
Inbreeding Effects on the Performance and Genomic Prediction for Polysomic Tetraploid Potato Offspring Grown at High Nordic Latitudes

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
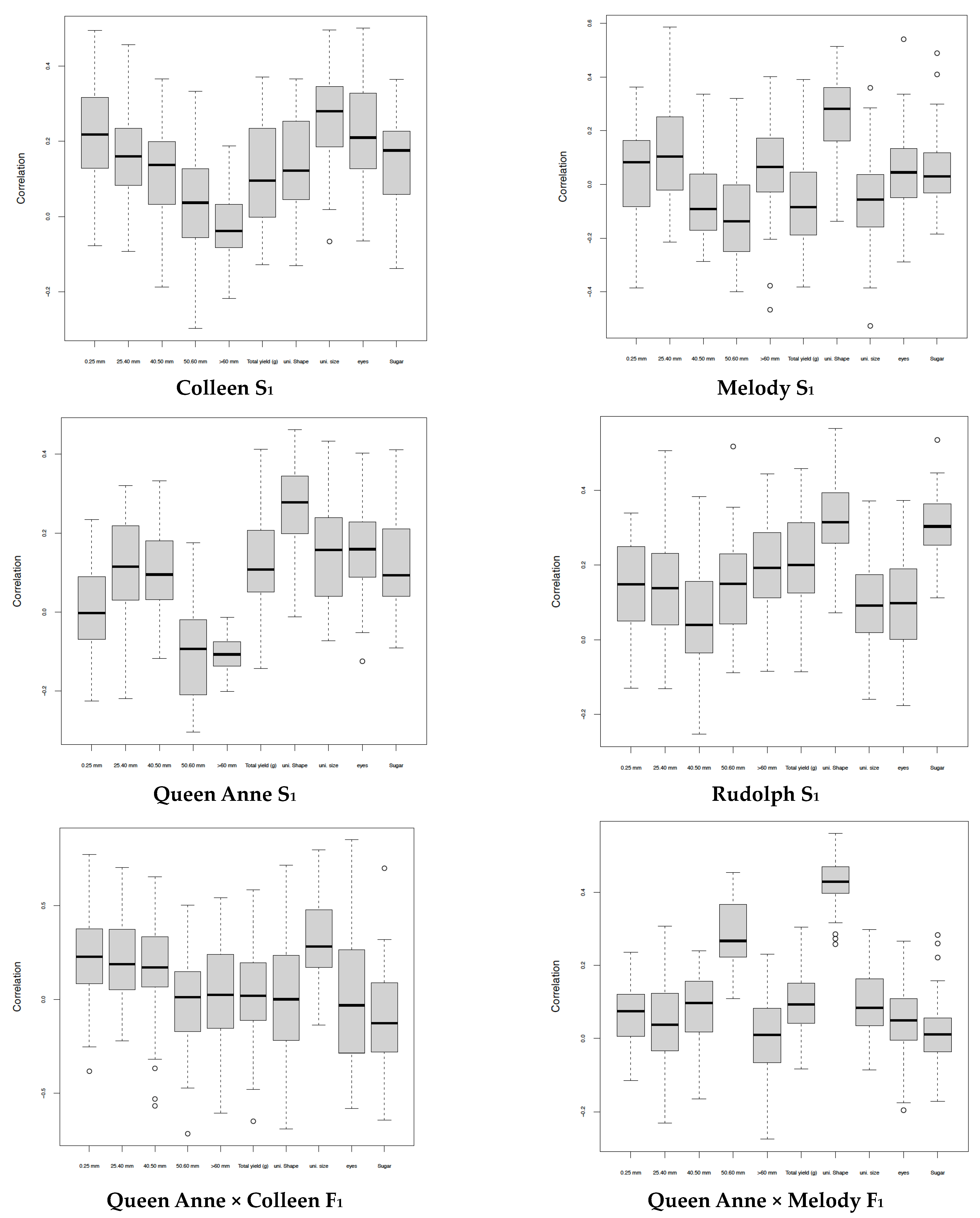
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Svačina, R.; Sourdille, P.; Kopecký, D.; Bartoš, J. Chromosome pairing in polyploid grasses. Front. Plant Sci. 2020, 11, 1056. [Google Scholar] [CrossRef]
- Manrique-Carpintero, N.C.; Coombs, J.J.; Pham, G.M.; Laimbeer, F.P.E.; Braz, G.T.; Jiang, J.; Veilleux, R.E.; Buell, C.R.; Douches, D.S. Genome reduction in tetraploid potato reveals genetic load, haplotype variation, and loci associated with agronomic traits. Front. Plant Sci. 2018, 9, 944. [Google Scholar] [CrossRef] [Green Version]
- Lian, Q.; Tang, D.; Bai, Z.; Qi, J.; Lu, F.; Huang, S.; Zhang, C. Acquisition of deleterious mutations during potato polyploidization. J. Integr. Plant Biol. 2019, 61, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, P.; Tang, D.; Yang, Z.; Lu, F.; Qi, J.; Tawari, N.R.; Shang, Y.; Li, C.; Huang, S. The genetic basis of inbreeding depression in potato. Nat. Genet. 2019, 51, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Labroo, M.R.; Endelman, J.B.; Gemenet, D.C.; Werner, C.R.; Gaynor, R.C.; Covarrubias-Pazaran, G.E. Clonal breeding strategies to harness heterosis insights from stochastic simulation. bioRxiv 2022. [Google Scholar] [CrossRef]
- Charlesworth, D.; Willis, J. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Busbice, T.H.; Wilsie, C.P. Inbreeding depression and heterosis in autotetraploids with application to Medicago sativa L. Euphytica 1966, 15, 52–67. [Google Scholar] [CrossRef]
- Glendinning, D.R. Estimation of inbreeding in potato pedigrees. Potato Res. 1997, 40, 277–284. [Google Scholar] [CrossRef]
- Malecot, G. Les Mathámatiques de L’hérédité; Masson et Cie: Paris, France, 1948. [Google Scholar]
- Kempthorne, O. An Introduction to Genetic Statistics; Iowa State University Press: Ames, Iowa, 1957. [Google Scholar]
- Mendoza, H.A.; Haynes, F.L. Some aspects of breeding and inbreeding in potatoes. Am. Potato J. 1973, 50, 216–222. [Google Scholar] [CrossRef]
- Hagberg, A.; Tedin, O. Inter- and intra-clonal crosses and inbreeding in potatoes. Hereditas 1951, 37, 280–287. [Google Scholar] [CrossRef]
