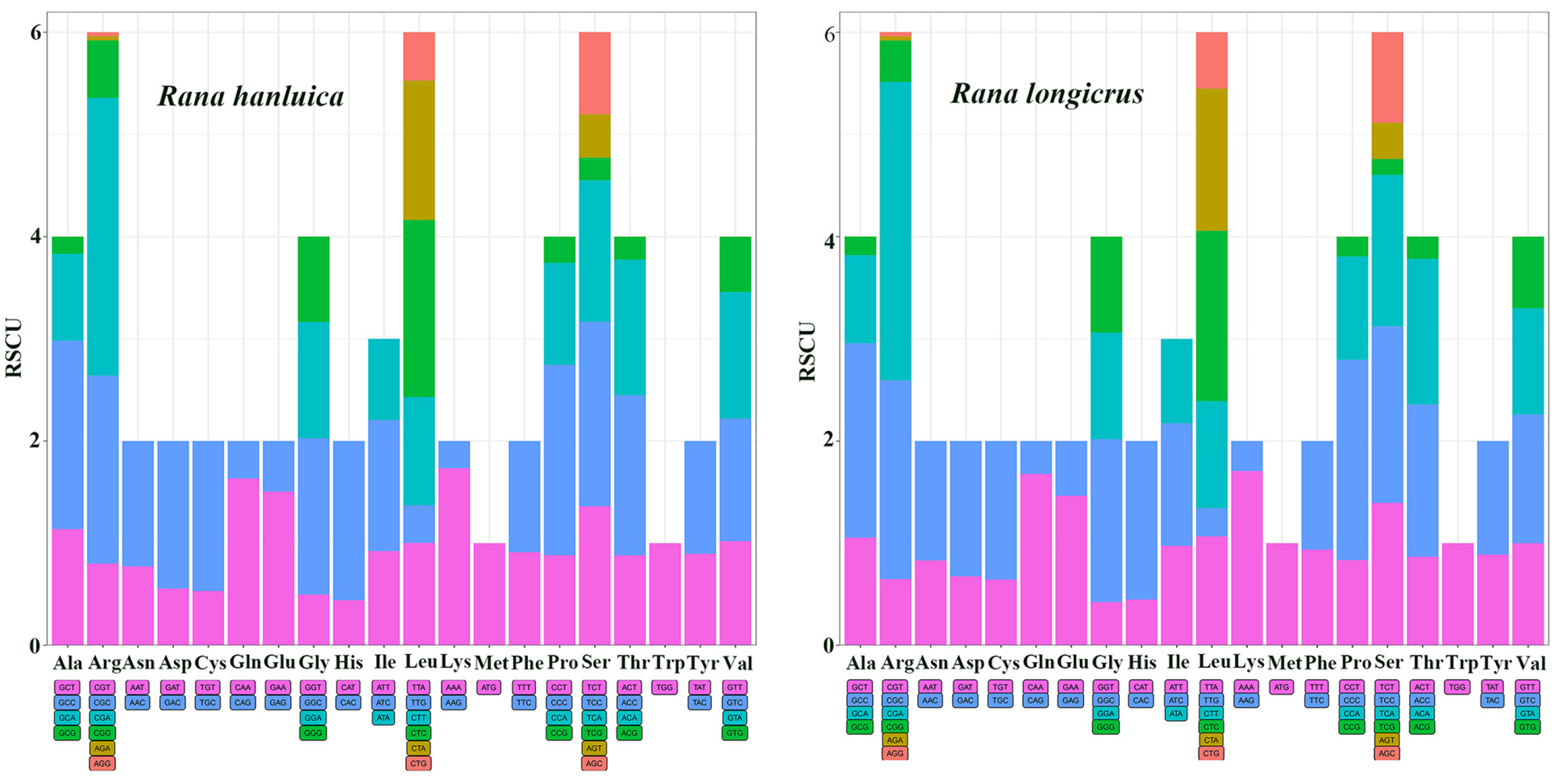
Subsequently, we evaluated the RSCU in the 13 PCGs to determine their preference for specific synonymous codons, as summarized in Table S3. Our results showed that T and A were more frequent than G and C, indicating a strong A or T bias in the third codon position. The most common amino acids in these two Rana mitogenomes were Arg, Leu, and Ser (Figure 2).
