
Forensic Analysis and Genetic Structure Construction of Chinese Chongming Island Han Based on Y Chromosome STRs and SNPs

Abstract
:1. Introduction
2. Materials and Methods
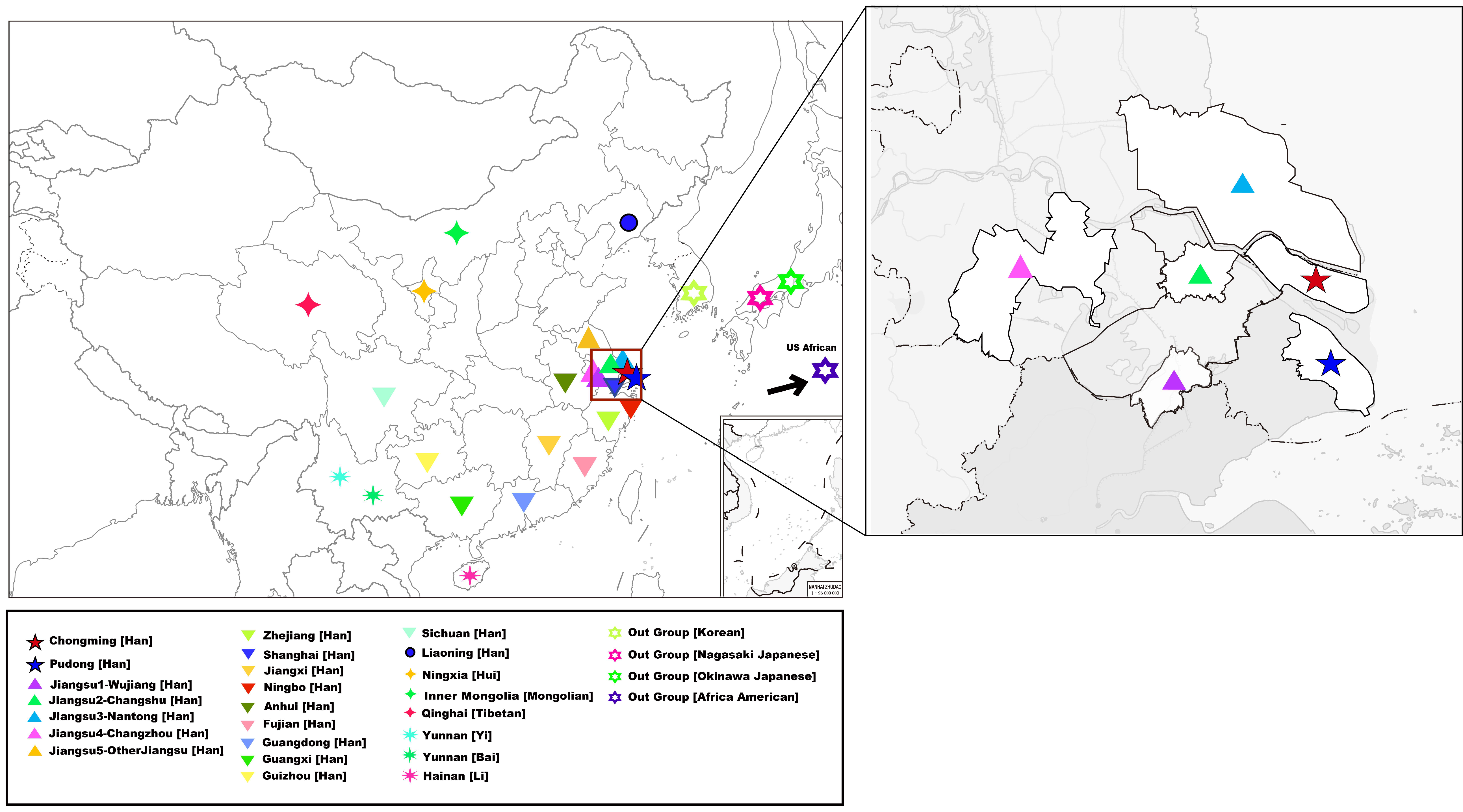
2.1. DNA Sample Preparation
2.2. DNA Typing of 38 Y-STR Locus and 24 Y-SNP Markers
2.3. Data Analysis
2.3.1. Y-STR Analytic Methods
2.3.2. Y-SNP Analytic Methods
2.3.3. Y-STR and Y-SNP Joint Analytic Methods
3. Results
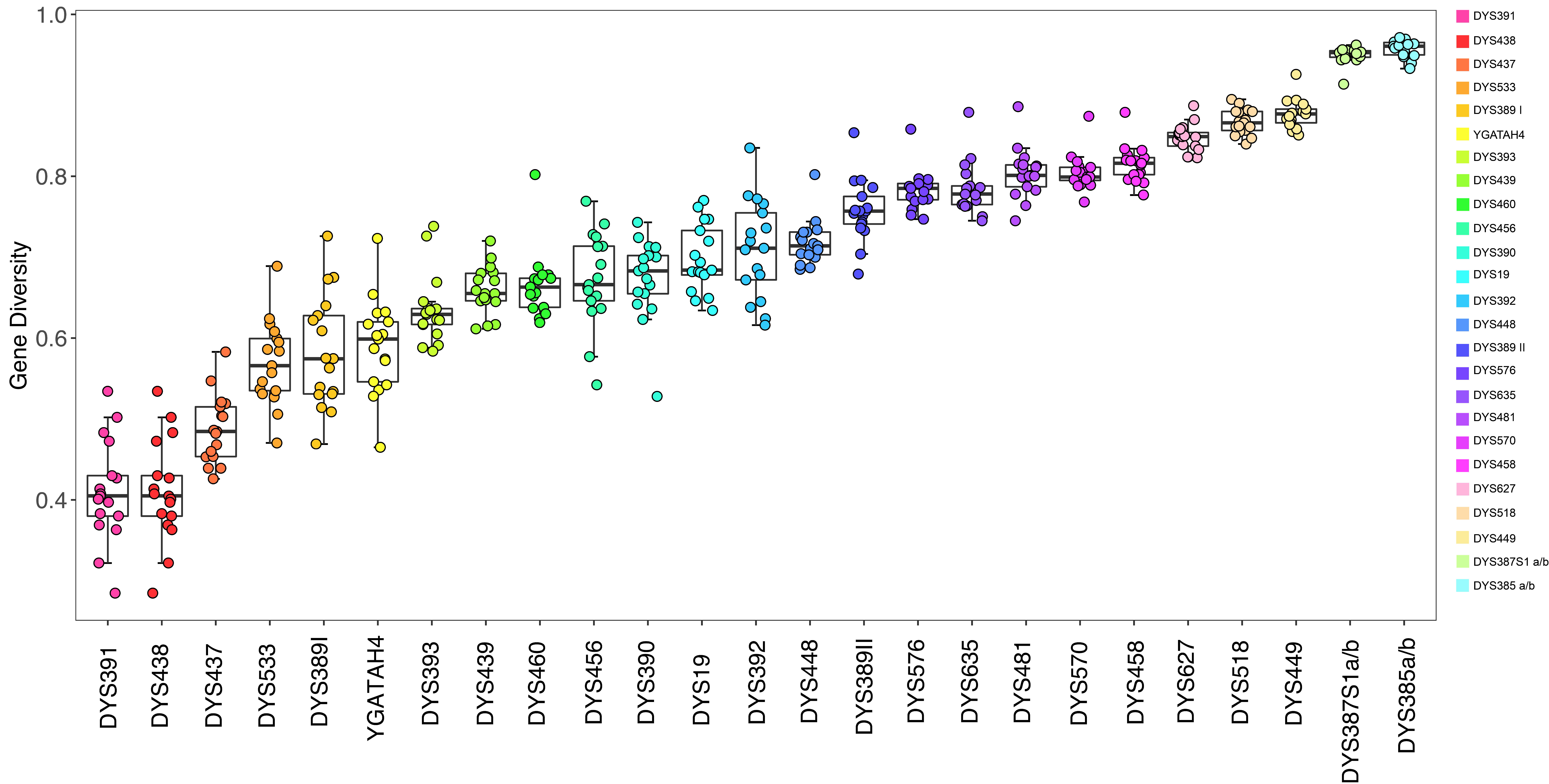
3.1. Genetic Diversity and Haplotype Diversity of Y-STRs
3.1.1. Allelic Diversity Analysis
3.1.2. Haplotype Diversity Analysis
3.1.3. Variation Analysis
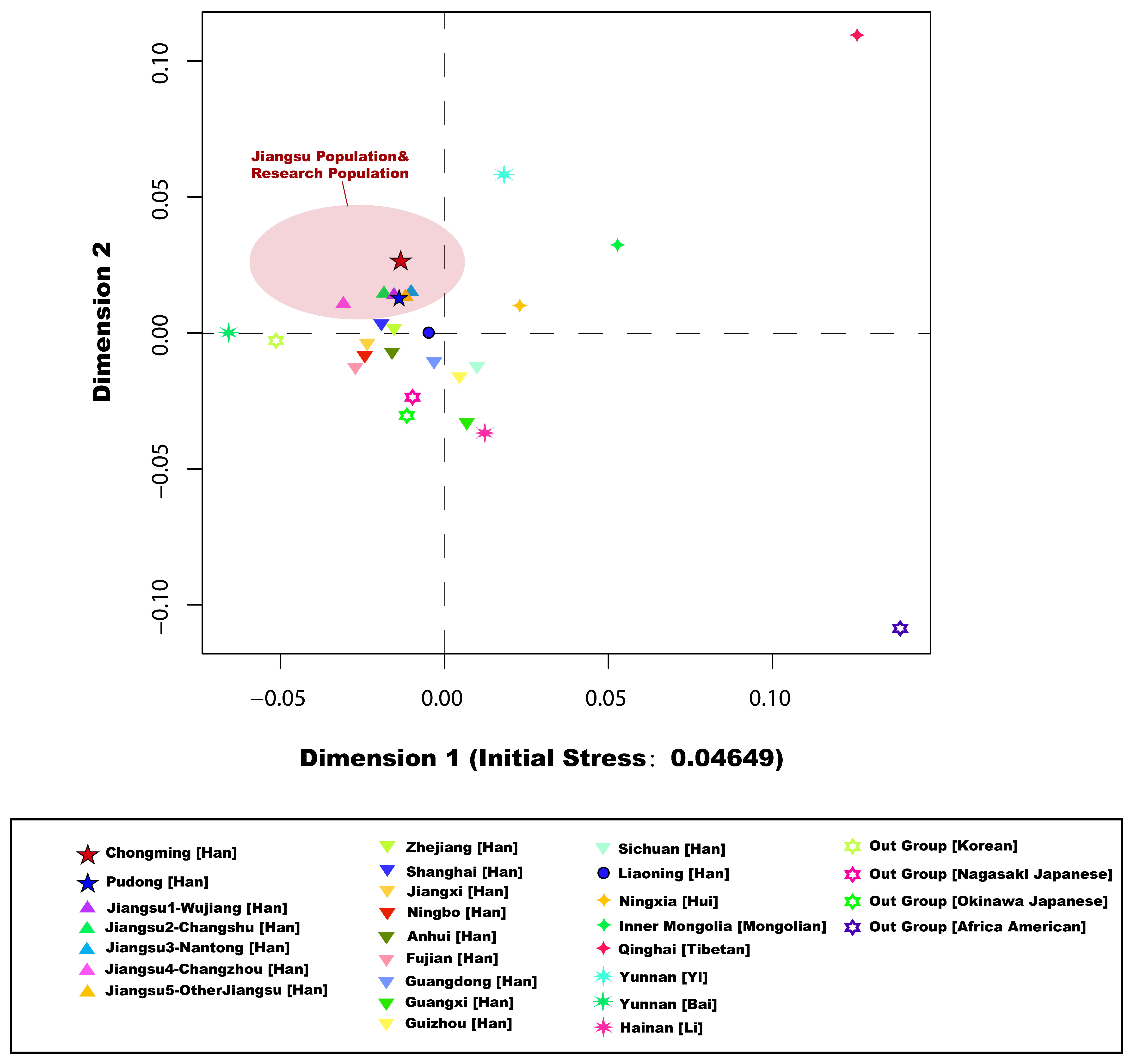
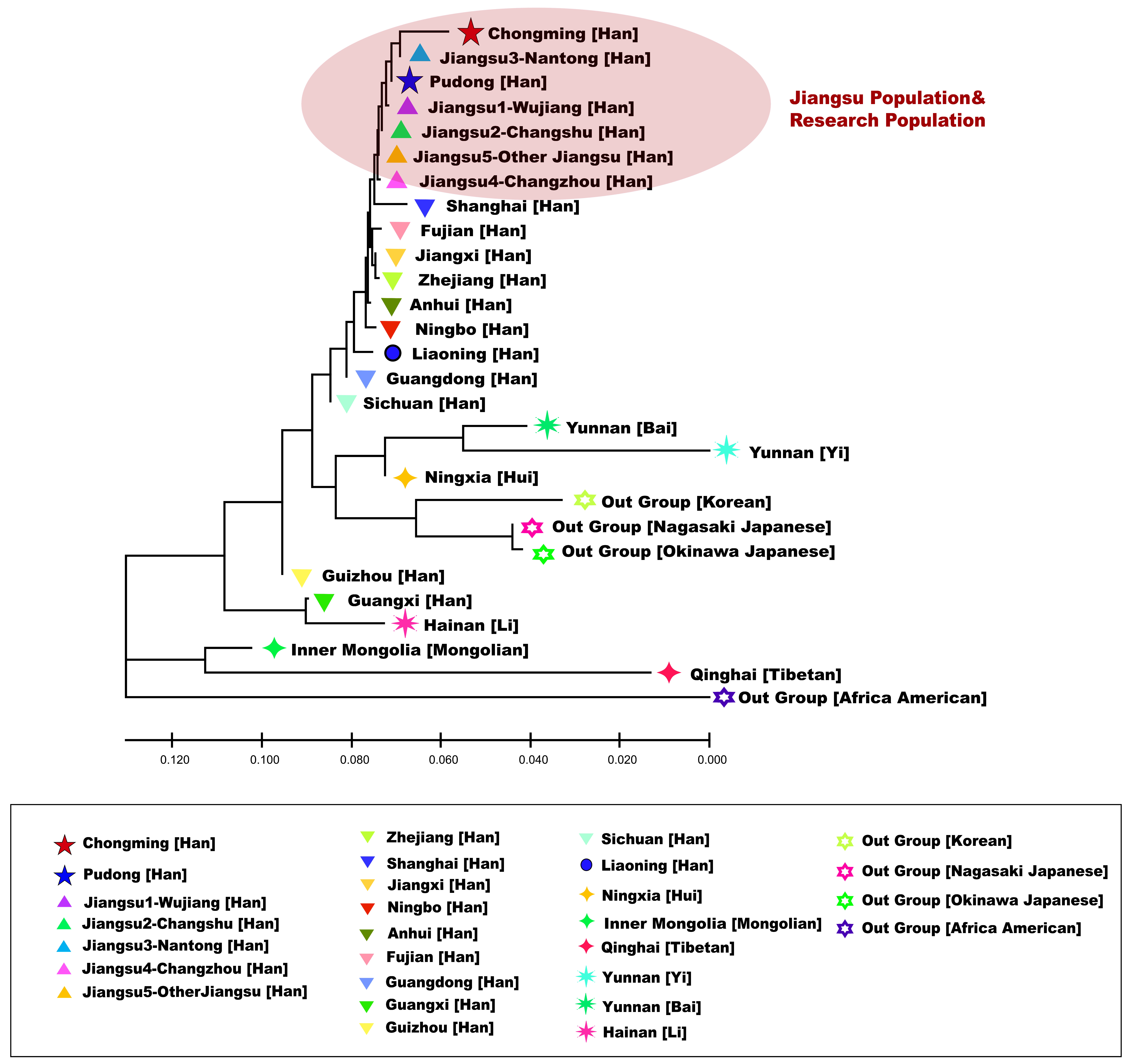
3.2. Genetic Affinity Analysis
3.3. Y-SNP Analysis
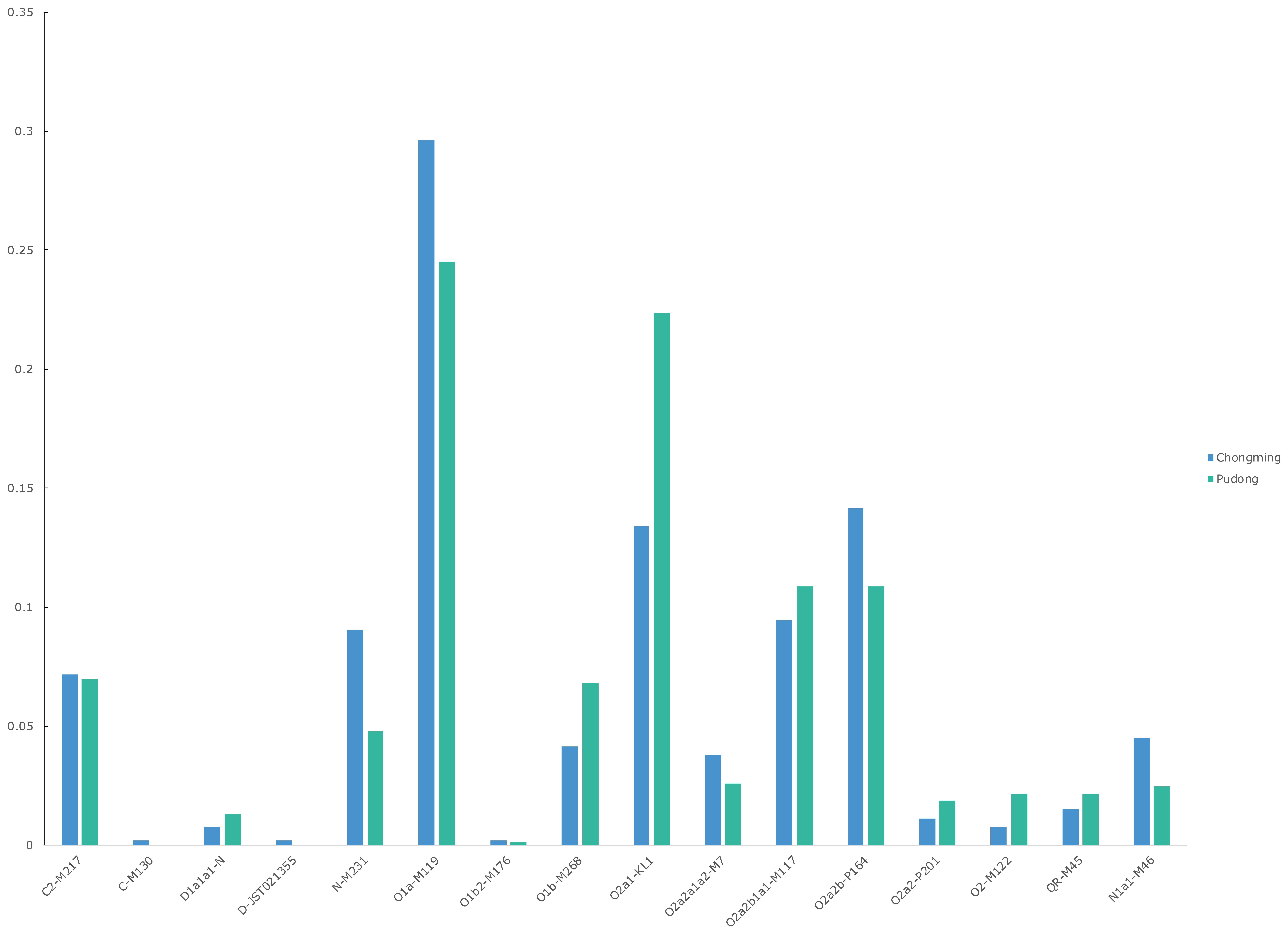
3.3.1. Y-Chromosomal Haplogroup Distribution
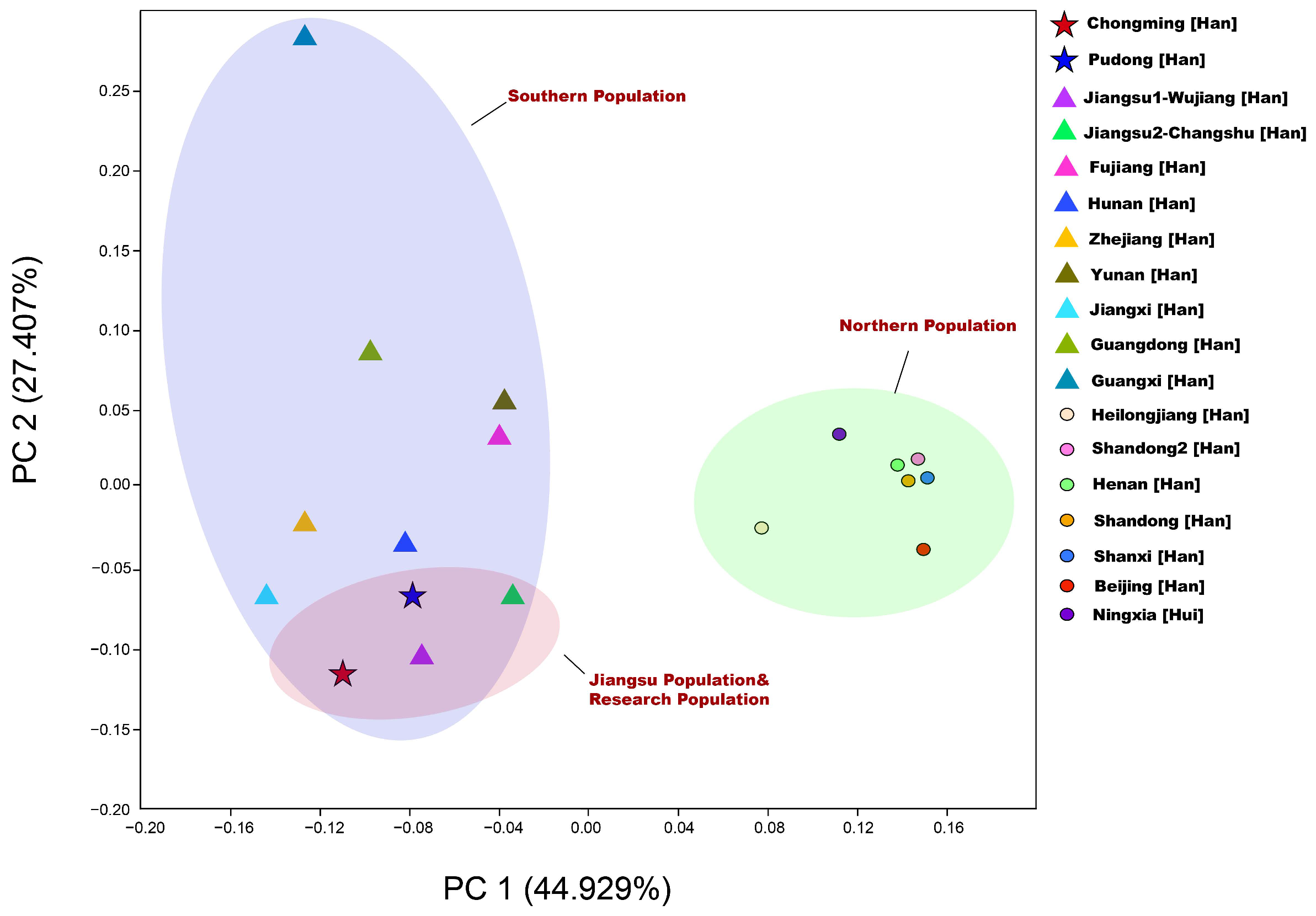
3.3.2. Principal Component Analysis
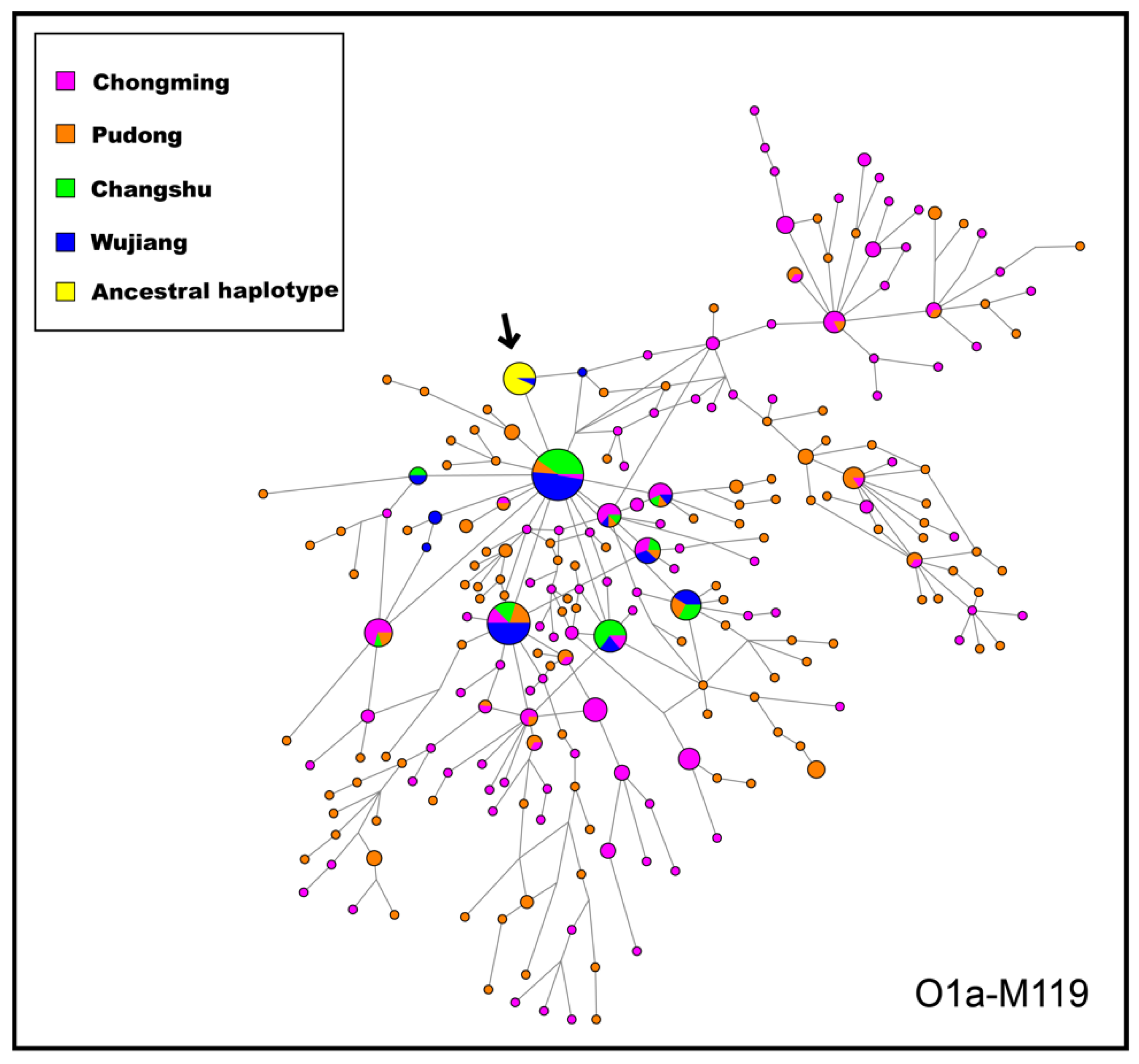
3.4. Network Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jobling, M.A.; Tyler-Smith, C. Human Y-Chromosome Variation in the Genome-Sequencing Era. Nat. Rev. Genet. 2017, 18, 485–497. [Google Scholar] [CrossRef]
- Pinto, N.; Gusmão, L.; Amorim, A. Mutation and Mutation Rates at Y Chromosome Specific Short Tandem Repeat Polymorphisms (STRs): A Reappraisal. Forensic Sci. Int. Genet. 2014, 9, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Wang, Q.; Long, Q.; Ng, B.L.; Swerdlow, H.; Burton, J.; Skuce, C.; Taylor, R.; Abdellah, Z.; Zhao, Y.; et al. Human Y Chromosome Base-Substitution Mutation Rate Measured by Direct Sequencing in a Deep-Rooting Pedigree. Curr. Biol. 2009, 19, 1453–1457. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Kubo, S. Y-SNP and Y-STR Analysis in a Japanese Population. Leg. Med. 2008, 10, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Cortellini, V.; Verzeletti, A.; Cerri, N.; Marino, A.; De Ferrari, F. Y-Chromosome Polymorphisms and Ethnic Group—A Combined STR and SNP Approach in a Population Sample from Northern Italy. Croat. Med. J. 2013, 54, 279–285. [Google Scholar] [CrossRef]
- Zhou, Z. Chongming County Annals; Shanghai People’s Publishing House: Shanghai, China, 1989. [Google Scholar]
- Nicogossian, A.; Kloiber, O.; Stabile, B. The Revised World Medical Association’s Declaration of Helsinki 2013: Enhancing the Protection of Human Research Subjects and Empowering Ethics Review Committees. World Med. Health Policy 2014, 6, 1–3. [Google Scholar] [CrossRef]