- Mendoza, H.A.; Haynes, F.L. Genetic relationship among potato cultivars grown in the United States. HortScience 1974, 9, 328–330. [Google Scholar] [CrossRef]
- Chen, Z.J. Molecular mechanisms of polyploidy and hybrid vigor. Trends Plant Sci. 2010, 15, 57–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, H.A.; Haynes, F.L. Genetic basis of heterosis for yield in the autotetraploid potato. Theor. Appl. Genet. 1974, 45, 21–25. [Google Scholar] [CrossRef]
- Bonierbale, M.W.; Plaisted, R.L.; Tanksley, S.D. A test of the maximum heterozygosity hypothesis using molecular markers in tetraploid potatoes. Theor. Appl. Genet. 1993, 86, 481–491. [Google Scholar] [CrossRef]
- Muthoni, J.; Shimelis, H.; Melis, R. Production of hybrid potatoes: Are heterozygosity and ploidy levels important? Aust. J. Crop Sci. 2019, 13, 687–694. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, Z.; Tang, D.; Zhu, Y.; Wang, P.; Li, D.; Zhu, G.; Xiong, X.; Shang, Y.; Li, C.; et al. Genome design of hybrid potato. Cell 2021, 84, 3873–3883.e12. [Google Scholar] [CrossRef]
- Hirsch, C.N.; Hirsch, C.D.; Felcher, K.; Coombs, J.; Zarka, D.; Van Deynze, A.; De Jong, W.; Veilleux, R.E.; Jansky, S.; Bethke, P.; et al. Retrospective view of North American potato (Solanum tuberosum L.) breeding in the 20th and 21st centuries. G3 2013, 3, 1003–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramu, P.; Esuma, W.; Kawuki, R.; Rabbi, I.Y.; Egesi, C.; Bredeson, J.V.; Bart, R.S.; Verma, J.; Buckler, E.S.; Lu, F. Cassava haplotype map highlights fixation of deleterious mutations during clonal propagation. Nat. Genet. 2017, 49, 959–963. [Google Scholar] [CrossRef] [Green Version]
- Karunarathna, N.L. Genomic background selection to reduce the mutation load after random mutagenesis. Sci. Rep. 2021, 11, 19404. [Google Scholar] [CrossRef]
- Momo, S.; Jannink, J.-L. Incorporating selfing to purge deleterious alleles in a cassava genomic selection program. bioRxiv 2020. [Google Scholar] [CrossRef]
- Federer, W. Augmented designs. Hawaii. Plant. Recorder. 1956, 55, 191–208. [Google Scholar] [CrossRef] [Green Version]
- Mann, D.J.; Lammerink, J.P.; Coles, G.D. Predicting potato crisp darkening: Two methods for analysis of glucose. N. Z. J. Crop Hortic. Sci. 1991, 19, 199–201. [Google Scholar] [CrossRef]