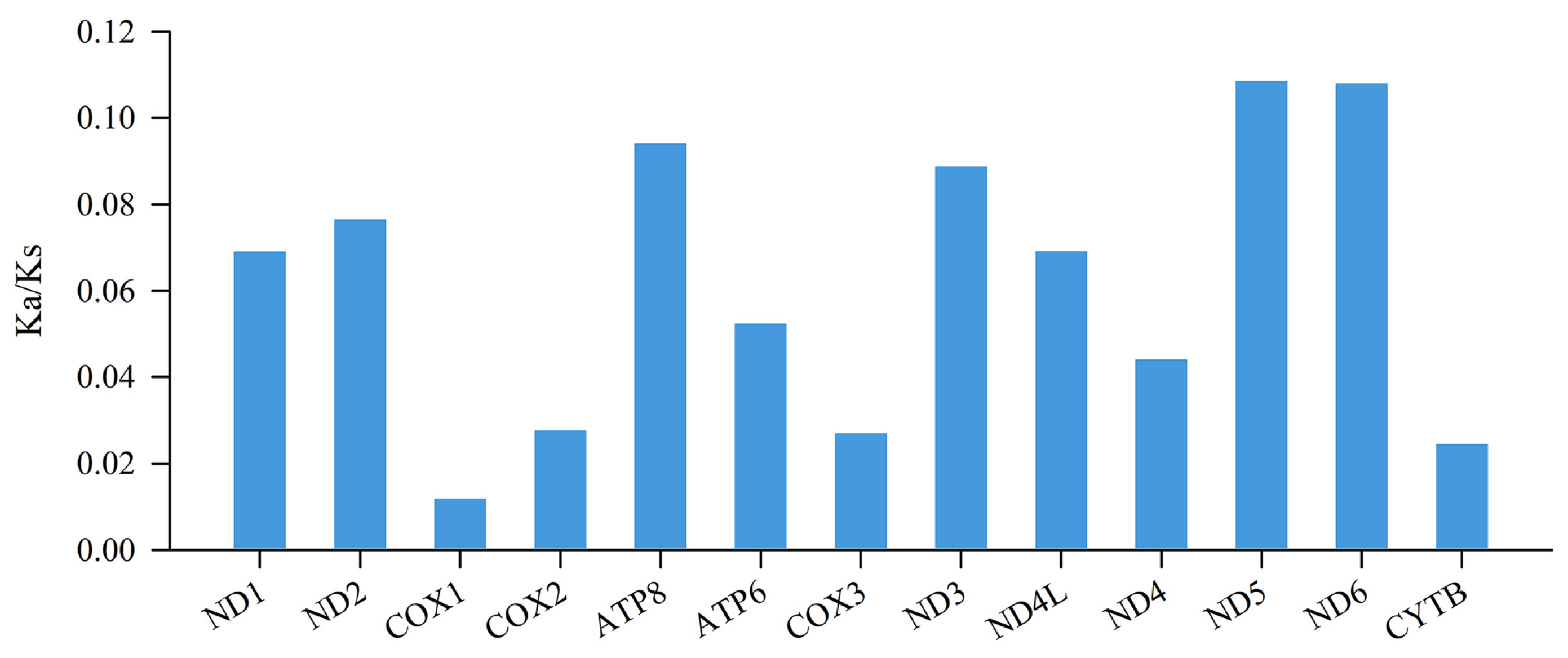
Synonymous and non-synonymous substitutions are important in understanding evolution [52], with selection pressure analysis being an indispensable part of evolutionary analysis. The Ka/Ks ratios of all 13 PCGs were much lower than those in the Rana species (Figure 3), indicating that the PCGs were under purifying selection [26]. COX1 had the lowest Ka/Ks ratio (0.012), suggesting that this gene was under the strongest selective pressure. In contrast, ND5 exhibited the highest Ka/Ks ratio (0.108), indicating that it was under the least selective pressure. The ratio varied among the 13 PCGs reflecting the different functional constraints among these genes [53]. In addition, we failed to detect significant adaptive mutations in the 13 mitogenomic PCGs; therefore, purifying selection may be the predominant force governing mitogenome evolution in Rana species [54].
