
Domestication over Speciation in Allopolyploid Cotton Species: A Stronger Transcriptomic Pull

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, RNA Extraction and Sequencing
2.2. Reference Preparation, Mapping, and Differential Gene Expression Analysis
2.3. Weighted Co-Expression Gene Network Analysis
3. Results
3.1. RNA-Seq Sample Quality and Removal
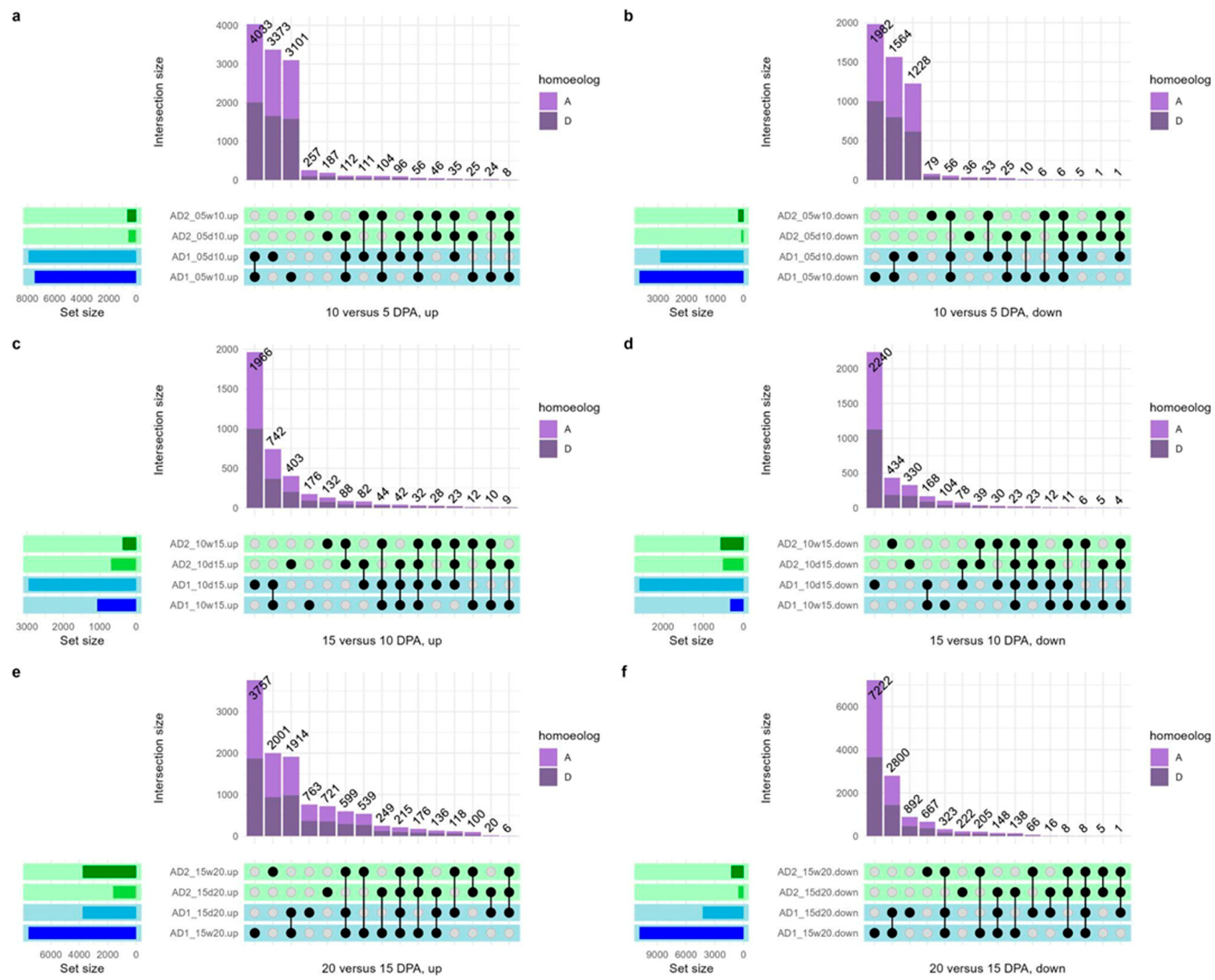
3.2. Differential Expression of Convergent Domesticates and Their Wild Progenitors
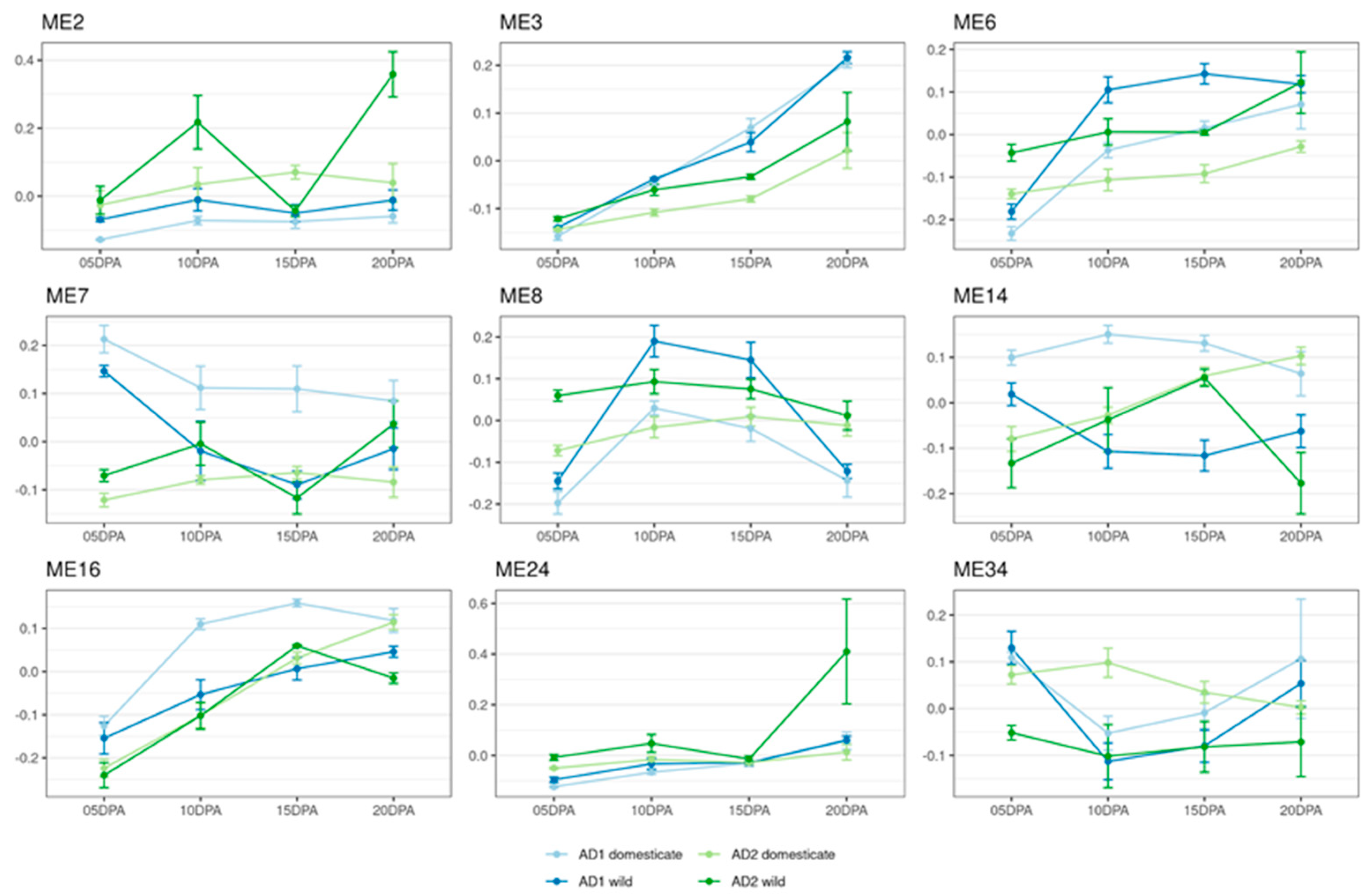
3.3. Meta-Coexpression Network Analysis Fiber Development in the Two Allopolyploid Species
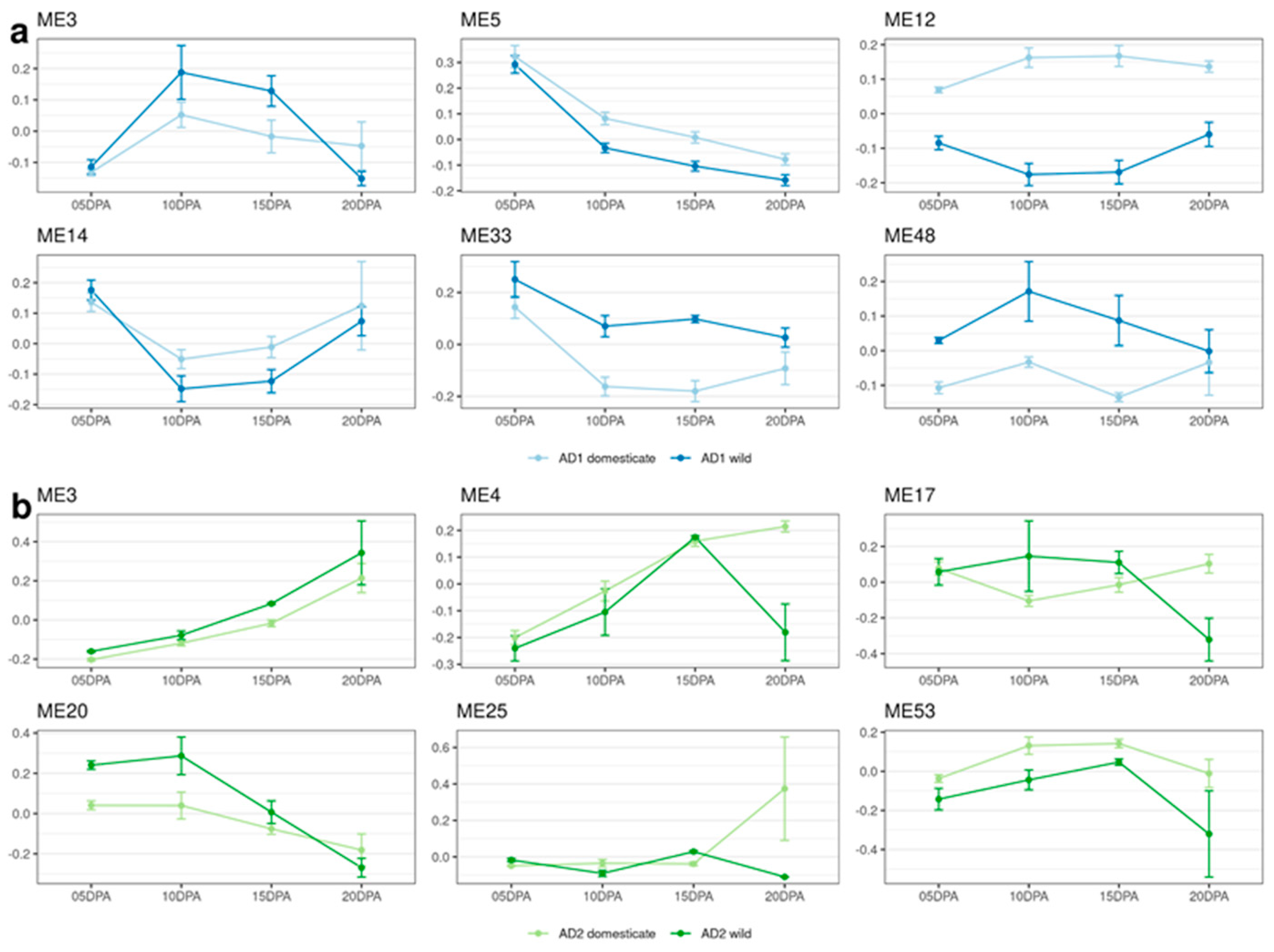
3.4. Comparison between the Separate Species-Networks for G. hirsutum and G. barbadense
3.5. Homoeolog Module Separation
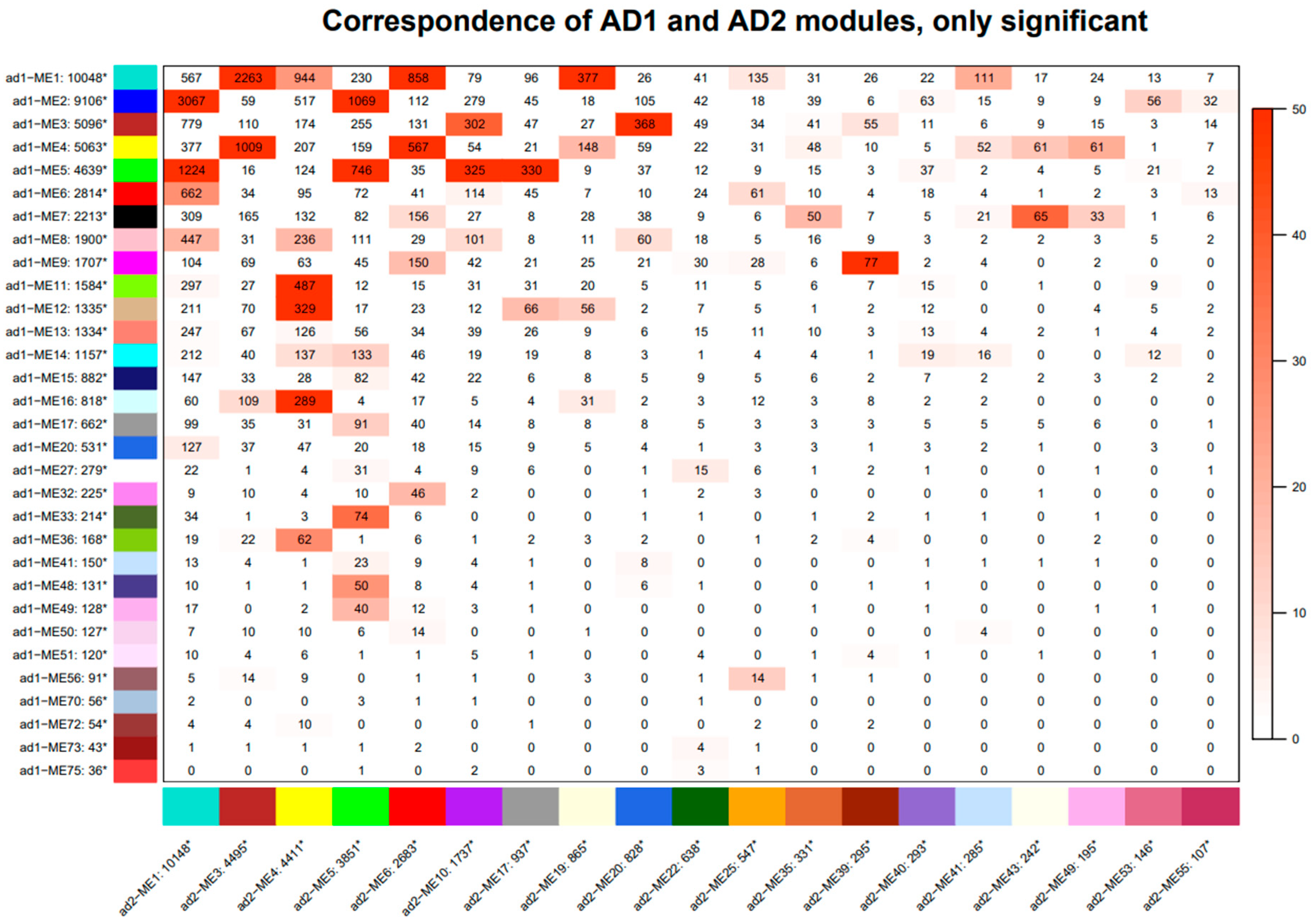
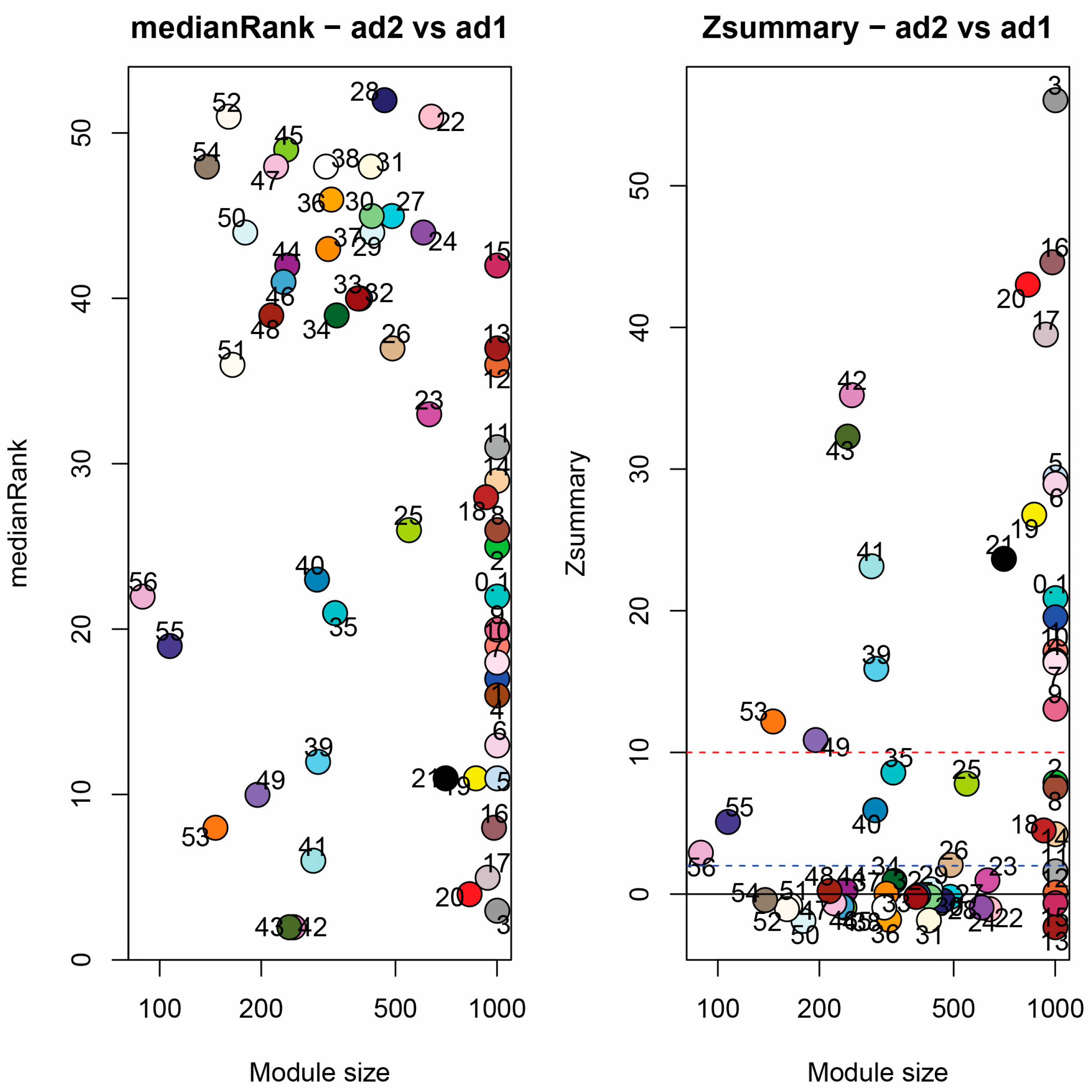
3.6. Module Correspondence and Preservation
4. Discussion
4.1. Independent Domestication Has Uniquely Impacted Two Polyploid Cotton Species
4.2. Comparative Expression Analysis Supports Independent Domestication Mechanisms
4.3. Network Analysis Shows Substantial Differences between G. hirsutum and G. barbadense
4.4. Polyploidy and Homoeolog Responses to Evolutionary Transitions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diamond, J. Evolution, Consequences and Future of Plant and Animal Domestication. Nature 2002, 418, 700–707. [Google Scholar] [CrossRef]
- Meyer, R.S.; Purugganan, M.D. Evolution of Crop Species: Genetics of Domestication and Diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Teletchea, F. Animal Domestication: A Brief Overview. In Animal Domestication; Teletchea, F., Ed.; IntechOpen: London, UK, 2019; ISBN 9781838801748. [Google Scholar]
- Doebley, J.F.; Gaut, B.S.; Smith, B.D. The Molecular Genetics of Crop Domestication. Cell 2006, 127, 1309–1321. [Google Scholar] [CrossRef] [Green Version]
- Purugganan, M.D. Evolutionary Insights into the Nature of Plant Domestication. Curr. Biol. 2019, 29, R705–R714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purugganan, M.D.; Fuller, D.Q. The Nature of Selection during Plant Domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Gross, B.L.; Olsen, K.M. Genetic Perspectives on Crop Domestication. Trends Plant Sci. 2010, 15, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Purugganan, M.D. What Is Domestication? Trends Ecol. Evol. 2022, 37, 663–671. [Google Scholar] [CrossRef]