- Song, M.; Song, F.; Wang, S.; Hou, Y. Developmental Validation of the Yfiler Platinum PCR Amplification Kit for Forensic Genetic Caseworks and Databases. Electrophoresis 2021, 42, 126–133. [Google Scholar] [CrossRef]
- Yin, C.; Ren, Y.; Adnan, A.; Tian, J.; Guo, K.; Xia, M.; He, Z.; Zhai, D.; Chen, X.; Wang, L.; et al. Title: Developmental Validation of Y-SNP Pedigree Tagging System: A Panel via Quick ARMS PCR. Forensic Sci. Int. Genet. 2020, 46, 102271. [Google Scholar] [CrossRef]
- Willuweit, S.; Roewer, L. The New Y Chromosome Haplotype Reference Database. Forensic Sci. Int. Genet. 2015, 15, 43–48. [Google Scholar] [CrossRef]
- Y Chromosome Consortium. A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups. Genome Res. 2002, 12, 339–348. [Google Scholar] [CrossRef]
- Karafet, T.M.; Mendez, F.L.; Meilerman, M.B.; Underhill, P.A.; Zegura, S.L.; Hammer, M.F. New Binary Polymorphisms Reshape and Increase Resolution of the Human Y Chromosomal Haplogroup Tree. Genome Res. 2008, 18, 830–838. [Google Scholar] [CrossRef] [PubMed]
- van Oven, M.; Van Geystelen, A.; Kayser, M.; Decorte, R.; Larmuseau, M.H. Seeing the Wood for the Trees: A Minimal Reference Phylogeny for the Human Y Chromosome. Hum. Mutat. 2014, 35, 187–191. [Google Scholar] [CrossRef]
- Nei, M. Analysis of Gene Diversity in Subdivided Populations. Proc. Natl. Acad. Sci. USA 1974, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Tajima, F. DNA Polymorphism Detectable by Restriction Endonucleases. Genetics 1981, 97, 145–163. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Chen, T.F.; Zeng, Z.F.; Zhang, Y.W.; Tang, Z.; Su, K.Y.; Fan, C.Y.; Li, S.L. Genetic Structure Analysis of Y-Chromosome STR and SNP in Population of Wujiang Area, Suzhou City. Fa Yi Xue Za Zhi 2019, 35, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Tian, J.Z.; Zhang, Y.W.; Yin, C.Y.; Tang, Z.; Li, S.; Yao, J.; Huang, J.W.; Li, S.L. Genetic Structure Analysis of Y Chromosome STR and SNP in Population of Changshu. Fa Yi Xue Za Zhi 2020, 36, 538–544. [Google Scholar] [CrossRef]
- Zhou, Y.; Shao, C.; Li, L.; Zhang, Y.; Liu, B.; Yang, Q.; Tang, Q.; Li, S.; Xie, J. Genetic Analysis of 29 Y-STR Loci in the Chinese Han Population from Shanghai. Forensic Sci. Int. Genet. 2018, 32, e1–e4. [Google Scholar] [CrossRef]
- Liu, Y.; Jin, H.Y.; Song, D.L.; Wang, C.Z.; Shi, M.S.; Wang, C.C. 27 Y-Chromosomal STR Haplotypic Structure for the Chinese Han Population from Changchun, Northeastern China. Leg. Med. 2021, 48, 101813. [Google Scholar] [CrossRef]