3.3. Transfer RNA and Ribosomal RNA Genes
Twenty-two tRNA genes were scattered across the two Rana mitogenomes: 65 bp for tRNA-Cys and 73 bp for tRNA-Asn. All tRNA genes appeared highly A + T biased; the AT-skew was slightly positive, and the GC-skew was negative in the 22 tRNA genes of the two Rana species (Table 1). Among these tRNA genes, 8 (tRNA-Pro, tRNA-Gln, tRNA-Glu, tRNA-Ala, tRNA-Asn, tRNA-Tyr, tRNA-Cys, and tRNA-Ser) were encoded by L-strands, and the other 14 tRNA genes were encoded by H-strands (Table S2). Furthermore, we found that the lengths of all rRNA genes (2509 bp in R. hanluica and 2506 bp in R. longicrus) were similar to those of other Rana species (e.g., 2510 bp in R. omeimontis, 2505 bp in R. kukunoris, and 2511 bp in R. pyrenaica) [30,33,46]. The 12S and 16S rRNA genes are located between tRNA-Phe and tRNA-Leu and are separated by tRNA-Val, which is a common feature in vertebrate mitogenomes [54]. The A + T content of the two rRNA genes was 55.4% in R. hanluica, which was lower than that in R. longicrus (55.9%). Additionally, they had a positive AT skew and a negative GC skew.
3.4. Noncoding Regions
The noncoding regions in the mitogenome of both Rana species included the CR and a few intergenic spacers. The CR is located between tRNA-Leu and CYTB, which harbors several conserved blocks for the replication and transcription of the mitogenome. The CR of R. hanluica was 3602 bp, which was longer than that of R. longicrus (1807 bp) (Table 1). CR contains signals for the initiation and regulation of mitogenomic transcription and replication [55]. Metazoan mitogenomes usually have CRs that vary in length among species [56]. The A + T content (78.8%) of the CR in R. hanluica is the highest known to date among Rana species. In addition, both R. hanluica and R. longicrus showed negative AT-skew and GC-skew in the CR.
3.5. Phylogenetic Analysis
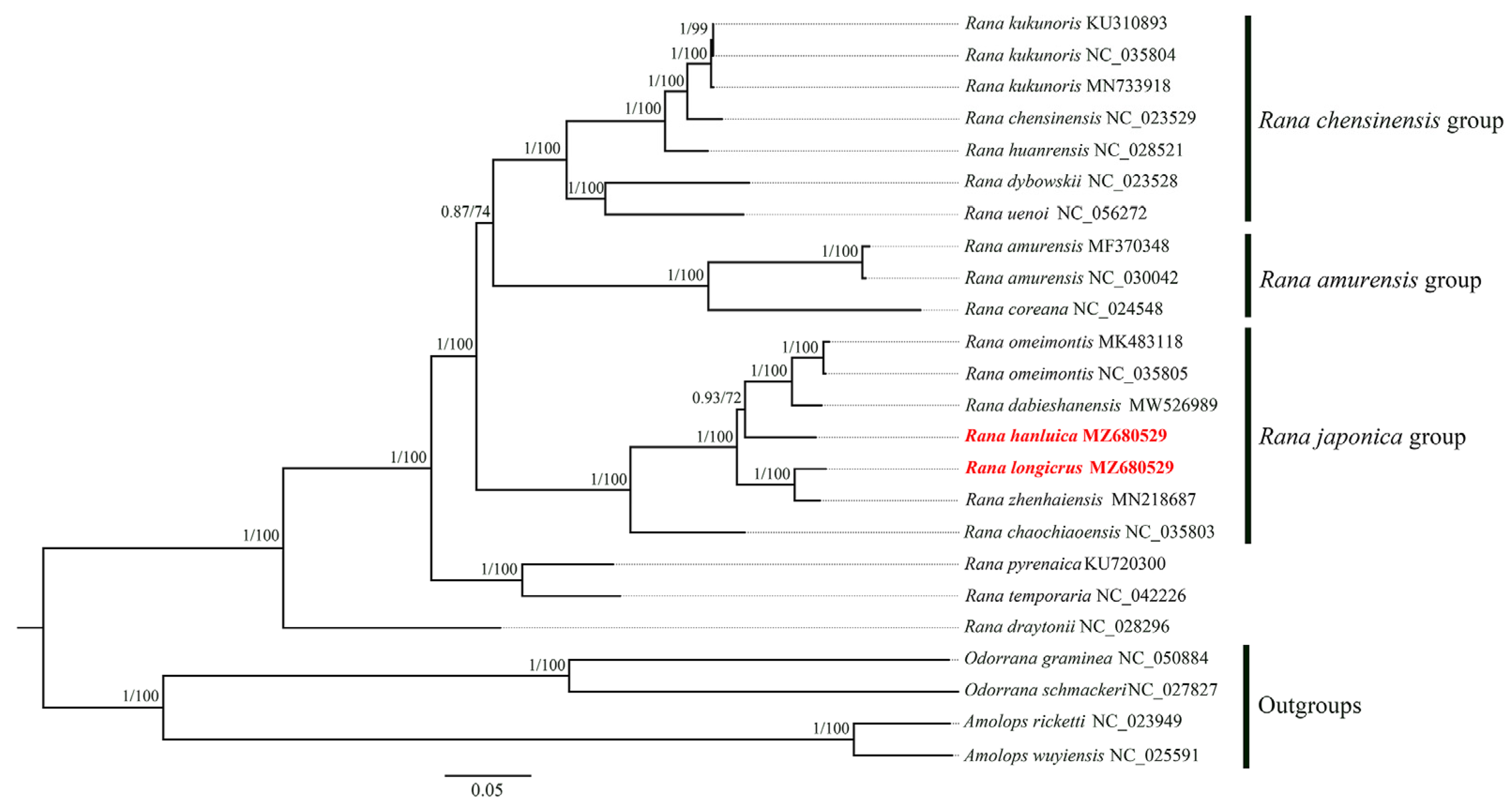
The final concatenated 13 PCGs and two rRNA genes for the 22 Rana samples were 13,921 bp in size, including 6695 variable sites of which 942 were singleton sites. The two phylogenetic reconstruction methods (ML and BI) yielded identical tree topologies that favored the clades and/or relationships of the family Ranidae (Figure 4).