- Olsen, K.M.; Wendel, J.F. A Bountiful Harvest: Genomic Insights into Crop Domestication Phenotypes. Annu. Rev. Plant Biol. 2013, 64, 47–70. [Google Scholar] [CrossRef] [Green Version]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and Processes in Crop Domestication: An Historical Review and Quantitative Analysis of 203 Global Food Crops. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef]
- Sakuma, S.; Salomon, B.; Komatsuda, T. The Domestication Syndrome Genes Responsible for the Major Changes in Plant Form in the Triticeae Crops. Plant Cell Physiol. 2011, 52, 738–749. [Google Scholar] [CrossRef] [Green Version]
- Konishi, S.; Izawa, T.; Lin, S.Y.; Ebana, K.; Fukuta, Y.; Sasaki, T.; Yano, M. An SNP Caused Loss of Seed Shattering during Rice Domestication. Science 2006, 312, 1392–1396. [Google Scholar] [CrossRef] [Green Version]
- Paran, I.; van der Knaap, E. Genetic and Molecular Regulation of Fruit and Plant Domestication Traits in Tomato and Pepper. J. Exp. Bot. 2007, 58, 3841–3852. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Huang, W.; Gao, J.-P.; Yang, J.; Shi, M.; Zhu, M.-Z.; Luo, D.; Lin, H.-X. Genetic Control of Rice Plant Architecture under Domestication. Nat. Genet. 2008, 40, 1365–1369. [Google Scholar] [CrossRef]
- Swanson-Wagner, R.; Briskine, R.; Schaefer, R.; Hufford, M.B.; Ross-Ibarra, J.; Myers, C.L.; Tiffin, P.; Springer, N.M. Reshaping of the Maize Transcriptome by Domestication. Proc. Natl. Acad. Sci. USA 2012, 109, 11878–11883. [Google Scholar] [CrossRef] [Green Version]
- Moyers, B.T.; Morrell, P.L.; McKay, J.K. Genetic Costs of Domestication and Improvement. J. Hered. 2018, 109, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Constable, G.; Llewellyn, D.; Walford, S.A.; Clement, J.D. Cotton Breeding for Fiber Quality Improvement. In Industrial Crops: Breeding for Bioenergy and Bioproducts; Cruz, V.M.V., Dierig, D.A., Eds.; Springer: New York, NY, USA, 2015; pp. 191–232. ISBN 9781493914470. [Google Scholar]
- Yuan, D.; Grover, C.E.; Hu, G.; Pan, M.; Miller, E.R.; Conover, J.L.; Hunt, S.P.; Udall, J.A.; Wendel, J.F. Parallel and Intertwining Threads of Domestication in Allopolyploid Cotton. Adv. Sci. 2021, 8, 2003634. [Google Scholar] [CrossRef] [PubMed]
- Westengen, O.T.; Huamán, Z.; Heun, M. Genetic Diversity and Geographic Pattern in Early South American Cotton Domestication. Theor. Appl. Genet. 2005, 110, 392–402. [Google Scholar] [CrossRef]