- Lang, M.; Liu, H.; Song, F.; Qiao, X.; Ye, Y.; Ren, H.; Li, J.; Huang, J.; Xie, M.; Chen, S.; et al. Forensic Characteristics and Genetic Analysis of Both 27 Y-STRs and 143 Y-SNPs in Eastern Han Chinese Population. Forensic Sci. Int. Genet. 2019, 42, e13–e20. [Google Scholar] [CrossRef]
- Incerti, D.; Thom, H.; Baio, G.; Jansen, J.P. R You Still Using Excel? The Advantages of Modern Software Tools for Health Technology Assessment. Value Health 2019, 22, 575–579. [Google Scholar] [CrossRef]
- Siegert, S.; Roewer, L.; Nothnagel, M. Shannon’s Equivocation for Forensic Y-STR Marker Selection. Forensic Sci. Int. Genet. 2015, 16, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Mulero, J.J.; Chang, C.W.; Calandro, L.M.; Green, R.L.; Li, Y.; Johnson, C.L.; Hennessy, L.K. Development and Validation of the AmpFlSTR Yfiler PCR Amplification Kit: A Male Specific, Single Amplification 17 Y-STR Multiplex System. J. Forensic Sci. 2006, 51, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.; Zhong, C.; Nguyen, V.; Ge, J.; Lagace, R.E.; Short, M.L.; Mulero, J.J. Developmental Validation of the Yfiler(®) Plus PCR Amplification Kit: An Enhanced Y-STR Multiplex for Casework and Database Applications. Forensic Sci. Int. Genet. 2016, 24, 164–175. [Google Scholar] [CrossRef]
- Thompson, J.M.; Ewing, M.M.; Frank, W.E.; Pogemiller, J.J.; Nolde, C.A.; Koehler, D.J.; Shaffer, A.M.; Rabbach, D.R.; Fulmer, P.M.; Sprecher, C.J.; et al. Developmental Validation of the PowerPlex® Y23 System: A Single Multiplex Y-STR Analysis System for Casework and Database Samples. Forensic Sci. Int. Genet. 2013, 7, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Yang, S.; Yang, M.; Jia, D.; Han, X.; Wang, W.; Jin, L.; Li, L.; Li, S. Analysis of Y-Chromosome Short Tandem Repeat Loci on 1082 Nantong Han Individuals in Eastern China. Forensic Sci. Int. Genet. 2016, 23, e18–e19. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Wang, S.; Zhang, J.; Zhang, J.; Yang, Z.; Zhang, S.; Li, C. Genetic Characterization of 27 Y-STR Loci Analyzed in the Nantong Han Population Residing along the Yangtze Basin. Forensic Sci. Int. Genet. 2019, 39, e10–e13. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Jin, M.; Ji, G.; Zhang, J.; Zhang, J.; Yang, Z.; Chen, C.; Zhang, S.; Li, C. Forensic Characteristics of 36 Y-STR Loci in a Changzhou Han Population and Genetic Distance Analysis among Several Chinese Populations. Forensic Sci. Int. Genet. 2019, 40, e268–e270. [Google Scholar] [CrossRef]
- Wang, H.; Ba, H.; Yang, C.; Zhang, J.; Tai, Y. Inner and Inter Population Structure Construction of Chinese Jiangsu Han Population Based on Y23 STR System. PLoS ONE 2017, 12, e0180921. [Google Scholar] [CrossRef]