The phylogenetic analysis in this study showed that the trees generated via both ML and BI were identical to each other in topological structure (Bayesian posterior probability ≥ 0.87; bootstrap value ≥ 72%), and they both supported the following classification and phylogenetic relationships of the Rana species. Three clades corresponded to previously recognized species groups, namely the R. chensinensis group, R. amurensis group, and R. japonica group. The R. japonica group was the sister group of a clade comprising the R. chensinensis and R. amurensis groups. Our results showed that R. longicrus and R. zhenhaiensis clustered on one branch, R. hanluica was close to R. dabieshanensis and R. omeimontis, and these five Rana species were classified in the R. japonica group (Figure 4). Other results suggested that R. pyrenaica and R. temporaria were clustered into a clade, and these, along with R. draytonii, were the two main conspicuous subclades (Figure 4), which may not be consistent with the other study [30].
Overall, the reconstructed phylogeny at the genus level in our study verified that R. longicrus and R. hanluica belong to the R. japonica group, which is consistent with the results of Wang et al. [57] and Wan et al. [17]. Further studies are required to clarify the phylogenetic position of the genus Rana. Although we did not fully resolve the taxonomic status within the genus Rana, this study provides valuable reference material for elucidating the taxonomy of the genus Rana.
4. Conclusions
We present the complete mitogenomes of R. hanluica and R. longicrus, with sizes of 19,395 and 17,833 bp, respectively. The mitogenome structures of the two Rana species share many features with most previously reported ranid frogs, including 37 genes in the mitogenomes (13 PCGs, 22 tRNA genes, and two rRNA genes). The phylogenetic tree was analyzed by rebuilding topological trees (ML and BI) with a dataset of 13 PCGs and two rRNA genes, which showed that R. longicrus and R. hanluica were classified in the R. japonica group. Our results provide valuable insights for further studies on the evolutionary genetics, species delimitation, and phylogeny of the genus Rana.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/genes14091786/s1; Table S1: GenBank accession numbers of the ranid species used in the phylogenetic trees; Table S2: The mitogenome characteristics and location of the two Rana species; Table S3: RSCU information for the mitochondrial protein-coding genes of R. hanluica and R. longicrus; and Figure S1: Mitogenome coverage map of (A) Rana hanluica and (B) Rana longicrus.
Author Contributions
Conceptualization, G.-H.D. and F.Q.; Data curation, Y.-M.W. and C.-Y.Z.; Formal analysis, Y.-M.W., C.-Y.Z., S.-T.L. and G.-H.D.; Methodology, Y.-M.W., C.-Y.Z. and G.-H.D.; Resources, G.-H.D.; Software, S.-T.L.; Supervision, G.-H.D. and F.Q.; Visualization, Y.-M.W., S.-T.L. and G.-H.D.; Writing—Original draft, Y.-M.W., C.-Y.Z. and G.-H.D.; Writing—Review and editing, Y.-M.W., C.-Y.Z., G.-H.D. and F.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This study received no specific grant from any funding agency.
Institutional Review Board Statement
The experimental procedures followed in this study complied with the animal welfare and research laws in China and were approved by the Ethics Committee of Lishui University (permit no. AREC-LSU202104002DGH).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: NCBI (accession: MZ680528, MZ680529, SRR15045243, and SRR15045242).