- Selga, S.; Reslow, R.; Pérez-Rodríguez, P.; Ortiz, R. The power of genomic estimated breeding values for selection when using a finite population size in genetic improvement of tetraploid potato. G3 2022, 12, jkab362. [Google Scholar] [CrossRef] [PubMed]
- Huaman, Z.; Williams, J.T.; Salhuana, W.; Vincent, L. Descriptors for the Cultivated Potato and the Maintenance and Distribution of Germplasm Collections; International Board for Plant Genetic Resources: Rome, Italy, 1977. [Google Scholar]
- Selga, C.; Koc, A.; Chawade, A.; Ortiz, R. A bioinformatics pipeline to identify a subset of SNPs for genomics-assisted potato breeding. Plants 2021, 10, 30. [Google Scholar] [CrossRef]
- Oliveira, I.B.; Resende, M.F.R., Jr.; Ferrão, L.F.V.; Amadeu, R.R.; Endelman, J.B.; Kirst, M.; Coelho, A.S.G.; Munoz, P.R. Genomic prediction of autotetraploids: Influence of relationship matrices, allele dosage, and continuous genotyping calls in phenotype prediction. G3 2019, 9, 1189–1198. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, R.; Crossa, J.; Reslow, F.; Perez-Rodriguez, P.; Cuevas, J. Genome-based genotype environment prediction enhances potato (Solanum tuberosum L.) improvement using pseudo-diploid and polysomic tetraploid modeling. Front. Plant Sci. 2022, 13, 785196. [Google Scholar] [CrossRef]
- Cotes, T.J.M.; Ñústez, L.C.E. Propuesta para el análisis de diseños aumentados en fitomejoramiento: Un caso en papa. Rev. Latinoam. Papa 2001, 12, 15–34. [Google Scholar] [CrossRef]
- Gardner, C.O.; Eberhart, S.A. Analysis and interpretation of the variety cross diallel and related populations. Biometrics 1966, 22, 439–452. [Google Scholar] [CrossRef]
- Slater, A.T.; Cogan, N.O.; Forster, J.W.; Hayes, B.J.; Daetwyler, H.D. Improving genetic gain with genomic selection in autotetraploid potato. Plant Genome 2016, 9, plantgenome2016.02.0021. [Google Scholar] [CrossRef] [Green Version]
- Amadeu, R.R.; Cellon, C.; Olmstead, J.W.; Garcia, A.A.F.; Resende, M.F.R.; Munoz, P.R. AGHmatrix: R package to construct relationship matrices for autotetraploid and diploid species: A blueberry example. Plant Genome 2016, 9, plantgenome2016.01.0009. [Google Scholar] [CrossRef]
- Pérez, P.; de los Campos, G. Genome-wide regression and prediction with the BGLR statistical package. Genetics 2014, 198, 483–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 9 January 2023).