- Wang, C.-Z.; Zhang, J.-S.; Li, X.-B.; Bai, R.-F.; Shi, M.-S.; Wang, C.-C. Haplotype Analysis of 36 Y-STR Loci in a Chinese Han Population from Anhui Province, Eastern China. Int. J. Leg. Med. 2020, 134, 2063–2065. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Zhang, C.; Li, R.; Yang, Y.; Ou, X.; Tong, D.; Sun, H. Genetic Polymorphisms and Mutation Rates of 27 Y-Chromosomal STRs in a Han Population from Guangdong Province, Southern China. Forensic Sci. Int. Genet. 2016, 21, 5–9. [Google Scholar] [CrossRef]
- Sun, H.; Su, K.; Fan, C.; Long, F.; Liu, Y.; Sun, J.; Mo, X.; Ge, Y.; Zhang, L.; Zhai, L.; et al. Y-STRs’ Genetic Profiling of 1953 Individuals from Two Chinese Han Populations (Guizhou and Shanxi). Forensic Sci. Int. Genet. 2019, 38, e8–e10. [Google Scholar] [CrossRef] [PubMed]
- Guo, F. Population Genetics for 17 Y-STR Loci in Northern Han Chinese from Liaoning Province, Northeast China. Forensic Sci. Int. Genet. 2017, 29, e35–e37. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.-Y.; Zhang, Z.-Q.; Tang, P.-Z.; Song, D.-L.; Zheng, X.-K.; Zhou, Y.-J.; Liu, M.-N. Forensic and Phylogenetic Analyses of Populations in the Tibetan-Yi Corridor Using 41 Y-STRs. Int. J. Leg. Med. 2021, 135, 783–785. [Google Scholar] [CrossRef]
- Wu, W.; Pan, L.; Hao, H.; Zheng, X.; Lin, J.; Lu, D. Population Genetics of 17 Y-STR Loci in a Large Chinese Han Population from Zhejiang Province, Eastern China. Forensic Sci. Int. Genet. 2011, 5, e11–e13. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Pan, L.; Tang, P.; Zhou, Y.; Liu, M.; Luo, X. Technical Note: Developmental Validation of a Novel 41-Plex Y-STR System for the Direct Amplification of Reference Samples. Int. J. Leg. Med. 2021, 135, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Bian, Y.; Zhang, S.; Zhu, R.; Zhou, W.; Gao, Y.; Li, C. Population Genetics Study Using 26 Y-Chromosomal STR Loci in the Hui Ethnic Group in China. Forensic Sci. Int. Genet. 2017, 28, e26–e27. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Song, F.; Li, J.; Lang, M.; Luo, H.; Wang, Z.; Wu, J.; Li, C.; Tian, C.; Wang, W.; et al. Genetic Substructure and Forensic Characteristics of Chinese Hui Populations Using 157 Y-SNPs and 27 Y-STRs. Forensic Sci. Int. Genet. 2019, 41, 11–18. [Google Scholar] [CrossRef]
- Fan, H.; Wang, X.; Chen, H.; Zhang, X.; Huang, P.; Long, R.; Liang, A.; Song, T.; Deng, J. Population Analysis of 27 Y-Chromosomal STRs in the Li Ethnic Minority from Hainan Province, Southernmost China. Forensic Sci. Int. Genet. 2018, 34, e20–e22. [Google Scholar] [CrossRef]