Acknowledgments
We would like to thank Lei Feng, Zhi-Qiang Chen and Yu-Fan Wang for their field assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cai, Y.; Shen, S.; Lu, L.; Storey, K.B.; Yu, D.; Zhang, J. The complete mitochondrial genome of Pyxicephalus adspersus: High gene rearrangement and phylogenetics of one of the world’s largest frogs. PeerJ 2019, 7, e7532. [Google Scholar] [CrossRef]
- Li, F.; Lv, Y.; Wen, Z.; Bian, C.; Zhang, X.; Guo, S.; Shi, Q.; Li, D. The complete mitochondrial genome of the intertidal spider (Desis jiaxiangi) provides novel insights into the adaptive evolution of the mitogenome and the evolution of spiders. BMC Ecol. Evol. 2021, 21, 72. [Google Scholar] [CrossRef]
- Zhang, J.; Luu, B.E.; Yu, D.; Zhang, L.; Al-Attar, R.; Storey, K.B. The complete mitochondrial genome of Dryophytes versicolor: Phylogenetic relationship among Hylidae and mitochondrial protein-coding gene expression in response to freezing and anoxia. Int. J. Biol. Macromol. 2019, 132, 461–469. [Google Scholar] [CrossRef]
- Wang, Z.; Shi, X.; Tao, Y.; Wu, Q.; Bai, Y.; Guo, H.; Tang, D. The complete mitochondrial genome of Parasesarma pictum (Brachyura: Grapsoidea: Sesarmidae) and comparison with other Brachyuran crabs. Genomics 2019, 111, 799–807. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Z.; Tang, D.; Xu, X.; Tao, Y.; Ji, C.; Wang, Z. Characterization and comparison of the mitochondrial genomes from two Alpheidae species and insights into the phylogeny of Caridea. Genomics 2020, 112, 65–70. [Google Scholar] [CrossRef]
- Chen, G.; Wang, B.; Liu, J.; Xie, F.; Jiang, J. Complete mitochondrial genome of Nanorana pleskei (Amphibia: Anura: Dicroglossidae) and evolutionary characteristics. Curr. Zool. 2011, 57, 785–805. [Google Scholar] [CrossRef]
- Xin, Z.; Liu, Y.; Zhu, X.; Wang, Y.; Zhang, H.; Zhang, D.; Zhou, C.; Tang, B.; Liu, Q. Mitochondrial genomes of two Bombycoidea insects and implications for their phylogeny. Sci. Rep. 2017, 7, 6544. [Google Scholar] [CrossRef] [PubMed]
- AmphibiaWeb. Available online: https://amphibiaweb.org (accessed on 20 July 2023).
- Yan, F.; Jiang, K.; Chen, H.; Fang, P.; Jin, J.; Li, Y.; Wang, S.; Murphy, R.W.; Che, J.; Zhang, Y. Matrilineal history of the Rana longicrus species group (Rana, Ranidae, Anura) and the description of a new species from Hunan, southern China. Asian Herpetol. Res. 2011, 2, 61–71. [Google Scholar]
- Boulenger, G.A. A monograph of the South Asian, Papuan, Melanesian, and Australian frogs of the genus Rana. Rec. Ind. Mus. 1920, 20, 1–126. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, S.; Zhu, H.; Li, P.; Yang, B.; Ma, J. Phylogeny and biogeography of South Chinese brown frogs (Ranidae, Anura). PLoS ONE 2017, 12, e175113. [Google Scholar] [CrossRef]
- Yuan, Z.; Zhou, W.; Chen, X.; Poyarkov, N.A.; Chen, H.; Jang-Liaw, N.; Chou, W.; Matzke, N.J.; Iizuka, K.; Min, M.; et al. Spatiotemporal diversification of the true frogs (genus Rana): A historical framework for a widely studied group of model organisms. Syst. Biol. 2016, 65, 824–842. [Google Scholar] [CrossRef] [PubMed]
- Fei, L.; Hu, S.Q.; Ye, C.Y.; Huang, Y.Z. Amphibia Vol. 3. Anura Ranidae. Fauna Sinica; Science Press: Beijing, China, 2009; pp. 1–22. [Google Scholar]
- Che, J.; Pang, J.; Zhao, E.; Matsui, M.; Zhang, Y. Phylogenetic relationships of the Chinese brown frogs (genus Rana) inferred from partial mitochondrial 12S and 16S rRNA gene sequences. Zool. Sci. 2007, 24, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Fei, L.; Ye, C.Y. On taxonomic status and relationships of Rana japonica group. China (Amphibia: Anura: Radidae). Cultum. Herpetol. Sin. 2000, 8, 74–80. [Google Scholar]
- Yang, B.; Zhou, Y.; Min, M.; Matsui, M.; Dong, B.; Li, P.; Fong, J.J. Diversity and phylogeography of Northeast Asian brown frogs allied to Rana dybowskii (Anura, Ranidae). Mol. Phylogenet. Evol. 2017, 112, 148–157. [Google Scholar] [CrossRef]