- Hoopes, G.M.; Zarka, D.; Feke, A.; Acheson, K.; Hamilton, J.P.; Douches, D.; Buell, C.R.; Farré, E.M. Keeping time in the dark: Potato diel and circadian rhythmic gene expression reveals tissue-specific circadian clocks. Plant Direct 2022, 6, e425. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Li, G.; Jiang, Y.; Duan, S.; Liu, J.; Xu, J.; Jin, L. Genes related to circadian rhythm are involved in regulating tuberization time in potato. Hortic. Plant J. 2022, 8, 369–380. [Google Scholar] [CrossRef]
- Zhou, T.; Song, B.; Liu, T.; Shen, Y.; Dong, L.; Jing, S.; Xie, C.; Liu, J. Phytochrome F plays critical roles in potato photoperiodic tuberization. Plant J. 2019, 98, 42–54. [Google Scholar] [CrossRef]
- Rodríguez-Falcón, M.; Bou, J.; Prat, S. Seasonal control of tuberization in potato: Conserved elements with the flowering response. Annu. Rev. Plant Biol. 2006, 57, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Golmirzaie, A.; Ortiz, R.; Atlin, G.; Iwanaga, M. Inbreeding and true seed in tetrasomic potato. I. Selfing and open pollination in Andean landraces (Solanum tuberosum Gp. Andigena). Theor. Appl. Genet. 1998, 97, 1125–1128. [Google Scholar] [CrossRef]
- Golmirzaie, A.; Bretschneider, K.; Ortiz, R. Inbreeding and true seed in tetrasomic potato. II. Selfing and sib-mating in heterogeneous hybrid populations of Solanum tuberosum. Theor. Appl. Genet. 1998, 97, 1129–1132. [Google Scholar] [CrossRef]
- Krantz, F.A. Potato breeding methods. In Technical Bulletin; University of Minnesota Department of Agriculture: St Paul, MN, USA, 1924; Volume 25. [Google Scholar]
- Krantz, F.A.; Hutchins, A.E. Technical Bulletin; University of Minnesota Department of Agriculture: St Paul, MN, USA, 1929; Volume 58. [Google Scholar]
- Mendiburu, A.O.; Peloquin, S.J. The significance of 2n gametes in potato breeding. Theor. Appl. Genet. 1977, 49, 53–61. [Google Scholar] [CrossRef]
- Batista, L.G.; Mello, V.H.; Souza, A.P.; Margarido, G.R.A. Genomic prediction with allele dosage information in highly polyploid species. Theor. Appl. Genet. 2022, 135, 723–739. [Google Scholar] [CrossRef]
- Amadeu, R.R.; Ferrão, L.F.V.; Oliveira, I.d.B.; Benevenuto, J.; Endelman, J.B.; Munoz, P.R. Impact of dominance effects on autotetraploid genomic prediction. Crop Sci. 2020, 60, 656–665. [Google Scholar] [CrossRef]
- Ortiz, R.; Reslow, F.; Crossa, J.; Cuevas, J. Heritable variation, genetic and phenotypic correlations for tuber traits and host plant resistance to late blight for potato breeding in Scandinavian testing sites. Agriculture 2021, 11, 1287. [Google Scholar] [CrossRef]
- Endelman, J.B.; Carley, C.A.S.; Bethke, P.C.; Coombs, J.J.; Clough, M.E.; da Silva, W.L.; De Jong, W.S.; Douches, D.S.; Frederick, C.M.; Haynes, K.G.; et al. Genetic variance partitioning and genome-wide prediction with allele dosage information in autotetraploid potato. Genetics 2018, 209, 77–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuevas, J.; Reslow, F.; Crossa, J.; Ortiz, R. Modeling genotype × environment interaction for single and multi-trait genomic prediction in potato (Solanum tuberosum L.). G3 2022, 13, jkac322. [Google Scholar] [CrossRef] [PubMed]