- Percy, R.G.; Wendel, J.F. Allozyme Evidence for the Origin and Diversification of Gossypium barbadense L. Theor. Appl. Genet. 1990, 79, 529–542. [Google Scholar] [CrossRef] [Green Version]
- Viot, C.R.; Wendel, J.F. Evolution of the Cotton Genus, Gossypium, and Its Domestication in the Americas. CRC Crit. Rev. Plant Sci. 2023, 42, 1–33. [Google Scholar] [CrossRef]
- Wendel, J.F.; Brubaker, C.L.; Percival, A.E. Genetic Diversity in Gossypium hirsutum and the Origin of Upland Cotton. Am. J. Bot. 1992, 79, 1291–1310. [Google Scholar] [CrossRef] [Green Version]
- Brubaker, C.L.; Koontz, J.A.; Wendel, J.F. Bidirectional Cytoplasmic and Nuclear Introgression in the New World Cottons Gossypium barbadense and G. hirsutum (Malvaceae). Am. J. Bot. 1993, 80, 1203–1208. [Google Scholar] [CrossRef]
- Zhang, J.; Percy, R.G.; McCarty, J.C. Introgression Genetics and Breeding between Upland and Pima Cotton: A Review. Euphytica 2014, 198, 1–12. [Google Scholar] [CrossRef]
- Lu, Q.; Shi, Y.; Xiao, X.; Li, P.; Gong, J.; Gong, W.; Huang, J. Transcriptome Analysis Suggests That Chromosome Introgression Fragments from Sea Island Cotton (Gossypium barbadense) Increase Fiber Strength in Upland Cotton. G3 Genes 2017, 7, 3469–3479. [Google Scholar]
- Grover, C.E.; Yoo, M.-J.; Lin, M.; Murphy, M.D.; Harker, D.B.; Byers, R.L.; Lipka, A.E.; Hu, G.; Yuan, D.; Conover, J.L.; et al. Genetic Analysis of the Transition from Wild to Domesticated Cotton (Gossypium hirsutum L.). G3 Genes Genomes Genet. 2020, 10, 731–754. [Google Scholar]
- Gallagher, J.P.; Grover, C.E.; Hu, G.; Jareczek, J.J.; Wendel, J.F. Conservation and Divergence in Duplicated Fiber Coexpression Networks Accompanying Domestication of the Polyploid Gossypium hirsutum L. G3 Genes Genomes Genet. 2020, 10, 2879–2892. [Google Scholar] [CrossRef]
- Chen, X.; Guo, W.; Liu, B.; Zhang, Y.; Song, X.; Cheng, Y.; Zhang, L.; Zhang, T. Molecular Mechanisms of Fiber Differential Development between G. barbadense and G. hirsutum Revealed by Genetical Genomics. PLoS ONE 2012, 7, e30056. [Google Scholar] [CrossRef] [Green Version]
- Mei, H.; Qi, B.; Han, Z.; Zhao, T.; Guo, M.; Han, J.; Zhang, J.; Guan, X.; Hu, Y.; Zhang, T.; et al. Subgenome Bias and Temporal Postponement of Gene Expression Contributes to the Distinctions of Fiber Quality in Gossypium Species. Front. Plant Sci. 2021, 12, 819679. [Google Scholar] [CrossRef]
- Jiao, Y.; Long, Y.; Xu, K.; Zhao, F.; Zhao, J.; Li, S.; Geng, S.; Gao, W.; Sun, P.; Deng, X.; et al. Weighted Gene Co-Expression Network Analysis Reveals Hub Genes for Fuzz Development in Gossypium hirsutum. Genes 2023, 14, 208. [Google Scholar] [CrossRef]