- Wan, H.; Lyu, Z.; Qi, S.; LIU, L.; Wang, Y. A new species of the Rana japonica group (Anura, Ranidae, Rana) from China, with a taxonomic proposal for the R. johnsi group. Zookeys 2020, 942, 141–158. [Google Scholar] [CrossRef]
- Zhu, S.; Zhang, X.; Zhang, S.; Zhang, R.; Liu, S.; Xiong, R. Molecular identification of a Rana hanluica population distributed in Youyang County, Chongqing City in China. Open J. Nat. Sci. 2018, 6, 1–8. [Google Scholar] [CrossRef]
- Pope, C.H.; Boring, A.M. A survey of Chinese Amphibia. Peking Nat. Hist. Bull. 1940, 15, 13–86. [Google Scholar]
- Wu, Y.; Yan, F.; Stuart, B.L.; Prendini, E.; Suwannapoom, C.; Dahn, H.A.; Zhang, B.; Cai, H.; Xu, Y.; Jiang, K.; et al. A combined approach of mitochondrial DNA and anchored nuclear phylogenomics sheds light on unrecognized diversity, phylogeny, and historical biogeography of the torrent frogs, genus Amolops (Anura: Ranidae). Mol. Phylogenet. Evol. 2020, 148, 106789. [Google Scholar] [CrossRef]
- Jin, J.; Yu, W.; Yang, J.; Song, Y.; DePamphilis, C.W.; Yi, T.; Li, D. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Meng, G.; Li, Y.; Yang, C.; Liu, S. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Yang, Z. Computational Molecular Evolution; Oxford University Press: Oxford, UK; New York, NY, USA, 2006. [Google Scholar]
- Dong, B.; Zhou, Y.; Yang, B. The complete mitochondrial genome of the Rana huanrensis (Anura: Ranidae). Mitochondrial DNA A 2016, 27, 4551–4552. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yin, W.; Xia, R.; Lei, G.; Fu, C. Complete mitochondrial genome of a brown frog, Rana kunyuensis (Anura: Ranidae). Mitochondrial DNA A 2016, 27, 34–35. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lei, G.; Fu, C. Complete mitochondrial genomes of two brown frogs, Rana dybowskii and Rana cf. chensinensis (Anura: Ranidae). Mitochondrial DNA A 2016, 27, 155–156. [Google Scholar] [CrossRef] [PubMed]
- Peso-Fernández, M.; Ponti De La Iglesia, R.; Ponz Segrelles, G.; Gonzalez Martinez, R.; Arcones Segovia, A.; Vieites, D.R. The complete mitochondrial genome of the Endangered European brown frog Rana pyrenaica through RNAseq. Mitochondrial DNA B 2016, 1, 394–396. [Google Scholar] [CrossRef]
- Liu, P.; Wang, H.; Zhao, W. Sequencing and analysis of the complete mitochondrial genome of Rana amurensis (Anura: Ranidae). Mitochondrial DNA B 2017, 2, 424–425. [Google Scholar] [CrossRef]
- Chen, J. The complete mitochondrial genome of common terrestrial frog (Rana temporaria). Mitochondrial DNA B 2018, 3, 978–979. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, M.; Deng, L.; Xu, Z.; Shi, H.; Jia, X.; Lai, Z.; Ruan, Q.; Chen, W. Characteristics of the mitochondrial genome of Rana omeimontis and related species in Ranidae: Gene rearrangements and phylogenetic relationships. Ecol. Evol. 2020, 10, 12817–12837. [Google Scholar] [CrossRef]
- Wang, J.; Li, Z.; Gao, H.; Liu, Z.; Teng, L. The complete mitochondrial genome of the Rana kukunoris (Anura: Ranidae) from Inner Mongolia, China. Mitochondrial DNA B 2020, 5, 586–587. [Google Scholar] [CrossRef] [PubMed]
- Suk, H.Y.; Jeon, J.Y.; Kim, D.; Cha, S.; Min, M. The complete mitochondrial genome information of Rana uenoi (Amphibia, Anura, Ranidae) and the phylogenetic implication. Mitochondrial DNA B 2021, 6, 689–690. [Google Scholar] [CrossRef]
- Huang, M.; Zhao, Q.; Wu, Y. Characterization of the complete mitochondrial genome of Zhenhai brown frog Rana zhenhaiensis (Anura: Ranidae). Mitochondrial DNA B 2019, 4, 3204–3205. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Duan, R.; Kong, X.; Wang, H.; Zhu, H. The complete mitochondrial genome of Amolops wuyiensis (Anura: Ranidae). Mitochondrial DNA A 2016, 27, 1705–1706. [Google Scholar]