- Song, M.; Wang, Z.; Zhang, Y.; Zhao, C.; Lang, M.; Xie, M.; Qian, X.; Wang, M.; Hou, Y. Forensic Characteristics and Phylogenetic Analysis of Both Y-STR and Y-SNP in the Li and Han Ethnic Groups from Hainan Island of China. Forensic Sci. Int. Genet. 2019, 39, e14–e20. [Google Scholar] [CrossRef]
- Wang, C.; Su, M.; Li, Y.; Chen, L.; Jin, X.; Wen, S.; Tan, J.; Shi, M.; Li, H. Genetic Polymorphisms of 27 Yfiler® Plus Loci in the Daur and Mongolian Ethnic Minorities from Hulunbuir of Inner Mongolia Autonomous Region, China. Forensic Sci. Int. Genet. 2019, 40, e252–e255. [Google Scholar] [CrossRef]
- Zhu, B.; Wu, Y.; Shen, C.; Yang, T.; Deng, Y.; Xun, X.; Tian, Y.; Yan, J.; Li, T. Genetic Analysis of 17 Y-Chromosomal STRs Haplotypes of Chinese Tibetan Ethnic Group Residing in Qinghai Province of China. Forensic Sci. Int. 2008, 175, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Bai, P.; Zhu, W.; Chen, D.; Wang, H.; Jin, B.; Zhang, L.; Liang, W. Genetic Portrait of 27 Y-STR Loci in the Tibetan Ethnic Population of the Qinghai Province of China. Forensic Sci. Int. Genet. 2018, 34, e18–e19. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Shen, C.; Qian, G.; Shi, R.; Dang, Y.; Zhu, J.; Huang, P.; Xu, Y.; Zhao, Q.; Ma, J.; et al. Genetic Polymorphisms for 11 Y-STRs Haplotypes of Chinese Yi Ethnic Minority Group. Forensic Sci. Int. 2006, 158, 229–233. [Google Scholar] [CrossRef]
- Fan, G.-Y.; An, Y.-R.; Peng, C.-X.; Deng, J.-L.; Pan, L.-P.; Ye, Y. Forensic and Phylogenetic Analyses among Three Yi Populations in Southwest China with 27 Y Chromosomal STR Loci. Int. J. Leg. Med. 2019, 133, 795–797. [Google Scholar] [CrossRef] [PubMed]
- Watahiki, H.; Fujii, K.; Fukagawa, T.; Mita, Y.; Kitayama, T.; Mizuno, N. Polymorphisms and Microvariant Sequences in the Japanese Population for 25 Y-STR Markers and Their Relationships to Y-Chromosome Haplogroups. Forensic Sci. Int. Genet. 2019, 41, e1–e7. [Google Scholar] [CrossRef] [PubMed]
- Uchihi, R.; Yamamoto, T.; Usuda, K.; Yoshimoto, T.; Tanaka, M.; Tokunaga, S.; Kurihara, R.; Tokunaga, K.; Katsumata, Y. Haplotype Analysis with 14 Y-STR Loci Using 2 Multiplex Amplification and Typing Systems in 2 Regional Populations in Japan. Int. J. Leg. Med. 2003, 117, 34–38. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, N.Y.; Hong, S.B.; Cho, N.S.; Kim, J.J.; Han, M.S.; Kim, W. Genetic Polymorphisms of 16 Y Chromosomal STR Loci in Korean Population. Forensic Sci. Int. Genet. 2008, 2, e9–e10. [Google Scholar] [CrossRef]