- Zou, X.; Liu, A.; Zhang, Z.; Ge, Q.; Fan, S.; Gong, W.; Li, J.; Gong, J.; Shi, Y.; Tian, B.; et al. Co-Expression Network Analysis and Hub Gene Selection for High-Quality Fiber in Upland Cotton (Gossypium hirsutum) Using RNA Sequencing Analysis. Genes 2019, 10, 119. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Chen, Q.; Chen, Q.; Zheng, K.; Cai, Y.; Long, Y.; Zhao, J.; Guo, Y.; Sun, F.; Qu, Y. Analysis of Transcriptome Data and Quantitative Trait Loci Enables the Identification of Candidate Genes Responsible for Fiber Strength in Gossypium barbadense. G3 2022, 12, jkac167. [Google Scholar] [CrossRef]
- Tu, L.-L.; Zhang, X.-L.; Liang, S.-G.; Liu, D.-Q.; Zhu, L.-F.; Zeng, F.-C.; Nie, Y.-C.; Guo, X.-P.; Deng, F.-L.; Tan, J.-F.; et al. Genes Expression Analyses of Sea-Island Cotton (Gossypium barbadense L.) during Fiber Development. Plant Cell Rep. 2007, 26, 1309–1320. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Sun, H.; Hao, P.; Wang, H.; Wang, C.; Ma, L.; Wei, H.; Yu, S. Transcriptome Analysis Reveals Differences in the Mechanisms of Fiber Initiation and Elongation between Long- and Short-Fiber Cotton (Gossypium hirsutum L.) Lines. BMC Genom. 2019, 20, 633. [Google Scholar] [CrossRef] [Green Version]
- Li, P.-T.; Wang, M.; Lu, Q.-W.; Ge, Q.; Rashid, M.H.O.; Liu, A.-Y.; Gong, J.-W.; Shang, H.-H.; Gong, W.-K.; Li, J.-W.; et al. Comparative Transcriptome Analysis of Cotton Fiber Development of Upland Cotton (Gossypium hirsutum) and Chromosome Segment Substitution Lines from G. hirsutum × G. barbadense. BMC Genom. 2017, 18, 705. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wu, M.; Liu, G.; Pei, W.; Zhai, H.; Yu, J.; Zhang, J.; Yu, S. Identification of Candidate Genes for Fiber Length Quantitative Trait Loci through RNA-Seq and Linkage and Physical Mapping in Cotton. BMC Genom. 2017, 18, 427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naoumkina, M.; Thyssen, G.N.; Fang, D.D. RNA-Seq Analysis of Short Fiber Mutants Ligon-Lintless-1 (Li1) and -2 (Li2) Revealed Important Role of Aquaporins in Cotton (Gossypium hirsutum L.) Fiber Elongation. BMC Plant Biol. 2015, 15, 65. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, M.K.; Kim, H.J.; Tang, Y.; Naoumkina, M.; Fang, D.D. Comparative Transcriptome Analysis of Short Fiber Mutants Ligon-Lintless 1 and 2 Reveals Common Mechanisms Pertinent to Fiber Elongation in Cotton (Gossypium hirsutum L.). PLoS ONE 2014, 9, e95554. [Google Scholar] [CrossRef]