- Li, Y.; Wu, X.; Zhang, H.; Yan, P.; Xue, H.; Wu, X. The complete mitochondrial genome of Amolops ricketti (Amphidia, Anura, Ranidae). Mitochondrial DNA A 2016, 27, 242–243. [Google Scholar] [CrossRef]
- Jin, X.; Li, W.; Hu, S.; Li, W.; Yang, J. The complete mitochondrial genome of large odorous frog, Odorrana graminea (Amphibia: Ranidae) and phylogenetic analysis. Mitochondrial DNA B 2020, 5, 3139–3140. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Pereira, S.L. Mitochondrial genome organization and vertebrate phylogenetics. Genet. Mol. Biol. 2000, 23, 745–752. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Wu, X.; Xue, H.; Yan, P.; Wu, X. A novel mitogenomic rearrangement for Odorrana schmackeri (Anura: Ranidae) and phylogeny of Ranidae inferred from thirteen mitochondrial protein-coding genes. Amphib.-Reptil. 2014, 35, 331–343. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, J.; Zhai, X.; Zhu, Y.; Chen, X. Complete mitochondrial genome of the green odorous frog Odorrana margaretae (Anura: Ranidae). Mitochondrial DNA 2015, 26, 487–488. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, J.; Liu, J.; Zhou, M.; Li, B.; Ouyang, B. Three new Ranidae mitogenomes and the evolution of mitochondrial gene rearrangements among Ranidae species. Asian Herpetol. Res. 2018, 9, 85–98. [Google Scholar]
- Jeffrey, L.B. Survey and summary animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef]
- Jiang, L.; Zhao, L.; Cheng, D.; Zhu, L.; Zhang, M.; Ruan, Q.; Chen, W. The complete mitochondrial genome sequence of the Sichuan Digging Frog, Kaloula rugifera (Anura: Microhylidae) and its phylogenetic implications. Gene 2017, 626, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Kurisaki, M.; Hashiguchi, Y.; Kumazawa, Y. Variation and evolution of polyadenylation profiles in sauropsid mitochondrial mRNAs as deduced from the high-throughput RNA sequencing. BMC Genom. 2017, 18, 665. [Google Scholar] [CrossRef]
- Mayfield, J.E.; McKenna, J.F. AT rich sequences in vertebrate DNA. Chromosoma 1978, 67, 157–163. [Google Scholar] [CrossRef]
- Cai, Y.; Li, Q.; Zhang, J.; Storey, K.B.; Yu, D. Characterization of the mitochondrial genomes of two toads, Anaxyrus americanus (Anura: Bufonidae) and Bufotes pewzowi (Anura: Bufonidae), with phylogenetic and selection pressure analyses. PeerJ 2020, 8, e8901. [Google Scholar] [CrossRef]
- Shahzadi, I.; Mehmood, F.; Ali, Z.; Malik, M.S.; Waseem, S.; Mirza, B.; Ahmed, I.; Waheed, M.T. Comparative analyses of chloroplast genomes among three Firmiana species: Identification of mutational hotspots and phylogenetic relationship with other species of Malvaceae. Plant Gene 2019, 19, 100199. [Google Scholar]
- Zhang, X.; Liu, P.; Xu, S.; Rizo, E.Z.; Zhang, Q.; Dumont, H.J.; Han, B. Geographic Variation of Phyllodiaptomus tunguidus Mitogenomes: Genetic Differentiation and Phylogeny. Front. Genet. 2021, 12, 711992. [Google Scholar] [CrossRef]
- Chen, L.; Lin, Y.; Xiao, Q.; Lin, Y.; Du, Y.; Lin, C.; Ward-Fear, E.; Hu, C.; Qu, Y.; Li, H. Characterization of the complete mitochondrial genome of the many-lined sun skink (Eutropis multifasciata) and comparison with other Scincomorpha species. Genomics 2020, 113, 2526–2536. [Google Scholar] [CrossRef] [PubMed]
- Kurabayashi, A.; Ueshima, R. Complete sequence of the mitochondrial DNA of the primitive opisthobranch gastropod Pupa strigosa: Systematic implication of the genome organization. Mol. Biol. Evol. 2000, 17, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Wolstenholme, D.R. Animal mitochondrial DNA: Structure and evolution. Int. Rev. Cyto. 1992, 141, 173–216. [Google Scholar]
- Wang, C.; Qian, L.; Zhang, C.; Guo, W.; Pan, T.; Wu, J.; Wang, H.; Zhang, B. A new species of Rana from the Dabie Mountains in eastern China (Anura, Ranidae). Zookeys 2017, 724, 135. [Google Scholar] [CrossRef]
Figure 1.