- Kayser, M.; Brauer, S.; Schädlich, H.; Prinz, M.; Batzer, M.A.; Zimmerman, P.A.; Boatin, B.A.; Stoneking, M. Y Chromosome STR Haplotypes and the Genetic Structure of U.S. Populations of African, European, and Hispanic Ancestry. Genome Res. 2003, 13, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Wang, C.-C.; Shrestha, R.; Wang, L.-X.; Zhang, M.; He, Y.; Kidd, J.R.; Kidd, K.K.; Jin, L.; Li, H. Inferring Population Structure and Demographic History Using Y-STR Data from Worldwide Populations. Mol. Genet. Genom. 2015, 290, 141–150. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Su, K.; He, Z.; Zhai, D.; Guo, K.; Chen, X.; Jin, L.; Li, S. Genetic Reconstruction and Forensic Analysis of Chinese Shandong and Yunnan Han Populations by Co-Analyzing Y Chromosomal STRs and SNPs. Genes 2020, 11, 743. [Google Scholar] [CrossRef]
- Verma, P.J. Data Analysis in Management with SPSS Software; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-81-322-0785-6. [Google Scholar]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Balaresque, P.; Poulet, N.; Cussat-Blanc, S.; Gerard, P.; Quintana-Murci, L.; Heyer, E.; Jobling, M.A. Y-Chromosome Descent Clusters and Male Differential Reproductive Success: Young Lineage Expansions Dominate Asian Pastoral Nomadic Populations. Eur. J. Hum. Genet. 2015, 23, 1413–1422. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.R.; Epplen, J.T.; Pena, S.D. Testing Deficiency Paternity Cases with a Y-Linked Tetranucleotide Repeat Polymorphism. EXS 1993, 67, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Kayser, M. Forensic Use of Y-Chromosome DNA: A General Overview. Hum Genet 2017, 136, 621–635. [Google Scholar] [CrossRef] [PubMed]
- Corach, D.; Filgueira Risso, L.; Marino, M.; Penacino, G.; Sala, A. Routine Y-STR Typing in Forensic Casework. Forensic Sci. Int. 2001, 118, 131–135. [Google Scholar] [CrossRef]
- Guo, F.; Li, J.; Chen, K.; Tang, R.; Zhou, L. Population Genetic Data for 27 Y-STR Loci in the Zhuang Ethnic Minority from Guangxi Zhuang Autonomous Region in the South of China. Forensic Sci. Int. Genet. 2017, 27, 182–183. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, L.; Qian, E.; Long, F.; Cui, W.; Ji, A.; Zhang, F.; Zou, K.; Huang, J.; Li, C. Genetic Polymorphisms and Haplotypic Structure Analysis of the Guizhou Gelao Ethnic Group Based on 35 Y-STR Loci. Leg. Med. 2020, 43, 101666. [Google Scholar] [CrossRef]