- Islam, M.S.; Fang, D.D.; Thyssen, G.N.; Delhom, C.D.; Liu, Y.; Kim, H.J. Comparative Fiber Property and Transcriptome Analyses Reveal Key Genes Potentially Related to High Fiber Strength in Cotton (Gossypium hirsutum L.) Line MD52ne. BMC Plant Biol. 2016, 16, 36. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Fan, L.; Li, P.; Zou, X.; Zhang, Z.; Fan, S.; Gong, J.; Yuan, Y.; Shang, H. Co-Expression Network and Comparative Transcriptome Analysis for Fiber Initiation and Elongation Reveal Genetic Differences in Two Lines from Upland Cotton CCRI70 RIL Population. PeerJ 2021, 9, e11812. [Google Scholar] [CrossRef]
- Hovav, R.; Chaudhary, B.; Udall, J.A.; Flagel, L.; Wendel, J.F. Parallel Domestication, Convergent Evolution and Duplicated Gene Recruitment in Allopolyploid Cotton. Genetics 2008, 179, 1725–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamblin, T.; LeGendre, M.; Collette, M.R.; Lee, G.L.; Moody, A.; De Supinski, B.R.; Futral, S. The Spack Package Manager: Bringing Order to HPC Software Chaos. In Proceedings of the SC15: International Conference for High-Performance Computing, Networking, Storage and Analysis, Austin, TX, USA, 15–20 November 2015; pp. 1–12. [Google Scholar]
- Paterson, A.H.; Wendel, J.F.; Gundlach, H.; Guo, H.; Jenkins, J.; Jin, D.; Llewellyn, D.; Showmaker, K.C.; Shu, S.; Udall, J.; et al. Repeated Polyploidization of Gossypium Genomes and the Evolution of Spinnable Cotton Fibres. Nature 2012, 492, 423–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, J.T.; Gingle, A.R.; Udall, J.A. PolyCat: A Resource for Genome Categorization of Sequencing Reads from Allopolyploid Organisms. G3 2013, 3, 517–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Erratum: Near-Optimal Probabilistic RNA-Seq Quantification. Nat. Biotechnol. 2016, 34, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Series B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Krassowski, M.; Arts, M.; Lagger, C.; Max. Krassowski/Complex-Upset: v1.3.5; 2022. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 9783319242774. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef] [Green Version]
- Bache; Wickham. Magrittr: A Forward-Pipe Operator for R; R package Version.
- Dowle; Srinivasan; Gorecki. Chirico Package “Data. Table”; Extension of Data.
- Pantano DEGreport: Report of DEG Analysis; R Package Version, NJ, USA.