Circular map of the complete mitochondrial genomes of (A) Rana hanluica and (B) Rana longicrus.
Figure 1.
Circular map of the complete mitochondrial genomes of (A) Rana hanluica and (B) Rana longicrus.

Figure 2.
Codon number and relative synonymous codon usage of protein-coding genes in R. hanluica and R. longicrus mitogenomes.
Figure 2.
Codon number and relative synonymous codon usage of protein-coding genes in R. hanluica and R. longicrus mitogenomes.

Figure 3.
The evolutionary rate indicated by the ratios of non-synonymous to synonymous substitutions (Ka/Ks) for each PCG among the 16 representative Rana mitogenomes.
Figure 3.
The evolutionary rate indicated by the ratios of non-synonymous to synonymous substitutions (Ka/Ks) for each PCG among the 16 representative Rana mitogenomes.

Figure 4.
Phylogenetic tree for the genus Rana using BI and ML analyses based on nucleotide sequences. Bayesian posterior probability (BPP) and bootstrap values (BP) of each node are shown as BPP/BP, with maxima of 1.00/100. Samples sequenced in the present study are highlighted in red.
Figure 4.
Phylogenetic tree for the genus Rana using BI and ML analyses based on nucleotide sequences. Bayesian posterior probability (BPP) and bootstrap values (BP) of each node are shown as BPP/BP, with maxima of 1.00/100. Samples sequenced in the present study are highlighted in red.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The mitogenome composition of the two Rana species.
| Region | Size (bp) | A + T Content (%) | G + C Content (%) | AT−Skew | GC−Skew | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| RH | RL | RH | RL | RH | RL | RH | RL | RH | RL | |
| Whole genome | 19,395 | 17,833 | 59.1 | 55.6 | 40.9 | 44.4 | −0.045 | −0.007 | −0.314 | −0.328 |
| PCGs | 11,279 | 11,288 | 53.9 | 54 | 46.1 | 46 | −0.088 | −0.082 | −0.322 | −0.321 |
| rRNA genes | 2509 | 2509 | 55.4 | 55.9 | 44.6 | 44.1 | 0.157 | 0.162 | −0.15 | −0.148 |
| tRNA genes | 1530 | 1530 | 57.6 | 56.9 | 42.4 | 43.1 | 0.015 | 0.024 | 0.045 | 0.032 |
| CR | 3602 | 1930 | 78.8 | 64.2 | 21.2 | 35.8 | −0.134 | −0.031 | −0.251 | −0.294 |
RH: Rana hanluica; RL: Rana longicrus.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Y.-M.; Zhang, C.-Y.; Luo, S.-T.; Ding, G.-H.; Qiao, F. Characterization and Comparison of the Two Mitochondrial Genomes in the Genus Rana. Genes 2023, 14, 1786. https://doi.org/10.3390/genes14091786
AMA Style
Wang Y-M, Zhang C-Y, Luo S-T, Ding G-H, Qiao F. Characterization and Comparison of the Two Mitochondrial Genomes in the Genus Rana. Genes. 2023; 14(9):1786. https://doi.org/10.3390/genes14091786
Chicago/Turabian StyleWang, Yan-Mei, Chi-Ying Zhang, Si-Te Luo, Guo-Hua Ding, and Fen Qiao. 2023. "Characterization and Comparison of the Two Mitochondrial Genomes in the Genus Rana" Genes 14, no. 9: 1786. https://doi.org/10.3390/genes14091786
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.