- Zhou, H.; Ren, Z.; Zhang, H.; Wang, J.; Huang, J. Genetic Profile of 17 Y Chromosome STRs in the Guizhou Han Population of Southwestern China. Forensic Sci. Int. Genet. 2016, 25, e6–e7. [Google Scholar] [CrossRef]
- Zhong, H.; Shi, H.; Qi, X.B.; Duan, Z.Y.; Tan, P.P.; Jin, L.; Su, B.; Ma, R.Z. Extended Y Chromosome Investigation Suggests Postglacial Migrations of Modern Humans into East Asia via the Northern Route. Mol. Biol. Evol. 2011, 28, 717–727. [Google Scholar] [CrossRef]
- Yan, S.; Wang, C.-C.; Li, H.; Li, S.-L.; Jin, L.; Schurr, T.; Santos, F.; Quintana-Murci, L.; Bertranpetit, J.; Comas, D.; et al. An Updated Tree of Y-Chromosome Haplogroup O and Revised Phylogenetic Positions of Mutations P164 and PK4. Eur. J. Hum. Genet. 2011, 19, 1013–1015. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; He, Z.; Wang, Y.; He, X.; Zhang, X.; Xia, M.; Zhai, D.; Chang, K.; Chen, X.; Chen, X.; et al. Improving the Regional Y-STR Haplotype Resolution Utilizing Haplogroup-Determining Y-SNPs and the Application of Machine Learning in Y-SNP Haplogroup Prediction in a Forensic Y-STR Database: A Pilot Study on Male Chinese Yunnan Zhaoyang Han Population. Forensic Sci. Int. Genet. 2022, 57, 102659. [Google Scholar] [CrossRef] [PubMed]
- Rootsi, S.; Zhivotovsky, L.A.; Baldovič, M.; Kayser, M.; Kutuev, I.A.; Khusainova, R.; Bermisheva, M.A.; Gubina, M.; Fedorova, S.A.; Ilumäe, A.-M.; et al. A Counter-Clockwise Northern Route of the Y-Chromosome Haplogroup N from Southeast Asia towards Europe. Eur. J. Hum. Genet. 2007, 15, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Ke, Y.; Su, B.; Xiao, J.; Chen, H.; Huang, W.; Chen, Z.; Chu, J.; Tan, J.; Jin, L.; Lu, D. Y-Chromosome Haplotype Distribution in Han Chinese Populations and Modern Human Origin in East Asians. Sci. China Ser. C Life Sci. 2001, 44, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zheng, H.; Bei, J.-X.; Sun, L.; Jia, W.; Li, T.; Zhang, F.; Seielstad, M.; Zeng, Y.-X.; Zhang, X.; et al. Genetic Structure of the Han Chinese Population Revealed by Genome-Wide SNP Variation. Am. J. Hum. Genet. 2009, 85, 775–785. [Google Scholar] [CrossRef] [PubMed]
- Gan, R.-J.; Pan, S.-L.; Mustavich, L.F.; Qin, Z.-D.; Cai, X.-Y.; Qian, J.; Liu, C.-W.; Peng, J.-H.; Li, S.-L.; Xu, J.-S.; et al. Pinghua Population as an Exception of Han Chinese’s Coherent Genetic Structure. J. Hum. Genet. 2008, 53, 303–313. [Google Scholar] [CrossRef]
- Auton, A.; Abecasis, G.R.; Altshuler, D.M.; Durbin, R.M.; Abecasis, G.R.; Bentley, D.R.; Chakravarti, A.; Clark, A.G.; Donnelly, P.; Eichler, E.E.; et al. A Global Reference for Human Genetic Variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef]