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]
- Langfelder, P.; Luo, R.; Oldham, M.C.; Horvath, S. Is My Network Module Preserved and Reproducible? PLoS Comput. Biol. 2011, 7, e1001057. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.; Hovav, R.; Grover, C.E.; Faigenboim-Doron, A.; Kadmon, N.; Page, J.T.; Udall, J.A.; Wendel, J.F. Evolutionary Conservation and Divergence of Gene Coexpression Networks in Gossypium (cotton) Seeds. Genome Biol. Evol. 2016, 8, 3765–3783. [Google Scholar]
- Yip, A.M.; Horvath, S. Gene Network Interconnectedness and the Generalized Topological Overlap Measure. BMC Bioinform. 2007, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexa, A.; Rahnenfuhrer, J. TopGo: Enrichment Analysis for Gene Ontology; 2016. [Google Scholar]
- Lee, C.M.; Kafle, K.; Belias, D.W.; Park, Y.B.; Glick, R.E.; Haigler, C.H.; Kim, S.H. Comprehensive Analysis of Cellulose Content, Crystallinity, and Lateral Packing in Gossypium hirsutum and Gossypium barbadense Cotton Fibers Using Sum Frequency Generation, Infrared and Raman Spectroscopy, and X-ray Diffraction. Cellulose 2015, 22, 971–989. [Google Scholar] [CrossRef]
- Salnikov, V.V.; Grimson, M.J.; Seagull, R.W.; Haigler, C.H. Localization of Sucrose Synthase and Callose in Freeze-Substituted Secondary-Wall-Stage Cotton Fibers. Protoplasma 2003, 221, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-R.J. Cytoskeletal Motors in Arabidopsis. Sixty-One Kinesins and Seventeen Myosins. Plant Physiol. 2004, 136, 3877–3883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustine, R.C.; Vidali, L.; Kleinman, K.P.; Bezanilla, M. Actin Depolymerizing Factor Is Essential for Viability in Plants, and Its Phosphoregulation Is Important for Tip Growth. Plant J. 2008, 54, 863–875. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, D.J. Plant Expansins: Diversity and Interactions with Plant Cell Walls. Curr. Opin. Plant Biol. 2015, 25, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Suarez, C.; Carroll, R.T.; Burke, T.A.; Christensen, J.R.; Bestul, A.J.; Sees, J.A.; James, M.L.; Sirotkin, V.; Kovar, D.R. Profilin Regulates F-Actin Network Homeostasis by Favoring Formin over Arp2/3 Complex. Dev. Cell 2015, 32, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Tuttle, J.R.; Nah, G.; Duke, M.V.; Alexander, D.C.; Guan, X.; Song, Q.; Chen, Z.J.; Scheffler, B.E.; Haigler, C.H. Metabolomic and Transcriptomic Insights into How Cotton Fiber Transitions to Secondary Wall Synthesis, Represses Lignification, and Prolongs Elongation. BMC Genom. 2015, 16, 477. [Google Scholar] [CrossRef] [Green Version]
- Salih, H.; Gong, W.; He, S.; Sun, G.; Sun, J.; Du, X. Genome-Wide Characterization and Expression Analysis of MYB Transcription Factors in Gossypium hirsutum. BMC Genet. 2016, 17, 129. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Huang, G.-Q.; Zou, D.; Yan, J.-Q.; Li, Y.; Hu, S.; Li, X.-B. The Cotton (Gossypium hirsutum) NAC Transcription Factor (FSN1) as a Positive Regulator Participates in Controlling Secondary Cell Wall Biosynthesis and Modification of Fibers. New Phytol. 2018, 217, 625–640. [Google Scholar] [CrossRef] [Green Version]
- McCurdy, D.W.; Kovar, D.R.; Staiger, C.J. Actin and Actin-Binding Proteins in Higher Plants. Protoplasma 2001, 215, 89–104. [Google Scholar] [CrossRef]
- Desprez, T.; Juraniec, M.; Crowell, E.F.; Jouy, H.; Pochylova, Z.; Parcy, F.; Höfte, H.; Gonneau, M.; Vernhettes, S. Organization of Cellulose Synthase Complexes Involved in Primary Cell Wall Synthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 15572–15577. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Talbot, M.; Llewellyn, D.J. Pectin Methylesterase and Pectin Remodelling Differ in the Fibre Walls of Two Gossypium Species with Very Different Fibre Properties. PLoS ONE 2013, 8, e65131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovav, R.; Udall, J.A.; Chaudhary, B.; Hovav, E.; Flagel, L.; Hu, G.; Wendel, J.F. The Evolution of Spinnable Cotton Fiber Entailed Prolonged Development and a Novel Metabolism. PLoS Genet. 2008, 4, e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, B.; Hovav, R.; Flagel, L.; Mittler, R.; Wendel, J.F. Parallel Expression Evolution of Oxidative Stress-Related Genes in Fiber from Wild and Domesticated Diploid and Polyploid Cotton (Gossypium). BMC Genom. 2009, 10, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Magwanga, R.O.; Cai, X.; Zhou, Z.; Wang, X.; Wang, Y.; Zhang, Z.; Jin, D.; Guo, X.; Wei, Y.; et al. Deep Transcriptome Analysis Reveals Reactive Oxygen Species (ROS) Network Evolution, Response to Abiotic Stress, and Regulation of Fiber Development in Cotton. Int. J. Mol. Sci. 2019, 20, 1863. [Google Scholar] [CrossRef] [Green Version]
- Seagull, R.W. Cytoskeletal Involvement in Cotton Fiber Growth and Development. Micron 1993, 24, 643–660. [Google Scholar] [CrossRef]
- Bartolini, F.; Gundersen, G.G. Formins and Microtubules. Biochim. Biophys. Acta 2010, 1803, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; von Mollard, G.F.; Kovaleva, V.; Stevens, T.H.; Raikhel, N.V. The Plant Vesicle-Associated SNARE AtVTI1a Likely Mediates Vesicle Transport from the Trans-Golgi Network to the Prevacuolar Compartment. Mol. Biol. Cell 1999, 10, 2251–2264. [Google Scholar] [CrossRef] [Green Version]
- Meagher, R.B.; Fechheimer, M. The Arabidopsis Cytoskeletal Genome. Arab. Book 2003, 2, e0096. [Google Scholar] [CrossRef] [Green Version]