- Zingaretti, L.M.; Gezan, S.A.; Ferrão, L.F.V.; Osorio, L.F.; Monfort, A.; Muñoz, P.R.; Whitaker, V.M.; Pérez-Enciso, M. Exploring deep learning for complex trait genomic prediction in polyploid outcrossing species. Front. Plant Sci. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Atlin, G. Inbreeding in TPS progenies: Implications for breeding and seed production strategy. In Proceedings of the Present and Future Strategies for Potato Breeding and Improvement: Report of the XXVI Planning Conference, Lima, Peru, 12–14 December 1983; Centro Internacional de la Papa: Lima, Peru, 1985; pp. 71–85. [Google Scholar]


{kind=link}
| Cultivar or Offspring | Tuber Weight (4-Plant Plot, g) | Tuber Uniformity Z | Tuber Eye Depth Y | Tuber Flesh Reducing Sugar | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| <25 mm | 25–40 mm | 40–50 mm | 50–60 mm | >60 mm | Total | Shape | Size | |||
| Cultivars (S0) | ||||||||||
| Colleen | 34 | 280 | 528 | 1118 | 810 | 2771 | 5.3 | 5.3 | 5.0 | 0.00 |
| Melody | 23 | 452 | 844 | 1349 | 637 | 3304 | 5.3 | 6.6 | 5.0 | 0.00 |
| Queen Anne | 43 | 513 | 1010 | 721 | 161 | 2247 | 6.1 | 5.6 | 6.0 | 0.22 |
| First inbred generation (S1) | ||||||||||
| Colleen | 39 | 219 | 240 | 176 | 51 | 725 | 5.2 | 5.2 | 4.6 | 0.32 |
| Melody | 66 | 291 | 298 | 103 | 59 | 781 | 5.4 | 4.9 | 4.9 | 0.22 |
| Queen Anne | 60 | 249 | 139 | 67 | 11 | 497 | 6.7 | 5.4 | 5.4 | 0.47 |
| Rudolph | 26 | 133 | 248 | 284 | 214 | 889 | 5.0 | 4.6 | 4.0 | 0.92 |
| Hybrid offspring (F1) | ||||||||||
| Queen Anne × Colleen | 80 | 257 | 378 | 312 | 136 | 1162 | 6.0 | 4.6 | 4.7 | 0.69 |
| Queen Anne × Melody | 53 | 318 | 497 | 552 | 169 | 1546 | 6.1 | 4.7 | 4.9 | 0.42 |
| LSD0.05 | 27 | 104 | 145 | 189 | 136 | 378 | 0.9 | 0.5 | 0.4 | 0.41 |
| Statistical significance (P > Fc) of contrasts | ||||||||||
| S0 vs. S1 | 0.1086 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.8898 | <0.0001 | <0.0001 | 0.0031 |
| S0 vs. F1 | 0.0008 | 0.0008 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0228 | <0.0001 | 0.0002 | 0.0012 |
| S1 vs. F1 | 0.0002 | 0.0014 | <0.0001 | <0.0001 | 0.0059 | <0.0001 | <0.0001 | <0.0001 | 0.1618 | 0.3104 |
| Training set | Testing sets | |||||
|---|---|---|---|---|---|---|
| ‘Queen Anne’ S1 | ‘Colleen’ S1 | ‘Melody’ S1 | ‘Queen Anne’ × ‘Colleen’ F1 | ‘Queen Anne’ × ‘Melody’ F1 | ‘Rudolph’ S1 | |
| ‘Queen Anne’ S1 | ||||||
| ‘Colleen’ S1 | ||||||
| ‘Melody’ S1 | ||||||
| ‘Queen Anne’ × ‘Colleen’ F1 | ||||||
| ‘Queen Anne’ × ‘Melody’ F1 | ||||||
| ‘Rudolph’ S1 | ||||||
| Training and Validating Offspring | Tuber Weight (4-Plant Plot, g) | Tuber Uniformity | Tuber Eye Depth | Tuber Flesh Reducing Sugar | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| <25 mm | 25–40 mm | 40–50 mm | 50–60 mm | >60 mm | Total | Shape | Size | |||
| Full-sibs S1 inbred offspring | ||||||||||
| A. Colleen | 0.227 | 0.145 | 0.120 | 0.032 | −0.022 | 0.105 | 0.136 | 0.266 | 0.217 | 0.142 |
| B. Melody | 0.050 | 0.119 | −0.048 | −0.120 | 0.061 | −0.070 | 0.246 | −0.055 | 0.045 | 0.056 |
| C. Queen Anne | −0.007 | 0.143 | 0.122 | −0.196 | −0.053 | −0.143 | 0.109 | 0.186 | −0.125 | 0.218 |
| D. Rudolph | 0.137 | 0.136 | 0.061 | 0.138 | 0.202 | 0.206 | 0.316 | 0.097 | 0.090 | 0.297 |
| Full-sibs F1 hybrid offspring | ||||||||||
| Y. Queen Anne × Colleen | 0.218 | 0.213 | 0.157 | 0.002 | 0.026 | 0.036 | 0.009 | 0.320 | −0.001 | −0.096 |