- Sudmant, P.H.; Rausch, T.; Gardner, E.J.; Handsaker, R.E.; Abyzov, A.; Huddleston, J.; Zhang, Y.; Ye, K.; Jun, G.; Hsi-Yang Fritz, M.; et al. An Integrated Map of Structural Variation in 2,504 Human Genomes. Nature 2015, 526, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Sun, H.; Zhou, H.; Jin, L.; Li, S. EA-YPredictor: One New Software Developed to Predict Pedigree Haplogroup Based on Y-STR Haplotypes. Forensic Sci. Technol. 2020, 45, 117–124. [Google Scholar] [CrossRef]
- Ke, Y.; Su, B.; Song, X.; Lu, D.; Chen, L.; Li, H.; Qi, C.; Marzuki, S.; Deka, R.; Underhill, P.; et al. African Origin of Modern Humans in East Asia: A Tale of 12,000 Y Chromosomes. Science 2001, 292, 1151–1153. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time(s) a Haplotype Was Observed | Y Filer | PPY23 | Y Filer Plus | Y Filer Platinum | ||||
|---|---|---|---|---|---|---|---|---|
| PD | CM | PD | CM | PD | CM | PD | CM | |
| 1 | 573 | 388 | 630 | 462 | 657 | 508 | 669 | 522 |
| 2 | 30 | 27 | 26 | 15 | 16 | 8 | 10 | 4 |
| 3 | 9 | 9 | 1 | 6 | 2 | |||
| 4 | 6 | 3 | 1 | 2 | ||||
| 5 | 1 | 3 | 1 | |||||
| 6 | 3 | |||||||
| 7 | 1 | 1 | ||||||
| 9 | 1 | |||||||
| No. of haplotypes | 619 | 435 | 658 | 487 | 673 | 518 | 679 | 526 |
| FUH | 0.83164 | 0.73208 | 0.91437 | 0.87170 | 0.95356 | 0.95849 | 0.97097 | 0.98491 |
| HD | 0.99957 | 0.99854 | 0.99985 | 0.99946 | 0.99993 | 0.99990 | 0.99996 | 0.99997 |
| MP | 0.00189 | 0.00334 | 0.00160 | 0.00243 | 0.00152 | 0.00199 | 0.00149 | 0.00192 |
| DC | 0.89840 | 0.82075 | 0.95501 | 0.91887 | 0.97678 | 0.97736 | 0.98549 | 0.99245 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Tang, Z.; Wang, B.; Zhou, X.; Zhou, L.; Zhang, G.; Tian, J.; Zhao, Y.; Yao, Z.; Tian, L.; et al. Forensic Analysis and Genetic Structure Construction of Chinese Chongming Island Han Based on Y Chromosome STRs and SNPs. Genes 2022, 13, 1363. https://doi.org/10.3390/genes13081363
Zhang X, Tang Z, Wang B, Zhou X, Zhou L, Zhang G, Tian J, Zhao Y, Yao Z, Tian L, et al. Forensic Analysis and Genetic Structure Construction of Chinese Chongming Island Han Based on Y Chromosome STRs and SNPs. Genes. 2022; 13(8):1363. https://doi.org/10.3390/genes13081363
Chicago/Turabian StyleZhang, Xiao, Zhen Tang, Bin Wang, Xindao Zhou, Limin Zhou, Gongying Zhang, Junzhe Tian, Yiqi Zhao, Zhiqing Yao, Lu Tian, and et al. 2022. "Forensic Analysis and Genetic Structure Construction of Chinese Chongming Island Han Based on Y Chromosome STRs and SNPs" Genes 13, no. 8: 1363. https://doi.org/10.3390/genes13081363