- Preuss, M.L.; Kovar, D.R.; Lee, Y.-R.J.; Staiger, C.J.; Delmer, D.P.; Liu, B. A Plant-Specific Kinesin Binds to Actin Microfilaments and Interacts with Cortical Microtubules in Cotton Fibers. Plant Physiol. 2004, 136, 3945–3955. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Irani, N.G.; Friml, J. Clathrin-Mediated Endocytosis: The Gateway into Plant Cells. Curr. Opin. Plant Biol. 2011, 14, 674–682. [Google Scholar] [CrossRef]
- Huang, S.; Qu, X.; Zhang, R. Plant Villins: Versatile Actin Regulatory Proteins. J. Integr. Plant Biol. 2015, 57, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, J.; Zhang, L.; Zuo, K. A Cotton Annexin Protein AnxGb6 Regulates Fiber Elongation through Its Interaction with Actin 1. PLoS ONE 2013, 8, e66160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; He, S.; Wang, X.; Sun, J.; Zhang, Y.; Zhang, G.; Wu, L.; Li, Z.; Liu, Z.; Sun, G.; et al. Resequencing a Core Collection of Upland Cotton Identifies Genomic Variation and Loci Influencing Fiber Quality and Yield. Nat. Genet. 2018, 50, 803–813. [Google Scholar] [CrossRef]
- Qin, Y.-M.; Hu, C.-Y.; Zhu, Y.-X. The Ascorbate Peroxidase Regulated by H2O2 and Ethylene Is Involved in Cotton Fiber Cell Elongation by Modulating ROS Homeostasis. Plant Signal. Behav. 2008, 3, 194–196. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Sun, W.; Wang, J.; Zhao, C.; Zuo, K. GhbHLH18 Negatively Regulates Fiber Strength and Length by Enhancing Lignin Biosynthesis in Cotton Fibers. Plant Sci. 2019, 286, 7–16. [Google Scholar] [CrossRef]
- Niu, Q.; Tan, K.; Zang, Z.; Xiao, Z.; Chen, K.; Hu, M.; Luo, M. Modification of Phytosterol Composition Influences Cotton Fiber Cell Elongation and Secondary Cell Wall Deposition. BMC Plant Biol. 2019, 19, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, T.; Yao, H.; Wang, J.; Wang, J.; Xue, H.; Zuo, K. GhLTPG1, a Cotton GPI-Anchored Lipid Transfer Protein, Regulates the Transport of Phosphatidylinositol Monophosphates and Cotton Fiber Elongation. Sci. Rep. 2016, 6, 26829. [Google Scholar] [CrossRef] [Green Version]
- Shan, C.-M.; Shangguan, X.-X.; Zhao, B.; Zhang, X.-F.; Chao, L.-M.; Yang, C.-Q.; Wang, L.-J.; Zhu, H.-Y.; Zeng, Y.-D.; Guo, W.-Z.; et al. Control of Cotton Fibre Elongation by a Homeodomain Transcription Factor GhHOX3. Nat. Commun. 2014, 5, 5519. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.-X.; Chen, Y.; Zeng, W.; Li, Y.; Gao, L.; Li, D.-D.; Bacic, A.; Xu, W.-L.; Li, X.-B. The Cotton β-Galactosyltransferase 1 (GalT1) That Galactosylates Arabinogalactan Proteins Participates in Controlling Fiber Development. Plant J. 2017, 89, 957–971. [Google Scholar] [CrossRef] [Green Version]
- Thyssen, G.N.; Fang, D.D.; Turley, R.B.; Florane, C.B.; Li, P.; Mattison, C.P.; Naoumkina, M. A Gly65Val Substitution in an Actin, GhACT_LI1, Disrupts Cell Polarity and F-Actin Organization Resulting in Dwarf, Lintless Cotton Plants. Plant J. 2017, 90, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Burns, T.H.; Light, G.; Sun, Y.; Fokar, M.; Kasukabe, Y.; Fujisawa, K.; Maekawa, Y.; Allen, R.D. Xyloglucan Endotransglycosylase/hydrolase Genes in Cotton and Their Role in Fiber Elongation. Planta 2010, 232, 1191–1205. [Google Scholar] [CrossRef]
- Maris, A.; Kaewthai, N.; Eklöf, J.M.; Miller, J.G.; Brumer, H.; Fry, S.C.; Verbelen, J.-P.; Vissenberg, K. Differences in Enzymic Properties of Five Recombinant Xyloglucan Endotransglucosylase/hydrolase (XTH) Proteins of Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Madson, M.; Dunand, C.; Li, X.; Verma, R.; Vanzin, G.F.; Caplan, J.; Shoue, D.A.; Carpita, N.C.; Reiter, W.-D. The MUR3 Gene of Arabidopsis Encodes a Xyloglucan Galactosyltransferase That Is Evolutionarily Related to Animal Exostosins. Plant Cell 2003, 15, 1662–1670. [Google Scholar] [CrossRef] [Green Version]
- Amos, R.A.; Mohnen, D. Critical Review of Plant Cell Wall Matrix Polysaccharide Glycosyltransferase Activities Verified by Heterologous Protein Expression. Front. Plant Sci. 2019, 10, 915. [Google Scholar] [CrossRef] [Green Version]
- Lv, F.; Wang, H.; Wang, X.; Han, L.; Ma, Y.; Wang, S.; Feng, Z.; Niu, X.; Cai, C.; Kong, Z.; et al. GhCFE1A, a Dynamic Linker between the ER Network and Actin Cytoskeleton, Plays an Important Role in Cotton Fibre Cell Initiation and Elongation. J. Exp. Bot. 2015, 66, 1877–1889. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.; Grover, C.E.; Yuan, D.; Dong, Y.; Miller, E.; Conover, J.L.; Wendel, J.F. Evolution and Diversity of the Cotton Genome. In Cotton Precision Breeding; Rahman, M.-U., Zafar, Y., Zhang, T., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 25–78. ISBN 9783030645045. [Google Scholar]
- Wendel, J.F.; Grover, C.E. Taxonomy and Evolution of the Cotton Genus, Gossypium. In Cotton; Agronomy Monograph; American Society of Agronomy, Inc.: Madison, WI, USA; Crop Science Society of America, Inc.: Madison, WI, USA; Soil Science Society of America, Inc.: Madison, WI, USA, 2015; pp. 25–44. ISBN 9780891186267. [Google Scholar]
- Gallagher, J.P.; Grover, C.E.; Rex, K.; Moran, M.; Wendel, J.F. A New Species of Cotton from Wake Atoll, Gossypium Stephensii (Malvaceae). Syst. Bot. 2017, 42, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Grover, C.E.; Zhu, X.; Grupp, K.K.; Jareczek, J.J.; Gallagher, J.P.; Szadkowski, E.; Seijo, J.G.; Wendel, J.F. Molecular Confirmation of Species Status for the Allopolyploid Cotton Species, Gossypium Ekmanianum Wittmack. Genet. Resour. Crop Evol. 2015, 62, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Brubaker, C.L.; Wendel, J.F. Reevaluating the Origin of Domesticated Cotton (Gossypium hirsutum; Malvaceae) Using Nuclear Restriction Fragment Length Polymorphisms (RFLPs). Am. J. Bot. 1994, 81, 1309–1326. [Google Scholar] [CrossRef]