| Z. Queen Anne × Melody | 0.070 | 0.049 | 0.088 | 0.287 | 0.011 | 0.094 | 0.420 | 0.095 | 0.048 | 0.021 |
| Inbred full-sibs S1 (training population)—half-sib F1 hybrids (breeding population) | ||||||||||
| A.–Y. | 0.024 | 0.083 | 0.232 | 0.100 | 0.111 | 0.311 | 0.444 | 0.104 | 0.165 | 0.245 |
| B.–Z. | −0.088 | −0.013 | 0.140 | 0.059 | 0.108 | 0.100 | 0.217 | −0.085 | 0.060 | 0.014 |
| C.–Y. | 0.221 | −0.062 | −0.100 | −0.152 | −0.393 | −0.224 | 0.106 | 0.310 | 0.235 | 0.379 |
| C.–Z. | 0.159 | 0.076 | 0.156 | 0.208 | 0.100 | 0.264 | 0.135 | 0.026 | 0.030 | 0.112 |
| Half-sib F1 hybrids (one as a training population and the other as a breeding population) | ||||||||||
| Y.–Z. | 0.100 | −0.110 | 0.141 | −0.148 | −0.093 | −0.057 | −0.042 | −0.084 | 0.018 | −0.023 |
| Z.–Y. | 0.147 | −0.153 | 0.354 | −0.266 | −0.206 | −0.011 | 0.166 | −0.226 | 0.104 | −0.275 |
| Inbred S1 (training population)—non-related F1 (breeding population) | ||||||||||
| A.–Z. | 0.096 | 0.114 | −0.078 | −0.109 | 0.056 | −0.021 | 0.066 | −0.072 | 0.076 | 0.158 |
| B.–Y. | −0.170 | 0.090 | 0.079 | 0.171 | −0.151 | 0.282 | 0.105 | −0.008 | −0.082 | −0.098 |
| D.–Y. | 0.214 | 0.164 | 0.173 | 0.116 | −0.037 | 0.001 | −0.040 | −0.136 | 0.130 | 0.097 |
| D.–Z. | 0.106 | 0.136 | 0.160 | −0.029 | 0.003 | 0.123 | 0.225 | 0.064 | 0.024 | 0.012 |
| Among inbred S1 offspring (one as a training population and the other as a breeding population) | ||||||||||
| A.–B. | −0.227 | −0.074 | −0.015 | −0.132 | 0.202 | −0.058 | 0.171 | −0.118 | 0.028 | −0.281 |
| A.–C. | 0.042 | −0.139 | −0.096 | 0.131 | 0.077 | 0.093 | −0.013 | 0.035 | 0.193 | −0.077 |
| B.–C. | −0.083 | −0.050 | 0.083 | −0.052 | 0.076 | 0.099 | −0.098 | 0.166 | −0.134 | 0.185 |
| B.–A. | −0.173 | −0.013 | 0.034 | −0.091 | −0.014 | 0.004 | 0.140 | −0.055 | 0.093 | −0.127 |
| C.–A. | 0.081 | −0.111 | −0.125 | 0.053 | −0.002 | 0.054 | 0.018 | 0.116 | 0.165 | −0.083 |
| C.–B. | −0.185 | −0.164 | 0.044 | 0.108 | −0.058 | −0.042 | −0.076 | 0.067 | −0.028 | 0.193 |
| D.–A. | 0.154 | 0.148 | −0.033 | 0.048 | 0.110 | 0.118 | 0.230 | −0.050 | 0.094 | −0.053 |
| D.–B. | −0.097 | −0.151 | −0.013 | −0.008 | 0.024 | 0.014 | 0.002 | −0.066 | −0.295 | 0.146 |
| D.–C. | −0.080 | −0.066 | 0.019 | −0.069 | 0.174 | 0.030 | 0.006 | −0.179 | 0.015 | 0.136 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz, R.; Reslow, F.; Vetukuri, R.; García-Gil, M.R.; Pérez-Rodríguez, P.; Crossa, J. Inbreeding Effects on the Performance and Genomic Prediction for Polysomic Tetraploid Potato Offspring Grown at High Nordic Latitudes. Genes 2023, 14, 1302. https://doi.org/10.3390/genes14061302
Ortiz R, Reslow F, Vetukuri R, García-Gil MR, Pérez-Rodríguez P, Crossa J. Inbreeding Effects on the Performance and Genomic Prediction for Polysomic Tetraploid Potato Offspring Grown at High Nordic Latitudes. Genes. 2023; 14(6):1302. https://doi.org/10.3390/genes14061302
Chicago/Turabian StyleOrtiz, Rodomiro, Fredrik Reslow, Ramesh Vetukuri, M. Rosario García-Gil, Paulino Pérez-Rodríguez, and José Crossa. 2023. "Inbreeding Effects on the Performance and Genomic Prediction for Polysomic Tetraploid Potato Offspring Grown at High Nordic Latitudes" Genes 14, no. 6: 1302. https://doi.org/10.3390/genes14061302