- Kohel, R.J.; Richmond, T.R.; Lewis, C.F. Genetics of Flowering Response in Cotton. VI. Flowering Behavior of Gossypium hirsutum L. and G. barbadense L. Hybrids. Crop. Sci. 1974, 14, 696–699. [Google Scholar] [CrossRef]
- Lewis, C.F.; Richmond, T.R. The Genetics of Flowering Response in Cotton. II. Inheritance of Flowering Response in a Gossypium barbadense Cross. Genetics 1960, 45, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Bitocchi, E.; Rau, D.; Bellucci, E.; Rodriguez, M.; Murgia, M.L.; Gioia, T.; Santo, D.; Nanni, L.; Attene, G.; Papa, R. Beans (Phaseolus Ssp.) as a Model for Understanding Crop Evolution. Front. Plant Sci. 2017, 8, 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, B.P.; Haigler, C.H. Microtubules Exert Early, Partial, and Variable Control of Cotton Fiber Diameter. Planta 2021, 253, 47. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Wright, R.J.; El-Zik, K.M.; Paterson, A.H. Polyploid Formation Created Unique Avenues for Response to Selection in Gossypium (cotton). Proc. Natl. Acad. Sci. USA 1998, 95, 4419–4424. [Google Scholar] [CrossRef] [Green Version]
- Lacape, J.-M.; Nguyen, T.-B.; Courtois, B.; Belot, J.-L.; Giband, M.; Gourlot, J.-P.; Gawryziak, G.; Roques, S.; Hau, B. QTL Analysis of Cotton Fiber Quality Using Multiple Gossypium hirsutum Gossypium barbadense Backcross Generations. Crop Sci. 2005, 45, 123–140. [Google Scholar] [CrossRef]
- Said, J.I.; Song, M.; Wang, H.; Lin, Z.; Zhang, X.; Fang, D.D.; Zhang, J. A Comparative Meta-Analysis of QTL between Intraspecific Gossypium hirsutum and Interspecific G. hirsutum × G. barbadense Populations. Mol. Genet. Genom. 2015, 290, 1003–1025. [Google Scholar] [CrossRef]
- Yoo, M.-J.; Szadkowski, E.; Wendel, J.F. Homoeolog Expression Bias and Expression Level Dominance in Allopolyploid Cotton. Heredity 2013, 110, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, B.; Flagel, L.; Stupar, R.M.; Udall, J.A.; Verma, N.; Springer, N.M.; Wendel, J.F. Reciprocal Silencing, Transcriptional Bias and Functional Divergence of Homeologs in Polyploid Cotton (Gossypium). Genetics 2009, 182, 503–517. [Google Scholar] [CrossRef] [Green Version]
- Flagel, L.; Udall, J.; Nettleton, D.; Wendel, J. Duplicate Gene Expression in Allopolyploid Gossypium Reveals Two Temporally Distinct Phases of Expression Evolution. BMC Biol. 2008, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Hu, G.; Grover, C.E.; Conover, J.; Yuan, D.; Wendel, J.F. Unraveling Cis and Trans Regulatory Evolution during Cotton Domestication. Nat. Commun. 2019, 10, 5399. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Accession | Domestication Status |
|---|---|---|
| Gossypium hirsutum | CRB252 | domesticated |
| Maxxa | domesticated | |
| TM1 | domesticated | |
| TX665 | wild | |
| TX2094 | wild | |
| TX2095 | wild | |
| Gossypium barbadense | Pima S6 | domesticated |
| Pima S7 | domesticated | |
| Phy76 | domesticated | |
| GB0303 | wild | |
| GPS52 | wild | |
| K101 | wild |
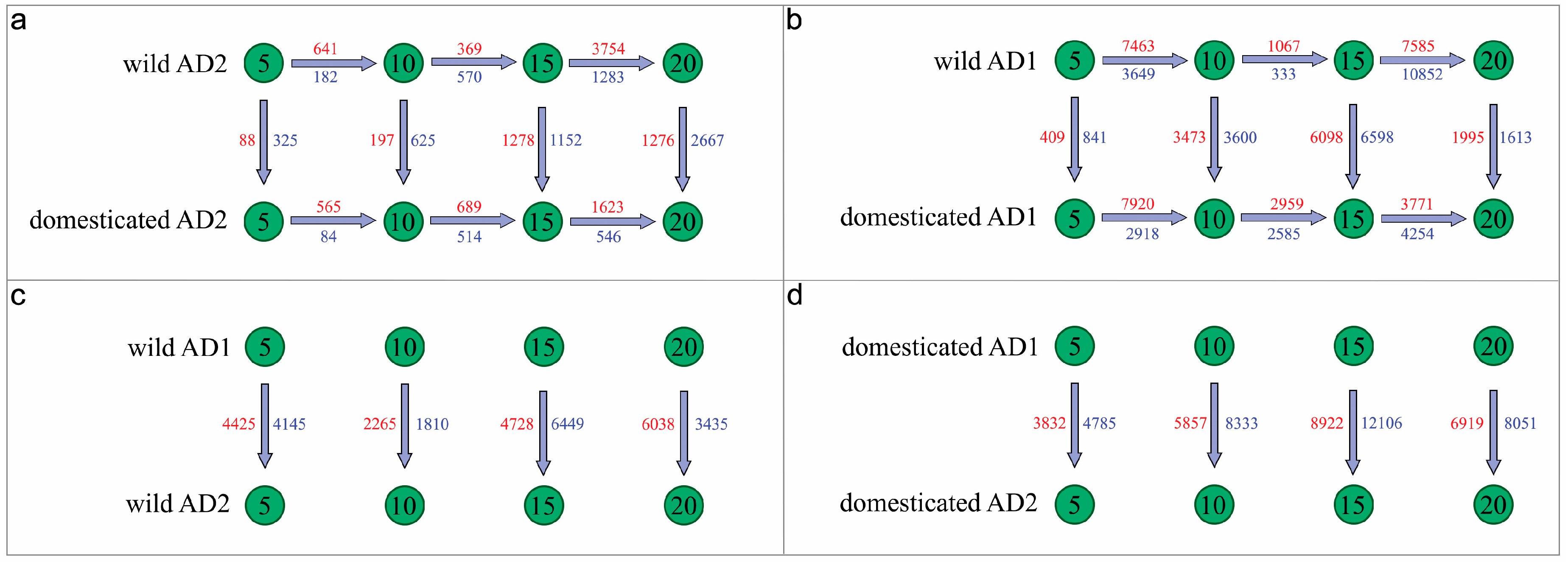
| Material | DPA | G. barbadense | G. hirsutum | Interspecies | DPA |
|---|---|---|---|---|---|
| wild | 10 vs. 05 | 823 (641:182) | 11,112 (7463:3649) | 8570 (4425:4145) | 5 |
| 15 vs. 10 | 939 (369:570) | 1400 (1067:333) | 4075 (2265:1810) | 10 | |
| 20 vs. 15 | 5037 (3754:1283) | 18,437 (7585:10852) | 11,177 (4728:6449) | 15 | |
| 9473 (6038:3435) | 20 | ||||
| domesticated | 10 vs. 05 | 649 (565:84) | 10,838 (7920:2918) | 8617 (3832:4785) | 5 |
| 15 vs. 10 | 1203 (689:514) | 5544 (2959:2585) | 14,190 (5857:8333) | 10 | |
| 20 vs. 15 | 2169 (1623:546) | 8025 (3771:4254) | 21,028 (8922:12106) | 15 | |
| 14,970 (6919:8051) | 20 | ||||
| wild versus domesticated | 5 | 413 (88:325) | 1250 (409:841) | ||
| 10 | 822 (197:625) | 7073 (3473:3600) | |||
| 15 | 2430 (1278:1152) | 12,696 (6098:6598) | |||
| 20 | 3943 (1276:2667) | 3608 (1995:1613) |
| Meta | AD1 | AD2 | AD1-AD2 Consensus | |
|---|---|---|---|---|
| Total module genes | 63,675 | 62,084 | 61,934 | 57,019 |
| Solo A-homoeolog | 19,976 (31.4%) | 19,857 (32.0%) | 23,726 (38.3%) | 22,171 (38.9%) |
| Solo D-homoeolog | 19,945 (31.3%) | 19,834 (31.9%) | 23,668 (38.2%) | 22,210 (39.0%) |
| Homoeolog pairs | 11,877 (37.3%) | 11,198 (36.1%) | 7270 (23.5%) | 6319 (22.2%) |
| Total modules | 58 | 76 | 56 | 165 |
| A-dominant module | 7 | 3 | 5 | 5 |
| D-dominant module | 5 | 1 | 5 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jareczek, J.J.; Grover, C.E.; Hu, G.; Xiong, X.; Arick II, M.A.; Peterson, D.G.; Wendel, J.F. Domestication over Speciation in Allopolyploid Cotton Species: A Stronger Transcriptomic Pull. Genes 2023, 14, 1301. https://doi.org/10.3390/genes14061301
Jareczek JJ, Grover CE, Hu G, Xiong X, Arick II MA, Peterson DG, Wendel JF. Domestication over Speciation in Allopolyploid Cotton Species: A Stronger Transcriptomic Pull. Genes. 2023; 14(6):1301. https://doi.org/10.3390/genes14061301
Chicago/Turabian StyleJareczek, Josef J., Corrinne E. Grover, Guanjing Hu, Xianpeng Xiong, Mark A. Arick II, Daniel G. Peterson, and Jonathan F. Wendel. 2023. "Domestication over Speciation in Allopolyploid Cotton Species: A Stronger Transcriptomic Pull" Genes 14, no. 6: 1301. https://doi.org/10.3390/genes14061301